Contents
Introduction
Hypoxia-inducible factor (HIF)
Glucose transporter (GLUT) proteins
Conclusion
Introduction
Cancer is the focus of scientific studies and
investigations into oral pathology, as it is a significant cause of
morbidity and mortality, and has a poor prognosis. This has
encouraged additional studies that are concerned with factors that
may alter the development of this disease. Oral squamous cell
carcinoma, or oral epidermoid carcinoma (OEC), is the most common
clinical entity among the malignant oral neoplasias, accounting for
∼90% of oral cancer cases (1,2).
OEC presents with heterogeneous clinical,
pathological and biological aspects, and its development and
progression are promoted by multiple alterations at a cellular and
molecular level in the squamous epithelium (3,4). A
loss of heterozygocity on specific chromosomes, microsatellite
instability and mutations in the tumor suppressor genes, p53 and
p16, which are important in cell cycle regulation, are associated
with distinct phases of tumor progression (5). In order to predict the potential
aggressiveness of OEC, factors in addition to the tumor
classification and staging system [the tumor node metastasis (TNM)
staging system] ought to be considered. Thus, scientific studies
have focused on the involvement of biomarkers in the progression of
OEC, by means of various laboratory methods (6,7).
The biological behavior of malignant neoplasias is
complex. The growth and dissemination of cancer depend, not only on
neoplastic cell proliferation, but also on the normal host tissue
responses; a variety of interactions have been observed between
tumor cells, the vascular network, the immune system and supportive
conjunctive tissue (8). In order to
understand the tumor biology, the energy metabolism of tumor cells
has been investigated (9).
Establishing molecular signatures and mechanisms, in
order to understand tumor progression, may facilitate the
identification of novel predictive and prognostic markers, in
addition to new therapeutic targets for the treatment of cancer
(5). This is due to the fact that
the TMN staging system and the histopathological degree of
differentiation are insufficient for predicting the prognosis of
OEC (10). Thus, pathways that have
an influence on tumor progression have been the targets of previous
studies, such as those involving HIF and GLUT proteins (8,11).
Solid tumors may present rapid growth, which exceeds
the necessary vascularization, and thus the supply of nutrients and
oxygen becomes insufficient for the tumor tissue. Under these
conditions of hypoxia, a signaling pathway, the HIF-1 pathway, is
activated, which acts as a modulator of an adaptive response to the
reduction of oxidative stress in the tumor microenvironment
(10). In a hypoxic tumor medium,
HIF may act on the genes that regulate glycolytic metabolism,
promoting an increase in the rate of glucose uptake by the cell
through the transcription of GLUTs. This uptake allows the cell to
obtain energy by means of glucose metabolism, as a metabolic change
from mitochondrial respiration to glycolysis is frequently observed
in the neoplastic cell. Therefore, the increase in glucose uptake
by the GLUTs is essential, as it facilitates an increased survival
time of the neoplastic cell (12).
The aim of the present study was to conduct a
literature review of oral squamous cell carcinoma, focusing on the
factors that influence the metabolism of oxygen during tumor growth
and progression, and emphasizing the roles of HIF and GLUTs in oral
carcinogenesis. A search strategy was implemented in PubMed,
selecting only English articles in the period from 2002 to 2012,
and which used the terms or combinations of the following
descriptors: Oral squamous cell carcinoma, HIF, hypoxia-related
proteins, GLUT proteins and tumor progression. A total of 36
scientific studies were selected, which involved laboratory
investigations, particularly those including immunohistochemistry,
uni- and multivariate analyses, and retrospective, comparative and
multi-centric studies. These studies were concerned with the role
of HIF and GLUTs in the carcinogenesis of oral squamous cells.
Hypoxia-inducible factor (HIF)
With rapid tumor growth and expansion, the
intratumoral regions may present with hypoxia, which is a state of
reduction in oxidative stress to levels below the normal limit, and
this is commonly observed in diverse malignant tumors.
