Introduction
Esophageal cancer (EC), which includes esophageal
squamous cell carcinoma (ESCC) and esophageal adenocarcinoma (EAC),
is the sixth most leading cause of cancer-related death worldwide
(1,2). The five-year survival rate of
patients with ESCC is less than 20%. In China, over 90% of EC cases
are ESCC, and EC is ranked as the sixth most frequent cancer and
the fourth leading cause of cancer-related death (3–5).
The low survival rate is likely due to increased resistance
(acquired or intrinsic) of tumor cells to chemo/radiotherapies
(6,7).
Recent study suggested that microRNAs (miRNAs) can
serve as biomarkers in predicting therapeutic outcomes in a variety
of solid tumors, including EC (6). In the context of ESCC, studies have
demonstrated that one set of miRNAs promotes the development of
radio resistance, while another set of miRNAs sensitizes EC cells
to radiation therapy treatments (6,8–10).
Therefore, an improved understanding of these miRNAs in responding
to ESCC radiotherapy remains demanding.
Exosomes are nanosized vesicles (30–150 nm diameter)
that are secreted by most cells. They are enclosed by a lipid
bilayer and carry various biomolecules, including proteins,
glycans, lipids, metabolites, RNAs and DNAs (11). Studies have shown that miRNAs are
the predominant RNA species transported by exosomes (12–15). Since exosomes can transport and
transfer bioactive molecules, extensive studies have focused on
their ability to regulate tumor growth and metastasis (16–19). Exosomal RNA has been demonstrated
as the potential cancer biomarker by regulating the communications
among different cells (12,20–22). For example, circUBE2Q2 regulates
gastric cancer progression via the circUBE2Q2-miR-370-3p-STAT3 axis
and promotes tumor metastasis through exosomal communications
(22). Exosomal miR-1245 from
colorectal cancer cells reprograms macrophages to tumor-associated
macrophages (TAMs) with high transforming growth factor β (TGF-β)
expression that enables tumor growth and metastasis (23). However, the roles of exosomal
miRNAs in ESCC remain rudimentary (24–26). Understanding dynamic changes of
exosomal miRNAs and their target genes will help improve the
therapeutic outcome in patients with EC.
In the present study, the expression signature of
plasma exosomal miRNAs in patients with ESCC was investigated
before and after radiotherapy and differentially expressed miRNAs
were identified. Target genes for up- and downregulated miRNAs were
then predicted and relevant biological pathways and potential
biomarkers for ESCC responding to radiotherapy were identified.
Finally, it was found that the changes of radiation-sensitized
miR-652 and miR-30a can regulate cancer cell migration rates
without changing proliferation in vitro. The present study
provided a reference for future identification of potential
biomarkers for cancer treatment through radiotherapy.
Materials and methods
Patients and sample selection
Plasma samples were collected from ESCC female
patients with pre- and post-radiotherapy (mean age, 42; stage II;
September 2020-December 2020), and were provided by the First
Hospital of Quanzhou. Briefly, female patients with ESCC at stage
II with the following characteristics were excluded from the
present study: i) metastasis, ii) poorly controlled blood pressure,
hyperlipidemia and diabetes, iii) alcohol consumption and cigarette
smoking and iv) infectious diseases. One week before receiving
radiotherapy, pre-radiotherapy plasma samples were collected. One
week after having received 50 Gy radiation (2.0 Gy/fraction, 5
fractions/week, for 5 weeks), patient plasma samples were
collected, termed as post-radiotherapy.
In the present study, 7 sets of pre- and post-
radiotherapy plasma samples were obtained from 7 female ESCC
patients at stage II (Table SI).
For each patient, plasma samples were analyzed as a pair for pre-
and post-radiotherapy. Plasma samples were used for the following
experiments: i) exosome characterizations (n=1, sample no. 37892),
ii) exosome miRNA sequencing (n=3, sample nos. 38380, 38632 and
38930, retained corresponding tumor tissues for H&E staining)
and iii) reverse transcription-quantitative (RT-q) PCR validation
for exosome miRNAs (n=3, sample nos. 38912, 38783 and 39221).
The present study was approved [approval no.
(2018)101] by the Institution Ethic Issue Committee of the First
Hospital of Quanzhou (Quanzhou, China). Signed informed consents
were obtained from female patients with ESCC who received
radiotherapy diagnosis.
Exosome isolation and RNA
purification
Cleared plasma were isolated from 5 ml blood by
ultracentrifugation-based assay as previously described (27). Next, exosomes were isolated from
cleared plasma by SmartSEC™ HT EV Isolation System for Serum &
Plasma (cat. no. SSEC096A-1; SBI, systembio.com). A total of
250–500 µl of cleared plasma were applied to each well of a filter
plate, incubated and centrifuged (3,000 g for 10 min at 4°C) to
elute the first fraction. Equal volumes of SmartSEC Isolation
Buffer were added and plasma samples were centrifuged (1,000 × g
for 30 sec at 4°C) again into a clean plate to elute the second
fraction. Depending on the volume of sample loaded, the majority of
exosomes was collected in either the first or second elution.
Isolated exosomes were immediately used for experiments or stored
at 4°C. For a long-period storage, exosomes were frozen at
−80°C.
Exosomal RNAs were purified using SeraMir™ Exosome
RNA Amplification kit (cat. no. RA808A-1; SBI). A total of 500 µl
serum and 120 µl ExoQuick Media were combined and placed at 4°C for
30 min. After centrifugation at 4,000 g for 2 min at 4°C, 350 µl
LYSIS Buffer was added to the exosome pellet and vortexed for 15
sec. A total of 200 µl 100% ethanol was added, samples were
centrifuged at 4,000 g for 1 min at 4°C, washed twice and were let
to dry. A total of 30 µl elution buffer were added directly to the
membrane in a spin column. Samples were centrifuged at 600 g for 2
min at 4°C, and increased to 4,000 g for 1 min at 4°C. Exosome RNAs
(30–40 µl) were obtained. The quality and quantity of these RNA
samples were determined by a NanoDrop spectrophotometer (ND-2000C;
Thermo Fisher Scientific Inc.). Agilent RNA 6000 Nano assay
(Agilent Technologies, Inc.) and agarose gel electrophoresis were
used to check purity of the RNA samples.
Transmission electron microscopy
(TEM), Nanoparticle tracking analysis (NTA) and western blot
assay
The isolated exosomes were centrifuged using an
airfuge at 30,000 g with 25 pounds per square inch for 45 min at
4°C. Exosomes were fixed with 2.5% glutaraldehyde at 4°C overnight.
After washing, vesicles were loaded onto formvar/carbon-coated
grids, negatively stained with aqueous phosphotungstic acid for 60
sec and images were captured with a transmission electron
microscope (H7500, 100XCII; JEOL, Ltd.) at 80 kV.
Purified exosomes were diluted to 106–107
particles/ml in PBS for NTA using a Nanosight 300 equipped with
v3.2.16 analytical software (Malvern Instruments, Inc.) (28,29). Two videos (25 sec each) were
recorded for each sample and the software was used to estimate
concentration and size of the particles. The recordings were
performed at room temperature and were monitored manually. For
analysis, the detection threshold was set to 6. Calibration was
carried out using 100 nm polystyrene latex microspheres (Magsphere
Inc. magsphere.com) diluted to PBS and then two videos were
recorded.
Aliquots (5–10 µl) of isolated exosomes were
dispensed into wells of a 96-well plate, and the assay was
performed as recommended by the manufacturer instructions (Pierce
BCA Protein Assay kit; cat. no. lL-61105; Thermo Fisher Scientific,
Inc.). Total protein concentrations were determined using a linear
standard curve established with bovine serum albumin. Because
exosomes normally contain a common set of proteins, named
tetraspanins, including CD81, CD63 and CD9 (29), expression of CD81, CD63 and CD9
was validated by western blot analysis. Total proteins were then
separated by 12%-polyacrylamide gel electrophoresis. A total of 7.5
µl protein sample (5–25 µg proteins) was mixed with 2.5 µl 4X
lithium dodecyl sulfate sample loading buffer (Invitrogen; Thermo
Fisher Scientific, Inc.) and heated at 70°C for 10 min. Samples
were then loaded into pre-casted NuPAGE Novex 12% Bis-Tris 1.0 mm
mini-gels (Invitrogen; Thermo Fisher Scientific, Inc.). Then, 5 µl
pre-stained SDS-PAGE Standards (Bio-Rad Laboratories, Inc.) were
loaded in each gel run.
Electrophoresis was performed at room temperature
for ~45 min using a constant voltage (200 V) in 1X solution of
NuPAGE MOPS SDS running buffer (Invitrogen; Thermo Fisher
Scientific, Inc.) until the dye front reached the bottom of the
60-mm gel. After proteins were transferred to PVDF membrane, 5%
skimmed milk powder was sealed at room temperature for 2 h. Then,
blots were incubated overnight at 4°C with appropriate antibodies.
The following primary antibodies were used: CD9 (1:1,000; ~24 kDa;
cat. no. MA5-31980), CD81 (1:1,000; ~24 kDa; cat. no. MA5-13548);
CD63 (1:1,000; 40~60 kDa; cat. no. MA1-19281) and Ab specific for
GAPDH (1:1,000; ~37 kDa; AB_10977387; all from Thermo Fisher
Scientific, Inc.). The next day, blots were incubated with goat
anti-rabbit antibody (1:5,000; cat. no. ab205718) and goat
anti-mouse (1:5,000; cat. no. ab205719; both from Abcam)
horseradish peroxidase-conjugated IgG for 2 h at room temperature.
