Introduction
Previous studies have shown that expression of
angiogenic growth factors have a close correlation in the
progression of pancreatic cancer (1,2).
Pancreatic cancer cells produce multiple angiogenic growth factors.
They are, therefore, believed to be important sources of those
factors. Expression levels of angiogenic growth factors are likely
to be closely related to pancreatic cancer cell proliferation and
invasion. Different angiogenic growth factors may modulate each
other (3,4), particularly in cases where both
pro-angiogenic and anti-angiogeneic factors are involved. Vascular
endothelial growth factor (VEGF) and basic fibroblast growth factor
(bFGF) are by far the most important pro-angiogenic growth factors
(5,6), while endostatin has been found to be
the strongest anti-angiogenic factor (6,7). In
order to further study the modulatory effects between these
factors, sw1990, PCT-3 and Panc-1 pancreatic cancer cell lines were
infected with VEGF or bFGF siRNA, respectively. We found that VEGF
and bFGF siRNA significantly inhibited the secretion of endostatin
in PCT-3 and sw1990 cells. This study provided a basis to establish
anti-angiogenesis therapy in patients with pancreatic cancer. The
study was approved by the ethics committee of Hebei Medical
University (Shijiazhuang, China).
Materials and methods
Materials
Three human pancreatic cancer cell lines sw1990,
Panc-1 and PCT-3 were obtained from the laboratory of General
Surgery, Beijing Concord Hospital (China). VEGF siRNA, bFGF siRNA,
control siRNA (Fluorescein Conjugate) and siRNA transfection medium
were purchased from Santa Cruz Biotechnology, Inc (Santa Cruz, CA,
USA). Lipofectamine 2000 and the total RNA Extraction kit were
obtained from Invitrogen (Carlsbad, CA, USA). Endostatin ELISA kit
was obtained from R&D (Minneapolis, MN, USA). Polyclonal
endostatin antibody was obtained from Abcam (Cambridge, MA, USA).
Goat-anti-rabbit IgG-HRP was obtained from Santa Cruz
Biotechnology, Inc.
siRNA transfection in pancreatic cancer
cell lines
In preliminary experiments, sw1990, Panc-1 and PCT-3
cell lines were transfected with 2, 4, 6, 8 and 10 μl
control siRNA (Fluorescein Conjugate), respectively. Lipofectamine
2000 (2.5, 5 and 6 μl) were used to detect transfection
efficiency. Before transfection, each type of cancer cell
(2×105 per well) was seeded in six-well plates with 2 ml
antibiotic-free 1640 medium (10% FBS), at 37°C with 5%
CO2. When cells reached 60–80% confluence, cells were
transfected with a mixture of siRNA and Lipofectamine 2000 which
had been incubated at room temperature for 30 min. To determine
transfection efficiency, cells were visualized 24 h after
transfection by fluorescence microscopy.
RNA isolation and RT-PCR
Total RNA was isolated from the adult mouse heart.
In brief, cells were collected, lysed, and processed for total RNA
isolation at 4°C using an RNeasy Plus Mini Kit (Qiagen, Valencia,
CA, USA). The concentration of total RNA in each sample was
determined using a Nanodrop ND-1000 Spectrophotometer (Thermo
Scientific, Wilmington, DE, USA). The integrity of the extracted
RNA was confirmed by electrophoresis under denaturing conditions.
RT-PCR was performed using iScript cDNA Synthesis Kit (Bio-Rad,
Hercules, CA, USA) for the synthesis of a single-stranded cDNA
library. PCR reactions were performed using a Bio-Rad PCR machine.
The following primers were used: VEGF: 5′-AGCTACTGCCATCCAATCGC-3 ′,
5′-GGCGAATCCAATTCCAAGAG-3′; bFGF: 5′-AGCGGCTGTACTGCAAAAAC-3′,
5′-CCCAGGTCC TGTTTTGGAT-3′; Endostatin: 5′-CTCAATGCAGAGCAC
GATGT-3′, 5′-TGTTCTCAGGCTCTGAGGGT-3′; β-actin:
5′-GGCGGCACCACCATGTACCCT-3′, 5′-AGGGGCC GGACTCGTCATACT-3′. PCR
products were visualized on 1.5% agarose gel and pictures were
taken under UV lamp camera. The relative ratio was calculated using
the formula: (A) = Atarget gene /
Aβ-actin.