Characteristically, OEC is a locally aggressive neoplasm with rapid
progression, and the oxygen concentration is significantly reduced
in OEC tumors (13). All cells of
the body require oxygen to perform their normal metabolic
functions, which include oxidative phosphorylation. The capacity of
cancerous cells to adapt to hypoxia, whether transitory or
extensive, is essential for tumor survival. It has been suggested
that the invasive and metastatic nature of OEC is a consequence of
its adaptation to the hypoxic microenvironment (7,14). An
important mechanism for adaptation to reduced oxygen concentrations
is the regulation of HIF-1. Oxygenation in solid neoplasms depends
on the supply of oxygen and its consumption by the tumor cells.
Therefore, hypoxia is a common phenomenon in various types of
malignant neoplasms, contributing to the progression of cancer and
the selection of the more aggressive phenotype (7,15). The
high metabolic rates of tumor cells induce intratumoral hypoxia;
this hypoxic stress induces the expression of a complex of genes
that regulate homeostasis of the oxygen supply. In addition, HIF-1
is the master regulator of the transcription of these genes.
Therefore, HIF-1 mediates adaptive responses at cellular and
systemic levels for the maintenance of homeostasis, which is the
main mechanism whereby tumor cells respond to acidosis and hypoxic
stress (12).
HIF-1 is a basic, helix-loop-helix-PER-ARNT-SIM
(PAS) heterodimer composed of α and β subunits. HIF-1α is the
oxygen-sensitive subunit that dimerizes with the constitutively
expressed β subunit (HIF-1β) (10).
HIF-1β, also known as aryl hydrocarbon-receptor nuclease
translocator (ARNT), is a 91- to 94-kDa protein. HIF-1α is encoded
by a gene located on chromosome 14 (14q21-q24), and is stabilized
and accumulated in response to hypoxia. Under the conditions of a
normal oxygen supply, HIF-1α is not usually detected or stabilized
(7), and is hydroxylated at the
proline 402 and 564 (Pro-402 and -564) residues. Following
hydroxylation, the HIF-1α residues bond to the von Hippel-Lindau
tumor suppressor protein (pVHL) and are rapidly destroyed by an
enzyme, E3 ubiquitin ligase (Fig.
1) (16). Another mechanism
whereby HIF-1 is inhibited is by the HIF-1 inhibition factor, which
hydroxylates asparagine 803 by means of the transactivation domain,
and consequently blocks the p300-CREB-binding protein (CBP)
coactivator bond (7).
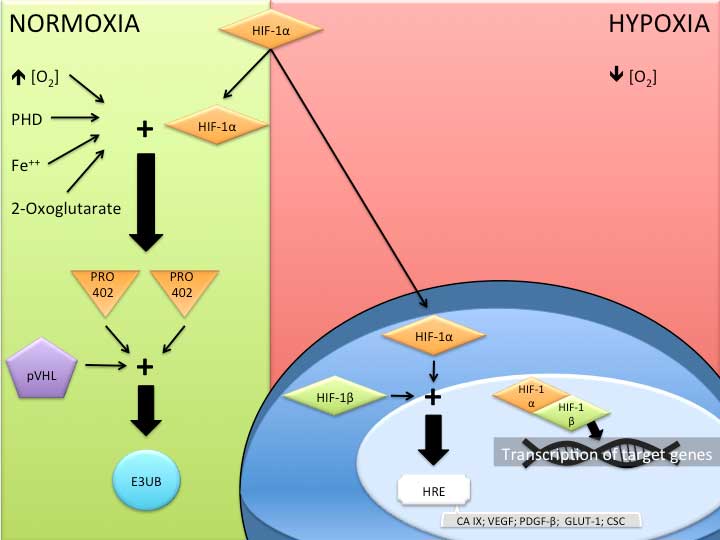 | Figure 1.Mechanism of action of hypoxia-induced
factor 1 (HIF-1) in normoxic and hypoxic environments. PHD, prolyl
hydroxylase domain; PRO 402, proline 402; pVHL, von Hippel-Lindau
tumor supressor; E3UB, E3 ubiquitin ligase; HRE, hypoxia response
element; CA IX, carbonic anhydrase IX; VEGF, vascular endothelial
growth factor; PDGF-β, platelet derived growth factor-β; GLUT 1,
glucose transporter 1; CSC, cancer stem cell. |
The suppression of tumor suppressor genes, such as
p53, phosphatase and tensin homolog (PTEN) and pVHL, may increase
the expression of HIF-1α in tumor cells, as well as the action of
oncogenes [including ras, SRC and phosphatidylinositide 3 kinase
(PI3K) proteins), growth factors (such as the endothelial growth
factor) and cytokines (including prostaglandin E2) (12). The role of HIF-1α in carcinogenesis
is to signal the message of hypoxia to the cell nucleus, which
promotes a response to hypoxic stress. This response involves
influencing the control of >100 genes associated with tumor
adaptation, such as glycolytic transport, and with alterations in
the tumor microenvironment, by stabilizing the pH and by
angiogenesis (7). The action of
HIF-1α on cancer stem cells (CSCs) has also been suggested, as a
high rate of survival, proliferation and expression of these cells
has been observed in a hypoxic medium when compared with cells that
are in homeostasis (10). In
addition, HIF-1α may control the expression of vascular endothelial
growth factor (VEGF) and erythropoietin (EPO) genes, resulting in
the promotion of tumor angiogenesis, as both stimulate endothelial
cell proliferation and migration, and EPO alone promotes an
increase in the proliferation and growth of various human
neoplasias. Thus, an increase in HIF-1α expression may influence
oral cancer progression by promoting tumor angiogenesis and by
direct stimulation of tumor cell growth (17,18).