After washed with Tris-Tween (0.05%) buffer saline, blots were
placed in a gel imager and treated with enhanced chemiluminescence
(ECL; Bio-Rad Laboratories, Inc.) super-sensitive solution for 1–3
min. The optical densities of bands were analyzed using ImageJ
software 1.53 (National Institutes of Health) (30).
Hematoxylin-eosin (H&E)
staining
Paraffin blocks of esophageal tumor tissues
(Table SI) were cut into 4-µm
thickness. Sections were deparaffinized in xylene and dehydrated in
alcohol. Tissues were stained with Harris' hematoxylin solution for
6 h at 60–70°C, and were then rinsed in tap water until the water
was colorless. Next, 10% acetic acid and 85% ethanol in water were
used to differentiate the tissue twice for 2 and 10 h each. After
rinsing with tap water, tissues were soaked in saturated lithium
carbonate solution for 12 h and then rinsed again with tap water.
Finally, staining was performed with eosin Y ethanol solution for
48 h. Images were captured by a light microscope (Nikon
Corporation).
MiRNA isolation, sequencing and
analysis
RNAs in exosomes were isolated from three patients
with pre- and post-radiotherapy (Table SI). Total RNA of each sample was
used to prepare the miRNA sequencing library, which included the
following steps: i) 3′-adaptor ligation, ii) 5′-adaptor ligation,
iii) cDNA synthesis, iv) PCR amplification and v) size selection of
~150 bp PCR amplicons (corresponding to ~22 nt miRNAs) (Cloud-Seq
Biotech, cloud-seq.com.cn). The libraries were denatured as
single-stranded DNA molecules, captured on Illumina flow cells,
amplified in situ as clusters and finally sequenced for 50
cycles on an Illumina HiSeq sequencer following the manufacturer's
instructions.
Raw data was generated after sequencing, image
analysis, base calling and quality filtering on Illumina sequencer.
Q30 was used to perform the quality control. The adaptor sequences
were trimmed and the adaptor-trimmed-reads (≥15 nt) were left by
cutadapt software (v1.9.2,MIT). Trimmed reads from all samples were
pooled, and miRDeep2 software (v2.0.0.5, MDC Berlin-Mitte) was used
to predict novel miRNAs. The trimmed reads were aligned to the
merged human pre-miRNA databases (known pre-miRNA from miRBase plus
the newly predicted pre-miRNAs) using Novoalign software (v3.02.12,
Novocraft Technologies Sdn Bhd) with at most one mismatch. The
numbers of mature miRNA mapped tags were defined as the raw
expression levels of that miRNA. The read counts were normalized by
TPM (tag counts per million aligned miRNAs) approach.
Differentially expressed miRNAs between two samples were filtered
through Fold change and P-value.
TE-1 cell culture and miRNA
mimics/inhibitor transfection
Human EC cell line TE-1, developed by the cell bank
at the Chinese Academy of Science, was reported to maintain
biological characteristics of ESCC (31). TE-1 cells were cultured in
RPMI-1640 with 10% exosome-free fetal bovine serum (FBS, both
Thermo Fisher Scientific, Inc.) and maintained in an incubator
containing 5% CO2 at 37°C. The exosome-free FBS was
produced by centrifugation (100,000 g) at 4°C overnight in order to
ensure the removal of any bovine-derived exosomes. TE-1 cells
(3×105) were seeded in a six-well plate and incubated 12
h at 37°C for attachment. Cells were then transfected at 37°C using
Lipofectamine™ 3000 (Thermo Fisher Scientific, Inc.) with mimics or
inhibitors (10 µl; 20 µM) according to the manufacturer's protocol
for 12, 24 and 72 h. 12, 24 and 72 h subsequent experiments were
performed. Transient overexpression and inhibition of
miR-30a-3p/miR-652-3p were performed by transfection with miRNA
oligonucleotides (miR-30a-3p mimics, CUUUCAGUCGGAUGUUUGCAGC,
miR-652-3p mimics: AAUGGCGCCACUAGGGUUGUG, miR-30a-3p inhibitors:
GCUGCAAACAUCCGACUGAAAG, miR-652-3p inhibitors:
CACAACCCUAGUGGCGCCAUU, mimics negative controls:
AGGCAAGCUGACCCUGAAGU, inhibitor negative control:
CAGUACUUUUGUGUAGUACAA, 2 OD miRNA oligonucleotides were dissolved
in ddH2O as the 20 µM solution, which were purchased
from the Shanghai GenePharma Co., Ltd.).
RT-qPCR
RNAs in exosomes were isolated from three patients
with pre- and post-radiotherapy (Table SI). After exosome RNA extraction
(SeraMir™ Exosome RNA Amplification kit, cat. no. RA808A-1,SBI),
cDNA for polyA miRNA RT-qPCR was synthesized using All-in-One miRNA
First-Strand cDNA Synthesis kit (GeneCopoeia, Inc.) according to
the manufacturer's protocols. Expression levels of mRNA from cells
were tested using RT-qPCR with specific primers in triplicates. U6
(Rnu6-1) was used as an endogenous control for miRNA. Quantitative
expression was conducted on the ABI 7500 real-time PCR system
(Applied Biosystems; Thermo Fisher Scientific, Inc.). qPCR was
performed using the qPCR SYBRGreen Mix (Bio-Rad Laboratories,
Inc.). The relative expression levels of miRNAs were calculated
using the 2−∆∆Cq method (32). The primers for RT-qPCR are shown
in Table SII.
Wound healing and Cell Counting Kit-8
(CCK-8) assay
TE-1 cells were treated with the miRNA mimics or
inhibitor for 72 h. Cells were harvested and seeded in a six-well
plate at a density of 2×105 cells/well. When cells
reached 70–80% confluence, medium was replaced with serum-free DMEM
(Thermo Fisher Scientific, Inc.). A scratch wound was generated
using a sterile 200-µl pipette tip, and floating cells were removed
by washing with 1X PBS. Images of the scratches were captured using
an inverted light microscope at ×100 magnification at 0, 24 and 72
h after scratching. The wound area was normalized as follows: wound
area (%)=An/A0 ×100, where A0
represents the area of initial wound area in control group,
An represents the remaining area of wound at 24 or 72 h.
The data were representative of three independent experiments in
triplicates. 96-well plates (2×104 TE-1 cells/well) were
used here to inoculate and culture TE-1 cells at 37°C with 5%
CO2. After being incubated for 0, 24, 48 and 72 h, 10 µl
CCK-8 solution (cat. no. CK04; Dojindo Laboratories, Inc.) was
added. After 2 h, the absorbance was assessed at 450 nm. The data
were representative of three independent experiments in
triplicate.
MiRNA target prediction and function
analysis
Target prediction of miRNAs was performed using
TargetScan Human7.2 (http://www.targetscan.org/vert_72/) and Funrich
software 3.1.3 (http://funrich.org/) (33).
Predicted target genes were imported into the
OmicShare tools, a free online platform for data analysis
(https://www.omicshare.com/tools). Gene
Ontology (GO; http://geneontology.org/), Kyoto Encyclopedia of Genes
and Genomes (KEGG; http://www.kegg.jp) and Disease Ontology (DO;
http://disease-ontology.org/) analysis
of the miRNAs-targets with differential expression levels was
performed. FunRich 3.1.3 (funrich.org/) was used to compare
functional analysis, clinical phenotype analysis and site of
expression (33). In the present
study, modules in the gene-miRNA-KEGG were mined with Cytoscape
3.9.1, and then functional enrichment analysis was applied to the
miRNAs in the modules (34).
Volcano map, radar map, Circular map, heatmap, Lollipop chart and
Sankey diagram construction were performed using the OmicShare
tools, a free online platform for data analysis (https://www.omicshare.com/tools).
Statistical analysis
The P-values were used to verify statistically
significant difference and were determined by unpaired Student's
t-test for assessing the significance of differences by SPSS
version 25 (IBM Corp.). P<0.05 was considered to indicate a
statistically significant difference.
