Introduction
Psoriasis is a complex autoimmune skin disorder that
affects ~2% of the global population, posing notable health
challenges due to its chronic nature and impact on quality of life
(1). In Taiwan, the prevalence of
psoriasis is notably lower, affecting ~0.24% of the population;
however, it remains a concern due to the debilitating symptoms it
can cause (2). The clinical
manifestations of psoriasis include scaling, thickened skin
(acanthosis) and the formation of distinct psoriatic plaques. This
condition is primarily driven by an overactive immune response,
particularly involving T-helper 17 (Th17) cells, which are crucial
in the proliferation of keratinocytes and the production of
pro-inflammatory cytokines (3).
IL-17A, secreted by Th17 cells, can induce C/CAAT-enhancer-binding
proteins (C/EBPs) that are translocated into the nucleus, and can
increase transcript levels of chemokines and inflammatory
cytokines, such as CXCL8 and IL-6 (4). Abnormal crosstalk between
keratinocytes and immune cells has been considered the main driver
of skin inflammation.
Heat shock proteins (HSPs), which are critical for
protein folding, assembly and intracellular trafficking, also serve
a role in cellular responses to stress, such as heat, oxidative
conditions or nutrient deprivation (5). When cells are exposed to high
temperature, oxidative stress or energy shortage, HSP expression is
increased to rescue cell damage. Specifically targeting the
regulation of HSP expression has been reported to protect against
inflammatory diseases including neuro-inflammation, hepatic
inflammation and psoriasis (6–8).
However, the role of HSP 70 in psoriasis remains controversial; for
example, HSP 70 has been shown to be released from exosomes and to
activate dendritic cells to secrete proinflammatory cytokines,
which may then induce overproliferation of keratinocytes (9). By contrast, a different study
reported that HSP 70 reduces psoriasis-like inflammation induced by
imiquimod (IMQ) in mice (8).
Brevilin A is also known to target multiple
pathways, including the JAK/STAT signaling pathway, which is
pivotal in immune response regulation, and has been implicated in
both inflammatory disease mechanisms and oncogenesis (10,11).
However, whether brevilin A inhibits skin inflammation remains to
be determined. Given its broad anti-inflammatory and
antiproliferative properties, the present study aimed to
investigate the effects of brevilin A on the regulation of
pro-inflammatory cytokines and to elucidate its underlying
mechanisms in the context of psoriasis, potentially offering a
novel therapeutic approach to managing this persistent skin
disorder.
Materials and methods
Cell culture and treatment
The human keratinocyte HaCaT cell line (Elabscience
Bionovation Inc.) was cultured in a Dulbecco's Modified Eagle's
Medium (cat. no. SH30022.02; HyClone; Cytiva) supplemented with 10%
fetal bovine serum (cat. no. SH30396.03; HyClone; Cytiva) and 1%
penicillin-streptomycin (Gibco; Thermo Fisher Scientific, Inc.).
Cells were incubated at 37°C in a humidified atmosphere containing
5% CO2. For treatment, cells were exposed to brevilin A
(CAS no. 16503-32-5; Chengdu Biopurify Phytochemicals Ltd.) at
concentrations of 1.25, 2.5 and 5 µM at 37°C for 9 or 24 h. IL-17A
(Peprotech, Inc.) was applied at a concentration of 100 ng/ml to
stimulate cells in co-treatment assays.
MTT assay for cell viability
Cell viability was assessed using the MTT assay (CAS
no. 298-93-1; Sigma-Aldrich; Merck KGaA). Briefly, HaCaT cells were
seeded at a density of 5×104 cells/well in a 96-well
plate, and were treated with the indicated concentrations of
brevilin A (0.625–20 µM) and/or apoptozole (CAS no. 1054543-47-3;
MedChemExpress) (0.009–20 µM) at 37°C for 24 h. Each concentration
of each substance was tested in triplicate wells (n=3). After 24 h
of treatment, MTT solution (5 mg/ml) was added and the cells were
incubated for an additional 4 h. Formazan crystals formed were
dissolved in isopropanol, and the absorbance was measured at 570 nm
using a microplate reader (SUNRISE; Tecan Group, Ltd).
ELISA for cytokine measurement
The levels of IL-6 (cat. no. 430504) and IL-8 (cat.
no. 431504) in the supernatant were measured using ELISA kits
(Biolegend, Inc.) according to the manufacturer's protocols. HaCaT
cells were seeded at a density of 2×105 cells/well in a
24-well plate. Samples from HaCaT cells were collected at 9 and 24
h following co-treatment with IL-17A (100 ng/ml) (12) and either brevilin A (1.25, 2.5 and
5 µM) or apoptozole (1.25, 2.5 and 5 µM). Dexamethasone (1 µg/ml;
Taiwan Biotech Co., Ltd.) was used as a positive control. Each
experimental condition was replicated in three wells to confirm the
consistency of the results.
Western blot analysis
Western blot analysis was performed to assess the
expression levels of HSP 70 and HSP 90 in HaCaT cells treated with
specific agents. The cells were harvested and lysed using RIPA
buffer (Bio Basic Inc.), which contains Tris-HCl (20 mM, pH 7.4),
NaCl (150 mM), NP-40 (1%), sodium deoxycholate (0.5% w/v), SDS
(0.1%), EDTA (1 mM) and EGTA (5 mM), with added protease
inhibitors. After incubation on ice for 30 min, the lysates were
centrifuged at 14,000 × g for 15 min at 4°C to eliminate cell
debris. The protein-containing supernatants were collected, and
protein concentrations were measured using a BCA protein assay kit
(Thermo Fisher Scientific, Inc.). The proteins (30 µg/lane) were
then separated by SDS-PAGE on 10% gels and transferred onto PVDF
membranes. These membranes were blocked with 5% non-fat milk
dissolved in PBS at room temperature for 1 h and were then probed
with primary antibodies for HSP 70 (1:1,000; cat. no. sc-32239;
Santa Cruz Biotechnology, Inc.), HSP 90 (1:1,000; cat. no.
sc-69703; Santa Cruz Biotechnology, Inc.), phosphorylated (p-)p38
(1:1,000; cat. no. 4511; Cell Signaling Technology, Inc.), p38
(1:1,000; cat. no. 8690; Cell Signaling Technology, Inc.), p-ERK
(1:1,000; cat no. 4370; Cell Signaling Technology, Inc.), ERK
(1:1,000; cat. no. 4695; Cell Signaling Technology, Inc.), p-JNK
(1:2,000; cat. no. 4668; Cell Signaling Technology, Inc.), JNK
(1:1,000; cat. no. 9252; Cell Signaling Technology, Inc.) and
β-actin (loading control; 1:5,000; cat. no. 3700; Cell Signaling
Technology, Inc.). Detection was performed using HRP-conjugated
anti-mouse IgG (1:7,000; cat. no. 7076; Cell Signaling Technology,
Inc.) or anti-rabbit IgG (1:7,000; cat. no. 7074; Cell Signaling
Technology, Inc.) secondary antibodies at room temperature for 1 h,
followed by enhanced chemiluminescence (MilliporeSigma). Western
blots were semi-quantified using ImageJ (version 1.53e; National
Institutes of Health).