It has been demonstrated that HIF-1α expression has
a significant effect on tumor progression. Patients with oral
neoplasias with an elevated rate of HIF-1α expression present a
3.49-fold lower survival rate when compared with such patients with
limited expression of this protein (7). HIF-1α is involved in tumor
carcinogenesis and progression; Koukourakis et al (19) observed that overexpression of HIF-1α
was correlated with a more aggressive behavior of oral carcinoma
and an elevated resistance to radiotherapy treatment. In addition,
Lin et al (18) observed
that an increase in HIF-1α expression was correlated with a poor
prognosis for cases of OEC, by univariate analysis.
Invasion and metastasis are key characteristics of
malignant neoplasias and represent the primary cause of
cancer-related mortality (13). It
has been suggested that HIF-1α is correlated with a higher rate of
aggressiveness and metastasis, promoting mutations and stimulating
angiogenesis by the activation of VEGF, inducing the proliferation,
differentiation and migration of vascular endothelial cells by
means of the increase in capillary permeability, as well as the
reduction in apoptosis (20). Tumor
cells interact with extracellular matrix (ECM) proteins by
molecular rearrangement on the cell surface, and cell adhesion and
integrin molecules promote binding to the ECM. Thus, tumor hypoxia
induces an accumulation of HIF-1α, which in turn increases the
expression of integrin α5 and fibronectin in the tumor cells,
facilitating binding to the ECM that is rich in fibronectin, and
providing cell invasion potential at the cell surface (13).
In addition to hypoxia, tumor cells suffer
acidosis-induced stress and an increase in interstitial fluid
pressure, and present a greater glucose requirement (12). Studies have demonstrated that
cancerogenous cells metabolize a substantial quantity of
extracellular glucose, and that a subset of cells may utilize
glutamine, a free amino acid abundant in muscle tissue that may act
as a source of energy (21–28). In general, neoplastic cells in a
hypoxic medium require the uptake of glucose, with the aim of
increasing energy levels by means of glucose metabolism (to obtain
energy in the form of ATP). Therefore, it is necessary for the
cells to facilitate the process of glucose uptake from the
extracellular medium. Various GLUTs have been observed in malignant
tumors, revealing an important role of GLUTs in the maintenance of
neoplastic cell survival, and tumor progression and growth
(10).
Glucose transporter (GLUT) proteins
Cancerogenous cells have a high rate of glucose
uptake and glycolytic metabolism, and thus, tumor cells exhibit
significantly different metabolic activity compared with that
involved in normal eukaryotic cell homeostasis. When there is a
limited oxygen supply, and it is therefore not possible to obtain
energy by mitochondrial respiration, normal eukaryotic cells
maximize their energy production by the combination of traditional
energy pathways, including glycolysis, the carboxylic acid cycle
and the electron transport chain. Thus, the cells efficiently
convert the glucose molecule into carbon dioxide and water,
maximizing ATP production and potentially reducing NADPH production
(29–31). Normal cells obtain only 10% of their
energy by glycolysis, with the remainder being the result of
mitochondrial respiratory activity. However, tumor cells obtain the
majority of their energy by glycolysis, maintaining elevated rates
of lactate production, which is sufficient for tumor cell survival
in a hypoxic environment. Glycolysis generates a net gain of only
two molecules of ATP per glucose molecule, a markedly smaller
amount of energy compared with the net gain of 38 molecules of ATP
produced by respiration. Thus, neoplastic cells require an
increased glucose uptake that is essential to obtain sufficient
energy (10).