Results
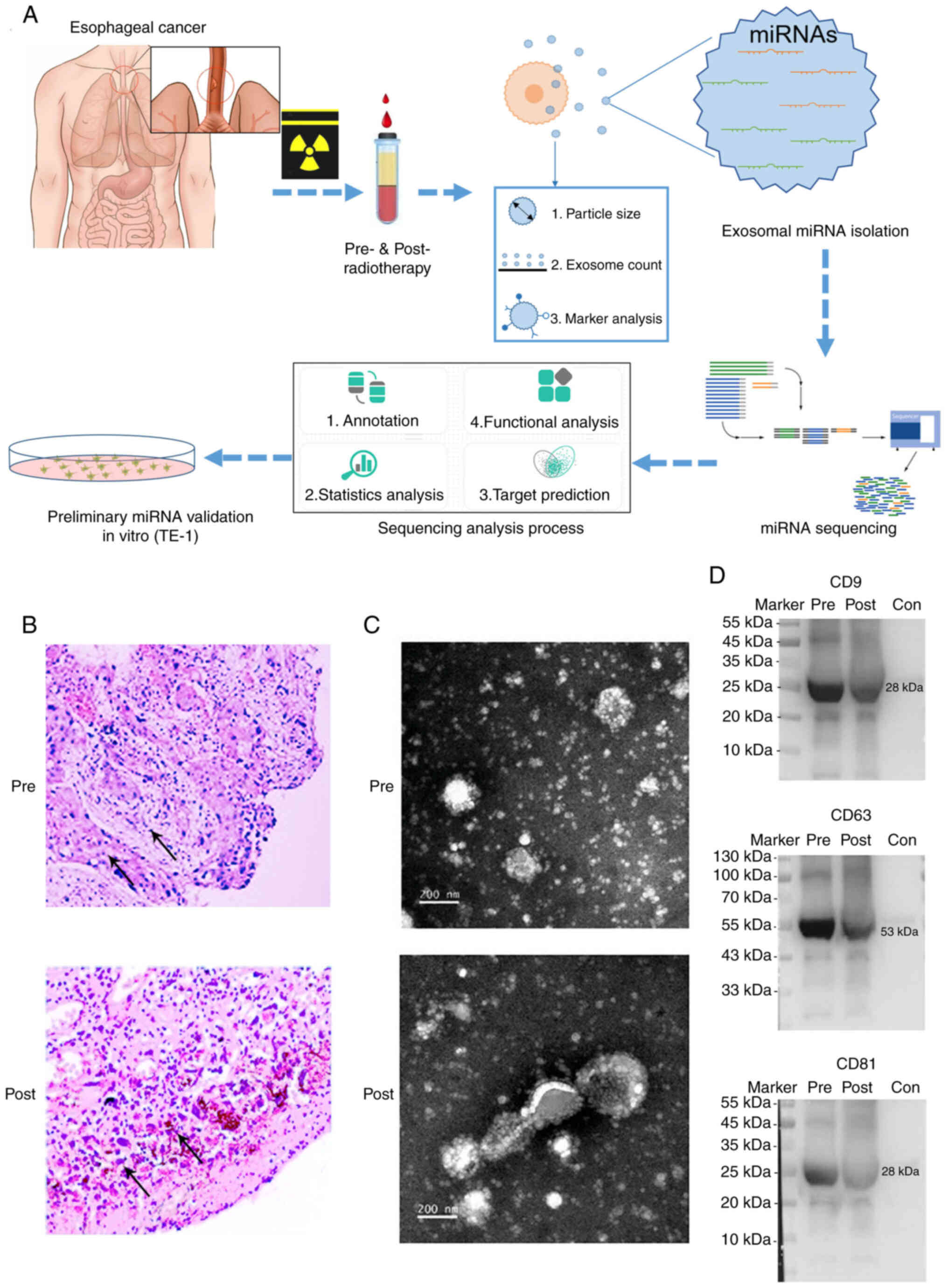
Characterizations of ESCC patient
tumor samples and plasma exosomes
To explore whether exosomal miRNAs respond to
radiation treatment of ESCC, plasma samples were obtained from
three female ESCC patients (mean age=42; stage II, without
metastasis), and exosomal miRNAs were extracted and sequenced
(Fig. 1A). Patients were treated
with radiotherapy (pre- and post-radiotherapy). Tumor samples from
patients for ESCC plasma exosomal miRNA sequencing were stained
with H&E (Table SI).
Microscopy revealed an irregular cell shape, large, irregular and
deeply stained nuclei, and single keratinocytes, confirming the
diagnosis of squamous cell carcinoma (Fig. 1B) (35). Following radiotherapy, tumor cells
with calcifications, larger and blurred nuclei, apoptosis and
necrosis were detected (Fig. 1B).
These changes revealed the typical radiological indicator of early
cancers (stage I–II; Fig. 1B)
(35).
Exosomes were isolated from pre- and
post-radiotherapy plasma samples of 7 patients with ESCC (Fig. 1C). Isolated exosomes were
characterized by TEM. Sizes of exosomes in pre- and post- groups
were 68.28±19.07 and 77.47±26.87 nm, and exosomal concentrations of
two groups were 2.12×1010 and 2.22×1010
particles/ml, respectively (Table
SIII and Fig. S1). High
resolution TEM examinations further showed cup-shaped vesicles of
extracted exosomes (Fig. 1C).
Moreover, expression of three established exosome markers, such as
CD63, CD9 and CD81 were readily identified using western blot
analysis (Fig. 1D) (29,36,37). These results indicated that
exosomes with a high quality are extracted from the plasma of ESCC
patients with pre- and post-radiotherapy.
Differential expression of miRNAs in
exosomes of plasma samples of pre- and post-radiotherapy
To determine whether profiles of exosomal miRNAs
from pre- and post-radiotherapy plasma samples may differ, total
RNAs were isolated from exosomes, and small RNA libraries were
prepared for miRNA sequencing and annotation (Fig. 1A).
Combining 3 samples from pre- and post-radiotherapy
groups and removing low signals, 726 miRNAs were obtained (Table SIV). A total of 8 miRNAs highly
expressed in the pre-radiotherapy group (fold-change
(pre-/post-)>1, log2fold-change>0 and
P<0.05) and 9 miRNAs highly expressed in the post-radiotherapy
group were detected (Fig. 2A and
B and Table SV). The
upregulated (fold-change (pre-/post-)>1,
log2fold-change>0, P<0.05) and
downregulated (fold-change (Pre-/Post-)<1,
log2fold-change<0, P<0.05) miRNAs with
the mean expression and fold-changes were shown in a Radar graph
(Fig. 2C). Certain well-studied