Enzyme-linked immunosorbent assay (ELISA)
determination of endostatin
Culture medium was collected after treatment with
different siRNAs or under control conditions. Endostatin
concentrations were determined according to ELISA kit manual
(R&D Systems). Each experiment was performed in triplicate.
Immunoblotting
Cell lysates were prepared by directly extracting
cells in lysis buffer containing 150 mM NaCl, 50 mM Tris-HCl, 1%
Triton, 0.5% NP40 and protease inhibitor cocktail (Roche, Mannheim,
Germany). Following centrifugation at 10,000 rpm for 10 min,
supernatants were collected. Protein concentration was quantified
using a BCA Protein Assay Kit (Thermo Fisher Scientific, Rockford,
IL, USA). Approximately 20 μg protein was dissolved with 4X
LDS sample buffer (Invitrogen) and separated on NuPAGE 4–12%
Bis-Tris gels (Invitrogen). The blots were visualized by enhanced
chemiluminescence and images were captured using a Kodak Image
Station 4000 R and quantified using Kodak MI SE software.
Statistical analyses
Each experiment was performed in triplicate and the
results are presented as mean ± standard error. The statistical
significance of differences between groups was assessed using
either a one-way ANOVA or two-tailed Student’s t-test with SPSS
(Chicago, IL, USA) 13.0 software. P<0.05 was considered to
indicate a statistically significant result.
Results
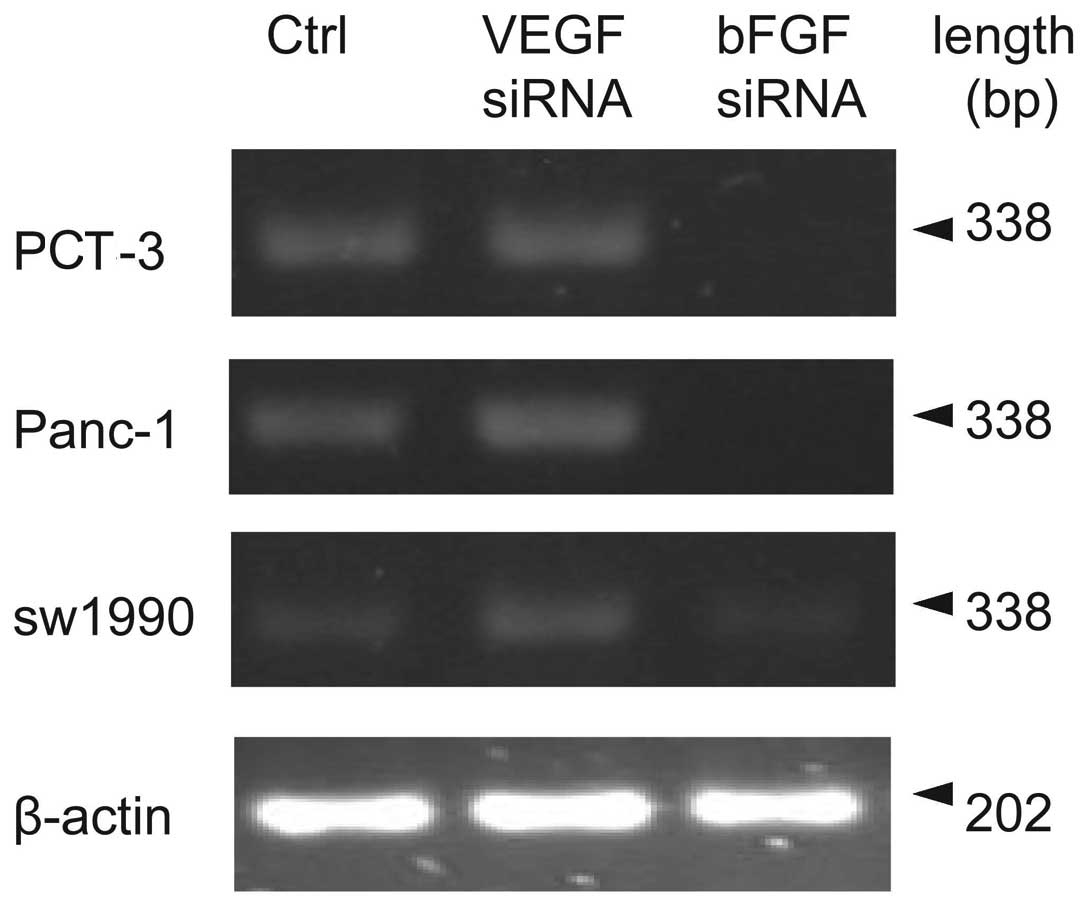
Effects of bFGF and VEGF siRNA
interference on bFGF mRNA expression in different pancreatic cell
lines
bFGF or VEGF siRNA was transfected into PCT-3,
Panc-1 and sw1990 cell lines. These are stable pancreatic cancer
cell lines that express bFGF and VEGF. Using RT-PCR, it was shown
that bFGF siRNA but not VEGF siRNA significantly reduced the bFGF
mRNA amount in the cell lines tested (P<0.01, compared to
control), as shown in Fig. 1 and
Table I. Notably. bFGF mRNA
expression was increased following VEGF siRNA treatment, especially
in Panc-1 cell line (P<0.05, compared to control).
