Introduction
Epithelial ovarian cancer (EOC) is the most lethal
of all gynecologic cancers and most women are diagnosed at an
advanced stage (1). Overall
mortality rates have remained relatively constant over the past
several decades and the cause of EOC is largely unknown.
The Kruppel-like factor 4 (KLF4) gene encodes
an epithelial cell-enriched, zinc finger-containing transcription
factor that has been shown to play important roles in cell
proliferation and differentiation (2,3). In
total, 70% of breast carcinomas have elevated KLF4 mRNA
levels, which are associated with a more aggressive phenotype
(4,5). By contrast, KLF4 expression is
frequently lost in various human cancers, including
gastrointestinal (6), bladder
(7) and advanced prostate cancer,
and KLF4 has been found to exert tumor-suppressive effects
(8). The dual and opposing roles of
KLF4 in tumorigenesis suggest that KLF4 is one of the
molecular elements that define the tissue-specific epithelial
carcinogenesis pathway.
Conversely, the ultimate vulnerability of cells to
apoptosis is determined by the relative ratio of various
pro-apoptotic and anti-apoptotic members of the Bcl-2 family
(9). The expression of Bcl-2
and Bax has been reported to be a prognostic factor in
ovarian cancer (10,11). In addition, KLF4
overexpression in a leukemia cell line affected the transcriptional
regulation of Bcl-2 and Bax, with effects on
apoptosis and cell growth (12).
The expression of KLF4 and its role in the
development of ovarian cancer has not previously been studied.
Therefore, to investigate the possible role of KLF4 in
ovarian carcinogenesis, we examined its expression in human ovarian
cancer tissues and measured Bcl-2 and Bax mRNA levels
in the same specimens. We also transduced the KLF4 gene into
ovarian cancer cells to investigate the changes in Bcl-2 and
Bax gene expression and the consequences in terms of cell
proliferation.
Materials and methods
Patients
This study was approved by the institutional review
board of Hanyang University Hospital (HYUHIRB-2009-R-50) and
written informed consent was obtained from each patient. The
patients included were women with surgically determined primary
advanced stage (III–IV) EOC who received debulking surgery at our
institution. Normal control samples obtained at the time of
salpingo-oophorectomy for benign indications were used for
comparative purposes. All tumor samples were snap frozen at the
time of surgery and stored at −70°C until use.
Real-time RT-PCR analysis
RNA from EOC, normal tissues and ovarian cancer
cells was isolated using an RNeasy extraction kit (Qiagen Inc.,
Valencia, CA, USA). Following quantification of RNA and
verification of its integrity, 1 μg samples were reverse
transcribed with an Advantage RT for PCR kit (BD Biosciences,
Clontech, Palo Alto, CA, USA). Primers were designed with the
Primers Express program (PE Applied Biosystems, Carlsbad, CA, USA):
KLF4 forward, 5′-ATCAGATGCAGCCGCAAGTCCC-3′ and reverse,
5′-TCT TCATGTGTAAGGCGAGGTGGTCC-3′ (GenBank accession no.
NM_004235.4); Bcl-2 forward, 5′-ATGTGTGTG GAGAGCGTCAA-3′ and
reverse, 5′-ACAGTTCCACAA AGGCATCC-3′ (GenBank accession no.
NM_000633.2); and Bax forward, 5′-GGGGACGAACTGGACAGTAA-3′
and reverse, 5′-CAGTTGAAGTTGCCGTCAGA-3′ (GenBank accession no.
NM_004324.3). Amplification of glyceraldehyde-3-phosphate
dehydrogenase (GAPDH) (forward, 5′-CAGCCTCAAGATCATCAGCA-3′ and
reverse, 5′-TGT GGTCATGAGTCCTTCCA-3′; GenBank accession no.
NM_002046.3) was used to normalize each reaction (amplification
product sizes 369, 136, 122 and 106 bps for KLF4,
Bcl-2, Bax and GAPDH, respectively). Real-time PCR
reactions were carried out in total volumes of 25 μl using
SYBR-Green Supermix (Bio-Rad, Hercules, CA, USA) with an iCycler™
Thermal Cycler (Bio-Rad). PCR conditions were 10 min at 95°C, 35
cycles of 95°C for 15 sec, 60°C for 45 sec and 72°C for 1 min.
Samples were run in triplicate in 96-well optical plates (Bio-Rad)
and the mean values were compared with normal controls to obtain
relative transcript levels.