miRNAs such as miR-106b, miR-18a, miR-200a, miR-30a/b, let-7b and
miR-370 exhibited differential expression, and certain new miRNAs
(miR-novel-chr5_22142, miR-novel-chr15_6643 and
miR-novel-chr7_25437) were detected as well. The sequencing results
were further verified using RT-qPCR. As examples, among tested
miRNAs, transcriptional levels of the upregulated (miR-652 and
miR-370) and downregulated (miR-30a and miR-484) miRNAs were
consistent with the exosomal sequencing results (Fig. 2D). These results suggested that
plasma exosomal miRNAs are sensitive and responding to the
radiotherapy of ESCC.
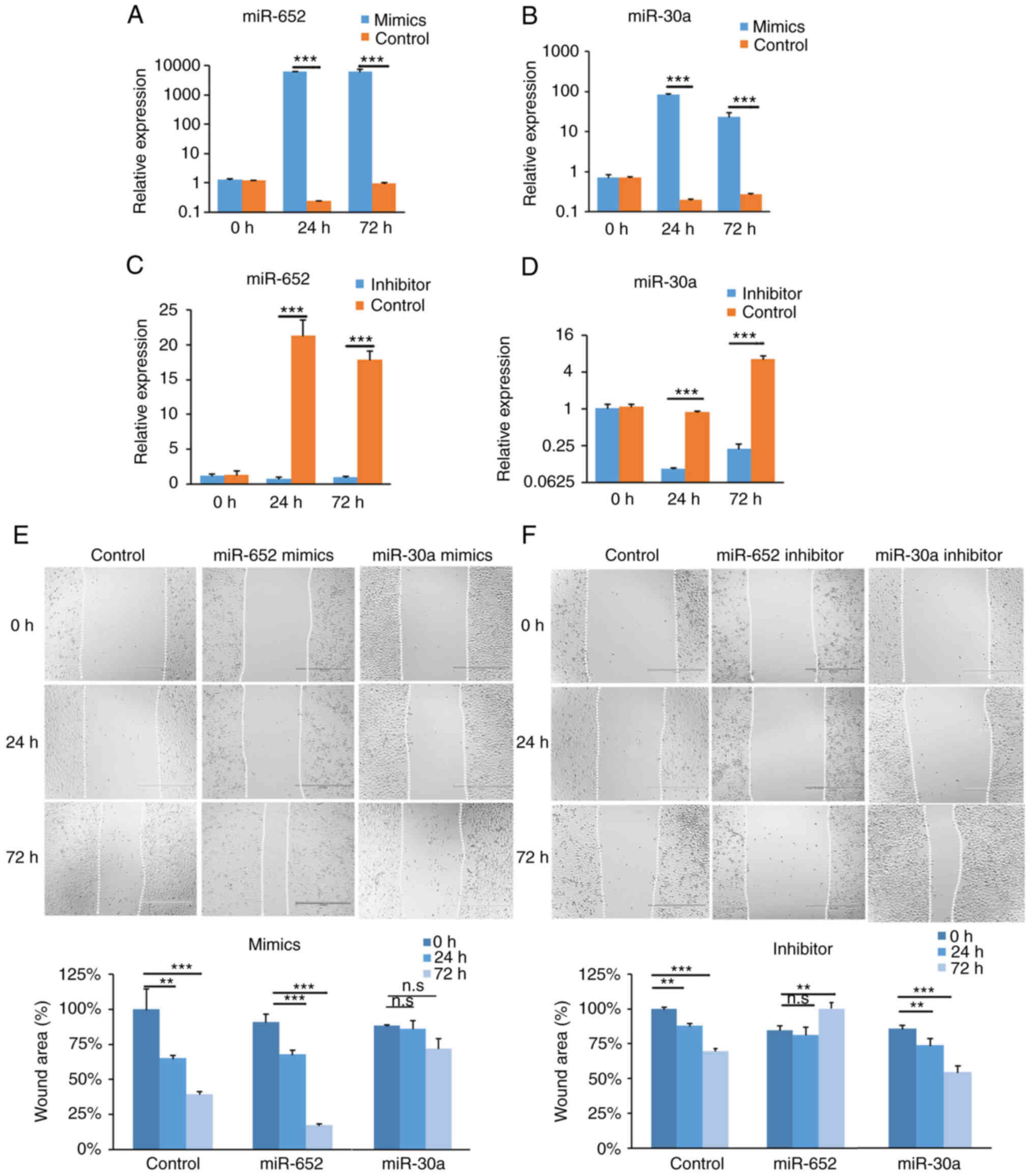
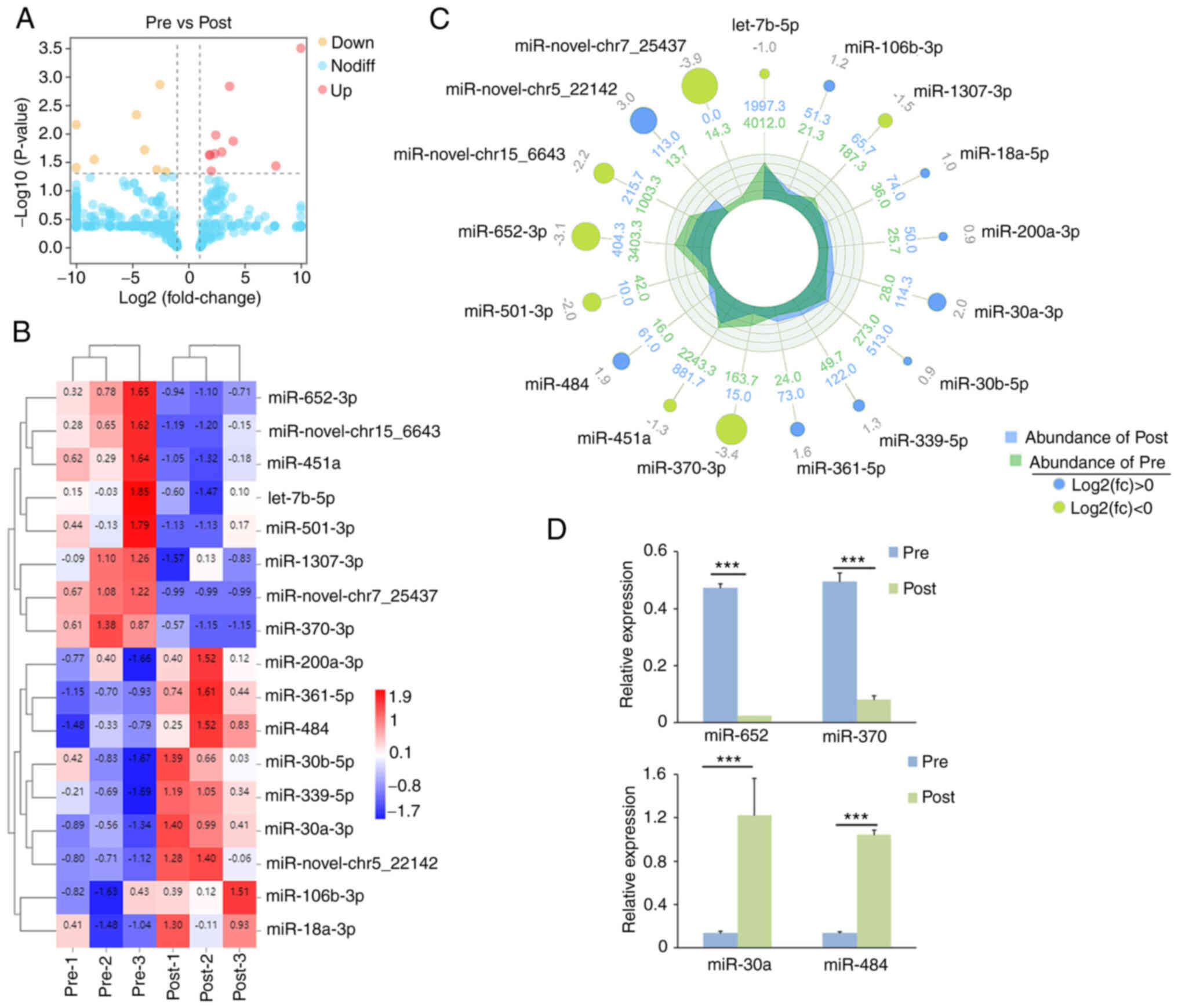 | Figure 2.Differential expression of miRNAs in
plasma exosomes of ESCC patients of pre- and post-radiotherapy. (A)
Volcano plot of whole miRNA expression levels in Pre- and
Post-groups. Each point represents a miRNA, ‘yellow’ dot means
downregulated and ‘red’ dot means upregulated (pre- vs.
post-miRNAs). (B) The heat map of normalized expression of miRNAs
in pre- and post-radiotherapy groups. The heat map was drawn with
relative expression levels of each miRNAs. Blue, white and red
indicate low, middle and high expression levels of miRNAs,
respectively. Color map was used to distinguish the difference of
expression. Each column represents an experimental condition (such
as different comparison groups or samples), and each row represents
the log2Ratio-value of a gene. Different
expression changes or expression amounts are illustrated in
different colors. Blue, white and red indicate low, middle and high
expression levels of miRNAs, respectively. (C) Radar circus of the
most significantly up- and downregulated genes of pre- and
post-radiotherapy groups. From outside to inside: miRNAs with
log2(fold-change), miRNA expression in pre-
and post-radiotherapy groups. (D) Validation of expression levels
of 4 up- and downregulated miRNAs using reverse
transcription-quantitative PCR in ESCC plasma exosomes. Values of
histogram represent the mean ± SEM (n=3), ***P<0.001; unpaired
Student's t-test). miRNA, microRNA; ESCC, esophageal squamous cell
carcinoma. |
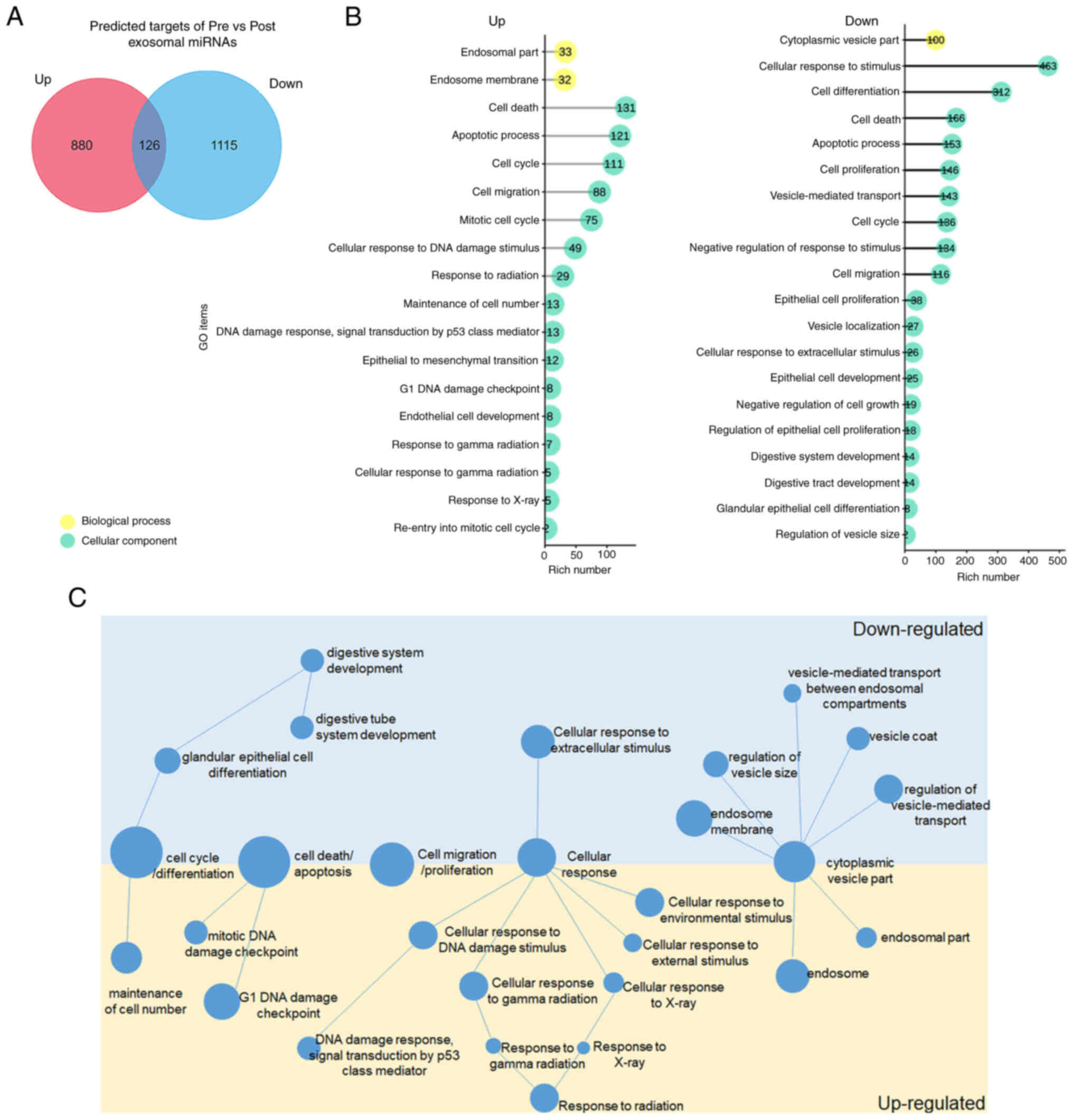
Target prediction and GO analysis of
up- and downregulated miRNAs
Since miRNAs normally function through silencing
their target genes, predicted targets of differentially expressed
exosomal miRNAs were next searched. A total of 1,106 genes were
predicted as targets of upregulated miRNAs, 1,241 genes as targets
of downregulated miRNAs and 126 genes were overlapped in two groups
using TargetScan Human7.2 and Funrich software 3.1.3 (Fig. 3A; Tables SVI and SVII).
