Introduction
Being a worldwide malignant liver tumor, gastric
cancer ranks the fourth in frequency among common human solid
tumors and the second leading cause of cancer-related mortality
(1). In China, gastric cancer
incidence ranks second only to lung cancer and accounts for
approximately half of the global incidence. In spite of the
improvement in surgical and multimodal therapy, the prognosis of
advanced gastric cancer remains poor due to the recurrence,
invasion and metastasis, with a 5-year survival rate of <30%. An
improved knowledge of the mechanisms underlying tumor metastasis is
warranted to discover novel paradigms for the diagnosis and
treatment of gastric cancer. Cancer metastasis is a highly complex
process that occurs through multiple steps, which include cell
invasion, cell migration, intravasation, transport through the
circulatory system, arrest at a secondary site, extravasation and
growth in a secondary organ (2).
Numerous metastasis-promoting genes are embedded in the primary
tumors and the ability to metastasize may be an inherent quality of
the tumor from the start. Additionally, multiple genes have been
reported to be involved in the metastasis of gastric cancer.
Matrix metalloproteinases (MMPs) have been regarded
as major critical molecules that assist tumor cells during
metastasis. MMPs, also designated matrixins, are proteinases that
participate in extracellular matrix (ECM) degradation. They
selectively cleave polypeptide bonds in ECM and remodel structural
proteins that are essential for the maintenance of connective
tissue integrity, such as collagens, aggrecan, fibronectin,
proteoglycan and laminin. To date, 24 different vertebrate MMPs
have been identified, 23 of which are found in humans (3). Among the numerous MMPs that have been
identified, MMP-2 encodes an enzyme which degrades type IV
collagen, the major structural component of basement membranes.
MMP-2 overexpression was found in a large proportion (94%) of the
gastric cancer tissues compared with the matched non-cancerous
tissues (4). An increase in MMP-2
levels and the presence of the active type of MMP-2 were closely
associated with the ability of invasion and metastasis of gastric
cancer (5).
Cluster of differentiation 147 (CD147), a 55-kDa
transmembrane glycoprotein, is located on the surface of human
tumor cells and normal keratinocytes (6). CD147 has been implicated in tumor
invasion and its elevated levels in cancer tissues have been
correlated with tumor progression in numerous malignant tumor
models (7). For example,
overexpression of CD147 has been demonstrated to enhance the
metastatic potential in human hepatoma cells (8). The silencing of CD147 expression in a
murine B16 melanoma model resulted in a reduced capability of the
tumor cells to metastasize to the draining lymph nodes (9). Gastric cancer tissue with higher CD147
expression also displayed an increased ability to invade into
lymphatic or venous vessels, or through the gastric wall (10). Downregulation of CD147 by RNAi led
to decreased cell proliferation, and invasive potential of SGC7901
cells (11). Therefore, control of
CD147 expression has considerable significance for regulation of
the metastatic capacity of gastric cancer.
Chitooligosaccharide (COS) is a natural alkaline
polysaccharose, an oligosaccharide formed by 2–10 amino-glucoses
through 1,4-glucosidic bond connection (12). The water-soluble COS possess various
biological activities, such as antitumor activity, antimicrobial
activity and antimutagenic activity (13). However, to date, no study has
reported the anti-metastatic effect of COS and its underlying
mechanism in human gastric cancer. In the present study, several
different gastric cancer cell lines were tested first for their
sensitivity to growth inhibition by COS. It was found that SGC-7901
was the most sensitive cell line among the tested cancer cell
lines. Then, the molecular mechanisms by which COS inhibited
SGC-7901 cell proliferation and metastasis were investigated.
Results presented here suggested that CD147/MMP-2 pathway played an
important role in the treatment of COS. We propose that COS has the
potential to be a novel chemotherapeutic agent for gastric
cancer.
Materials and methods
Materials
COS (1 kDa<MW<3 kDa) was obtained from the
Dalian Institute of Chemical Physics, the Chinese Academy of
Sciences (Dalian, China).
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
was purchased from Sigma-Aldrich (St. Louis, MO, USA) and Transwell
chambers were purchased from Corning Inc. (Corning, NY, USA). All
reagents used for cell culture were obtained from Gibco BRL (Grand
Island, NY, USA). Monoclonal goat anti-human CD147, monoclonal
rabbit anti-human MMP-2 and monoclonal mouse anti-human GADPH
antibodies were purchased from Santa Cruz Biotechnology, Inc.