Cell line transfection
Human ovarian cancer cell lines (SKOV3 and SNU251)
were purchased from the Korean Cell Line Bank (Seoul, Korea) and
transiently transfected with pCMV3xFLAG-KLF4 made by
subcloning the PCR-amplified coding region of KLF4. The
cells were plated in 6-well plates and incubated with 2 μg
aliquots of plasmid or empty vector in the presence of
Lipofectamine 2000 reagent (Invitrogen, Carlsbad, CA, USA).
Following incubation for 24–48 h, the cells were harvested for the
analysis of RNA, protein and cell numbers.
Cell proliferation assay
For assays examining FLAG-KLF4 abundance and
turnover, cell pellets were lysed in Laemmli buffer containing
β-mercaptoethanol (Bio-Rad). Samples were resolved by 4–12% NuPAGE
gel electrophoresis (Invitrogen), transferred to Hybond-P membranes
(Amersham Pharmacia Biotech, Arlington Heights, IL, USA) and
immunoblotted with anti-FLAG M2 antibody (1:1000) (F3165, Sigma,
St. Louis, MO, USA). Promega horseradish peroxidase-conjugated
anti-mouse immunoglobin G (W402B) antibody was used as a secondary
antibody. To ensure that lysates were loaded equally, the blots
were stripped and incubated with an anti-β-actin antibody (1:1000;
Sigma).
For the cell proliferation assays, cells were
transferred to 96-well microplates 24 or 48 h after transfection
and seeded at a density of approximately 1x105 cells per
well before the assay. Cell viability was subsequently determined
using an MTT cell proliferation assay kit (Cayman Chemical Company,
Ann Arbor, MI, USA). Absorbance was measured at 570 nm with a
microplate reader. The experiment was repeated 3 times and the data
were expressed as fold changes relative to empty vector-transfected
