Introduction
Telomerase activation is a critical step in human
carcinogenesis to maintain the telomere length; however, the
mechanisms for its activation and regulation remain to be
determined. The expression of hTERT, the catalytic subunit of the
telomerase enzyme, reveals a marked correlation to telomerase
activity in tumors. In the majority of tumors, active telomerase
requires the expression of hTERT. Furthermore, hTERT, which is
tightly regulated, is not detectable in most somatic cells where
telomerase activity is not present or is at basal level (1,2). hTERT
expression may be regulated by various epigenetic factors,
including promoter methylation or chromatin remodeling (3,4) and by
activators and repressors such as C-Myc, Sp1, USF1/2, p53, TGF-β
and CTCF (5–7).
One of the most crucial mechanisms by which hTERT
expression is regulated occurs throughout DNA methylation in the
hTERT promoter, which is embedded in a large CpG island.
Hypermethylation of hTERT was found to be necessary for its
expression in cancer cells (3,8). The
apparent opposition to the classic model of regulation was recently
clarified by the observation that methylation plays a dual role
(9,10). hTERT methylation prevents the
binding and thus the repressive effect of CTCF. However, a small
hypomethylated region around the transcription start site allows a
weak transcription of hTERT, despite the hypermethylation of the
border regions. In most normal tissues, the telomerase activity is
maintained at a basal level and it has been shown that these normal
somatic cells have a hypomethylated promoter (3,11,12).
Certain proliferative somatic cells, such as colorectal crypts as
well as gastric and endometrial cells, revealed telomerase activity
and hTERT expression concomitantly with the presence of hTERT
methylation (3, JB, unpublished data). An increase of telomerase
activity was observed in samples of normal, transitional and tumor
mucosa from patients with sporadic colorectal cancer (13). Since in the colon hTERT methylation
was detected in normal and tumor tissues, we aimed to investigate
whether a relationship exists between the level of telomerase
activity and the degree of hTERT promoter methylation in colorectal
tissues. Similarly, a potential relationship between hTERT promoter
methylation and telomere length was also determined.
Materials and methods
Tissue samples
This study involved 11 patients who underwent
surgery for colorectal cancer at the Lleida Arnau de Vilanova
Hospital. Of the 11 patients, 7 were male and 4 were female and
their average age was 75.40 years (range 65–82). Surgically
resected samples of cancer tissue and transitional and normal
mucosa were immediately frozen in liquid nitrogen and stored at
−80°C. The transitional mucosa was defined as having a
macroscopically normal appearance and lying <1 cm away from the
tumor, while the normal mucosa was obtained 10 cm away from the
tumor. The research protocol was approved by the Hospital Clinic
Research Ethics Committee of Lleida. Microdissection of two normal
colon samples was performed as previously described in order to
independently analyze the colon and stromal cells (14).
Fluorescent-telomeric repeat
amplification protocol assay
Telomerase activity was measured using a
quantitative system of the fluorescent-telomeric repeat
amplification protocol assay (TRAP-F), using a TRAPeze®
telomerase detection kit (InterGen) (15). The analysis was performed as
described in a previous study (13).
Telomere length measurement by Southern
blotting
The telomere restriction fragment (TRF) length was
determined by Southern blotting (Telo TAGGG Telomere length assay,
Roche Diagnostics GmbH, Mannheim, Germany) in the tumor,
transitional and normal tissues. The analysis was performed as
previously described (16). The
telomere length was expressed in kbp. The telomere length ratio
(TLR) was determined as the quotient between the TL in the tumor
tissue and the TL in normal mucosa from the same patient. Telomere
shortening and elongation were defined as TLs of carcinoma <80%
and >120% of the corresponding normal mucosa, respectively.
Methylation-sensitive single-strand
conformation analysis (MS-SSCA)
Bisulfite modification of 1 μg of genomic DNA (or 10
ng of DNA extracted from microdissected normal tissues) was
performed using the EpiTect® Bisulfite kit (Qiagen)
according to the manufacturer’s instructions. One twenty-fifth of
modified DNA was used to amplify a 220 bp fragment of the hTERT
promoter (17). PCR products were
analyzed by methylation-sensitive single-strand conformation
analysis (MS-SSCA) as previously described (18).