(Santa Cruz, CA, USA).
Cell culture
The SGC-7901 human gastric cancer cell line was
obtained from the Type Culture Collection of Chinese Academy of
Sciences (Shanghai, China). AGS and NCI-N87 human gastric cancer
cells were purchased from American Type Culture Collection
(Rockville, MD, USA). All the cells were grown in RPMI-1640
(Invitrogen Life Technologies, Carlsbad, CA, USA) supplemented with
10% fetal calf serum at 37°C in a 5% CO2 incubator.
Plasmid construction and
transfection
For CD147 overexpression, CD147 was constructed into
a pEGFP-C1 eukaryotic expression vector (Clontech, Heidelberg,
Germany). Transfection was carried out using Lipofectamine 2000
(Invitrogen Life Technologies), according to the manufacturer’s
instructions. The cells were selected in the presence of 500 mg/l
G418 (Invitrogen Life Technologies). Stable clones were selected
for at least 4 weeks before single colonies were picked and
analyzed for CD147 expression by western blotting. The empty vector
pEGFP-C1 was also transfected into SGC7901 cells, which served as
the control group.
MTT assay
Cells were seeded into 96-well plates at a density
of 1×105 cells/100 μl/well. After 24 h growth, cells
were treated with different concentrations of COS (50, 100, 250,
500 and 1,000 μg/ml) for 24, 48 and 72 h. Following the addition of
100 μl MTT (0.5 mg/ml; Sigma-Aldrich), cells were incubated at 37°C
for 1 h. The formazan deposits that formed were solubilized in
DMSO, and the absorbance of each well was measured at 570 nm in an
EMax Precision microplate reader (Molecular Devices Instruments,
Sunnyvale, CA, USA).
Wound-healing assay
Cells were cultured in a 24-well plate with 100%
confluency. A micro-pipette tip was used to scratch a line in the
cell monolayer. The medium was removed and the monolayer was washed
with warm phosphate-buffered saline three times. Then growth medium
containing different concentrations of COS was added to each well.
Following incubation for 48 h, cell migration was observed and
photographs were taken under a light microscope (Olympus BX41;
Olympus Corporation, Tokyo, Japan).
Transwell assay
Cell migration through Matrigel-coated filters was
measured by using Transwell chambers (Costar Corporation,
Tewksbury, MA, USA) with 8-μm-pore polycarbonate filters coated
with Matrigel matrix. SGC-7901 cells were seeded in the upper
compartment of each invasion chamber and incubated in the presence
of COS for 48 h. The lower well was filled to the top (500 μl) with
RPMI-1640 containing 10% fetal calf serum (Hangzhou Sijiqing
Bioengineering Material Co., Ltd., Hangzhou, China) as a
chemoattractant. Subsequently, non-migrating cells on the upper
surface of the membrane were removed by gently scrubbing with a
cotton swab, and the invading cells on the lower surface were fixed
with 100% methanol and stained with crystal violet (0.1%; Beyotime
Institute of Biotechnology, Nantong, China). The number of cells
was counted under a light microscope (Olympus BX41; Olympus
Corporation) at a magnification of ×100. Images were captured and
subjected to computer-assisted image analysis using a computer
coupled to the Olympus BX41 light microscope (Olympus Corporation)
using AnalySis software (Olympus Corporation).
Quantitative real-time polymerase chain
reaction (qPCR)
Total RNA of cells with or without COS intervention
was extracted using TRIzol reagent (Invitrogen Life Technologies).
Subsequently, 2 μg of total RNA was reverse transcribed in a total
volume of 20 μl containing 200 units of SuperScript II RNase H-
Reverse Transcriptase (Invitrogen Life Technologies), 50 mM
Tris-HCl (pH 8.3), 75 mM KCl, 3 mM MgCl2, 10 mM
dithiothreitol, 500 μM dNTPs (each), 500 ng oligo (dT) 23 primer
and 40 units of RNaseOUT, at 42°C for 50 min. This was then
followed by inactivating at 70°C for 15 min. qPCR was conducted by
LightCycler 480 SYBR Green I Master (Roche Diagnostics,
Indianapolis, IN, USA) with 1 mM primers. The expression of GADPH
was used as endogenous control for the normalization of gene
expression. All reactions were performed in duplicate. The forward
and reverse primers were 5′-CCCCAAAACGGACAAAGAC-3′ and
5′-CTTCAGCACAAACAGGTTGC-3′, respectively, for human MMP-2;
5′-CCATGCTGGTCTGCAAGTCAG-3′ and 5′-CCGTTCATGAGGGCCTTGTC-3′,
respectively, for human CD147; and 5′-CCAACCGCGAGAAGATGA-3′ and
5′-CCAGAGGCGTACAGGGATAG-3′, respectively, for human GADPH.