cells cultured for 24 h.
Statistical analysis
All data were analyzed using the Student’s t-test,
with P<0.05 considered to indicate a statistically significant
result. Data are expressed as the means ± standard deviation (SD)
of triplicate measurements.
Results
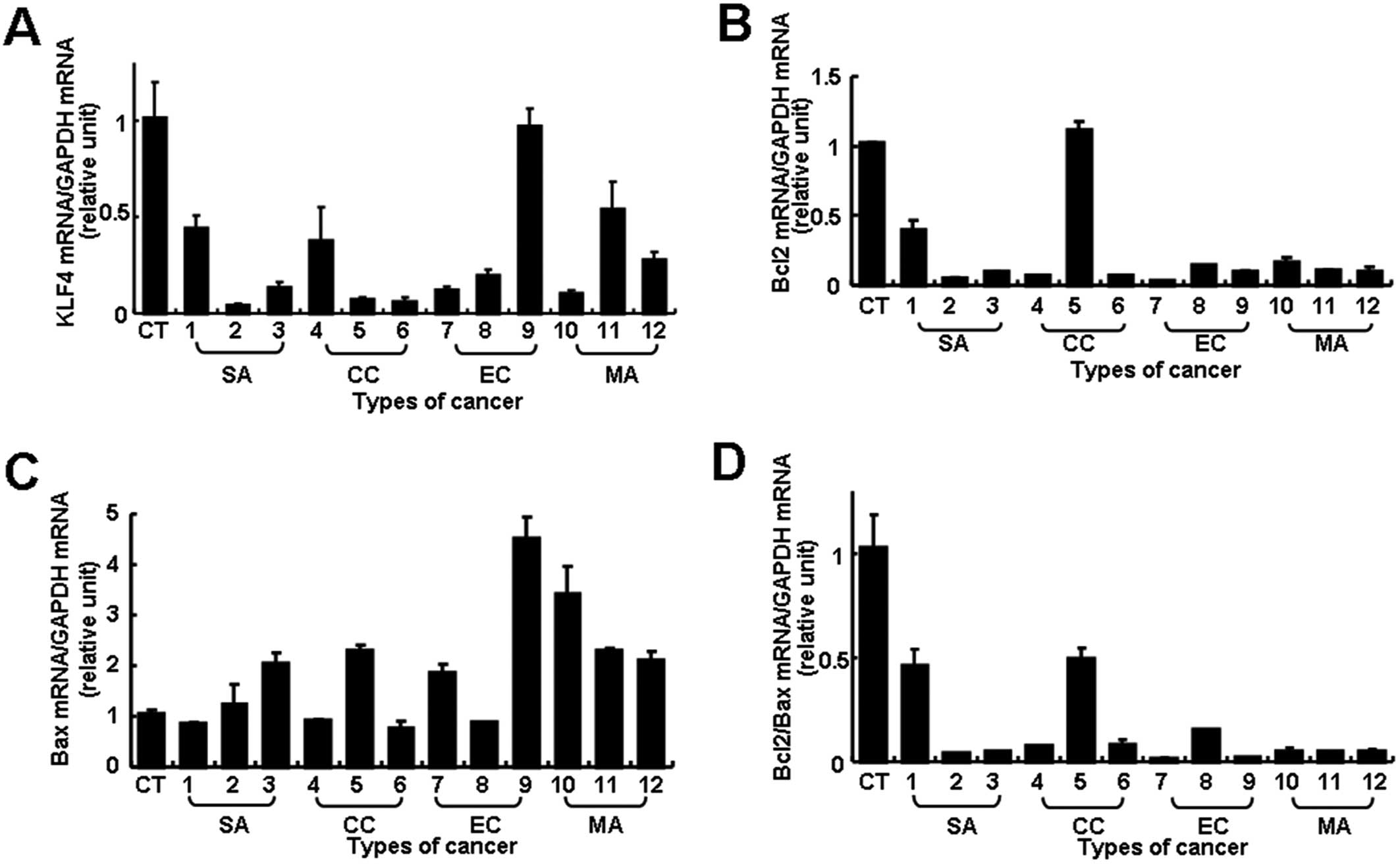
Using real-time RT-PCR, we analyzed mRNA expression
in pathology specimens from 3 normal ovaries and 12 cases of
advanced EOC, including 3 cases each of serous, clear cell and
endometrioid carcinoma and 3 mucinous cystadenocarcinomas.
KLF4 transcript levels were substantially
lower in most of the EOC samples (11 of 12 specimens) than in the
normal controls, although the extent of the difference varied
between tumors (Fig. 1A). The level
of expression in most samples was less than half the normal level
and it was particularly low (less than 0.3-fold of normal level) in
7 EOC samples. Bcl-2 or Bax mRNA expression also
varied between tumors and there was no correlation with
histological type (Fig. 1B and C).
However, an analysis of the Bcl-2/Bax ratio showed a
significantly reduced ratio, more than 2-fold lower than normal in
all 12 cases and even lower in 10 cases (more than 5-fold less than
normal) (Fig. 1D). Thus, both
reduced KLF4 transcript levels and lower Bcl-2/Bax
ratios were found in most of the EOC samples, but the extents of
the changes were not always correlated.
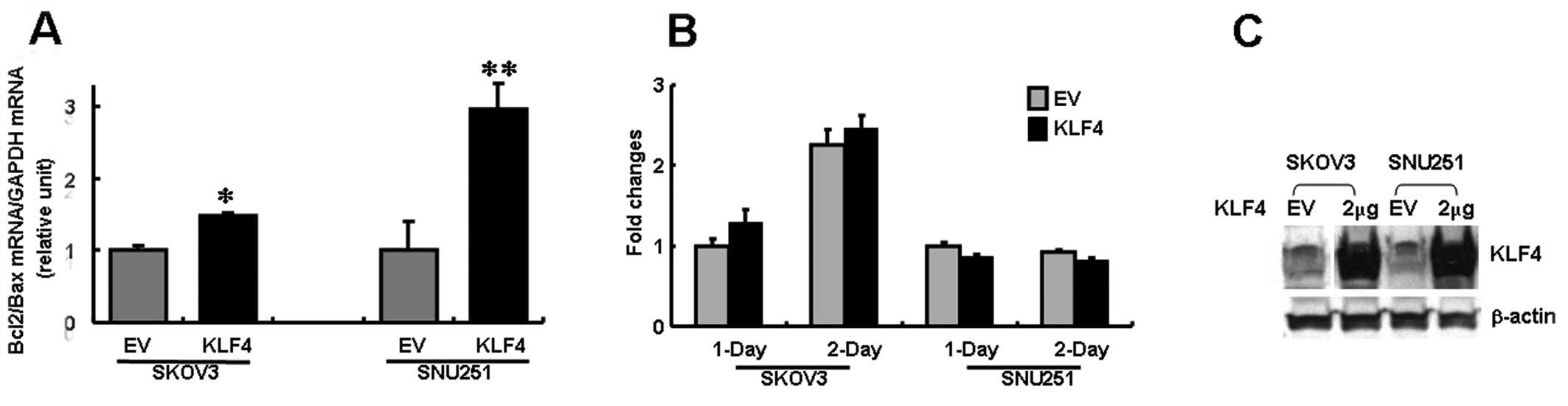
In order to examine the biological effects of
KLF4 expression in ovarian cancer cells, SKOV3 and SNU251
cells were transfected with FLAG-KLF4. An immunoblot
analysis confirmed marked expression of KLF4 in the
transfected cells (Fig. 2C).