In vivo psoriasis model
BALB/c female mice (n=30; age, 8 weeks; weight, 20±2
g), sourced from the National Laboratory Animal Center (Taipei,
Taiwan), were maintained under specific pathogen-free conditions at
a steady temperature of 23±1°C, 50–60% relative humidity and a 12-h
light/dark cycle, and had free access to food and water. The animal
study protocols were approved by the Animal Care and Use Committee
of Kaohsiung Veterans General Hospital (IACUC no. 2021-A039;
Kaohsiung, Taiwan). The mice were allocated into six groups
(n=5/group): Control, DMSO (vehicle control), brevilin A (5, 10 and
20 mg/kg) and dexamethasone (1 mg/kg) (12). All groups, with the exception of
the control group, received a daily topical application of 62.5 mg
5% IMQ cream (Aldara; 3M Pharmaceuticals) on their shaved backs and
right ears for 5 consecutive days. The DMSO group received daily
intraperitoneal injections of 0.5% DMSO in 200 µl normal saline.
The brevilin A groups were administered daily intraperitoneal
injections of brevilin A at their respective doses in 200 µl normal
saline, whereas the dexamethasone group received daily injections
of 1 mg/kg dexamethasone in 200 µl normal saline. On day 6, all
mice were euthanized for subsequent analyses. The procedure
involved placing the mice in a chamber gradually filled with
CO2 at a flow rate of 30% chamber volume/min. After the
gas was introduced, the mice were observed for 3 min to ensure
complete euthanasia, with death confirmed by the absence of a
heartbeat, cessation of respiratory movements and pupil dilation.
Following euthanasia, skin samples were harvested for histological
and immunohistochemical analysis to evaluate epithelial
hyperproliferation and HSP 70 expression.
Histological analysis and
immunohistochemistry
The dorsal skin from each mouse was removed, fixed
in 10% formalin at room temperature for 24 h, embedded in paraffin,
and sectioned into 5-µm slices. These sections were subsequently
stained with hematoxylin and eosin (H&E) at room temperature to
examine epithelial hyperproliferation and the infiltration of
immune cells. Tissue sections were deparaffinized in xylene (three
changes, 5 min each) and rehydrated through a graded series of
ethanol: 100% (1 min), 95% (1 min), 80% (1 min) and 70% (1 min),
followed by a 1-min rinse in distilled water. Sections were then
stained with hematoxylin for 5 min, washed in water for another 5
min, and counterstained with eosin Y for 5 min. Subsequently, the
sections were dehydrated in 95% ethanol (10 sec), followed by four
changes of 100% ethanol (10 sec for the first change and 1 min for
the subsequent changes). Finally, sections were cleared in xylene
(three changes, 5 min each) and mounted with a coverslip.
For immunohistochemistry (IHC), tissue sections were
deparaffinized in xylene (two changes, 10 min each) and rehydrated
through a graded series of ethanol: 100, 95, 80 and 70%, followed
by a 5-min rinse in tap water. Antigen retrieval was performed
using heat-induced epitope retrieval at 95°C for 30 min with either
citrate buffer (cat. no. CBB500; Scytek Laboratories, Inc.) for HSP
70 and IL-6, or Tris-EDTA buffer (cat. no. TES999; Scytek
Laboratories, Inc.) for HSP 90. After antigen retrieval, the
sections were washed in TBS-0.05% Tween 20 (TBST; cat. no. TBT999;
Scytek Laboratories, Inc.) for 5 min. To block endogenous
peroxidase activity, sections were incubated in Hydrogen Peroxide
Block (cat. no. TA-060-HP; Epredia; Thermo Fisher Scientific, Inc.)
for 10 min at room temperature, followed by two TBST washes (3 min
each). Non-specific binding was blocked using either MS Blocking A
and B (cat. no. D52; OriGene Technologies, Inc.) or Protein Block
(cat. no. TA-060-PBQ; Epredia; Thermo Fisher Scientific, Inc.) for
10–30 min at room temperature. Primary antibody incubation was
performed in a humidified chamber at room temperature for 50 min in
the dark. The following primary antibodies were used: HSP 70 (1:50;
cat. no. sc-32239; Santa Cruz Biotechnology, Inc.), IL-6 (1:50;
cat. no. ab9324; Abcam) and HSP 90 (1:300; cat. no. 4877; Cell
Signaling Technology, Inc.), diluted in antibody dilution buffer
(cat. no. ADB250; Roche Tissue Diagnostics). After primary antibody
incubation, the sections were washed twice in TBST (3 min each).
For signal amplification, either Mouse Antibody Enhancer (cat. no.
D52; OriGene Technologies, Inc.) or Rabbit Antibody Enhancer (cat.
no. D39; OriGene Technologies, Inc.) was applied for 15 min,
followed by another two washes with TBST (3 min each). Detection
was performed using HRP-conjugated anti-mouse (cat. no. D52;
OriGene Technologies, Inc.) or anti-rabbit (cat. no. D39; OriGene
Technologies, Inc.) secondary antibodies, each incubated for 15 min
at room temperature. After two additional TBST washes (3 min each),
the signal was developed using DAB (cat. no. RE7270-K; Novolink;
Leica Biosystems) for 5 min. The sections were rinsed in tap water
for 5 min and then counterstained with hematoxylin (cat. no.
3801522; Leica Biosystems) for 2 min. After a final wash with tap
water (5 min), the sections were dehydrated in 100% ethanol (two
changes, 5 min each) and cleared in xylene (two changes, 5 min
each). Finally, the slides were mounted using Surgipath Micromount
mounting medium (cat. no. 3801731; Leica Biosystems).
Images of the H&E- and IHC-stained sections were
captured using an APEXVIEW APX100 Digital Imaging System (Olympus
Corporation) and a light microscope, and were analyzed with ImageJ
software (version 1.5.3; National Institutes of Health). For HSP
70, HSP 90 and IL-6 staining analysis, tissue samples were stained,
and 3–5 regions of the same size were randomly selected from each
sample for semi-quantification. The average staining intensity for
each group was calculated, and statistical differences between
groups were analyzed using appropriate statistical methods. This
approach ensured that the quantification was unbiased, reproducible
and reflective of the staining patterns across the samples.
Statistical analysis
Data were analyzed using GraphPad Prism software
(version: 6.01, Dotmatics). Differences between groups were
evaluated using one-way ANOVA followed by Tukey's post hoc test for
multiple comparisons. P<0.05 was considered to indicate a
statistically significant difference.