Glucose is transported into the cell by means of
GLUTs, which are present in all type of cells, and have a variable
availability in the tissue distribution and a variable affinity for
glucose. GLUTs are a family of proteins that mediate glucose
transport through the membrane without depending on energy
(26,29). At present, various isoforms of GLUT
have been described, and the expression of these is cell-specific
and subject to extracellular medium control. Functionally, the
GLUTs regulate the movement of glucose between the extracellular
medium and the intracellular compartments, maintaining the glucose
supply available for cell metabolism (32). The GLUT family was originally
proposed to comprise 12 members (Table
I); however, novel forms of GLUTs have been described,
resulting in a total of 14 known GLUTs (33) that have different affinities for
glucose and other hexoses, such as fructose (24,25).
This family of transmembrane proteins appears to be regulated by
proto-oncogenes, which are present in normal cells, and growth
factors. The transport stimulation mechanisms include the
translocation of GLUTs to the plasma membrane, and the activation
of the transporters at the presynaptic terminals in the plasma
membrane, which are regulated by a PI3-kinase-dependent signaling
pathway (22,25).
 | Table I.The GLUT family. |
Table I.
The GLUT family.
| Author, year | Isoform | Main tissue
localization | Transport |
|---|
| Muecker, 1985; Gould,
1991 | GLUT-1 | Erythrocytes, brain
and ubiquitous | Glucose |
| Fukamoto, 1988;
Gould, 1991 | GLUT-2 | Liver, pancreas,
intestine and kidney | Glucose (low
affinity) and fructose |
| Kayano, 1988;
Gould, 1991 | GLUT-3 | Brain | Glucose (high
affinity) |
| Kukamoto, 1989;
James, 1989 | GLUT-4 | Heart, muscle, WAT,
BAT and brain | Glucose (high
affinity) |
| Kayano, 1990;
Davidson, 1992 | GLUT-5 | Intestine, testes
and kidney | Fructose and
glucose (very low affinity) |
| Doege, 2000;
Lisinski, 2001 | GLUT-6 | Brain, spleen and
leucocytes | Glucose |
| Joost e Thorens,
2001 | GLUT-7 | ND | ND |
| Carayannopoulos,
2000; Doege, 2000; Ibberson, 2000; Lisinski, 2001 | GLUT-8 | Testes, brain and
other tissues | Glucose |
| Phay, 2000 | GLUT-9 | Liver and
kidney | ND |
| Dawson, 2001;
McVie-Wylie, 2001 | GLUT-10 | Liver and
pancreas | Glucose |
| Doege, 2001; Wu,
2002; Sasaki, 2001 | GLUT-11 | Heart and
muscle | Glucose (low
affinity) and fructose |
| Rogers, 2002 | GLUT-12 | Heart, prostate,
muscle, small intestine, | ND WAT and
brain |
The first GLUTs of this family to be identified were
GLUT-1 and -3, which are expressed at variable levels in various
human tissues, suggesting that these constitutive isoforms may be
responsible for basal glucose uptake. GLUT-2 is expressed in the
hepatocytes, where it mediates glucose transport through the
membrane. GLUT-4 is expressed in tissues in which glucose uptake is
regulated by insulin, including adipose tissue, and skeletal and
myocardial muscle, by acute insulin stimulation. The high-capacity
transporters, such as GLUT-2 and -4, are normally restricted to
cells that do not divide. GLUT-5 is expressed in the small
intestine and sperm cells, acting as a fructose transporter
(22). GLUT-1 is not detected in
large proportions in the cells of normal tissues, with the
exception of erythrocytes, germinative cells of the testes, renal
tubules, the perineurium of the peripheral nerves and endothelial
cells of the blood-brain barrier. Its overexpression has been
observed in various types of malignant tumors, such as carcinomas
of the colon, lung, breast, esophagus, stomach, ovary and biliary
vesicle, as well as malignant mesothelioma, and squamous cell
carcinoma of the head and neck (25).