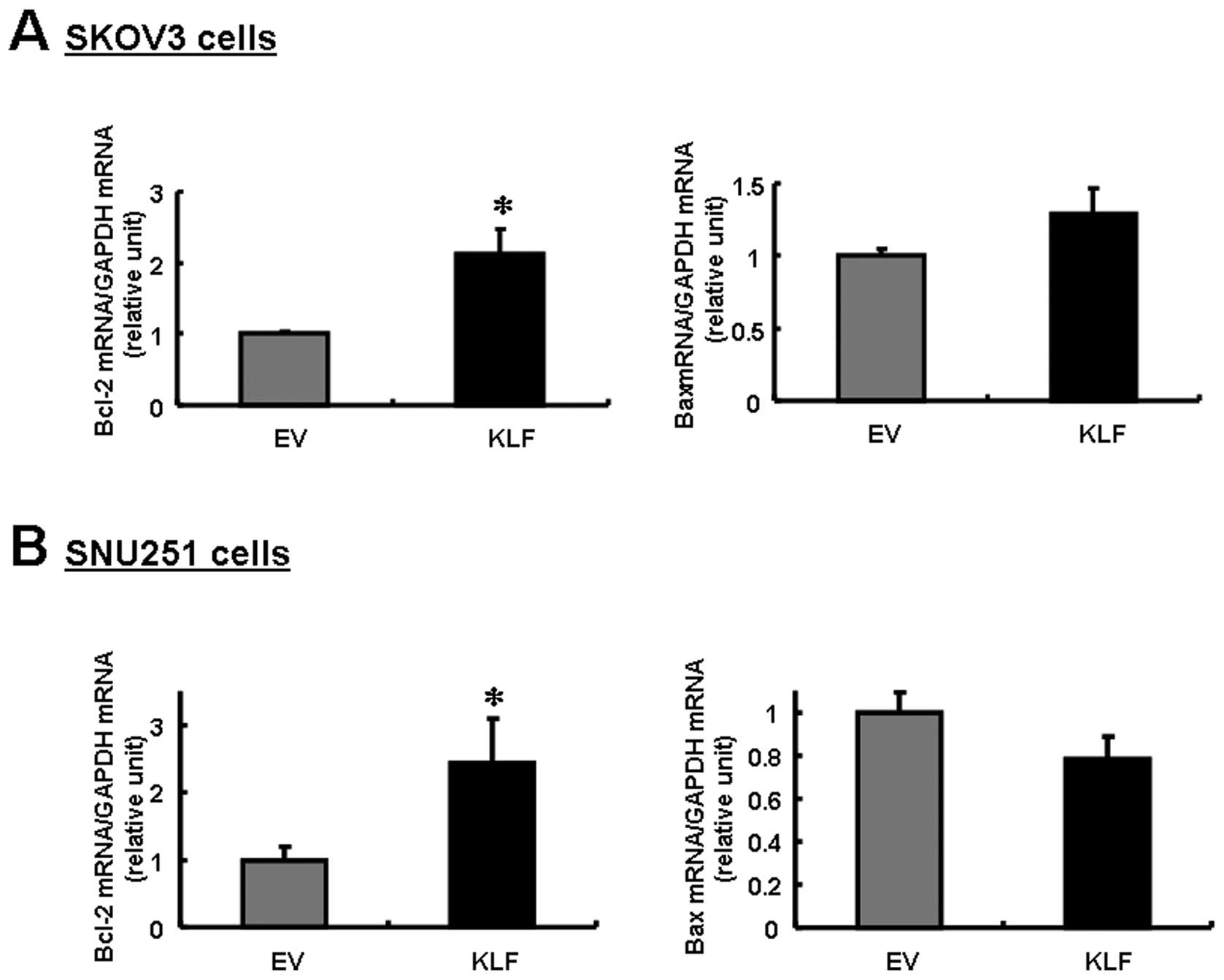
Notably, Bcl-2 expression was upregulated in both cell
lines, whereas Bax was downregulated in SNU251 cells and
slightly upregulated in SKOV3 cells (Fig. 3). The differential regulation of
Bcl-2 and Bax expression resulted in a decreased
Bcl-2/Bax ratio in both cell lines, but the decrease was
particularly significant in the SNU251 cells (P<0.03; Fig. 2A). This result suggests that the
decreased expression of KLF4 in EOC is correlated with the
reduced Bcl-2/Bax ratio, which is a known prognostic factor
in EOC.