Western blot analysis
Whole cell lysates from cultured cells were
harvested with cell lysis buffer. Equal amounts of protein (20 μg)
were separated by SDS-PAGE produced in-house and transferred to a
polyvinylidene difluoride membrane (Millipore, Bedford, MA, USA).
The membrane was blocked with 5% skimmed milk in Tris-buffered
saline and then incubated with primary antibodies for 1 h at room
temperature. Horseradish peroxidase-conjugated monoclonal goat
anti-rabbit, goat anti-mouse and donkey anti-goat secondary
antibodies and an enhanced chemiluminescence kit (all Beyotime
Institute of Biotechnology) were used for detection.
Gelatin zymography
Gelatin zymography was performed to determine the
activity of MMP-2. Briefly, protein in medium was then separated in
10% SDS-PAGE gel containing 1 mg/ml gelatin (Sigma-Aldrich) at 4°C.
After running, the gel was incubated in 2.5% Triton X-100 (Beyotime
Institute of Biotechnology) in deionized water for renaturing with
gentle agitation for 30 min at room temperature. Subsequently, the
gel was incubated in developing buffer (50 mM Tris-HCl, 0.2 M NaCl,
5 mM CaCl2 and 0.02% Brij35) overnight with gentle
shaking. The gel was stained with Coomassie blue R-250 (Beyotime
Institute of Biotechnology) for 30 min and then washed.