Results
Brevilin A inhibits IL-17A-induced
IL-6 and IL-8 levels
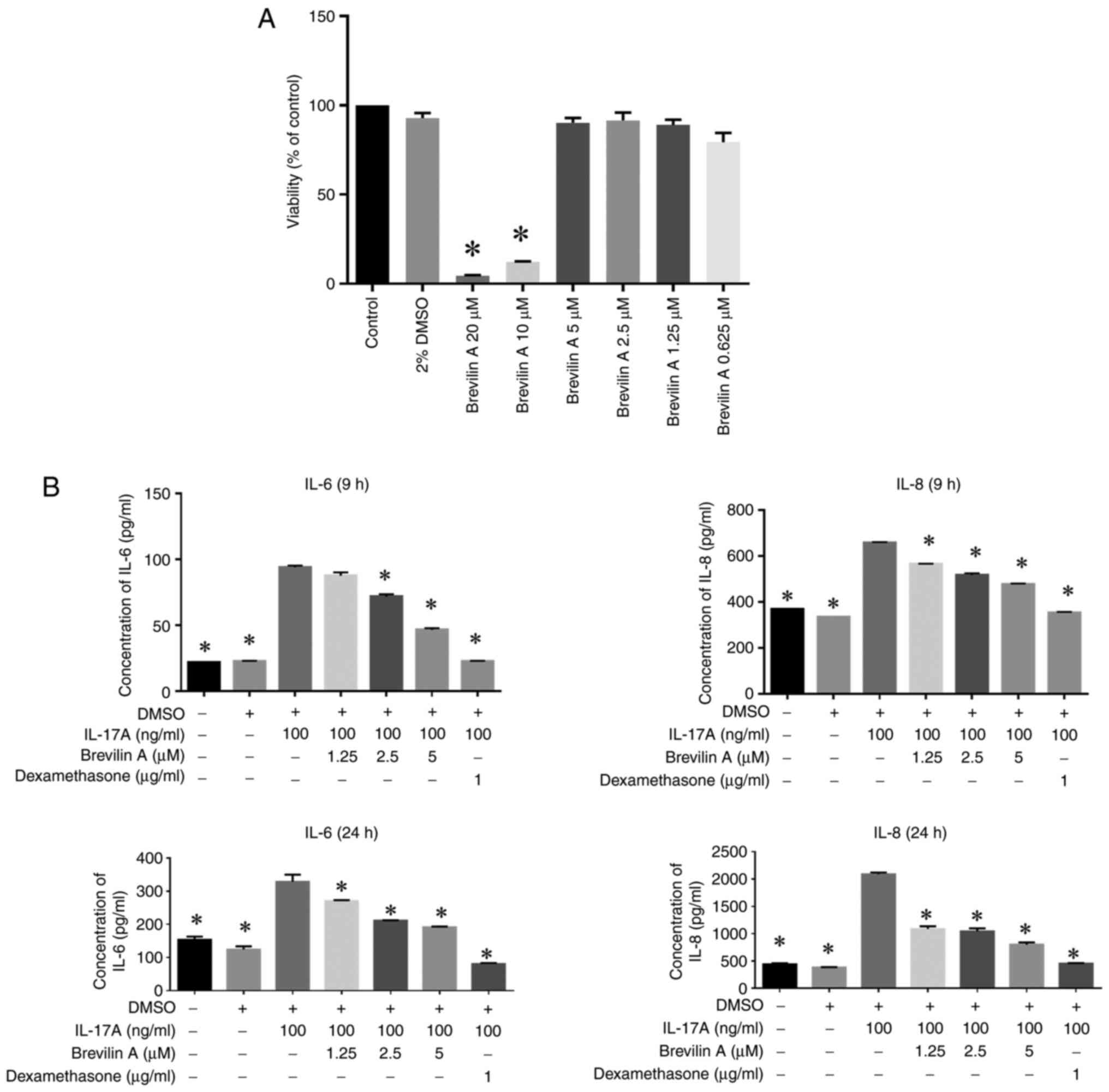
To evaluate the cytotoxicity and anti-inflammatory
effects of brevilin A on HaCaT cells, an MTT assay and ELISA were
performed. The results demonstrated that brevilin A exhibited
cytotoxic effects at higher concentrations, since 10 and 20 µM
significantly reduced cell viability compared with that in the
control group, whereas lower concentrations (0.625, 1.25, 2.5, and
5 µM) had no significant impact on viability (Fig. 1A). Additionally, brevilin A
effectively suppressed IL-17A-induced pro-inflammatory cytokine
production in HaCaT cells in a dose-dependent manner. At both 9 and
24 h, brevilin A significantly reduced the levels of IL-6 and IL-8,
with the effects being comparable to or exceeding those of
dexamethasone (12), a positive
control (Fig. 1B). These findings
indicated the potential of brevilin A as an anti-inflammatory agent
capable of modulating keratinocyte responses in inflammatory
conditions.
Brevilin A increases HSP 70 and HSP 90
expression in keratinocytes
HSPs regulate inflammatory responses in autoimmune
diseases and are upregulated during inflammation (13). However, the specific function of
HSPs in skin inflammation remains ambiguous. In the current study,
the influence of brevilin A on the expression levels of HSP 70 and
HSP 90 in keratinocytes was examined. Initially, HaCaT cells were
co-treated with IL-17A (100 ng/ml) and 5 µM brevilin A. The results
showed that HSP 70 and HSP 90 experssion levels were significantly
upregulated after co-culture with IL-17A and 5 µM brevilin A for 6
and 9 h (Fig. 2A-C). Subsequently,
to explore dose-dependent effects, HaCaT cells were treated with
different doses of brevilin A (1.25, 2.5 and 5 µM) for 9 h. The
results showed that the expression levels of HSP 70 and HSP 90 were
significantly increased after stimulation with 5 µM brevilin A
(Fig. 2D and E). The current study
also investigated whether brevilin A alone induces HSP 70 and HSP
90 expression. HaCaT cells were treated with different doses of
brevilin A for 9 h, and it was revealed that brevilin A (5 µM)
alone was sufficient to significantly elevate HSP 70 and HSP 90
levels (Fig. 2F and G).
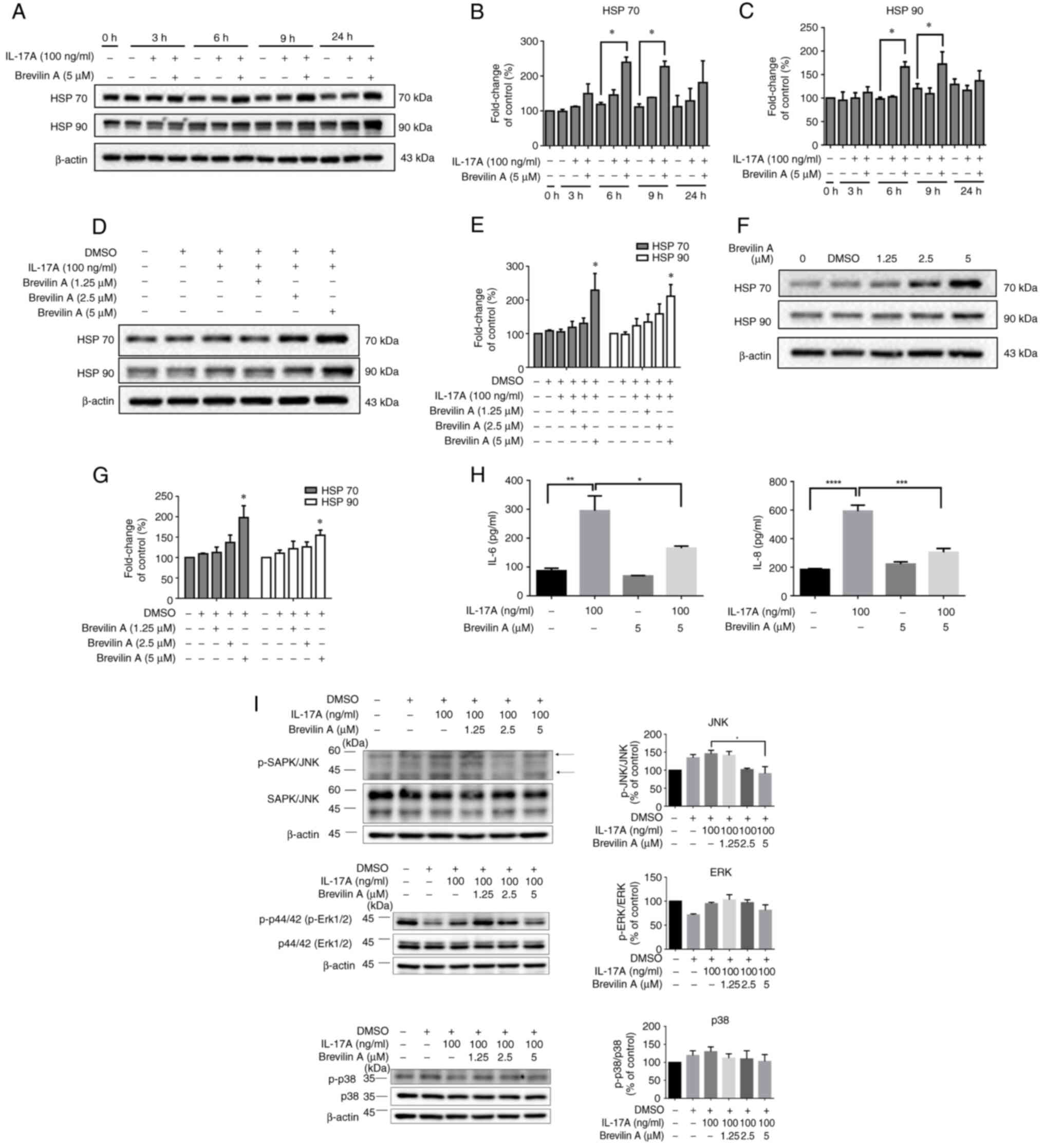 | Figure 2.Brevilin A increases HSP 70 and HSP
90 expression in HaCaT cells. (A) HaCaT cells were treated with
IL-17A alone (100 ng/ml), or were co-treated with IL-17A and
brevilin A (5 µM) for different durations (0, 3, 6, 9, and 24 h).