Functions of predicted targets of up- and
downregulated miRNAs were next analyzed by GO analysis, and
abundant items in Biological Process, Cellular Component and
Molecular Function were identified (P<0.05; Figs. S2 and S3; Tables SVIII and SIX). It was found that targets of both
up- and downregulated miRNAs play a role in cell
cycle/differentiation, cell death/apoptosis, cell
migration/proliferation, cellular response and cytoplasmic vesicle
part (Fig. 3B and C). Moreover,
targets of upregulated miRNAs were abundant in maintenance of cell
number, DNA damage process, multiple cellular response, endosome
and endosome part, while targets of downregulated miRNAs were
involved in vesicle part, glandular epithelial cell differentiation
and digestive system development (Fig. 3B and C). These results suggested
the distinct functions of targets of up- and downregulated plasma
exosomal miRNAs.
KEGG and DO analysis
Interactions between up- and downregulated miRNAs
and their predicted targets were next studied using KEGG analyses
(Figs. 4 and S4; Tables SX and SXI). Based on the Venn graph, 65 of
309 pathways were significantly abundant (P<0.05) in upregulated
miRNA targets, 42 of 300 pathways were significantly abundant
(P<0.05) in downregulated miRNA targets, and 12 pathways were
overlapped in two groups (Fig.
4A).
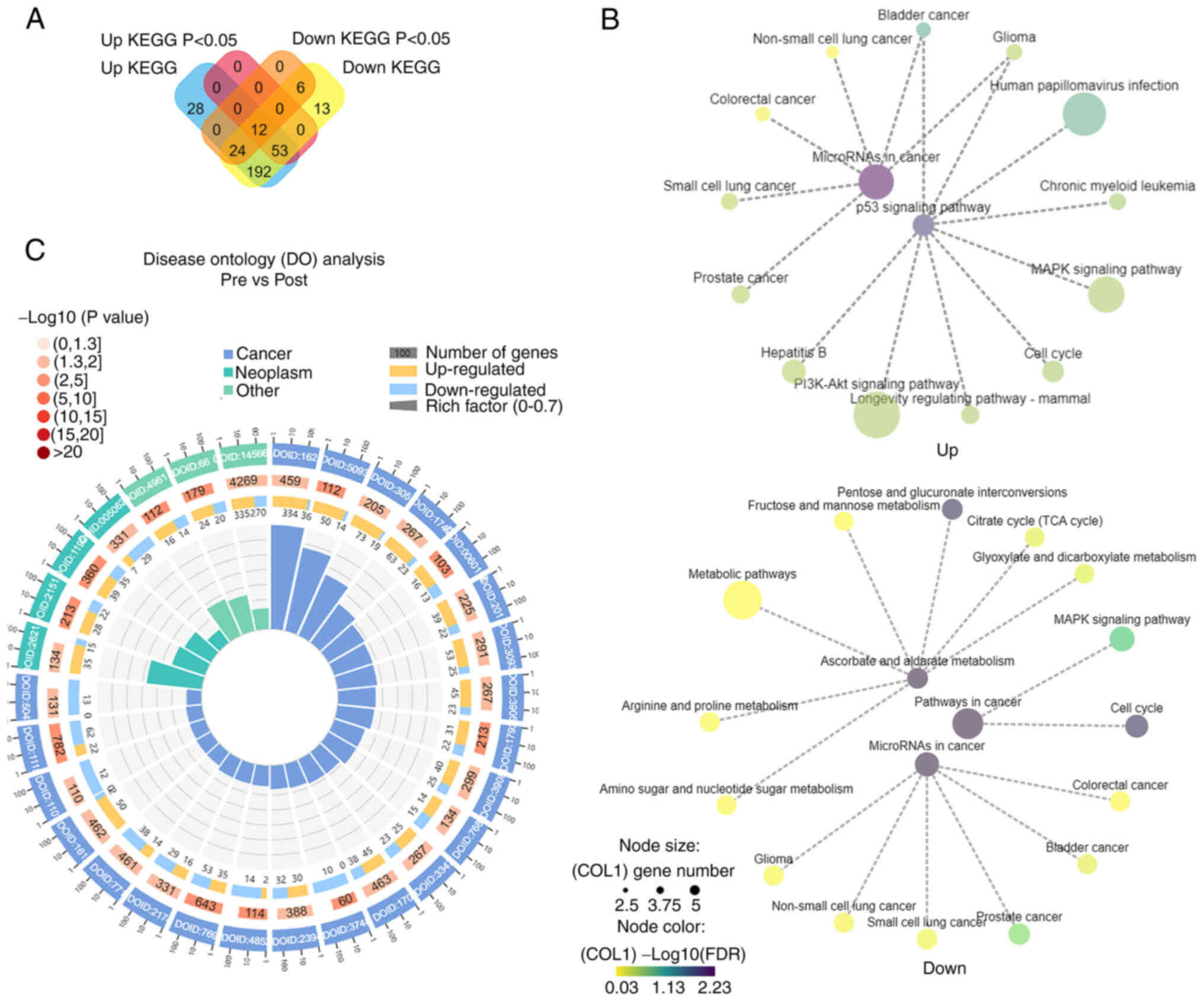 | Figure 4.KEGG and DO analysis in up- and
downregulated miRNAs. (A) Venn diagrams of the enriched KEGG
pathways for the targets of up- and downregulated miRNAs. (B) The
networks of target genes for downregulated vs. upregulated miRNAs
in signal pathways. The colors from yellow, green to purple
represent the values of -log10(FDR), and the
node sizes represent the gene numbers. (C) Comparison analysis of
DO analysis in miRNAs of pre-and post-radiotherapy. Classified as 3
functions: Cancer, neoplasm and other diseases. From outside to
inside: i) enrichment classification, outside the circle is the
coordinate ruler of gene numbers. Different colors represent
different categories; ii) P-values of this classification in
background genes. The more genes, the longer the bars, the smaller
the values, the redder the color is; iii) bar chart of upregulated
gene proportion, dark purple represents upregulated gene
proportion, light purple represents downregulated gene proportion;
iv) Rich Factor value of each classification (the number of
foreground genes in this classification divided by the number of
background genes), and each small bar of background auxiliary line
represents 0.1. KEGG, Kyoto Encyclopedia of Genes and Genomes; DO,
Disease Ontology. |
Based on the KEGG correlation network, it was found
that top 15 pathways of targets of downregulated miRNAs play a role
in multiple cancer processes, such as prostate and lung cancer, and
the p53 signaling pathway. Top 15 pathways of targets of
downregulated miRNAs function in pathways associated with cancer
such as cell cycle and the MAPK signaling pathway, and ascorbate
and aldarate metabolism (Fig.
4B). These KEGG analyses suggested that altered miRNAs of pre-
and post-radiotherapy may function in different pathways.
Interactions between miRNAs and their predicted
targets were next investigated using DO analysis. The targets of
upregulated miRNAs were enriched in digestive system cancers such
as pancreas adenocarcinoma, pancreatic ductal adenocarcinoma and
pancreatic carcinoma (Figs. 4C
and S5; Table SXII). The targets of
downregulated miRNAs were enriched in central nervous system cancer
such as cerebral ventricle cancer, cerebrum cancer, synovial
sarcoma, malignant glioma, brain cancer and atypical teratoid
rhabdoid tumor (Figs. 4C and
S5; Table SXIII). These results suggested
the up- and downregulated plasma exosomal miRNAs play different
roles in cancer processes.
Cell migration and proliferation
analyses of miR-652 and miR-30a in TE-1 cells
Our miRNA sequencing and RT-qPCR results showed that
miR-652 and miR-30a are sensitive and responding to the
radiotherapy of ESCC. Previous studies have shown that miR-652
inhibits proliferation and invasion of ESCC by targeting FGFR1 and
other genes, and miR-30a may negatively regulates FoxD1 in ESCC
(38–41). Thus, miR-652 and miR-30a were
selected and their roles in migration in TE-1 cells, a human EC
cell line, were examined using wound-healing assays (31,42,43) (Fig.
1A).
The expression of miR-652 and miR-30a was altered by
overexpressing and knocking down in TE-1 cells (31). Wound-healings were detected at 0,
24 and 72 h after scratching (Fig.
5). Compared with those at 0 h, the normalized wound sizes of
the miR-652 overexpression group (miRNA mimics) were significantly
decreased by 23.28% at 24 h, and by 73.54% at 72 h (Fig. 5A and E). Conversely, the
normalized wound sizes of the miRNA inhibitor group were
significantly increased by 15.42% at 72 h (Fig. 5C and F). Moreover, to investigate
the miRNA effect in proliferation, a CCK-8 assay was performed in
TE-1 cells, which is widely used to examine cell growth or
proliferation (44,45). Proliferation of TE-1 cells was not
significantly changed by miR-652 overexpression and inhibition
(Fig. S6A and B). These results
suggested that altering miR-652 expression affects TE-1 cell
migration but not proliferation.
The potential role of miR-30a in migration and
proliferation (Fig. 5) was next
examined. Compared with the control group, the normalized wound
sizes of the miR-30a mimics group showed no change (Fig. 5B and E). Conversely, the
normalized wound sizes of the inhibitor group were significantly
decreased by 11.50% at 24 h and by 29.94% at 72 h (Fig. 5D and F). Proliferation of TE-1
cells was not significantly changed by miR-30a overexpression and
inhibition (Fig. S6C and D).
These results indicated that inhibition of miR-30a directly
prevents cancer cell migration in vitro.