Gelatinolytic bands were observed as clear zones against the blue
background and the intensity of the bands was evaluated using
ImageMaster software (Amersham Pharmacia Bioscience, NJ, USA).
Statistical analyses
Data are expressed as the mean values ± standard
deviation from at least three experiments. Statistical comparisons
were based on Student’s t-test or analysis of variance. P<0.05
was considered to indicate a statistically significant
difference.
Results
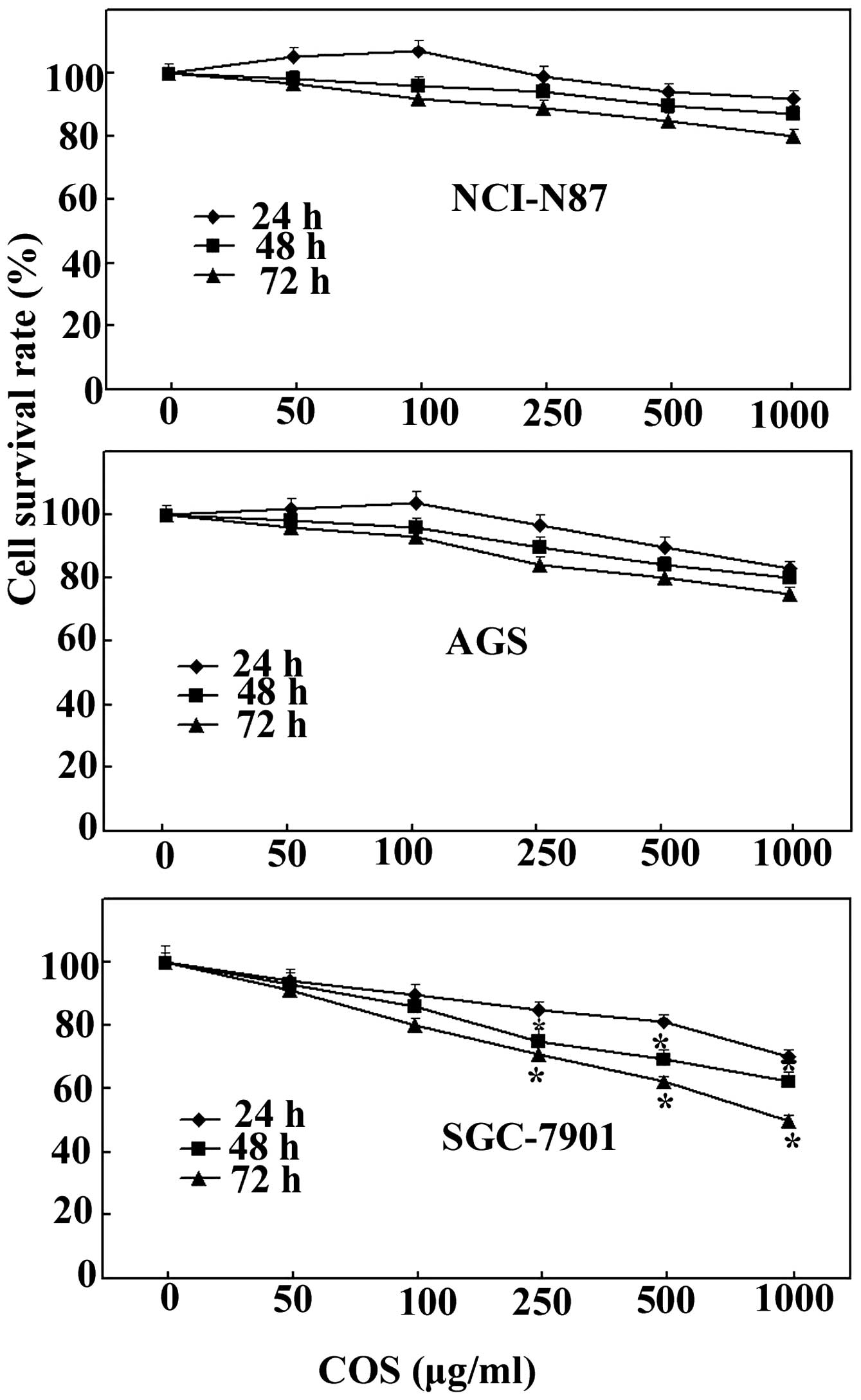
Effects of COS on cell proliferation in
AGS, SGC-7901 and NCI-N87 cells
To investigate the antiproliferative effect of COS
on various human gastric cancer cells, an MTT assay was carried out
following treatment with different concentrations of COS ranging
from 50 to 1000 μg/ml. As shown in Fig.
1, COS treatment marginally inhibited cell growth in AGS and
NCI-N87 cells, but significantly inhibited cell growth in SGC-7901
cells. COS inhibition of SGC-7901 cell growth was dose and
time-dependent; however, the effect became significant only after
≥48 h of treatment. Notably, COS concentrations >250 μg/ml
exhibited a marked inhibitory effect on cell proliferation compared
with low doses of COS (50 and 100 μg/ml). Therefore, SGC-7901 cells
were treated with COS at concentrations of 250, 500 and 1,000 μg/ml