 | Table IbFGF mRNA relative expression in
different pancreatic cell lines following VEGF and bFGF siRNA
treatment, respectively. |
Table I
bFGF mRNA relative expression in
different pancreatic cell lines following VEGF and bFGF siRNA
treatment, respectively.
| | PCT-3
| Panc-1
| sw1990
|
|---|
| Group | n | Mean value | Fold | Mean value | Fold | Mean value | Fold |
|---|
| Control | 3 | 0.42±0.02 | | 0.62±0.03 | | 0.32±0.03 | |
| VEGF siRNA | 3 | 0.50±0.05 | 1.19 | 0.81±0.05a | 1.31 | 0.36±0.06 | 1.12 |
| bFGF siRNA | 3 | 0.08±0.01b | 0.19 | 0.10±0.04b | 0.16 | 0.15±0.03b | 0.47 |
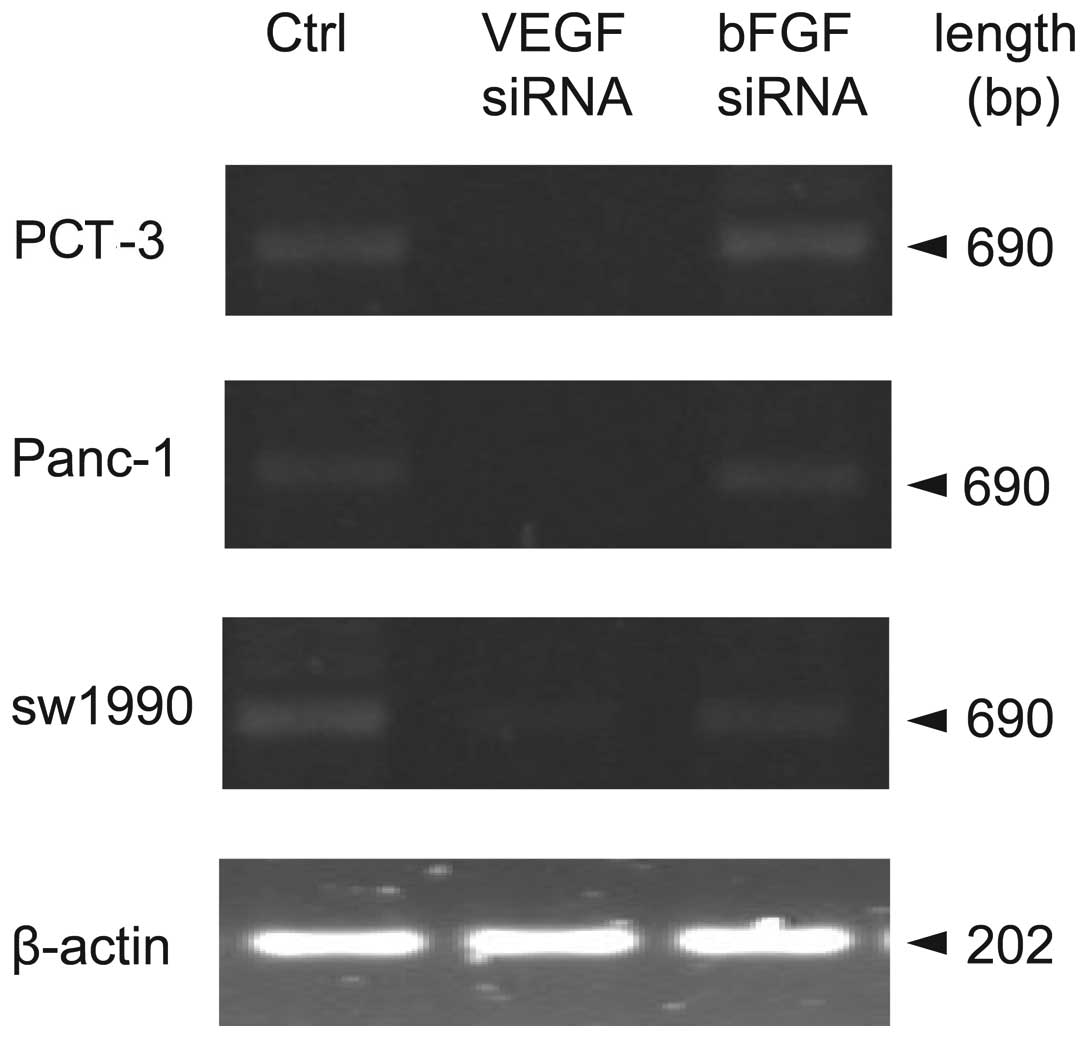
Effects of bFGF and VEGF siRNA
interference on VEGF mRNA expression in different pancreatic cell
lines
To test the knockdown effects of both types of siRNA
on expression level of VEGF, a similar strategy to detect VEGF
expression was used. RT-PCR results (Fig. 2 and Table II) showed bFGF siRNA did not change
VEGF mRNA expression in PCT-3 Panc-1 or sw1990 cells (P>0.05,
compared to control). However, VEGF mRNA expression was