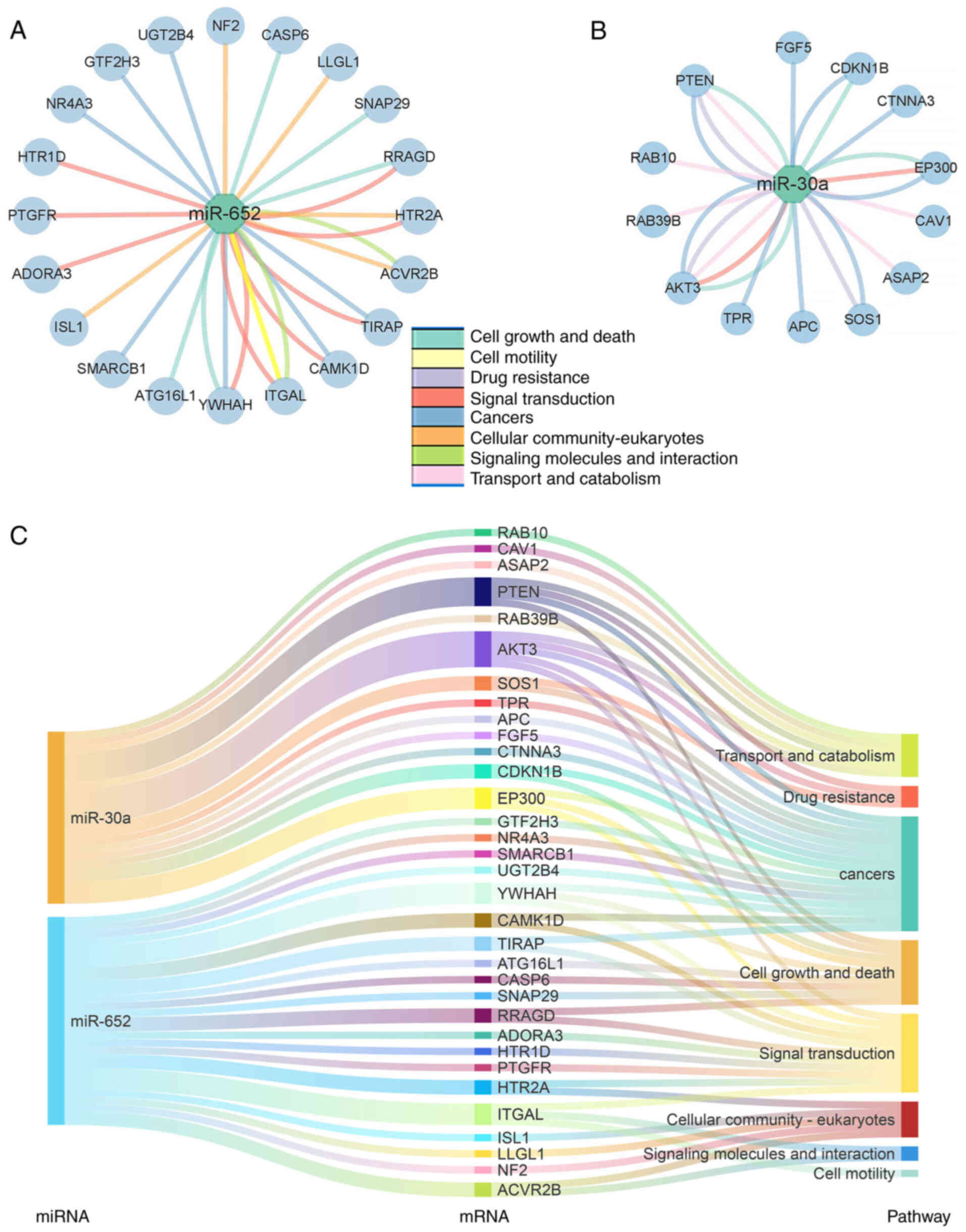
Potential roles of target genes of
miR-652 and miR-30a
It was found that the radiotherapy sensitive miR-652
and miR-30a may play roles in migration of TE-1 cells. Target genes
of miR-652 and miR-30a were next analyzed using the KEGG analysis.
A total of 8 different molecular processes of their targets
including cell growth and death, cell motility, drug resistance,
signal transduction, cancers, cellular community-eukaryotes,
signaling molecules and interaction and transport and catabolism
were chosen (Fig. 6). Among these
targets, 20 target genes of miR-652 and 13 target genes of miR-30a
were enriched in the 8 aforementioned molecular processes (Fig. 6A and B).
To visualize the potential role of predicted
targets, Sankey diagrams were constructed to illustrate a
relationship of genes, pathways and miRNAs (Fig. 6C). Certain previously demonstrated
target-miRNA pairs such as PTEN-miR30a, AKT3-miR30a,
EP300-miR652, YWHAH-miR652 and ITGAL-miR652 were readily
detected (Fig. 6C). These
gene-miRNA pairs may play an essential role in responding to
radiotherapy in ESCC.
Discussion
Radiotherapy is a major therapeutic way for ESCC
(46–48). Studies have shown that miRNAs
derived from plasma exosomes can be used to evaluate the curative
effect of radiotherapy (49–51). Previous studies have shown that
some specific miRNAs are sensitized by radiotherapy in ESCC
(52,53). In the present study, to identify
exosomal miRNAs that may be responding to radiotherapy in
non-metastatic ESCC, differentially expressed exosomal miRNAs were
analyzed in plasma samples of patients with pre- and
post-radiotherapy using miRNA sequencing and RT-qPCR. It was
identified that upregulated miRNAs play specific roles in
responding to radiation and DNA damage, and downregulated miRNAs
play roles in exosome transduction. Among them, upregulated miR-652
and downregulated miR-30a directly altered migration but not
proliferation of human EC cells in vitro. The present study
indicated that certain exosomal miRNAs are responding to
radiotherapy in ESCC patients, and provided new plasma biomarkers
for future evaluation of diagnosis, treatment and prognosis of
ESCC.
Accumulating studies have shown that circulating
exosomes are associated with cancer progression and therapeutic
reactions, for instance in ESSC (54–57). Moreover, radiation exposure
increases the levels of intracellular free radical species,
followed by DNA strand breaks and subsequent dysfunction of the
mitochondria, endoplasmic reticulum and other organelles (58). These radiation-induced cellular
events lead to activation of pro-apoptotic signaling and eventually
to tumor cell killing (59).
Thus, the present study aimed to identify miRNAs in exosomes that
may be responding to radiotherapy in ESSC. A total of 8 and 9
miRNAs with up- and downregulated expression in pre- and
post-radiotherapy plasma samples were identified, respectively.
Certain identified miRNAs in the present study have also been shown
as novel circulating factors in responding to radiotherapy in other
types of cancer (52,53,60–62). Moreover, a higher level of plasma
miR-339-5p, which was also identified in the present study, has
been shown to be associated with radiotherapy sensitivity and
favorable survival by targeting Cdc25A gene in ESCC
(53). Therefore, radiotherapy
causes significant alterations in expression levels of exosomal
miRNAs in ESCC plasma.
Furthermore, because miRNAs normally negatively
regulate expression levels of target genes, it was revealed that
targets of upregulated exosomal miRNAs are enriched in DNA damage
check point and cellular response to radiation, while targets of
downregulated miRNAs are enriched in endosome transport and
digestive system development. For instance, target genes for
upregulated miRNAs after radiotherapy are involved in the p53
signaling pathway and are enriched in digestive system cancers
based on the present study. These results suggested that radiation
sensitive miRNAs in plasma exosomes may be developed into
non-invasive biomarkers for evaluating the radiation-induced
esophageal toxicity and therapeutic effect in the future (63). More specimen and detailed target
examinations are required to uncover biomarkers of miRNA-target
pairs in the future studies.
Among identified exosomal miRNAs in the present
study, miR-652 and miR-30a have been demonstrated to regulate cell
migration in cancers of the digestive system by other laboratories
(64–67). It appears that miR-652 directly
inhibits tumor invasion by targeting FGFR1, PLD1 and other
genes (38,39). The present study also showed the
effect of miR-652 and miR-30a on cell migration but not
proliferation in TE-1 cells, an EC cell line. Moreover, our
miRNA-target pair analyses indicated that targets for miR-652 and
miR-30a are enriched in multiple biological and cellular processes,
which may correlate with cancer cell migration and invasion. In
addition, it is likely that miR-652 and miR-30a play a general role
in cancer development. Studies have shown that miR-652 executes a
tumor-promoter function in non-small cell lung cancer through
direct binding and regulating the expression of Lgl1
(68), and miR-30a plays a
central role in regulating the PTEN/AKT pathway in lung
adenocarcinoma and liver cancer cells (69,70). The present findings provided a
reference for further identification of miRNA-target interaction
networks in responding to radiotherapy of ESCC. The future studies
will be to validate miRNA-target regulations and to examine the
biological meanings of these interactions.
In summary, certain exosomal miRNAs sensitive to
radiotherapy were identified in the present study, which may be
developed into biomarkers for evaluation of the effect of
radiotherapy for human ESCC. Identifying and validating
interactions of miRNAs and their targets in responding to
radiotherapy will help us to design radiotherapy strategies in an
improved way and optimize treatment effects in the future.
Supplementary Material
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the Fundamental Research
Funds for the Central Universities (grant no. ZQN-1020), the
Scientific Research Funds of Huaqiao University (grant nos.
16BS815, 19BS303 and Z16Y0017), the Natural Science Foundation of
Fujian, China (grant nos. 2019J01071 and 605-52519051), the
Innovation Awards of Quanzhou Talents (grant no. 2018C057R),
Quanzhou City Science & Technology Program of China (grant no.