for 48 h in the following experiments.
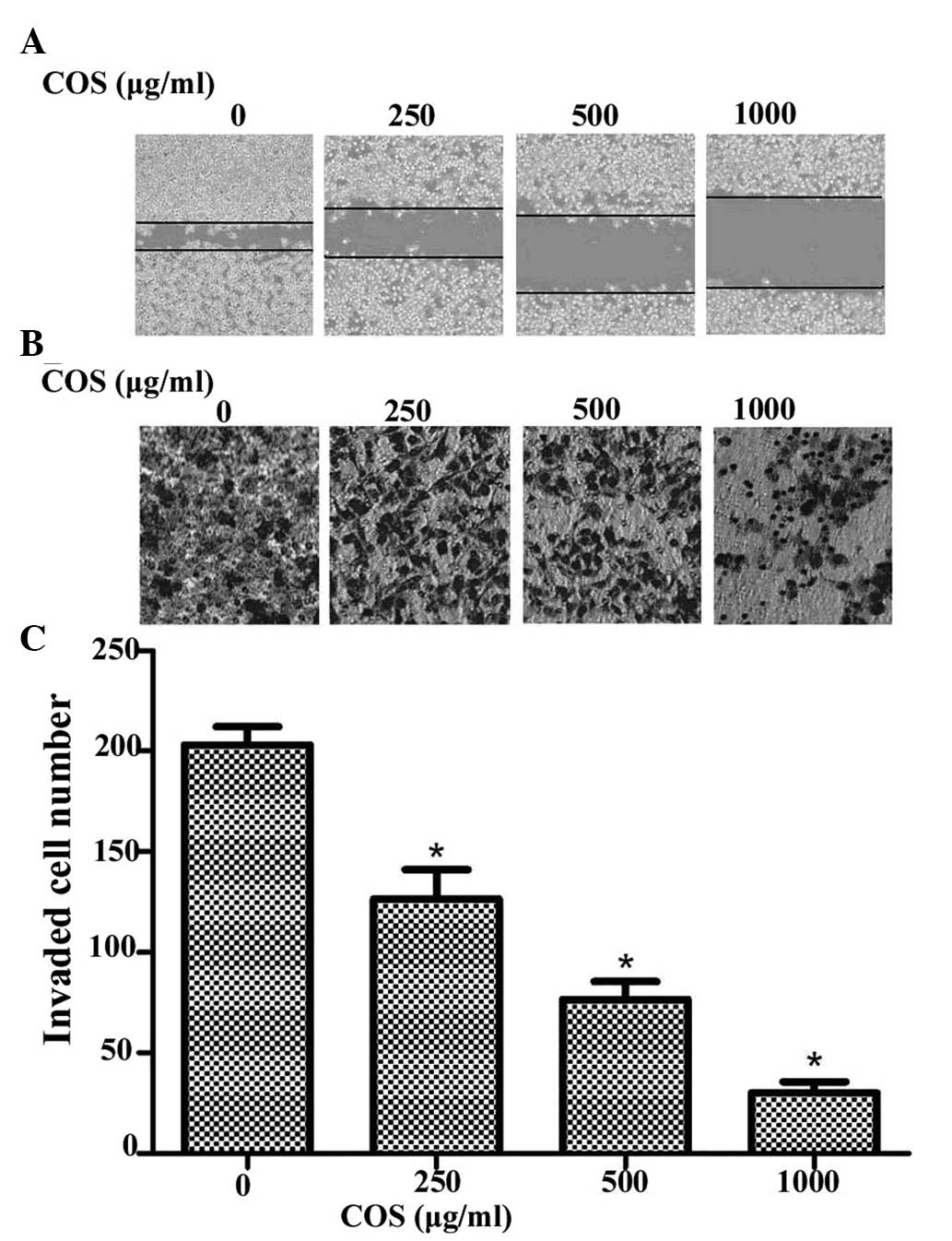
Effects of COS on cell migration and
invasion
In order to further assess the influence of COS on
SGC-7901 cells, cell migration and invasion assays were employed to
determine these two key factors of malignant progression and
metastasis. The wound healing assay showed that treatment of
SGC-7901 cells with increasing concentrations of COS after 48 h led
to a concentration-dependent decrease in wound-healing cell
migration (Fig. 2A). COS also
caused a dose-dependent decrease in the invasion of SGC-7901 cells
through the Matrigel chamber after 48 h (Fig. 2B). The number of invasive cells was
significantly decreased following COS treatment in a dose-dependent
manner (P<0.05, compared with the 0 μg/ml COS treatment group)
(Fig. 2C). These results indicate
that COS is a potential inhibitor of metastasis of gastric cancer
cells.
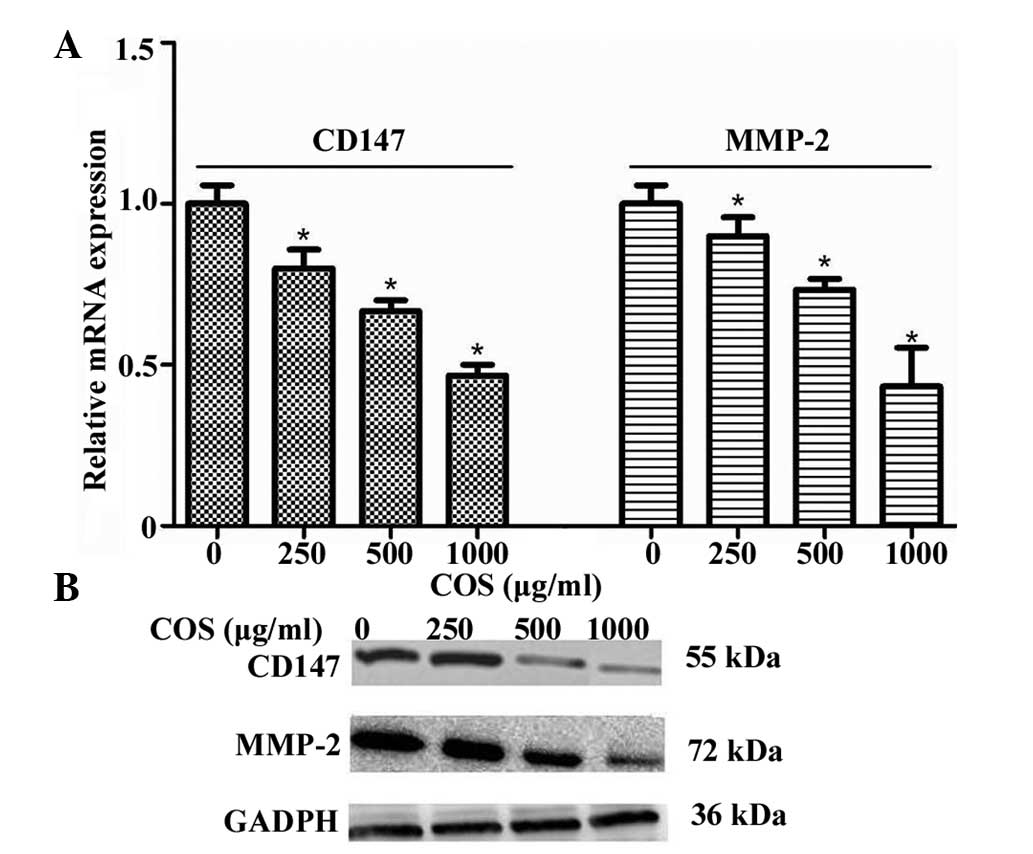
Effects of COS on CD147 and MMP-2
expression
Based on the abovementioned results, further studies
were carried out to examine the inhibitory effect of COS on CD147
mRNA and protein expression in SGC-7901 cells. Total RNA of cells
was isolated and qPCR was performed as described in Materials and