significantly reduced in the three pancreatic cell lines after VEGF
siRNA treatment (P<0.05, P<0.01 compared to control).
| Table IIVEGF mRNA relative expression in
different pancreatic cell lines following VEGF and bFGF siRNA
treatment, respectively. |
Table II
VEGF mRNA relative expression in
different pancreatic cell lines following VEGF and bFGF siRNA
treatment, respectively.
| | PCT-3
| Panc-1
| sw1990
|
|---|
| Group | n | Mean value | Fold | Mean value | Fold | Mean value | Fold |
|---|
| Control | 3 | 0.28±0.02 | | 0.24±0.02 | | 0.31±0.02 | |
| VEGF siRNA | 3 | 0.08±0.01b | 0.29 | 0.06±0.01b | 0.25 | 0.16±0.04a | 0.52 |
| bFGF siRNA | 3 | 0.26±0.03 | 0.93 | 0.22±0.03 | 0.92 | 0.29±0.02 | 0.94 |
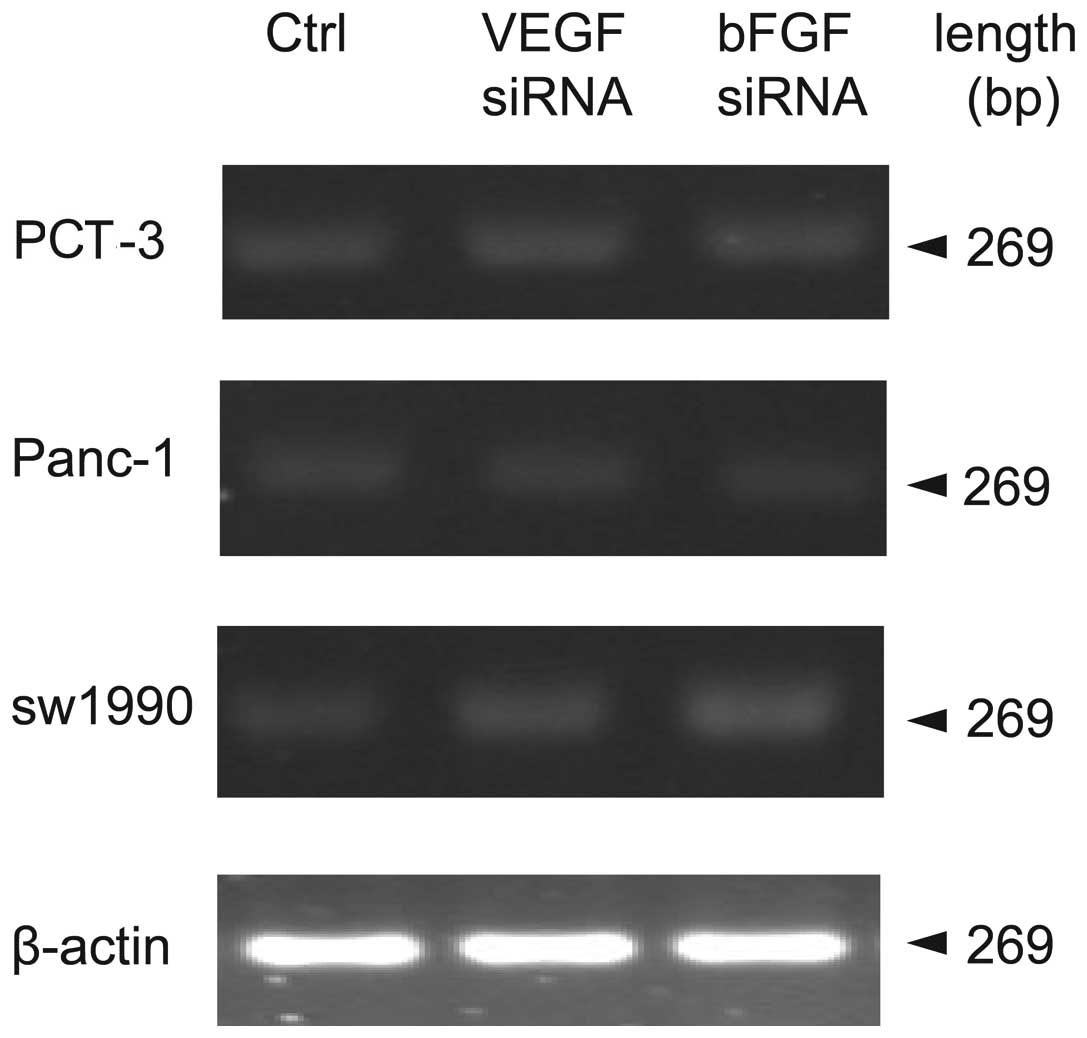
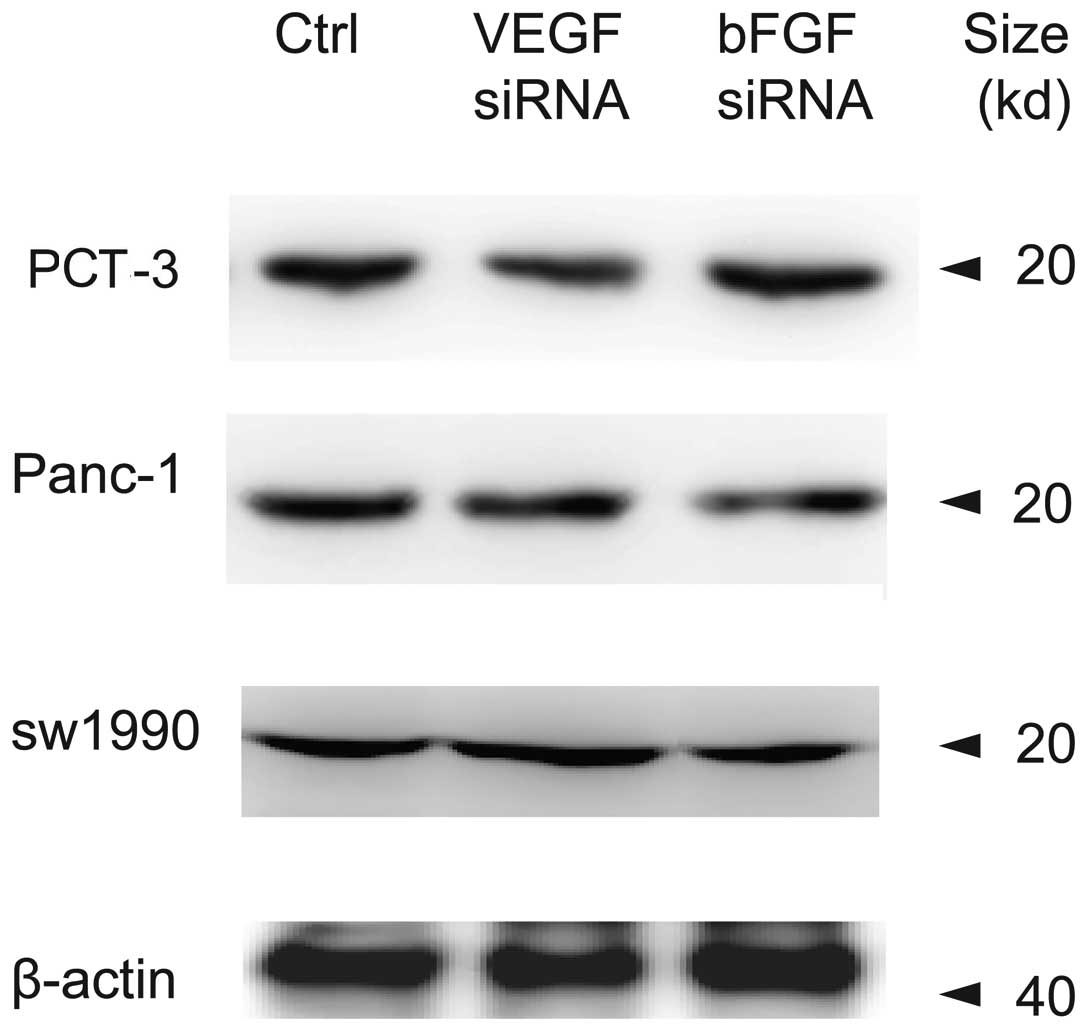
Effects of bFGF and VEGF siRNA
interference on endostatin mRNA and protein expression in different
pancreatic cell lines
Previous data showed that bFGF or VEGF siRNA was
able to dramatically reduce mRNA expression of bFGF or VEGF,
respectively. In order to determine the relationship between bFGF,