2018C057R) and the National Natural Science Foundation of China
(grant nos. 31771141 and 32100775).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
TS conceived and designed the experiments. WCa and
SD performed sample collection. NM, SD, WCh and WCa conducted RNA
extraction and analysis of results. NM, WCh and YL performed
bioinformatics analysis. NM and TS wrote and edited the manuscript.
NM and TS confirm the authenticity of all the raw data. All authors
have read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved [approval no.
(2018)101] by the Institution Ethic Issue Committee of the First
Hospital of Quanzhou (Quanzhou, China). Signed informed consents
were obtained from female patients with ESCC who received
radiotherapy diagnosis.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kelsen D: Principles and practice of
gastrointestinal oncology. Lippincott. Williams & Wilkins;
Philadelphia, PA: 2008
|
|
2
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lin Y, Totsuka Y, He Y, Kikuchi S, Qiao Y,
Ueda J, Wei W, Inoue M and Tanaka H: Epidemiology of esophageal
cancer in Japan and China. J Epidemiol. 23:233–242. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zheng RS, Sun KX, Zhang SW, Zeng HM, Zou
XN, Chen R, Gu XY, Wei WW and He J: Report of cancer epidemiology
in China, 2015. Zhonghua Zhong Liu Za Zhi. 41:19–28. 2019.(In
Chinese). PubMed/NCBI
|
|
5
|
He Z and Ke Y: Precision screening for
esophageal squamous cell carcinoma in China. Chin J Cancer Res.
32:673–682. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Malhotra A, Sharma U, Puhan S, Bandari NC,
Kharb A, Arifa PP, Thakur L, Prakash H, Vasquez KM and Jain A:
Stabilization of miRNAs in esophageal cancer contributes to
radioresistance and limits efficacy of therapy. Biochimie.
156:148–157. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Klug F, Prakash H, Huber PE, Seibel T,
Bender N, Halama N, Pfirschke C, Voss RH, Timke C, Umansky L, et
al: Low-dose irradiation programs macrophage differentiation to an
iNOS(+)/M1 phenotype that orchestrates effective T cell
immunotherapy. Cancer Cell. 24:589–602. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu J, Xue N, Guo Y, Niu K, Gao L, Zhang
S, Gu H, Wang X, Zhao D and Fan R: CircRNA_100367 regulated the
radiation sensitivity of esophageal squamous cell carcinomas
through miR-217/Wnt3 pathway. Aging (Albany NY). 11:12412–12427.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang YH, Wang QQ, Li H, Ye T, Gao F and
Liu YC: miR-124 radiosensitizes human esophageal cancer cell TE-1
by targeting CDK4. Genet Mol Res. 15:150278932016.
|
|
10
|
Chen Z, Hu X, Wu Y, Cong L, He X, Lu J,
Feng J and Liu D: Long non-coding RNA XIST promotes the development
of esophageal cancer by sponging miR-494 to regulate CDK6
expression. Biomed Pharmacother. 109:2228–2236. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mathieu M, Martin-Jaular L, Lavieu G and
Thery C: Specificities of secretion and uptake of exosomes and
other extracellular vesicles for cell-to-cell communication. Nat
Cell Biol. 21:9–17. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wei Z, Batagov AO, Schinelli S, Wang J,
Wang Y, Fatimy RE, Rabinovsky R, Balaj L, Chen CC, Hochberg F, et
al: Coding and noncoding landscape of extracellular RNA released by
human glioma stem cells. Nat Commun. 8:11452017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang H, Freitas D, Kim HS, Fabijanic K,
Li Z, Chen H, Mark MT, Molina H, Martin AB, Bojmar L, et al:
Identification of distinct nanoparticles and subsets of
extracellular vesicles by asymmetric flow field-flow fractionation.
Nat Cell Biol. 20:332–343. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Balandeh E, Mohammadshafie K, Mahmoudi Y,
Pourhanifeh MH, Rajabi A, Bahabadi ZR, Mohammadi AH, Rahimian N,
Hamblin MR and Mirzaei H: Roles of non-coding RNAs and angiogenesis
in glioblastoma. Front Cell Dev Biol. 9:7164622021. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang J, Li S, Li L, Guo C, Yao J and Mi
S: Exosome and exosomal microRNA: Trafficking, sorting, and
function. Genomics Proteomics Bioinformatics. 13:17–24. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Costa-Silva B, Aiello NM, Ocean AJ, Singh
S, Zhang H, Thakur BK, Becker A, Hoshino A, Mark MT, Molina H, et
al: Pancreatic cancer exosomes initiate pre-metastatic niche
formation in the liver. Nat Cell Biol. 17:816–826. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Peinado H, ković MA, Lavotshkin S, Matei
I, Costa-Silva B, Moreno-Bueno G, Hergueta-Redondo M, Williams C,
García-Santos G, Ghajar CM, et al: Corrigendum: Melanoma exosomes
educate bone marrow progenitor cells toward a pro-metastatic
phenotype through MET. Nat Med. 22:15022016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Peinado H, Aleckovic M, Lavotshkin S,
Matei I, Costa-Silva B, Moreno-Bueno G, Hergueta-Redondo M,
Williams C, García-Santos G, Ghajar C, et al: Melanoma exosomes
educate bone marrow progenitor cells toward a pro-metastatic
phenotype through MET. Nat Med. 18:883–891. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hoshino A, Costa-Silva B, Shen TL,
Rodrigues G, Hashimoto A, Mark MT, Molina H, Kohsaka S, Giannatale
AD, Ceder S, et al: Tumour exosome integrins determine organotropic
metastasis. Nature. 527:329–335. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Skog J, Wurdinger T, van Rijn S, Meijer
DH, Gainche L, Sena-Esteves M, Curry WT Jr, Carter BS, Krichevsky
AM and Breakefield XO: Glioblastoma microvesicles transport RNA and
proteins that promote tumour growth and provide diagnostic
biomarkers. Nat Cell Biol. 10:1470–1476. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Valadi H, Ekstrom K, Bossios A, Sjöstrand
M, Lee JJ and Lötvall JO: Exosome-mediated transfer of mRNAs and
microRNAs is a novel mechanism of genetic exchange between cells.
Nat Cell Biol. 9:654–659. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang J, Zhang X, Cao J, Xu P, Chen Z, Wang
S, Li B, Zhang L, Xie L, Fang L and Xu Z: Circular RNA UBE2Q2
promotes malignant progression of gastric cancer by regulating
signal transducer and activator of transcription 3-mediated
autophagy and glycolysis. Cell Death Dis. 12:9102021. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cooks T, Pateras IS, Jenkins LM, Patel KM,
Robles AI, Morris J, Forshew T, Appella E, Gorgoulis VG and Harris
CC: Mutant p53 cancers reprogram macrophages to tumor supporting
macrophages via exosomal miR-1246. Nat Commun. 9:7712018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen F, Chu L, Li J, Shi Y, Xu B, Gu J,
Yao X, Tian M, Yang X and Sun X: Hypoxia induced changes in miRNAs
and their target mRNAs in extracellular vesicles of esophageal
squamous cancer cells. Thorac Cancer. 11:570–580. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen F, Xu B, Li J, Yang X, Gu J, Yao X
and Sun X: Hypoxic tumour cell-derived exosomal miR-340-5p promotes
radioresistance of oesophageal squamous cell carcinoma via KLF10. J
Exp Clin Cancer Res. 40:382021. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Butz F, Eichelmann AK, Mayne GC, Wang T,
Bastian I, Chiam K, Marri S, Sykes PJ, Wijnhoven BP, Toxopeus E, et
al: MicroRNA profiling in oesophageal adenocarcinoma cell lines and
patient serum samples reveals a role for miR-451a in radiation
resistance. Int J Mol Sci. 21:88982020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zheng Y, Campbell EC, Lucocq J, Riches A
and Powis SJ: Monitoring the Rab27 associated exosome pathway using
nanoparticle tracking analysis. Exp Cell Res. 319:1706–1713. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang W, Peng P, Kuang Y, Yang J, Cao D,
You Y and Shen K: Characterization of exosomes derived from ovarian
cancer cells and normal ovarian epithelial cells by nanoparticle
tracking analysis. Tumour Biol. 37:4213–4221. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Simpson RJ, Jensen SS and Lim JW:
Proteomic profiling of exosomes: Current perspectives. Proteomics.
8:4083–4099. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Schneider CA, Rasband WS and Eliceiri KW:
NIH image to imageJ: 25 years of image analysis. Nat Methods.
9:671–675. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li BQ XJ, Li QZ, Li XY, Gu FZ and Mu XL: A
study on cell biological characters of human esophageal cancer cell
line TE-1. Chin J Mod Med. 13:33–35. 2011.(In Chinese).
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Fonseka P, Pathan M, Chitti SV, Kang T and
Mathivanan S: FunRich enables enrichment analysis of OMICs
datasets. J Mol Biol. 433:1667472021. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Otasek D, Morris JH, Boucas J, Pico AR and
Demchak B: Cytoscape automation: Empowering workflow-based network
analysis. Genome Biol. 20:1852019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Feng H, Hu YY, Jin P, Meng XK, Chen YB and
Zhang HM: Intensity-modulated radiotherapy combined with iodine-125
seed implantation in non-central recurrence of cervical cancer: A
case report and literature review. Oncol Letters. 14:4085–4091.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jeppesen DK, Fenix AM, Franklin JL,
Higginbotham JN, Zhang Q, Zimmerman LJ, Liebler DC, Ping J, Liu Q,
Evans R, et al: Reassessment of exosome composition. Cell.