It has been demonstrated that solid malignant tumors
with a rapid rate of growth and proliferation are characterized by
elevated rates of glucose use and uptake, due to the energy demand
required for uncontrolled proliferation, based on the increased
and/or atypical expression of multiple isoforms of GLUT (10,22,24,25,29).
This elevated expression of GLUTs is correlated with a risk of
metastasis and a worse prognosis (9,11,32,33).
Studies, such as the study by Weiner et al
(34), have highlighted the
importance of the correlation between the immunoreactivity of
GLUTs, including GLUT-1, and clinical and radiographical findings,
when the cytological findings are ambiguous. Ayala et al
(11) performed an
immunohistochemical marking of GLUT-1 and -3 in OEC, and observed
an elevated expression of GLUT-1 in the majority of cases; whereas
GLUT-3 was identified in 21.1% of the studied OEC samples, in
accordance with the fact that this protein is not found in the
normal oral epithelium. This suggested that GLUT-1 may be used as
an indicator of prognosis, due to its high immunoexpression that is
likely to be correlated with the increase in glycolytic metabolism
in more aggressive neoplastic cells. A study by Kunkel et al
(35) analyzed the immunoexpression
of GLUT-1 in 118 cases of OEC and observed an inversely
proportional correlation between the expression of this protein and
the survival rate of patients. This suggested that glucose
transport acted in a determinant manner for the promotion of the
tumor phenotype, and that GLUT-1 may be considered a negative
prognostic survival biomarker in patients with OECs. This was
concordant with the study by Ayala et al (11), in which it was proposed that this
protein be used as a potential marker of tumor progression.
Fukuzumi et al (22) observed the elevated immunoexpression
of GLUT-1 in oral squamous cell carcinoma, and suggested that this
protein may be important in the pathogenesis of the disease. The
atypical expression of GLUTs in oral squamous cell carcinoma does
not necessarily signify an increase in the rate of glucose
transport to the tumor cell, in order to promote substrate levels
for obtaining the energy that is important for tumor cell survival
and growth; it may involve the deregulation of gene expression in
specific tissues and cancerogenous cells. Studies have demonstrated
that an overexpression of GLUT-1 and -3 was associated with certain
solid tumors with greater potential malignancy and lower survival
rates (28,29,36).
Additionally, Ohba et al (25) demonstrated the use of GLUT-1 as an
indicator of the depth of invasion with a trend towards a worse
prognosis in patients, as a result of an increase in the
possibility of nodal metastasis.
The immunoexpression of GLUT-1 and -3 may be
associated with the clinical profile of a group of cells in a
subset of patients with oral squamous cell carcinoma, who present
with a distinctly poor prognosis. The increase in glycolytic
metabolism through the action of these proteins in more aggressive
tumor cells indicates a potential prognostic value that may enable
patients to be stratified with regard to risk (11). Fluorine-18 fluoro-2-deoxy-D-glucose
(FDG) is a glucose analog, which, like glucose, is present at high
levels in malignant tumors. Positron emission tomography (PET) with
FDG has become a novel, noninvasive, imaging diagnostic field. This
technique is utilized and favors the initial diagnosis, the
evaluation of the extension of the disease and prognosis, and the
treatment planning and monitoring, in addition to the detection of
recurrences of the disease (28).
Although it remains widely used, the TNM staging
system alone is not capable of establishing a true assessment of
the patient’s individual prognosis. In order to predict the
potential aggressiveness of oral neoplasias, factors additional to
the TNM staging system ought to be considered, such as the use of
immunohistochemical biomarkers, with the goal of determining
whether they exhibit a greater advantage as a prognostic factor
(7).
Studies investigating GLUT-1 and HIF-1 proteins
identified that these proteins presented with elevated expression
in oral OEC, and were correlated with a worse prognosis. This
suggests that GLUTs and HIF-1 may be used as markers of tumor
prognosis and progression (10).
Conclusion
Recent studies have encouraged the analysis of HIF-1
and GLUTs in OEC, assessing these proteins as potential prognostic
markers. This, in turn, enables the optimization of therapeutic
strategies in the treatment of mouth cancer, as these proteins may
be associated with the events of carcinogenesis and exert a
significant impact on the survival of patients affected by OEC.