Expression levels of HSP 70, HSP 90 and β-actin (loading control)
were detected by western blotting. Semi-quantitative analysis of
(B) HSP 70 and (C) HSP 90 expression levels. The intensity of HSP
70/HSP 90 bands was normalized to β-actin and expressed as fold
change relative to control (untreated) cells. (D) Western blot
analysis of the expression levels of HSP 70 and HSP 90 in HaCaT
cells treated with IL-17A (100 ng/ml) and various concentrations of
brevilin A (1.25, 2.5 and 5 µM) for 9 h. β-actin was used as a
loading control. (E) Semi-quantitative analysis of HSP 70 and HSP
90 band intensity was performed using ImageJ. The intensity of
bands was normalized to β-actin and expressed as fold change
relative to control (untreated) cells. (F) Western blot analysis of
the expression levels of HSP 70 and HSP 90 in HaCaT cells treated
with various concentrations of brevilin A (1.25, 2.5 and 5 µM) for
24 h. β-actin was used as a loading control. (G) Semiquantitative
analysis of HSP 70 and HSP 90 band intensity was performed using
ImageJ. The intensity of bands was normalized to β-actin and
expressed as fold change relative to control (untreated) cells. (H)
HaCaT cells were treated with IL-17A (100 ng/ml) alone, brevilin A
(5 µM) alone, or a combination of IL-17A and brevilin A for 9 h.
The secretion of IL-6 and IL-8 was measured by ELISA. (I) Brevilin
A modulates the MAPK signaling pathway in IL-17A-treated cells.
Western blot analysis was performed to assess the phosphorylation
levels of SAPK/JNK, ERK and p38 MAPK after treatment with IL-17A
(100 ng/ml) and different concentrations of brevilin A (1.25, 2.5
and 5 µM) for 9 h. Total SAPK/JNK, ERK and p38 protein levels were
used as controls, and β-actin served as the loading control.
Semi-quantification of p-SAPK/JNK, p-ERK and p-p38 was normalized
to total protein levels and presented as a percentage of the
control group. Data are presented as the mean ± SEM (n=3).
*P<0.05, **P<0.01, ***P<0.001, ****P<0.0001 vs. control
or as indicated. HSP, heat shock protein; p-, phosphorylated. |
To further confirm that brevilin A has the ability
to counteract IL-17A-induced inflammatory cytokine production, the
effects of IL-17A, brevilin A and their combination were examined
on IL-6 and IL-8 levels. The results demonstrated that IL-17A
significantly induced the production of IL-6 and IL-8, while
brevilin A alone did not increase IL-6 or IL-8 levels. In the
IL-17A and brevilin A co-treatment group, the levels of IL-6 and
IL-8 were significantly reduced compared with in the IL-17A
treatment group (Fig. 2H). These
findings indicated that brevilin A may exert a protective effect by
preventing keratinocytes from producing inflammatory cytokines in
response to IL-17A stimulation.
To further explore the molecular mechanisms
underlying the effects of brevilin A on IL-17A-induced responses,
the activation of the MAPK signaling pathway was examined; this
pathway serves a critical role in inflammation and stress
responses. Co-treatment with IL-17A and brevilin A (1.25, 2.5 and 5
µM) for 9 h reduced the phosphorylation of JNK in a dose-dependent
manner compared with in the IL-17A treatment group, whereas the
phosphorylation of ERK and p38 remained largely unaffected
(Fig. 2I). Brevilin A did not
significantly affect p-ERK or p-p38 levels, suggesting that its
regulatory effect is selective for the JNK pathway. These results
indicated that brevilin A may selectively inhibit IL-17A-induced
JNK phosphorylation without affecting ERK or p38, highlighting its
potential role in modulating inflammatory signaling through the JNK
pathway. These findings suggested that brevilin A specifically
suppresses the IL-17A-induced activation of the JNK pathway, which
may contribute to its effects on HSP 70 and HSP 90 expression and
its anti-inflammatory properties. All of the experiments were
performed in triplicate to ensure the reliability and
reproducibility of the results. These findings highlight the
capability of brevilin A to induce HSP 70 and HSP 90 expression
independently and in conjunction with IL-17A in keratinocytes,
suggesting a potential protective or regulatory role in
inflammatory responses within the skin.
Brevilin A increases HSP 70 expression
in keratinocytes and alleviates inflammation in a mouse model of
IMQ-induced psoriasis
To further investigate the effects of brevilin A on
inducing HSP 70 expression, an animal model of IMQ-induced
psoriasis was generated. Briefly, 8-week-old BALB/c mice were
randomly divided into six groups (n=5/group): Control (untreated),
DMSO (vehicle control), brevilin A (5, 10 and 20 mg/kg) and
dexamethasone (1 mg/kg) groups. BALB/c mice were first treated with
different doses (5, 10 or 20 mg/kg) of brevilin A for 5 days.
Epithelial cell hyperproliferation, and HSP 70, HSP 90 and IL-6
expression were then detected using H&E staining and IHC.
Representative images of the skin on the backs of the mice showed
that brevilin A treatment reduced erythema and scaling compared
with that in the vehicle control group (Fig. 3A). H&E staining revealed
decreased epithelial cell hyperproliferation and immune cell
infiltration in the brevilin A-treated groups and the
dexamethasone-treated group compared with that in the vehicle
control group, suggesting a reduction of skin inflammatory
responses (Fig. 3B). Furthermore,
IHC demonstrated that brevilin A treatment led to a dose-dependent
increase in HSP 70 expression in epithelial cells, with the highest
expression observed in the 20 mg/kg group (Fig. 3C and F). In addition, in the
dexamethasone treatment group, the expression of HSP 70 was
significantly increased compared with that in the vehicle control
group. By contrast, IHC staining of IL-6 showed a significant
reduction in its expression in the brevilin A-treated groups and
the dexamethasone-treated group, indicating its role in mitigating
inflammatory responses (Fig. 3D and
F). HSP 90 expression was moderately upregulated in brevilin
A-treated groups in a dose-dependent manner, with the highest
levels at 20 mg/kg (Fig. 3E and
F). Furthermore, dexamethasone treatment also increased HSP 90
expression conpared with that in the vehicle control group. These
results collectively indicated that brevilin A may exert protective
effects against psoriasis by alleviating inflammation and
upregulating HSP 70 expression in epithelial cells.