To evaluate the effect of KLF4 on cell
proliferation, we used the MTT assay for quantitative analysis. In
contrast to the known inhibition of cell growth and induction of
apoptosis by KLF4 in a number of non-gonadal cancer cells,
the in vitro assays indicated that cell proliferation
following KLF4 gene transfection was unaffected at both 24
or 48 h of culture (P>0.05; Fig.
2B).
Discussion
This study is the first to suggest that KLF4
may play an important role in the development and progression of
ovarian cancer. We found that both the expression of KLF4
and the Bcl-2/Bax ratio were downregulated in many advanced
EOC cases, and that KLF4 overexpression in ovarian cancer
cells resulted in an increased Bcl-2/Bax ratio, which is
known to indicate a favorable prognosis in ovarian cancer.
Several lines of evidence indicate that KLF4
has variable effects on cell cycle arrest and inhibition of
apoptosis depending on the cellular context (7,8,12,13).
Although the expression of KLF4 was found to be
downregulated in ovarian cancers in this study, which is consistent
with the results for many human cancers, we observed no change in
cell proliferation due to KLF4 overexpression (Fig. 2B). Given that the multiplication of
human Sertoli cells is not affected by a lack of KLF4
(14), it is likely that the
different effects of KLF4 on cell proliferation depend on
unidentified cellular factors.
In this study we found that Bax and
Bcl-2 expression levels varied among tumors, as demonstrated
in other studies (10,11). Thus, it has been suggested that the
ratio of Bcl-2 to Bax, rather than the absolute
concentration of either, is predictive of cell fate (9), and their relative expression has been
reported to be a better predictor of outcome for both
progression-free survival and overall survival (10,11).
The transcription of Bcl-2 and Bax in leukemia cells
is affected by KLF4 overexpression (12). We also demonstrated in this study
that the transgenic expression of KLF4 in ovarian cancer
cells modulates Bcl-2/Bax gene expression (Fig. 2A). Previously, a microarray study
showed that decreased KLF4 expression was associated with
chemotherapy resistance in ovarian cancers (15). In principle, chemotherapeutics act
on rapidly proliferating cells and promote cell cycle arrest or
apoptosis. Thus, lower expression of KLF4 linked to a lower
Bcl-2/Bax ratio may possibly contribute to resistance or
relapse in advanced EOC. This increases the possibility that a
crucial function of KLF4 is to increase the Bcl-2/Bax
ratio, at least in ovarian cells, and that this is essential for a
favorable prognosis in ovarian cancer.
Although further research should be devoted to
understanding why KLF4 regulates cell growth and apoptosis
in many other cancer cells (6–8) but
not in ovarian cancer cells, our data indicate that the
downregulation of KLF4 may be a frequent step in ovarian
carcinogenesis. The decreased expression of KLF4 in ovarian
cancer may modulate Bcl-2/Bax expression, a known prognostic
factor for cancer grade, although its exact role in ovarian
carcinogenesis needs to be clarified.
Acknowledgements
We thank Professor Hyun Kook (Medical
Research Center for Gene Regulation, Chonnam National University
Medical School, Gwangju, Korea) for the gift of the
pCMV3-FLAG-KLF4 plasmid construct. This study was supported
by a Korea Research Foundation Grant funded by the Korean
Government (KRF 2009-0074679 and 2009-0065769).