References
|
1.
|
Massano J, Regateiro FS, Januário G and
Ferreira A: Oral squamous cell carcinoma: review of prognostic and
predictive factors. Oral Surg Oral Med Oral Pathol Oral Radiol
Endod. 102:67–76. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Scully C and Bagan J: Oral squamous cell
carcinoma: overview of current understanding of aetiopathogenesis
and clinical implications. Oral Dis. 15:388–399. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Kademani D, Bell RB, Bagheri S, et al:
Prognostic factors in intraoral squamous cell carcinoma: the
influence of histologic grade. J Oral Maxillofac Surg.
63:1599–1605. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Haddad RI and Shin DM: Recent advances in
head and neck cancer. N Engl J Med. 359:1143–1154. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Glazer CA, Chang SS, Ha PK and Califano
JA: Applying the molecular biology and epigenetics of head and neck
cancer in everyday clinical practice. Oral Oncol. 45:440–446. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Eckert AW, Kappler M, Schubert J and
Taubert H: Correlation of expression of hypoxia-related proteins
with prognosis in oral squamous cell carcinoma patients. Oral
Maxillofac Surg. 16:189–196. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Oliveira LR and Ribeiro-Silva A:
Prognostic significance of immunohistochemical biomarkers in oral
squamous cell carcinoma. Int J Oral Maxillofac Surg. 40:298–307.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Nagpal JK and Das BR: Oral cancer:
reviewing the present understanding of its molecular mechanism and
exploring the future directions for its effective management. Oral
Oncol. 39:213–221. 2003. View Article : Google Scholar
|
|
9.
|
Moreno-Sánchez R, Rodriguez-Enriquez S,
Saavedra E, Marin-Hernández A and Gallardo-Pérez JC: The
bioenergetics of cancer: is glycolysis the main ATP supplier in all
tumor cells? Biofactors. 35:209–225. 2009.PubMed/NCBI
|
|
10.
|
Eckert AW, Lautner MH, Schütze A, Taubert
H, Schubert J and Bilkenroth U: Coexpression of hypoxia-inducible
factor-1α and glucose transporter-1 is associated with poor
prognosis in oral squamous cell carcinoma patients. Histopathology.
58:1136–1147. 2011.
|
|
11.
|
Ayala FR, Rocha RM, Carvalho KC, et al:
GLUT1 and GLUT3 as potential prognostic markers for Oral Squamous
Cell Carcinoma. Molecules. 15:2374–2387. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Denko NC: Hypoxia, HIF1 and glucose
metabolism in the solid tumour. Nat Rev Cancer. 8:705–713. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Ryu MH, Park HM, Chung J, Lee CH and Park
HR: Hypoxia-inducible factor-1alpha mediates oral squamous cell
carcinoma invasion via upregulation of alpha5 integrin and
fibronectin. Biochem Biophys Res Commun. 393:11–15. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Pérez-Sayáns M, Suárez-Peñaranda JM, Pilar
GD, Barros-Angueira F, Gándara-Rey JM and García-García A:
Hypoxia-inducible factors in OSCC. Cancer Lett. 313:1–8. 2011.
|
|
15.
|
Vaupel P and Mayer A: Hypoxia in cancer:
significance and impact on clinical outcome. Cancer Metastasis Rev.
26:225–239. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Semenza GL: HIF-1: upstream and downstream
of cancer metabolism. Curr Opin Genet Dev. 20:51–56. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Keith B, Johnson RS and Simon MC: HIF1α
and HIF2α: sibling rivalry in hypoxic tumour growth and
progression. Nat Rev Cancer. 12:9–22. 2011.
|
|
18.
|
Lin PY, Yu CH, Wang JT, et al: Expression
of hypoxia-inducible factor-1 alpha is significantly associated
with the progression and prognosis of oral squamous cell carcinomas
in Taiwan. J Oral Pathol Med. 37:18–25. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Koukourakis MI, Bentzen SM, Giatromanolaki
A, et al: Endogenous markers of two separate hypoxia response
pathways (hypoxia inducible factor 2 alpha and carbonic anhydrase
9) are associated with radiotherapy failure in head and neck cancer
patients recruited in the CHART Randomized Trial. J Clin Oncol.