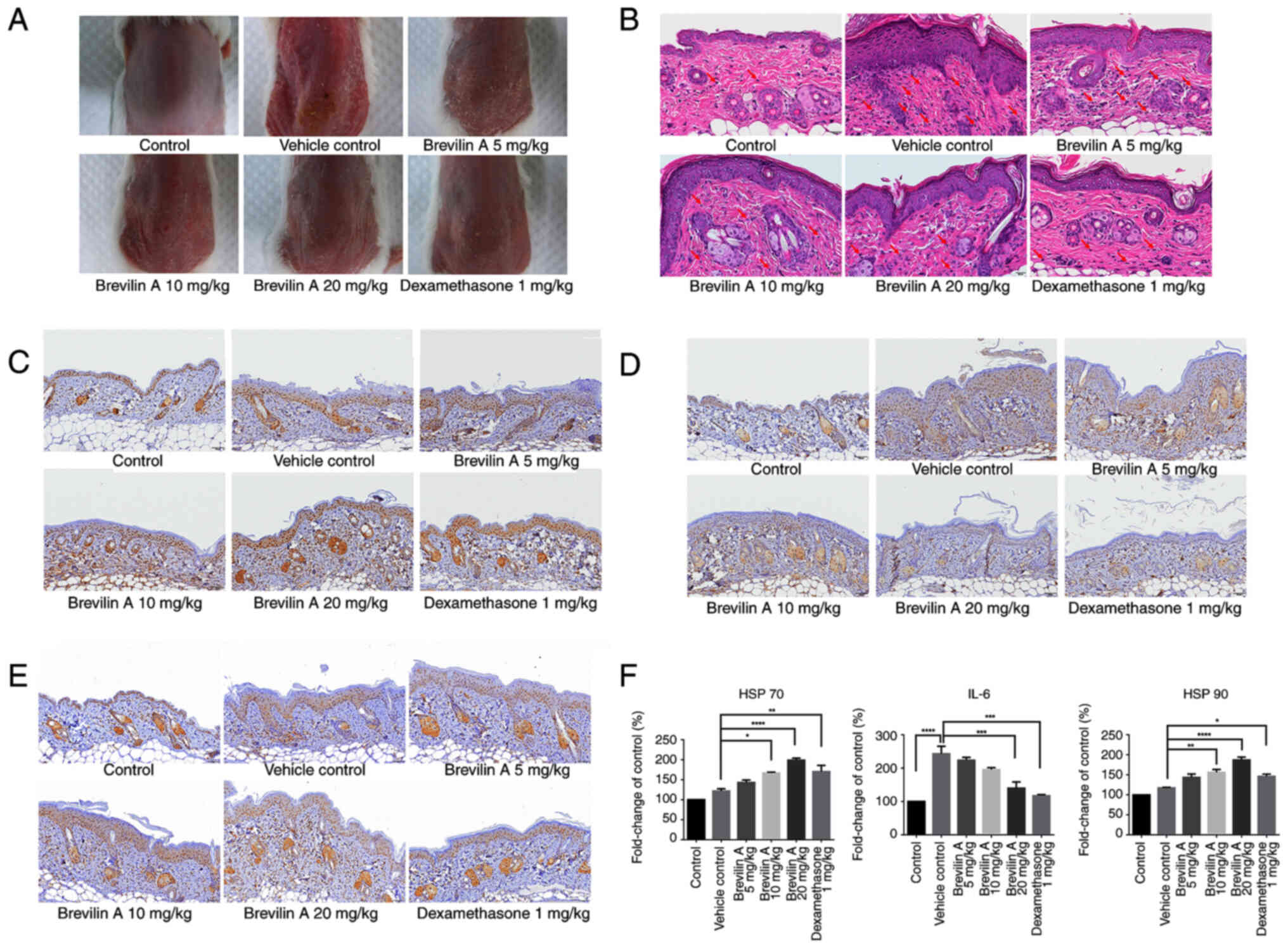 | Figure 3.Brevilin A reduces IMQ-induced skin
inflammation, decreases IL-6 expression and enhances HSP 70
expression in a mouse model of psoriasis. Briefly, 8-week-old
BALB/c mice (n=5/group) with IMQ-induced psoriasis were treated
with brevilin A (5, 10 and 20 mg/kg) or dexamethasone (1 mg/kg) by
intraperitoneal injection for 5 consecutive days. (A)
Representative images of the skin on the backs of mice showed
improvement in erythema and scaling in brevilin A- and
dexamethasone-treated groups compared with the vehicle control. (B)
Skin pathology was assessed using hematoxylin and eosin staining
(×400 magnification). Representative images show the degree of
epithelial hyperproliferation and immune cell infiltration, with
immune cells indicated by red arrows. Scale bar=20 µm. (C) IHC
staining of HSP 70 showed increased expression in brevilin
A-treated groups, with the highest expression observed at 20 mg/kg.
Scale bar=50 µm; ×200 magnification. (D) IHC staining of IL-6
revealed reduced expression in brevilin A-treated groups,
suggesting its anti-inflammatory effects. Scale bar=50 µm; ×200
magnification. (E) IHC staining of HSP 90 showed increased
expression in brevilin A-treated groups, with the highest
expression observed at 20 mg/kg. Scale bar=50 µm; ×200
magnification. (F) Semi-quantification of HSP 70, IL-6 and HSP 90
staining intensity are presented as the mean ± SEM. *P<0.05,
**P<0.01, ***P<0.001 and ****P<0.0001. IHC,
immunohistochemistry; IMQ, imiquimod |
An HSP 70 inhibitor reverses the
effects of brevilin A on IL-17A-induced proinflammatory cytokine
production
To elucidate the underlying mechanisms regarding the
effects of brevilin A on reducing IL-17A-induced IL-6 and IL-8
production through increasing HSP 70 expression, the present study
assessed the effects of an HSP 70-specific inhibitor, apoptozole.