VEGF and endostatin, endostatin mRNA expression was measured by
RT-PCR following knockdown of bFGF and VEGF. As shown in Fig. 3 and Table III, neither bFGF nor VEGF siRNA
changed endostatin mRNA expression in PCT-3, Panc-1 and sw-1990
cells (P>0.05, compared to control). To further determine
whether bFGF siRNA or VEGF siRNA affected endostatin protein
expression, western blot assay was employed and results (Fig. 4) showed that neither VEGF nor bFGF
siRNA changed endostatin protein expression levels in the cell
lines studied.
| Table IIIRelative expression of endostatin mRNA
in different pancreatic cell lines following VEGF siRNA and bFGF
siRNA treatment, respectively. |
Table III
Relative expression of endostatin mRNA
in different pancreatic cell lines following VEGF siRNA and bFGF
siRNA treatment, respectively.
| | PCT-3
| Panc-1
| sw1990
|
|---|
| Treatment | n | Mean value | Fold | Mean value | Fold | Mean value | Fold |
|---|
| Control | 3 | 0.32±0.01 | | 0.28±0.04 | | 0.31±0.03 | |
| VEGF siRNA | 3 | 0.30±0.01 | 0.94 | 0.26±0.05 | 0.93 | 0.34±0.06 | 1.10 |
| bFGF siRNA | 3 | 0.33±0.03 | 1.07 | 0.32±0.07 | 1.14 | 0.33±0.04 | 1.06 |
Effects of bFGF and VEGF siRNA
interference on endostatin concentration in culture supernatant in
different pancreatic cell lines
Previous studies showed that the level of exocrine
endostatin played important biological roles in the
micro-environment of pancreatic cancer cells (8). In order to evaluate the influence of
knocking down bFGF and VEGF on the secreted endostatin level, an
ELISA technique was used to detect endostatin concentrations in the
culture supernatant of different pancreatic cell lines with
different treatments. Endostatin concentrations were significantly
reduced following treatment with bFGF and VEGF siRNA in the cell
lines studied (P<0.01, compared to control; Table IV).
| Table IVConcentrations of endostatin in the
supernatant of different pancreatic cell lines following bFGF and
VEGF siRNA treatments, respectively. |
Table IV
Concentrations of endostatin in the
supernatant of different pancreatic cell lines following bFGF and
VEGF siRNA treatments, respectively.