Results
The methylation status of the hTERT promoter was
analyzed, and the telomerase activity and telomere length were
determined in normal, transitional and tumor samples from 11
patients who underwent surgery for a sporadic colorectal cancer.
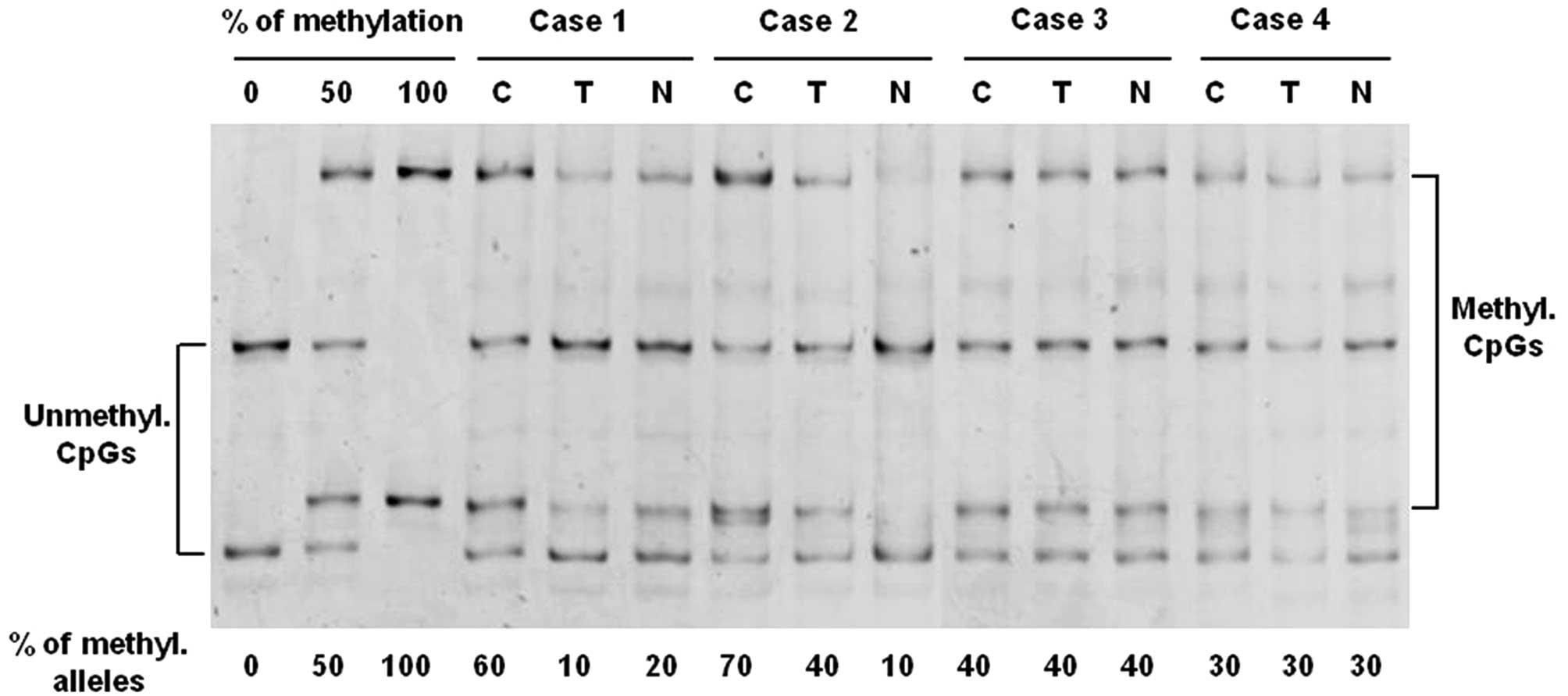
The analysis of hTERT promoter methylation was performed by MS-SSCA
after bisulfite modification of genomic DNAs. Four cases are shown
in Fig. 1. In all samples, a
mixture of unmethylated and fully methylated alleles at varying
ratios was observed. In cases 1 and 2, a significant difference in
the level of hTERT methylation between tumor and normal tissues was
detected. For example, in case 2, the level of methylation was
approximately 70% in the tumor sample, whereas the methylation
level reached only 40% in the transitional mucosa and only 10% in
the normal mucosa. A similar level of methylation was observed in
normal and tumor samples in cases 3 and 4. As shown in Table I, the mean level of hTERT
methylation was higher in tumor samples (51%) compared with
transitional (35%) and normal (20%) samples. After separation by
microdissection of epithelial cells and stromal cells from two
normal colon tissues, no methylation was detected in the hTERT
promoter of stromal cells (data not shown). Therefore, the hTERT
methylation observed in the normal colorectal tissues evidently
originated from the colonic cells.