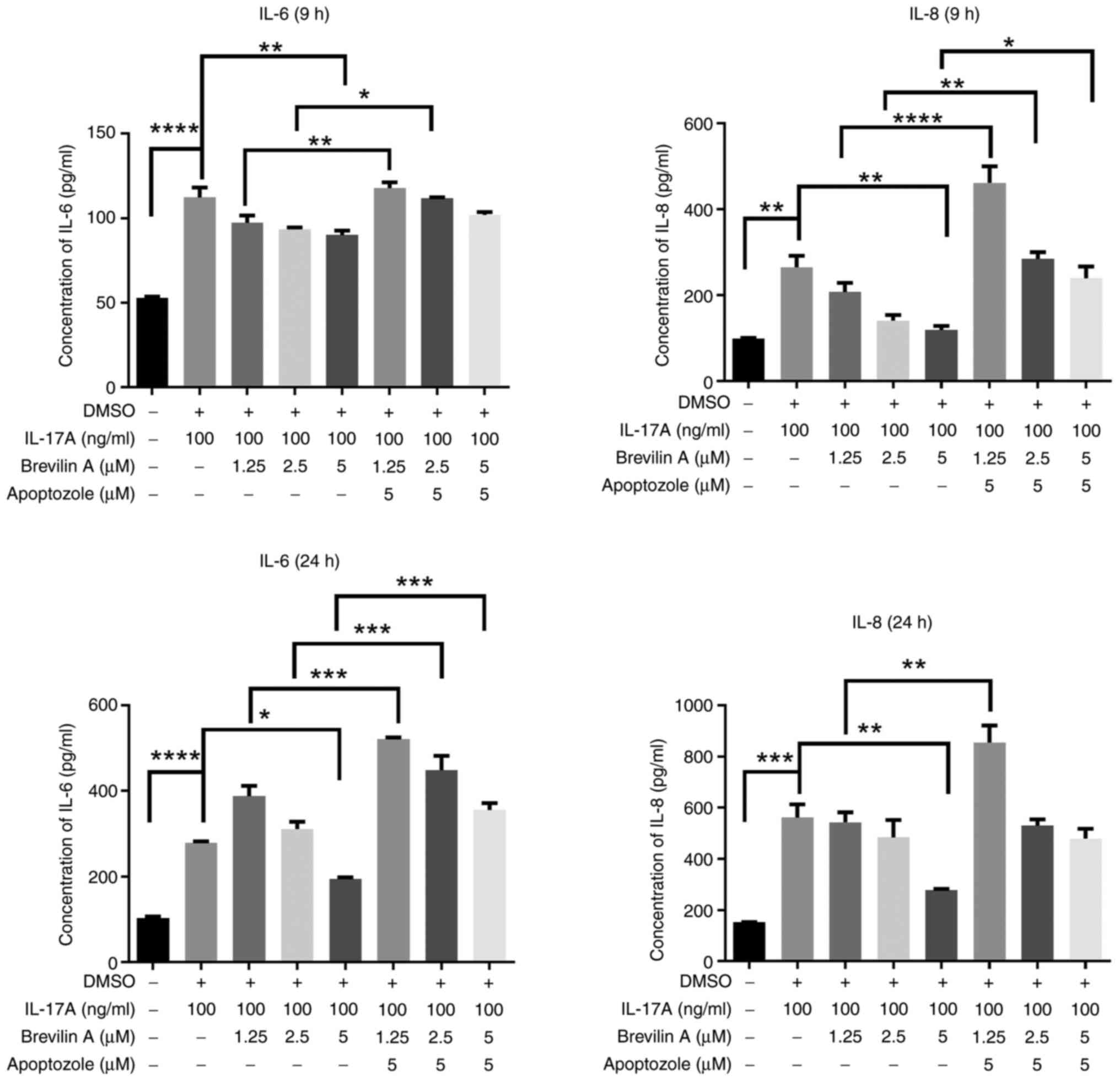
Initially, it was revealed that reatment with 20 µM apoptozole for
24 h significantly reduced the viability of HaCaT cells; therefore,
1.25, 2.5 and 5 µM apoptozole were used in subsequent experiments
(Fig. 4). Next, the current study
analyzed HaCaT cells co-treated with different doses of apoptozole,
100 ng/ml IL-17A and 5 µM brevilin A for 9 and 24 h; the results
showed that co-treating cells with apoptozole and IL-17A did not
increase the levels of IL-6 or IL-8 compared with those in the
IL-17A treatment group; however, after co-treating cells with
apoptozole (2.5 and 5 µM), IL-17A and brevilin A, the levels of
IL-6 and IL-8 were significantly higher in a dose-dependent manner
compared with those in the IL-17A + brevilin A group (Fig. 5). To determine whether the HSP
70-specific inhibitor could reverse the anti-inflammatory effects
of brevilin A on cytokine expression, HaCaT cells were co-treated
with 5 µM apoptozole and different doses of brevilin A for 9 and 24
h. The results showed that 5 µM brevilin A significantly reduced
IL-17A-induced IL-6 and IL-8 production compared with that in the
IL-17A-group, whereas the 5 µM brevilin A + 5 µM apoptozole group
showed a reversal of this reduction, restoring IL-6 levels at 24 h
and IL-8 levels at 9 h (Fig. 6).
These data indicated that HSP 70 may serve a major role in the
anti-inflammatory effects of brevilin A treatment.
Discussion
HSP 70 is known to serve a role in suppressing
disease progression and exerting immunomodulatory effects (14); however, the role of HSP70 in
psoriasis treatment remains controversial, as it can either
suppress inflammation by inhibiting pro-inflammatory cytokines, or
exacerbate the disease by promoting immune activation and
keratinocyte survival, depending on its localization and function.
The present study demonstrated that brevilin A reduced
IL-17A-induced IL-6 and IL-8 levels in HaCaT cells, and the
expression levels of HSP 70 and HSP 90 were upregulated in the
brevilin A-treated group. In the in vivo experiment,
brevilin A reduced epithelial cell hyperproliferation and immune
cell infiltration, and increased HSP 70 expression. The mechanistic
experiment showed that an HSP 70-specific inhibitor, apoptozole,
reversed the effects of brevilin A on reducing IL-17A-induced IL-6
and IL-8 levels in HaCaT cells. Taken together, these results
suggested that brevlin A may inhibit IL-17A-induced proinflammatory
cytokine production, acting through HSP 70 uprgulation.
In the pathogenesis of psoriasis, proinflammatory
cytokines have an important role in disease progression and
comorbidities. For example, IL-6 is a major cytokine that induces
the differentiation of naïve T cells into Th17 cells, and its
expression is upregulated in the serum and skin lesions of patients
with psoriasis (15). Another
cytokine, IL-8, which increases the infiltration of
polymorphonuclear cells, and induces keratinocyte proliferation and
angiogenesis, is also involved in skin inflammation (16). The levels of both of these cytokine
were increased in keratinocytes after IL-17A stimulation in the
present study. By contrast, brevilin A reduced IL-17A-induced
production of IL-6 and IL-8. Previous studies have shown that
brevilin A, a sesquiterpene lactone isolated from Centipeda
minima, exhibits anti-inflammatory and anticancer properties
(10,11,17–19).
Brevilin A has also been reported to inhibit NF-κB activation,
which is a crucial pathway in the inflammatory response (11). Additionally, brevilin A has been
shown to suppress the production of other proinflammatory
cytokines, such as PGE2 and IL-1β, in the cartilage tissue of an
osteoarthritis mouse model (20).
These data suggested that brevilin A may be a promising drug for
the treatment of psoriasis and other IL-17A-induced inflammatory
diseases.
HSPs are cellular chaperones that respond to stress
and intracellular protein misfolding. Notably, the role of HSPs in
anti-inflammatory responses and immunomodulation has previously
been reported (14,21). Among the HSP superfamily, HSP 70 is
crucial due to its widespread influences in protein folding,
protection against stress and modulating immune responses. HSP 70
regulates classical intracellular signaling pathways related to the
expression of proinflammatory cytokines, such as MAPK and NF-κB. By
inhibiting the binding of NF-κB to inflammation-related genes, and
inhibiting IκB-α phosphorylation, HSP 70 is able to downregulate
the levels of proinflammatory cytokines (22). The results of the present study,
alongside supporting evidence from the literature (23), highlight the modulatory role of HSP
70 in the inflammatory response driven by IL-17-mediated pathways.
HSP 70 appears to regulate the levels of the key pro-inflammatory
cytokines IL-6 and IL-8 through its influence on critical signaling
pathways, such as NF-κB and MAPK. Notably, HSP 70 has been shown to
inhibit NF-κB activation under specific conditions, leading to a
suppression of IL-6 and IL-8 production (24); however, under cellular stress
conditions, HSP 70 may stabilize certain signaling intermediates,
which can enhance cytokine expression (25). These observations suggest that HSP
70 predominantly modulates the levels of IL-6 and IL-8, rather than
being directly influenced by these cytokines. This finding
underscores the importance of HSP 70 as a pivotal regulator within
the IL-17-driven inflammatory axis and provides valuable insights
into its potential as a therapeutic target for chronic inflammatory
diseases. Nevertheless, further research is warranted to elucidate
the precise mechanisms underlying these interactions in
disease-specific contexts, such as psoriasis and autoimmune
disorders.