methods. As shown in Fig. 3A, the
levels of CD147 mRNA were significantly and dose-dependently
suppressed by COS. Pretreatment of SGC-7901 cells with COS also
decreased the protein expression of CD147 in a
concentration-dependent manner, which was detected by western blot
analysis (Fig. 3B). Furthermore,
the change of MMP-2 at the levels of mRNA and protein expression
displayed a similar trend to that of CD147 (Fig. 3A and B). In particular, 1,000 μg/ml
COS exhibited the greatest inhibitory effect on CD147 and MMP-2
expression.
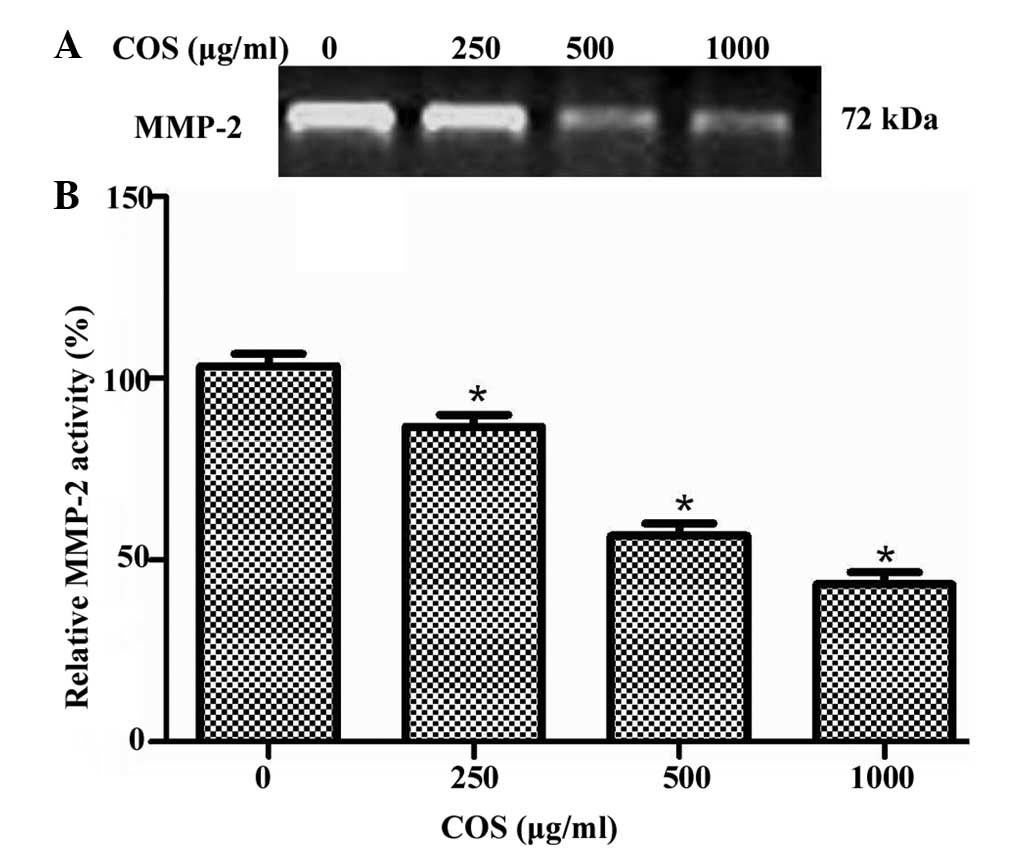
Effects of COS on MMP-2 activity
Increased MMP-2 activity is considered to be
important for the increased capability of gastric cancer cells to
traverse the membrane and invade and metastasize to distant sites
(14). Therefore, the present study
analyzed the effect of COS on the activity of MMP-2 cells. SGC-7901
cells were treated with different concentrations of COS (0, 250,
500 and 1,000 μg/ml), and the activity level of MMP-2 was
determined by gelatin zymography assay. As shown in Fig. 4A, the inhibitory effect on enzymatic
activity of MMP-2 was increased with increasing concentrations of
COS. Quantification analysis indicated that the MMP-2 activity
reduced by 17.6, 37.5 and 52.3% when cells were treated with 250,