 | Table IMean age, methylation rate, telomerase
activity and telomere length in tumor, transitional and normal
mucosa. |
Table I
Mean age, methylation rate, telomerase
activity and telomere length in tumor, transitional and normal
mucosa.
| Characteristics | Mean | Range |
|---|
| Age | 75.4 years | 65–82 years |
| Methylation rate |
| Tumor mucosa | 51% | 10–100% |
| Transitional
mucosa | 35% | 10–50% |
| Normal mucosa | 20% | 5–40% |
| Telomerase
activity |
| Tumor mucosa | 44.20 | 0–164 |
| Transitional
mucosa | 1.12 | 0–8 |
| Normal mucosa | 0.53 | 0–3 |
| Telomere length |
| Tumor mucosa | 7.66 kbp | 5.66–10.64 kbp |
| Transitional
mucosa | 8.82 kbp | 6.80–10.95 kbp |
| Normal mucosa | 9 kbp | 6.99–11.02 kbp |
Since all of the samples in this study contained at
least 40–50% of colorectal cells, it is evident that in a certain
number of samples only a subset of colorectal cells (normal or
tumoral) harbored a methylated hTERT promoter. Therefore, in order
to avoid insignificant methylation levels, a low level of hTERT
methylation was arbitrarily considered when the percentage of
methylation was <25%. As shown in Table II, the samples were divided into
two categories: low level (<25%) and high level (≥25%) of
methylation. In tumors, all but one of the samples exhibited a high
level of methylation. The percentage of cases with a high level of
methylation decreased to 73% in transitional samples, and 36% in
normal samples.
| Table IILow and high level of hTERT
methylation in tumor and normal colorectal tissues. |
Table II
Low and high level of hTERT
methylation in tumor and normal colorectal tissues.
| Met <25 (%) | Met ≥25 (%) | Total (%) |
|---|
| Tumor mucosa |
| TA− | 0 (0) | 2 (18) | 2 (18) |
| TA+ | 1 (9) | 8 (73) | 9 (82) |
| Total | 1 (9) | 10 (91) | 11 (100) |
| Transitional
mucosa |
| TA− | 2 (18) | 3 (27) | 5 (45) |
| TA+ | 1 (9) | 5 (46) | 6 (55) |
| Total | 3 (27) | 8 (73) | 11 (100) |
| Normal mucosa |
| TA− | 4 (36) | 3 (27) | 7 (64) |
| TA+ | 3 (27) | 1 (9) | 4 (36) |
| Total | 7 (64) | 4 (36) | 11(100) |
The mean telomerase activity was 40–80 times higher
in tumor samples than in transitional and normal mucosa,
respectively, whereas the mean level of hTERT methylation was only
1.5–2.5 times higher in tumor samples (Table I). In normal and transitional
tissues, a low level of telomerase activity was observed even in
the presence of a high level of methylation (Table III).
| Table IIIMeans of telomerase activity and
telomere length in tumor and normal colorectal tissues according to
the degree of hTERT methylation. |
Table III
Means of telomerase activity and
telomere length in tumor and normal colorectal tissues according to
the degree of hTERT methylation.
| Met <25% | Met ≥25% |
|---|
| Tumor mucosa |
| Telomerase
activity | 15 | 47.1 |
| Telomere length | 5.66 kpb | 7.87 kpb |
| Transitional
mucosa |
| Telomerase
activity | 0.13 | 1.75 |
| Telomere length | 7.31 kpb | 8.46 kpb |
| Normal mucosa |
| Telomerase
activity | 0.40 | 0.75 |
| Telomere length | 8.55 kpb | 9.69 kpb |
A reduction in the mean telomere length was observed
between normal (9 kbp), transitional (8.82 kbp), and tumor mucosa
(7.66 kbp) (Table I). These results
confirmed that tumor cells have shorter telomeres compared to
normal cells (19). Notably, in the
tumor, transitional and normal samples, the mean telomere length
was always shorter in the samples with a low level of methylation
than in those with a high level of methylation (Table III).