177:428–445.e18. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kowal J, Arras G, Colombo M, Jouve M,
Morath JP, Primdal-Bengtson B, Dingli F, Loew D, Tkach M and Théry
C: Proteomic comparison defines novel markers to characterize
heterogeneous populations of extracellular vesicle subtypes. Proc
Natl Acad Sci USA. 113:E968–E977. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lu M, Song Y, Fu W, Liu Y, Huai S, Cui X,
Pang L, Yang L and Wei Y: MicroRNA and target mRNA selection
through invasion and cytotoxicity cell modeling and bioinformatics
approaches in esophageal squamous cell carcinoma. Oncol Rep.
38:1181–1189. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhen C, Huang J and Lu J: MicroRNA-652
inhibits the biological characteristics of esophageal squamous cell
carcinoma by directly targeting fibroblast growth factor receptor
1. Exp Ther Med. 18:4473–4480. 2019.PubMed/NCBI
|
|
40
|
Yan Q, Liu L, Yang H, Xu C, Wang Z, Wang
Q, Wu Z, Wu C, Dong L, Wang J and Wu M: Long non-coding RNA
OIP5-AS1 inhibits the proliferation and migration of esophageal
squamous carcinoma cells by targeting FOXD1/miR-30a-5p axis and the
effect of micro- and nano-particles on targeting transfection
system. J Biomed Nanotechnol. 17:1380–1391. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wada M, Goto Y, Tanaka T, Okada R, Moriya
S, Idichi T, Noda M, Sasaki K, Kita Y, Kurahara H, et al: RNA
sequencing-based microRNA expression signature in esophageal
squamous cell carcinoma: oncogenic targets by antitumor miR-143-5p
and miR-143-3p regulation. J Hum Genet. 65:1019–1034. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Duff D and Long A: Roles for RACK1 in
cancer cell migration and invasion. Cell Signal. 35:250–255. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Rodriguez LG, Wu X and Guan JL:
Wound-healing assay. Methods Mol Biol. 294:23–29. 2005.PubMed/NCBI
|
|
44
|
Wang F, Chen TS, Xing D, Wang JJ and Wu
YX: Measuring dynamics of caspase-3 activity in living cells using
FRET technique during apoptosis induced by high fluence low-power
laser irradiation. Lasers Surg Med. 36:2–7. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Morita Y, Naka T, Kawazoe Y, Fujimoto M,
Narazaki M, Nakagawa R, Fukuyama H, Nagata S and Kishimoto T:
Signals transducers and activators of transcription (STAT)-induced
STAT inhibitor-1 (SSI-1)/suppressor of cytokine signaling-1
(SOCS-1) suppresses tumor necrosis factor alpha-induced cell death
in fibroblasts. Proc Natl Acad Sci USA. 97:5405–5410. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chan WWL, Lam KO and Kwong DLW:
Radiotherapy for thoracic esophageal squamous cell carcinoma.
Methods Mol Biol. 2129:307–319. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zou B, Tu Y, Liao D, Xu Y, Wang J, Huang
M, Ren L, Zhu J, Gong Y, Liu Y, et al: Radical esophagectomy for
stage II and III thoracic esophageal squamous cell carcinoma
followed by adjuvant radiotherapy with or without chemotherapy:
Which is more beneficial? Thorac Cancer. 11:631–639. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Deng W, Yang J, Ni W, Li C, Chang X, Han
W, Zhou Z, Chen D, Feng Q, Liang J, et al: Postoperative
radiotherapy in pathological T2-3N0M0 thoracic esophageal squamous
cell carcinoma: Interim report of a prospective, phase III,
randomized controlled study. Oncologist. 25:e701–e708. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Al-Mayah AH, Irons SL, Pink RC, Carter DR
and Kadhim MA: Possible role of exosomes containing RNA in
mediating nontargeted effect of ionizing radiation. Radiat Res.
177:539–545. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Su LL, Chang XJ, Zhou HD, Hou LB and Xue
XY: Exosomes in esophageal cancer: A review on tumorigenesis,
diagnosis and therapeutic potential. World J Clin Cases. 7:908–916.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Jin X, Chen Y, Chen H, Fei S, Chen D, Cai
X, Liu L, Lin B, Su H, Zhao L, et al: Evaluation of tumor-derived
exosomal miRNA as potential diagnostic biomarkers for early-stage
non-small cell lung cancer using next-generation sequencing. Clin
Cancer Res. 23:5311–5319. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Luo A, Zhou X, Shi X, Zhao Y, Men Y, Chang
X, Chen H, Ding F, Li Y, Su D, et al: Exosome-derived miR-339-5p
mediates radiosensitivity by targeting Cdc25A in locally advanced
esophageal squamous cell carcinoma. Oncogene. 38:4990–5006. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Sun Y, Wang J, Ma Y, Li J, Sun X, Zhao X,
Shi X, Hu Y, Qu F and Zhang X: Radiation induces NORAD expression
to promote ESCC radiotherapy resistance via EEPD1/ATR/Chk1
signalling and by inhibiting pri-miR-199a1 processing and the
exosomal transfer of miR-199a-5p. J Exp Clin Cancer Res.
40:3062021. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zhu L, Zhao L, Wang Q, Zhong S, Guo X, Zhu
Y, Bao J, Xu K and Liu S: Circulating exosomal miRNAs and cancer
early diagnosis. Clin Transl Oncol. 24:393–406. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Lv J, Zhao HP, Dai K, Cheng Y, Zhang J and
Guo L: Circulating exosomal miRNAs as potential biomarkers for
Barrett's esophagus and esophageal adenocarcinoma. World J
Gastroenterol. 26:2889–2901. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Avgeris M, Panoutsopoulou K, Papadimitriou
MA and Scorilas A: Circulating exosomal miRNAs: Clinical
significance in human cancers. Expert Rev Mol Diagn. 19:979–995.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Schwarzenbach H: The clinical relevance of
circulating, exosomal miRNAs as biomarkers for cancer. Expert Rev
Mol Diagn. 15:1159–1169. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Skladanowski A, Bozko P and Sabisz M: DNA
structure and integrity checkpoints during the cell cycle and their
role in drug targeting and sensitivity of tumor cells to anticancer
treatment. Chem Rev. 109:2951–2973. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Koukourakis MI: Radiation damage and
radioprotectants: New concepts in the era of molecular medicine.
Brit J Radiol. 85:313–330. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Li MH, Zou X, Xia T, Wang T, Liu P, Zhou
X, Wang S and Zhu W: A five-miRNA panel in plasma was identified
for breast cancer diagnosis. Cancer Med. 8:7006–7017. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Tang Y, Cui Y, Li Z, Jiao Z, Zhang Y, He
Y, Chen G, Zhou Q, Wang W, Zhou X, et al: Radiation-induced
miR-208a increases the proliferation and radioresistance by
targeting p21 in human lung cancer cells. J Exp Clin Canc Res.
35:72016. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Ham IH, Lee D and Hur H: Cancer-associated
fibroblast-induced resistance to chemotherapy and radiotherapy in
gastrointestinal cancers. Cancers (Basel). 13:11722021. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Xu T, Liao ZX, O'Reilly MS, Levy LB, Welsh
JW, Wang LE, Lin SH, Komaki R, Liu Z, Wei Q and Gomez DR: Serum
inflammatory miRNAs predict radiation esophagitis in patients
receiving definitive radiochemotherapy for non-small cell lung
cancer. Radiother Oncol. 113:379–384. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Zhang J, Qiu WQ, Zhu H, Liu H, Sun JH,
Chen Y, Shen H, Qian CL and Shen ZY: HOTAIR contributes to the
carcinogenesis of gastric cancer via modulating cellular and
exosomal miRNAs level. Cell Death Dis. 11:7802020. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Yao W, Guo P, Mu Q and Wang Y:
Exosome-derived Circ-PVT1 contributes to cisplatin resistance by
regulating autophagy, invasion, and apoptosis via miR-30a-5p/YAP1
axis in gastric cancer cells. Cancer Biother Radiopharm.
36:347–359. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Kulkarni B, Gondaliya P, Kirave P, Rawal
R, Jain A, Garg R and Kalia K: Exosome-mediated delivery of miR-30a
sensitize cisplatin-resistant variant of oral squamous carcinoma
cells via modulating Beclin1 and Bcl2. Oncotarget. 11:1832–1845.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Gao P, Wang D, Liu M, Chen S, Yang Z,
Zhang J, Wang H, Niu Y, Wang W, Yang J and Sun G: DNA
methylation-mediated repression of exosomal miR-652-5p expression
promotes oesophageal squamous cell carcinoma aggressiveness by
targeting PARG and VEGF pathways. PLoS Genet. 16:e10085922020.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Yang WH, Zhou CC, Luo M, Shi X, Li Y, Sun
Z, Zhou F, Chen Z and He J: MiR-652-3p is upregulated in non-small
cell lung cancer and promotes proliferation and metastasis by
directly targeting Lgl1. Oncotarget. 7:16703–16715. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Li WF, Dai H, Ou Q, Zuo GQ and Liu CA:
Overexpression of microRNA-30a-5p inhibits liver cancer cell
proliferation and induces apoptosis by targeting MTDH/PTEN/AKT
pathway. Tumor Biol. 37:5885–5895. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Sui J, Yang RS, Xu SY, Zhang YQ, Li CY,
Yang S, Yin LH, Pu YP and Liang GY: Comprehensive analysis of
aberrantly expressed microRNA profiles reveals potential biomarkers
of human lung adenocarcinoma progression. Oncol Rep. 38:2453–2463.
2017. View Article : Google Scholar : PubMed/NCBI
|