500 and 1,000 μg/ml of COS, respectively (Fig. 4B). Notably, 1,000 μg/ml COS
exhibited the greatest inhibitory effect on MMP-2 activity.
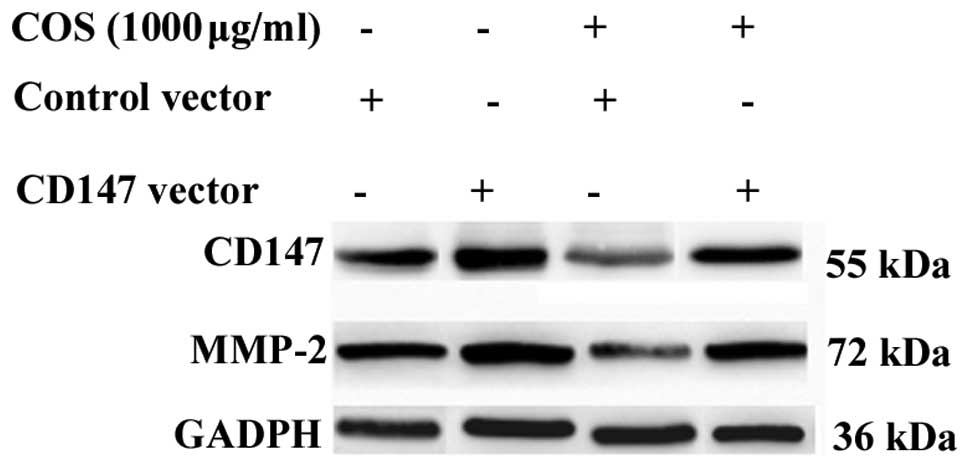
CD147 overexpression can attenuate the
effect of COS on MMP-2 inhibition
To further elucidate the mechanism by which COS
mediates MMP-2 inhibition through CD147, exogenous CD147 was
introduced into SGC-7901 cells. As shown in Fig. 5, SGC-7901 cells were treated for 48
h with COS at a concentration of 1000 μg/ml and then analyzed for
the indicated protein by western blotting. CD147 overexpression
activated the expression of MMP-2, and COS-induced MMP-2 inhibition
was reduced by CD147 overexpression. The results indicate that the
CD147/MMP-2 pathway is likely to be an important target of COS in
SGC-7901 cells.
Discussion
Chitosan, which is a linear polysaccharide composed
of randomly distributed β-(1–4)-linked
D-glucosamine (deacetylated unit) and N-acetyl-D-glucosamine
(acetylated unit), has been widely used in the food,
pharmaceutical, agricultural and environmental industries. In
recent years, chitosan has attracted interest when converted to
COS, as COS is not only water-soluble and of low molecular weight
but has also been demonstrated to exhibit various biological
functions (13,15,16).