| Group | n | sw1990 (ng/ml) | PCT-3 (ng/ml) | Panc-1 (ng/ml) |
|---|
| Control | 5 | 2.98±0.10 | 15.42±0.75 | 28.61±3.74 |
| VEGF siRNA | 5 | 0.63±0.12a | 11.07±0.30a | 13.67±2.25a |
| bFGF siRNA | 5 | 1.68±0.19a | 10.69±0.14a | 14.29±3.65a |
Discussion
Tumor spread and metastasis are based on
angiogenesis. Activation and modulation of angiogenesis are
dependent on the interaction between pro- and anti-angiogenic
factors (9). Several studies have
been performed and it has been recognized that different angiogenic
growth factors play important roles in the modulation of
neovascularization in all types of tumor progression.
Anti-angiogenic therapy is, therefore, one of the most important
antitumor therapeutic strategies (10). Anti-angiogenic therapy focuses on
two approaches: inhibition of pro-angiogenic factors, or
enhancement of anti-angiogenic factors. There are some
anti-angiogenic compounds under development in clinical trials.
Their therapeutic strategy is often focused on a single therapeutic
target. Studies on the interaction between pro-angiogenic factors
and anti-angiogenic factors are, however, still scarce. Studies on
the interaction may provide a strong basis for joint multi-target
anti-angiogenic therapy.
As a type of vasculature-lacking tumor, pancreatic
cancer shows atypical angiogenesis accompanied by endothelial cell
proliferation, and uneven distribution of vascular morphology
(11). This suggests that local
factors may account for the pancreatic angiogenic mechanism. A
large number of studies confirm that expression of many angiogenic
factors, such as VEGF, bFGF and endostatin are elevated in the
pancreatic cancer tissue (12),
suggesting angiogenic factors play critical roles in pancreatic
cancer. Our study shows that VEGF and bFGF siRNAs inhibit VEGF and
bFGF, respectively, and also modulate endostatin secretion. A
previous study showed that the level of exocrine endostatin was
higher than that of intracellular endostatin, suggesting endostatin
may play a biological role in the microenvironment of pancreatic
cancer cells (8). Our study further
indicates that siRNAs of VEGF and bFGF dramatically reduce
endostatin concentrations in the culture medium without affecting
intracellular endostatin mRNA and protein expression. This
suggested that the inhibitory effects of VEGF and bFGF siRNA on
endostatin secretion may occur following its transcription and
translation, and that modulating protein secretion may be its main
mechanism. Brammer et al(13) confirmed that collagen XVIII is
expressed in pancreatic cancer cells, and can be released into the
medium. Heljasvaara et al(14) showed that particular matrix
metalloproteinases (MMPs) can degrade collagen XVIII and generate
biological, suggesting that endostatin is hydrolysized from
collagen XVIII. Endostatin is expressed differently in variable
pancreatic cancer cell lines, which is modulated by TNF-α-dependent
elastase. A study by Nilsson et al(15) showed that estradiol and tamoxifen
regulate endostatin expression via MMP-2/MMP-9 in breast cancer. A
previous study (16) showed that
the p38MAPK pathway is involved in the modulation of MMP function,
suggesting that the MAPK pathway could be critical for VEGF/bFGF
biological function. Further research is required to confirm
this.
In pancreatic cancer cells, the regulation of
angiogenic factor secretion is complicated and this regulation is
closely related to tumor progression (17,18).
In the processes of pancreatic cancer occurrence, development and
metastasis, pro-angiogenic factors and anti-angiogenic factors
modulate each other while each factor has its tumor biological
functions (19,20). Further research is required to
elucidate the interaction mechanism. This will provide novel
therapeutic targets for anti-angiogenic treatment of pancreatic
cancer and other tumors. In the development of anti-angiogenic
treatment strategies, combined therapy for different targets may
yield better treatment results, and in-depth study of modulation
mechanisms will further improve the joint treatment effects. The
correlation between angiogenic factors in different cancer cell
lines may not be the same. Therefore, individualized treatment
should be considered when developing joint anti-angiogenic
treatment programs.
Acknowledgements
This study was supported in part by
Hebei Province Office of Science and Technology support projects,
China (12276104D-76 to C. Yan).
References
|
1
|
Yan CQ, Zhao YP, Dai MH, et al: The
relationship between the changes of proangiogenic factors serum
concentrations and progression of pancreatic carcinoma patients.