An increase of hTERT methylation and telomerase
activity, and a reduction of mean telomere length were observed in
the transitional mucosa in comparison to the distant normal mucosa.
However, the significant increase of methylation in the
transitional tissues was not concomitant with high telomerase
activity, as observed in the tumor samples (Tables II and III). These data indicate that hTERT
promoter methylation is not linearly correlated to telomerase
activity in colorectal tissues.
Discussion
A significant increase of hTERT methylation and
telomerase activity, and a reduction of mean telomere length were
observed from normal to tumor tissue. This observation also proves
to be true of transitional to tumor mucosa, but to a lesser extent.
A correlation between the degree of methylation and telomerase
activity was found in colorectal tumor samples. In other words,
tumors with a high degree of hTERT methylation revealed high
telomerase activity. An apparently different behavior occurred in
transitional and normal mucosa: the telomerase activity was low in
the two cases regardless of the methylation status of the hTERT
promoter. Nevertheless, when the level of hTERT methylation was
high, a higher level of telomerase activity was observed in
transitional and normal samples. Therefore, in tumor and normal
tissues, the higher the number of colorectal cells harboring an
hTERT methylated promoter, the more significant the level of
telomerase activity.
This study showed that hTERT promoter methylation is
required for hTERT expression, and thus telomerase activity, but it
is not sufficient to allow hTERT expression in normal colorectal
cells. The data observed in this study are in accordance with those
previously obtained in in vitro transcription experiments
with hTERT reporter gene constructs. The transcriptional activity
of the hTERT core promoter was found to be significantly higher in
cancer cells than in normal cells (9). Considering the significance of this
enzyme, it is clear that other factors should be recruited to
modulate hTERT expression.
hTERT methylation detected in normal mucosa may come
from the few stem cells and more largely from the daughter stem
cells, known as niche cells, contained in the colorectal crypts. In
previous studies, the cell expression of hTERT determined by
immunohistochemistry indicated that hTERT is mainly expressed at
the base of the crypts (20). The
fact that hTERT in transitional mucosa was more methylated than in
normal mucosa (mean of 35 versus 20%) indicates that some
significant changes occur in normal colorectal cells in the
vicinity of a tumor. This phenomenon may be due either to high cell
renewal or to the occurrence of epigenetic modifications in the
colorectal cells close to a cancer. Aberrant methylation of CpG
island promoters is known to be an early event in colorectal
carcinogenesis (21). Therefore, a
favorable hTERT methylation context may initiate the first phase of
carcinogenesis.
hTERT methylation was related, to a certain extent,
to the telomerase activity in normal and tumor colorectal tissues.
Low but significant levels of hTERT methylation and weak telomerase
activity are already present in the normal colorectal cells. A
slight increase of hTERT methylation, concomitantly with a great
change in telomerase activity level, occurs in tumor cells. In
conclusion, this study indicates that hTERT promoter methylation is
required but not sufficient for hTERT expression and telomerase
activity in colorectal cells.
Acknowledgements
This study was supported in part by grant
2006BE-00187 from the Generalitat de Catalunya (AGAUR).
References
|
1
|
Meyerson M, Counter CM, Eaton EN, et al:
hEST2, the putative human telomerase catalytic subunit gene, is
up-regulated in tumor cells and during immortalization. Cell.
90:785–795. 1997. View Article : Google Scholar
|
|
2
|
Nakamura TM, Morin GB, Chapman KB, et al:
Telomerase catalytic subunit homologs from fission yeast and human.