HSP 70 has also been shown to have roles in
apoptosis regulation, where it inhibits apoptosis by binding to
apoptosis-regulating proteins, such as Apaf-1 and AIF (26). HSP 70 also serves a role in
cellular signaling by modulating the JNK and p38 MAPK pathways,
contributing to its cytoprotective functions under stress
conditions (27). Moreover, HSP 70
is involved in antigen presentation, enhancing the immune response
by facilitating the maturation of dendritic cells and improving the
presentation of antigenic peptides on MHC class I molecules
(28). Furthermore, HSP 70
activates STAT3 and ERK signaling pathways in myeloid-derived
suppressor cells (MDSCs) to increase IL-10 expression and suppress
immune responses (14).
The present study focused on HSP 70 due to its
crucial roles in modulating immune responses and regulating
proinflammatory cytokine production, which are key pathways in
psoriasis pathogenesis. While HSP 90 also serves important roles in
protein folding and stabilizing various regulatory proteins
(29), its anti-inflammatory
functions are less direct compared with HSP 70. The present study
showed that brevilin A can induce HSP 70 expression, which may be
associated with reduced inflammatory responses in psoriasis. This
finding provided a strong basis for focusing on HSP 70 to
understand the therapeutic effects of brevilin A. Future research
could explore the potential synergistic effects of targeting both
HSP 70 and HSP 90; however, the present study highlighted the
critical role of HSP 70 in the effects of brevilin A, offering
insights into its potential as a treatment for psoriasis and other
IL-17A-induced inflammatory diseases.
In the present study, dynamic, time-dependent
variations in the expression of HSP 70 and HSP 90 were observed
following treatment with IL-17A and brevilin A. Specifically,
western blotting results showed significant upregulation at the 6-
and 9-h time points, whereas the 24-h data did not achieve
statistical significance. This discrepancy may be attributed to
peak expression periods occurring at earlier time points,
reflecting a transient response. By 24 h, cellular regulatory
mechanisms may have modulated the expression levels, leading to
variability between samples. Notably, the current study did not
observe significant downregulation of HSP 70 or HSP 90 at the 24-h
mark compared with in the control and IL-17-treated groups. These
findings align with the known dynamic nature of HSP expression,
which can vary based on treatment duration, stress intensity, and
cellular context (30). The
significant induction of HSP 70 and HSP 90 at earlier time points
underscores their role in the cellular response to IL-17A and
brevilin A treatment, even as their expression levels fluctuate
over time.
The present study adopted the approach of
simultaneous model creation and drug administration to better
simulate a clinical treatment scenario. This approach reflects
real-world therapeutic interventions, where treatment typically
begins at the early onset of disease or cellular stress, rather
than as a preventive measure. The aim was to evaluate the
therapeutic effects of the drug in modulating disease progression
or the inflammatory response during the early stages, which aligns
more closely with clinical practice. While a pretreatment approach
might be valuable for studying preventive effects, it does not
accurately represent the timing of most clinical interventions. For
future investigations, comparing pretreatment and simultaneous
administration strategies could provide further insights into the
influence of treatment timing on drug efficacy.
IL-17A induces inflammatory cytokines as a result of
activating several intracellular signaling pathways, such as NF-κB,
MAPK or C/EBPs (31). In the
present study, it was observed that brevilin A reduced the levels
of IL-17A-induced proinflammatory cytokines. Furthermore, HSP 70 is
known to regulate NF-κB and MAPK signaling pathways (32); therefore, in the current
mechanistic study, a specific HSP 70 inhibitor, apoptozole, which
blocks the interaction between HSP 70 and related intracellular
proteins (33), was used to
identify the possible mechanism underlying the effects of brevilin
A on reducing IL-17A-induced inflammatory responses in HaCaT cells.
The results demonstrated that apoptozole reversed the effects of
brevilin A on IL-6 and IL-8 levels, indicating that HSP 70 may
serve a major role in brevilin A-regulated signaling pathways.
However, the expression levels of NF-κB- and MAPK-related protein
in the apoptozole- and brevilin A-co-treated group remain to be
clarified in future studies.
The present study demonstrated that brevilin A
reduced IL-17A-induced IL-6 and IL-8 levels in HaCaT cells through
upregulating the HSP 70 expression. However, there are some
limitations in this study. First, despite evidence that brevilin A
ameliorated psoriasis in an IMQ-induced murine model, in
vivo data on the reversal of brevilin A treatment effects by
HSP 70 inhibitors were lacking. Second, HSP 70 could have
interacted with numerous intracellular proteins to inhibit
inflammatory responses, wheras the present study only demonstrated
that HSP 70 expression was upregulated after brevilin A treatment.
Other proteins that are also regulated by HSP 70 remain to be
investigated in the future. Third, the protein expression levels of
IL-8 could not be assessed in skin tissues from brevilin A-treated
mice. The present study focused on HSP 70 due to its
well-established roles in anti-inflammatory and cytoprotective
mechanisms, which are directly relevant to the therapeutic effects
of brevilin A on psoriasis. HSP 70 has been shown to modulate
inflammatory responses and enhance cellular protection under stress
conditions, making it a key target in the context of psoriasis
pathophysiology. While HSP 90 is another important member of the
HSP family, its primary functions are more closely associated with
protein stabilization and signal transduction, which were beyond
the scope of this investigation. The present findings on the
upregulation of HSP 70 further support its potential as a critical
mediator in the anti-inflammatory effects of brevilin A. Future
studies could explore the role of HSP 90 in psoriasis to provide a
more comprehensive understanding of HSPs in this disease model.
In conclusion, the present study demonstrated that
brevilin A reduced IL-17A-induced IL-6 and IL-8 levels through
upregulating HSP 70 expression. The results indicated that brevilin
A not only regulates immune cell activation, but can also inhibit
proinflammatory cytokine-induced inflammatory responses in
keratinocytes. These results suggested that brevilin A may be a
promising compound that could be applied to treat psoriasis or
other inflammatory skin disorders.
Acknowledgements
Not applicable.
Funding
This research was supported by the Taichung Veterans General
Hospital/Hungkuang University Joint Research Program (grant no.
TCVGH-HK1138004), the Kaohsiung Veterans General Hospital (grant
nos. KSVGH112-040; KSVGH113-043), the Ministry of Science and
Technology, Taiwan (grant no. MOST 110-2314-B-075B-003-MY2), and
the National Science and Technology Council, Taiwan (grant no. NSTC
112-2914-I-075A-002-A1).
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
CYY and SJY conceptualized the study. CLL, KYY, WNH,
and ECL designed the methodology. KCW, ECL, HSH, TYC and CLL
conducted the formal analysis. KCW, KYY, TYC, HSH, WNH and SJY
carried out the investigation. CLL and SJY prepared the original
draft of the manuscript, and CYY reviewed and edited it. SJY and
CYY were responsible for project administration and secured funding
for the study. CLL, SJY and TYC confirm the authenticity of all the
raw data. All authors read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
The animal experimental protocol was approved by the
Institutional Animal Care and Use Committee of Kaohsiung Veterans
General Hospital (2021-A039).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ogawa E, Sato Y, Minagawa A and Okuyama R:
Pathogenesis of psoriasis and development of treatment. J Dermatol.
45:264–272. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang TS, Hsieh CF and Tsai TF:
Epidemiology of psoriatic disease and current treatment patterns
from 2003 to 2013: A nationwide, population-based observational
study in Taiwan. J Dermatol Sci. 84:340–345. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Coimbra S, Figueiredo A, Castro E,
Rocha-Pereira P and Santos-Silva A: The roles of cells and
cytokines in the pathogenesis of psoriasis. Int J Dermatol.