References
|
1.
|
A JemalT MurrayE WardA SamuelsRC TiwariA
GhafoorEJ FeuerMJ ThunCancer statistics, 2005CA Cancer J
Clin551030200510.3322/canjclin.55.1.10
|
|
2.
|
LA Garrett-SinhaH EberspaecherMF SeldinB
de CrombruggheA gene for a novel zinc-finger protein expressed in
differentiated epithelial cells and transiently in certain
mesenchymal cellsJ Biol
Chem2713138431390199610.1074/jbc.271.49.31384
|
|
3.
|
JM ShieldsRJ ChristyVW YangIdentification
and characterization of a gene encoding a gut-enriched Kruppel-like
factor expressed during growth arrestJ Biol
Chem2712000920017199610.1074/jbc.271.33.200098702718
|
|
4.
|
KW FosterAR FrostP McKie-BellCY LinJA
EnglerWE GrizzleJM RuppertIncrease of GKLF messenger RNA and
protein expression during progression of breast cancerCancer
Res6064886495200011103818
|
|
5.
|
AY PandyaLI TalleyAR FrostTJ FitzgeraldV
TrivediM ChakravarthyDC ChhiengWE GrizzleJA EnglerH
KrontirasNuclear localization of KLF4 is associated with an
aggressive phenotype in early-stage breast cancerClin Cancer
Res1027092719200410.1158/1078-0432.CCR-03-048415102675
|
|
6.
|
AM GhalebJP KatzKH KaestnerJX DuVW
YangKruppel-like factor 4 exhibits antiapoptotic activity following
gamma-radiation-induced DNA
damageOncogene2623652373200710.1038/sj.onc.121002217016435
|
|
7.
|
S OhnishiS OhnamiF LaubK AokiK SuzukiY
KanaiK HagaM AsakaF RamirezT YoshidaDownregulation and growth
inhibitory effect of epithelial-type Kruppel-like transcription
factor KLF4, but not KLF5, in bladder cancerBiochem Biophys Res
Commun308251256200310.1016/S0006-291X(03)01356-112901861
|
|
8.
|
J WangRF PlaceV HuangX WangEJ NoonanCE
MagyarJ HuangLC LiPrognostic value and function of KLF4 in prostate
cancer: RNAa and vector-mediated overexpression identify KLF4 as an
inhibitor of tumor cell growth and migrationCancer
Res701018210191201010.1158/0008-5472.CAN-10-241421159640
|
|
9.
|
ZN OltvaiCL MillimanSJ KorsmeyerBcl-2
heterodimerizes in vivo with a conserved homolog, Bax, that
accelerates programmed cell
deathCell74609619199310.1016/0092-8674(93)90509-O8358790
|
|
10.
|
FJ de la TorreA GarciaA Gil-MorenoJ
PlanagumaJ ReventosS Ramon y CajalJ XercavinsApoptosis in
epithelial ovarian tumours Prognostic significance of clinical and
histopathologic factors and its association with the
immunohistochemical expression of apoptotic regulatory proteins
(p53, bcl-2 and bax)Eur J Obstet Gynecol Reprod
Biol1301211282007
|
|
11.
|
M SchuyerME van der BurgSC
Henzen-LogmansJH FieretJG KlijnMP LookJA FoekensG StoterEM
BernsReduced expression of BAX is associated with poor prognosis in
patients with epithelial ovarian cancer: a multifactorial analysis
of TP53, p21, BAX and BCL-2Br J
Cancer8513591367200110.1054/bjoc.2001.210111720475
|
|
12.
|
Z LiJ ZhaoQ LiW YangQ SongW LiJ LiuKLF4
promotes hydrogen-peroxide-induced apoptosis of chronic myeloid
leukemia cells involving the bcl-2/bax pathwayCell Stress
Chaperones15905912201010.1007/s12192-010-0199-520401760
|
|
13.
|
BD RowlandDS PeeperKLF4, p21 and
context-dependent opposing forces in cancerNat Rev
Cancer61123200610.1038/nrc178016372018
|
|
14.
|
M GodmannC KosanR BehrKruppel-like factor
4 is widely expressed in the mouse male and female reproductive
tract and responds as an immediate early gene to activation of the
protein kinase A in TM4 Sertoli
cellsReproduction139771782201010.1530/REP-09-053120051481
|
|
15.
|
AA JazaeriCS AwtreyGV ChandramouliYE
ChuangJ KhanC SotiriouO AprelikovaCJ YeeKK ZornMJ BirrerJC BarrettJ
BoydGene expression profiles associated with response to
chemotherapy in epithelial ovarian cancersClin Cancer
Res1163006310200510.1158/1078-0432.CCR-04-268216144934
|