Science. 277:955–959. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Guilleret I, Yan P, Grange F, Braunschweig
R, Bosman FT and Benhattar J: Hypermethylation of the human
telomerase catalytic subunit (hTERT) gene correlates with
telomerase activity. Int J Cancer. 101:335–341. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kyo S, Takakura M, Fujiwara T and Inoue M:
Understanding and exploiting hTERT promoter regulation for
diagnosis and treatment of human cancers. Cancer Sci. 99:1528–1538.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Goueli BS and Janknecht R: Regulation of
telomerase reverse transcriptase gene activity by upstream
stimulatory factor. Oncogene. 22:8042–8047. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Poole JC, Andrews LG and Tollefsbol TO:
Activity, function, and gene regulation of the catalytic subunit of
telomerase (hTERT). Gene. 269:1–12. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Renaud S, Loukinov D, Bosman FT,
Lobanenkov V and Benhattar J: CTCF binds the proximal exonic region
of hTERT and inhibits its transcription. Nucleic Acids Res.
33:6850–6860. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nomoto K, Maekawa M, Sugano K, et al:
Methylation status and expression of human telomerase reverse
transcriptase mRNA in relation to hypermethylation of the p16 gene
in colorectal cancers as analyzed by bisulfite PCR-SSCP. Jpn J Clin
Oncol. 32:3–8. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Renaud S, Loukinov D, Abdullaev Z, et al:
Dual role of DNA methylation inside and outside of CTCF-binding
regions in the transcriptional regulation of the telomerase hTERT
gene. Nucleic Acids Res. 35:1245–1256. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zinn RL, Pruitt K, Eguchi S, Baylin SB and
Herman JG: hTERT is expressed in cancer cell lines despite promoter
DNA methylation by preservation of unmethylated DNA and active
chromatin around the transcription start site. Cancer Res.
67:194–201. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dessain SK, Yu HY, Reddel RR,
Beijersbergen RL and Weinberg RA: Methylation of the human
telomerase gene CpG island. Cancer Res. 60:537–541. 2000.PubMed/NCBI
|
|
12
|
Devereux TR, Horikawa I, Anna CH, Annab
LA, Afshari CA and Barrett JC: DNA methylation analysis of the
promoter region of the human telomerase reverse transcriptase
(hTERT) gene. Cancer Res. 59:6087–6090. 1999.PubMed/NCBI
|
|
13
|
Bautista CV, Felis CP, Espinet JM, Garcia
JB and Salas JV: Telomerase activity is a prognostic factor for
recurrence and survival in rectal cancer. Dis Colon Rectum.
50:611–620. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Baisse B, Bian YS and Benhattar J:
Microdissection by exclusion and DNA extraction for multiple PCR
analyses from archival tissue sections. Biotechniques. 28:856–8.
860:8622000.PubMed/NCBI
|
|
15
|
Kim NW, Piatyszek MA, Prowse KR, et al:
Specific association of human telomerase activity with immortal
cells and cancer. Science. 266:2011–2015. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bautista CV, Felis CP, Rene Espinet JM,
Garcia JB and Salas JV: Telomere length is a prognostic factor for
overall survival in colorectal cancer. Colorectal Dis. September
27–2010.(E-pub ahead of print).
|
|
17
|
Clement G and Benhattar J: A methylation
sensitive dot blot assay (MS-DBA) for the quantitative analysis of
DNA methylation in clinical samples. J Clin Pathol. 58:155–158.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bian YS, Yan P, Osterheld MC, Fontolliet C
and Benhattar J: Promoter methylation analysis on microdissected
paraffin embedded tissues using bisulfite treatment and PCR-SSCP.
Biotechniques. 30:66–72. 2001.PubMed/NCBI
|
|
19
|
Garcia-Aranda C, De JC, Diaz-Lopez A, et
al: Correlations of telomere length, telomerase activity, and
telomeric-repeat binding factor 1 expression in colorectal
carcinoma. Cancer. 106:541–551. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yan P, Benhattar J, Seelentag W, Stehle JC
and Bosman FT: Immunohistochemical localization of hTERT protein in
human tissues. Histochem Cell Biol. 121:391–397. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim YH, Petko Z, Dzieciatkowski S, et al:
CpG island methylation of genes accumulates during the adenoma
progression step of the multistep pathogenesis of colorectal
cancer. Genes Chromosomes Cancer. 45:781–789. 2006. View Article : Google Scholar : PubMed/NCBI
|