51:389–398. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Furue M, Furue K, Tsuji G and Nakahara T:
Interleukin-17A and keratinocytes in psoriasis. Int J Mol Sci.
21:12752020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bukau B, Weissman J and Horwich A:
Molecular chaperones and protein quality control. Cell.
125:443–451. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Han X, Cheng X, Xu J, Liu Y, Zhou J, Jiang
L, Gu X and Xia T: Activation of TREM2 attenuates neuroinflammation
via PI3K/Akt signaling pathway to improve postoperative cognitive
dysfunction in mice. Neuropharmacology. 219:1092312022. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Huang YH, Wang FS, Wang PW, Lin HY, Luo SD
and Yang YL: Heat shock protein 60 restricts release of
mitochondrial dsRNA to suppress hepatic inflammation and ameliorate
Non-alcoholic fatty liver disease in mice. Int J Mol Sci.
23:5772022. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Seifarth FG, Lax JE, Harvey J, DiCorleto
PE, Husni ME, Chandrasekharan UM and Tytell M: Topical heat shock
protein 70 prevents imiquimod-induced psoriasis-like inflammation
in mice. Cell Stress Chaperones. 23:1129–1135. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mishra S, Kumar A, Varadwaj PK and Misra
K: Structure-based drug designing and simulation studies for
finding novel inhibitors of heat shock protein (HSP70) as
suppressors for psoriasis. Interdiscip Sci. 10:271–281. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Qu Z, Lin Y, Mok DK, Bian Q, Tai WC and
Chen S: Brevilin A, a natural sesquiterpene lactone inhibited the
growth of Triple-negative breast cancer cells via Akt/mTOR and
STAT3 signaling pathways. Onco Targets Ther. 13:5363–5373. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu L, Chen X, Jiang Y, Yuan Y, Yang L, Hu
Q, Tang J, Meng X, Xie C and Shen X: Brevilin A ameliorates acute
lung injury and inflammation through inhibition of NF-κB signaling
via targeting IKKα/β. Front Pharmacol. 13:9111572022. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shih MC, Li CL, Liao EC, Yen CY, Yen LJ,
Wang KC, Lu LY, Chou TY, Chen YC and Yu SJ: Inhibition of NLRP3
inflammasome activation by 3H-1,2-Dithiole-3-Thione: A potential
therapeutic approach for psoriasis treatment. Int J Mol Sci.
24:135282023. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Spierings J and van Eden W: Heat shock
proteins and their immunomodulatory role in inflammatory arthritis.
Rheumatology. 56:198–208. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Borges TJ, Wieten L, van Herwijnen MJ,
Broere F, van der Zee R, Bonorino C and van Eden W: The
anti-inflammatory mechanisms of Hsp70. Front Immunol. 3:952012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Blauvelt A: IL-6 Differs from TNF-α:
Unpredicted clinical effects caused by IL-6 blockade in psoriasis.
J Invest Dermatol. 137:541–542. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Konstantinova NV, Duong DM, Remenyik E,
Hazarika P, Chuang A and Duvic M: Interleukin-8 is induced in skin
equivalents and is highest in those derived from psoriatic
fibroblasts. J Invest Dermatol. 107:615–621. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu YF, Li WQ, Hu ND, Ai B, Xia HX, Guo X,
Chen Z and Xia H: Brevilin A ameliorates sepsis-induced
cardiomyopathy through inhibiting NLRP3 inflammation. Ann Med Surg
(Lond). 85:5952–5962. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Qin Q, Xu G, Zhan X, Wang Z, Wang Y, Liu
H, Hou X, Shi W, Ma J, Bai Z and Xiao X: Brevilin A inhibits NLRP3
inflammasome activation in vivo and in vitro by acting on the
upstream of NLRP3-induced ASC oligomerization. Mol Immunol.
135:116–126. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang R, Gao C, Yu M, Song J, Feng Z, Wang
R, Pan H, Liu H, Li W and Fan X: Mechanistic prediction and
validation of Brevilin A Therapeutic effects in Lung Cancer. BMC
Complement Med Ther. 24:2142024. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ruan Q, Wang C, Zhang Y and Sun J:
Brevilin A attenuates cartilage destruction in osteoarthritis mouse
model by inhibiting inflammation and ferroptosis via
SIRT1/Nrf2/GPX4 signaling pathway. Int Immunopharmacol.
124:1109242023. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zininga T, Ramatsui L and Shonhai A: Heat
shock proteins as immunomodulants. Molecules. 23:28462018.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cheng HM, Wu YC, Wang Q, Song M, Wu J,
Chen D, Li K, Wadman E, Kao ST, Li TC, et al: Clinical efficacy and
IL-17 targeting mechanism of Indigo naturalis as a topical agent in
moderate psoriasis. BMC Complement Altern Med. 17:4392017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tukaj S and Sitko K: Heat shock protein 90
(Hsp90) and Hsp70 as potential therapeutic targets in autoimmune
skin diseases. Biomolecules. 12:11532022. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Luo X, Zuo X, Zhou Y, Zhang B, Shi Y, Liu
M, Wang K, McMillian DR and Xiao X: Extracellular heat shock
protein 70 inhibits tumour necrosis factor-alpha induced
proinflammatory mediator production in fibroblast-like
synoviocytes. Arthritis Res Ther. 10:R412008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hulina-Tomašković A, Somborac-Bačura A,
Grdić Rajković M, Bosnar M, Samaržija M and Rumora L: Effects of
extracellular Hsp70 and cigarette smoke on differentiated THP-1
cells and human monocyte-derived macrophages. Mol Immunol.
111:53–63. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ravagnan L, Gurbuxani S, Susin SA, Maisse
C, Daugas E, Zamzami N, Mak T, Jäättelä M, Penninger JM, Garrido C
and Kroemer G: Heat-shock protein 70 antagonizes apoptosis-inducing
factor. Nat Cell Biol. 3:839–843. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Porto RR, Dutra FD, Crestani AP, Holsinger
RMD, Quillfeldt JA, Homem de Bittencourt PI Jr and de Oliveira
Alvares L: HSP70 facilitates memory consolidation of fear
conditioning through MAPK pathway in the hippocampus. Neuroscience.
375:108–118. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kettern N, Rogon C, Limmer A, Schild H and
Höhfeld J: The Hsc/Hsp70 co-chaperone network controls antigen
aggregation and presentation during maturation of professional
antigen presenting cells. PLoS One. 6:e163982011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wei H, Zhang Y, Jia Y, Chen X, Niu T,
Chatterjee A, He P and Hou G: Heat shock protein 90: Biological
functions, diseases, and therapeutic targets. MedComm (2020).
5:e4702024. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Diller KR: Stress protein expression
kinetics. Annu Rev Biomed Eng. 8:403–424. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Swaidani S, Liu C, Zhao J, Bulek K and Li
X: TRAF regulation of IL-17 cytokine signaling. Front Immunol.
10:12932019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen H, Wu Y, Zhang Y, Jin L, Luo L, Xue
B, Lu C, Zhang X and Yin Z: Hsp70 inhibits
lipopolysaccharide-induced NF-kappaB a activation by interacting
with TRAF6 and inhibiting its ubiquitination. FEBS Lett.
580:3145–3152. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Du S, Liu Y, Yuan Y, Wang Y, Chen Y, Wang
S and Chi Y: Advances in the study of HSP70 inhibitors to enhance
the sensitivity of tumor cells to radiotherapy. Front Cell Dev
Biol. 10:9428282022. View Article : Google Scholar : PubMed/NCBI
|