Zhonghua Wai Ke Za Zhi. 45:496–498. 2007.(In Chinese).
|
|
2
|
Dai MH, Yan CQ, Zhao YP and Song YM:
Relationship between changes of serum concentrations of
antiangiogenic factors and disease progression in patients of
pancreatic carcinoma. Zhonghua Wai Ke Za Zhi. 45:1199–1201.
2007.(In Chinese).
|
|
3
|
Yan CQ and Zhao YP: Expression of
angiogenic factors in pancreatic carcinoma cell and their
significance. Zhonghua Wai Ke Za Zhi. 47:787–790. 2009.(In
Chinese).
|
|
4
|
Yan CQ and Zhao YP: The significance of
VEGF siRNA and bFGF siRNA in invasion and proliferation of
pancreatic carcinoma cell. Zhonghua Wai Ke Za Zhi. 48:610–614.
2010.(In Chinese).
|
|
5
|
Sakurai T and Kudo M: Signaling pathways
governing tumor angiogenesis. Oncology. 81(Suppl 1): 24–29. 2011.
View Article : Google Scholar
|
|
6
|
Ohlund D, Ardnor B, Oman M, et al:
Expression pattern and circulating levels of endostatin in patients
with pancreas cancer. Int J Cancer. 122:2805–2810. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wigmore SJ: Endostatin in the pancreas. Br
J Cancer. 92:5–6. 2005. View Article : Google Scholar
|
|
8
|
Rahbari NN, Schmidt T, Falk CS, et al:
Expression and prognostic value of circulating angiogenic cytokines
in pancreatic cancer. BMC Cancer. 11:2862011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jain RK: Normalization of tumor
vasculature: an emerging concept in antiangiogenic therapy.
Science. 307:58–62. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Eichholz A, Merchant S and Gaya AM:
Anti-angiogenesis therapies: their potential in cancer management.
Onco Targets Ther. 3:69–82. 2010.PubMed/NCBI
|
|
11
|
Korc M: Pathways for aberrant angiogenesis
in pancreatic cancer. Mol Cancer. 2:82003. View Article : Google Scholar
|
|
12
|
Benckert C, Thelen A, Cramer T, et al:
Impact of microvessel density on lymph node metastasis and survival
after curative resection of pancreatic cancer. Surg Today.
42:169–176. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Brammer RD, Bramhall SR and Eggo MC:
Endostatin expression in a pancreatic cell line is modulated by a
TNFalpha-dependent elastase. Br J Cancer. 93:1024–1028. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Heljasvaara R, Nyberg P, Luostarinen J, et
al: Generation of biologically active endostatin fragments from
human collagen XVIII by distinct matrix metalloproteases. Exp Cell
Res. 307:292–304. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nilsson UW, Garvin S and Dabrosin C: MMP-2
and MMP-9 activity is regulated by estradiol and tamoxifen in
cultured human breast cancer cells. Breast Cancer Res Treat.
102:253–261. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhong J, Gencay MM, Bubendorf L, et al:
ERK1/2 and p38 MAP kinase control MMP-2, MT1-MMP, and TIMP action
and affect cell migration: a comparison between mesothelioma and
mesothelial cells. J Cell Physiol. 207:540–552. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xie L, Duncan MB, Pahler J, et al:
Counterbalancing angiogenic regulatory factors control the rate of
cancer progression and survival in a stage-specific manner. Proc
Natl Acad Sci USA. 108:9939–9944. 2011. View Article : Google Scholar
|
|
18
|
Albrecht I, Kopfstein L, Strittmatter K,
et al: Suppressive effects of vascular endothelial growth factor-B
on tumor growth in a mouse model of pancreatic neuroendocrine
tumorigenesis. PLoS One. 5:e141092010. View Article : Google Scholar
|
|
19
|
Pereira ER, Liao N, Neale GA and
Hendershot LM: Transcriptional and post-transcriptional regulation
of proangiogenic factors by the unfolded protein response. PLoS
One. 5:e125212010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Schuch G, Kisker O, Atala A and Soker S:
Pancreatic tumor growth is regulated by the balance between
positive and negative modulators of angiogenesis. Angiogenesis.
5:181–190. 2002. View Article : Google Scholar : PubMed/NCBI
|