24:727–735. 2006. View Article : Google Scholar
|
|
20.
|
Kyzas PA, Stefanou D, Batistatou A and
Agnantis NJ: Hypoxia-induced tumor angiogenic pathway in head and
neck cancer: an in vivo study. Cancer Lett. 225:297–304. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Demasi AP, Costa AF, Altemani A, Furuse C,
Araújo NS and Araújo VC: Glucose transporter protein 1 expression
in mucoepidermoid carcinoma of salivary gland: correlation with
grade of malignancy. Int J Exp Pathol. 91:107–113. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Fukuzumi M, Hamakawa H, Onishi A, Sumida T
and Tanioka H: Gene expression of GLUT isoforms and VHL in oral
squamous cell carcinoma. Cancer Lett. 161:133–140. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Grover-McKay M, Walsh SA, Seftor EA,
Thomas PA and Hendrix MJ: Role for glucose transporter 1 protein in
human breast cancer. Pathol Oncol Res. 4:115–120. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Krzeslak A, Wojcik-Krowiranda K, Forma E,
et al: Expression of GLUT1 and GLUT3 glucose transporters in
endometrial and breast cancers. Pathol Oncol Res. 18:721–728. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Ohba S, Fujii H, Ito S, et al:
Overexpression of GLUT-1 in the invasion front is associated with
depth of oral squamous cell carcinoma and prognosis. J Oral Pathol
Med. 39:74–78. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Parente P, Coli A, Massi G, Mangoni A,
Fabrizi MM and Bigotti G: Immunohistochemical expression of the
glucose transporters Glut-1 and Glut-3 in human malignant melanomas
and benign melanocytic lesions. J Exp Clin Cancer Res. 27:342008.
View Article : Google Scholar
|
|
27.
|
Richardson SM, Knowles R, Tyler J,
Mobasheri A and Hoyland JA: Expression of glucose transporters
GLUT-1, GLUT-3, GLUT-9 and HIF-1alpha in normal and degenerate
human intervertebral disc. Histochem Cell Biol. 129:503–511. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Tian M, Zhang H, Nakasone Y, Mogi K and
Endo K: Expression of Glut-1 and Glut-3 in untreated oral squamous
cell carcinoma compared with FDG accumulation in a PET study. Eur J
Nucl Med Mol Imaging. 31:5–12. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Sandulache VC and Myers JN: Altered
metabolism in head and neck squamous cell carcinoma: an opportunity
for identification of novel biomarkers and drug targets. Head Neck.
34:282–290. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Warburg O: On the origin of cancer cells.
Science. 123:309–314. 1956. View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Vander Heiden MG, Cantley LC and Thompson
CB: Understanding the Warburg effect: the metabolic requirements of
cell proliferation. Science. 324:1029–1033. 2009.PubMed/NCBI
|
|
32.
|
Macheda ML, Rogers S and Best JD:
Molecular and cellular regulation of glucose transporter (GLUT)
proteins in cancer. J Cell Physiol. 202:654–662. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Wood IS and Trayhurn P: Glucose
transporters (GLUT and SGLT): expanded families of sugar transport
proteins. Br J Nutr. 89:3–9. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34.
|
Weiner MF, Miranda RN, Bardales RH, et al:
Diagnostic value of GLUT-1 immunoreactivity to distinguish benign
from malignant cystic squamous lesions of the head and neck in
fine-needle aspiration biopsy material. Diagn Cytopathol.
31:294–299. 2004. View
Article : Google Scholar
|
|
35.
|
Kunkel M, Reichert TE, Benz P, et al:
Overexpression of Glut-1 and increased glucose metabolism in tumors
are associated with a poor prognosis in patients with oral squamous
cell carcinoma. Cancer. 97:1015–1024. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36.
|
Kunkel M, Moergel M, Stockinger M, et al:
Overexpression of GLUT-1 is associated with resistance to
radiotherapy and adverse prognosis in squamous cell carcinoma of
the oral cavity. Oral Oncol. 43:796–803. 2007. View Article : Google Scholar : PubMed/NCBI
|