The majority of these studies suggested that COS exhibited
antimetastatic activity in vivo and in vitro
(15,16). However, the underlying mechanisms
and the direct influence of COS on gastric cancer cells have not
been fully tested in detail. In the present study, we demonstrated
that COS treatment marginally inhibited cell growth in AGS and
NCI-N87 cells, but significantly inhibited cell growth in SGC-7901
cells. A similar result has been reported by Karagozlu et al
(17); however, the detailed
mechanisms remain unclear. In the current study, it was found that
SGC-7901 was the most sensitive cell line among the tested cancer
cell lines. The wound-healing and Transwell assays further
confirmed that COS could inhibit the metastatic process of SGC-7901
cells.
Expression of various MMPs has been found to be
upregulated in virtually every type of human cancer and correlates
with advanced stage, invasive and metastatic properties and, in
general, poor prognosis. Further upregulation of MMP expression, in
particular the gelatinases, which can degrade basement membrane
components, allows the tumor cells to invade into the adjacent
stroma and to break down the basement membranes associated with
capillaries and lymphatic vessels, allowing the tumor cells to
enter the circulation (18).
Therefore, compounds that have the potential to regulate MMPs are
considered to be attractive targets for therapeutic intervention.
Numerous studies have confirmed that the expression and activity of
MMPs could be mediated by COS. For example, Kim and Kim found that
COS suppressed the protein expression of MMP-2, and this inhibition
was caused by a decrease in the gene expression and transcriptional
activity of MMP-2 (19). MMP-9
inhibition in the presence of COS has been clearly observed in
HT1080 cells among tested molecular mass fractions (20). Furthermore, the inhibitory effect of
MMP-9 observed in HUVEC cells (21)
confirms that COS exerts its effect regardless of cell type. In the
present study, the expression of MMP-2 mRNA and protein, as
measured by qPCR and western blotting, was downregulated by COS in
SGC-7901 cells at concentrations of 250, 500 and 1,000 μg/ml
(P<0.05). It was also observed that COS caused a decline in the
enzymatic activity of MMP-2. These data demonstrate that COS can
significantly repress the invasion and migration ability of gastric
cancer cells in a dose-dependent manner, and this repression
strongly correlates with the inhibition of MMP-2.
As a tumor-associated antigen, CD147 forms
homo-oligomers in both heterotypic and homotypic cell-cell
interactions to induce production of MMPs. The functional
importance of CD147 has been demonstrated to be associated with its
ability to stimulate MMP expression. CD147 can induce the
production of MMP-1, MMP-2, MMP-3, MMP-9, MMP-14, and MMP-15
(22). Supporting its key role in
the processes of tumorigenesis and metastasis, CD147 has been
reported to be one of the most constantly upregulated mRNAs in
metastatic cells (23).
Downregulation of CD147 expression by RNA interference has been
demonstrated to inhibit MMP-2 expression and suppress cell
proliferation, invasion and tumorigenicity in vitro and
in vivo (24). In SGC-7901
gastric cancer cells, silencing the CD147 gene was found to
significantly decrease the proliferation and invasion of cells, and
downregulate the activity of MMP-2 (25). These studies support a model in
which CD147 in tumor cells stimulates MMP-2 production, thereby
leading to ECM degradation and increased tumor growth and
metastasis. To better understand how the COS regulated MMP-2, the
present study analyzed the change in CD147 expression levels
following treatment with different concentrations of COS ranging
from 250 to 1,000 μg/ml. The results revealed that the levels of
CD147 were significantly and dose-dependently suppressed by COS.
The change in MMP-2 expression levels displayed a similar trend to
that of CD147. These results therefore suggested that control of
CD147 expression by COS is involves in determining MMP-2 expression
in cancer metastasis.
As the regulation of gene expression of MMP-2 is
controlled by CD147, the current study next investigated whether
overexpression of CD147 could attenuate the effect of COS on MMP-2
inhibition. It was found that this effect could be effectively
attenuated by CD147 overexpression, suggesting that the
downregulating role of COS in MMP2 expression may be through the
upregulation of CD147. Further characterization of the effect of
COS on gastric cancer invasion and metastasis may lead to the
identification of new diagnostic markers and therapeutic targets.
We propose that these results may apply to a number of additional
cancer types other than gastric cancer, as CD147 and MMP-2 are
frequently upregulated in numerous other cancer types as well.
In conclusion, to the best of our knowledge, the
present study is the first to demonstrate that COS exhibits an
anti-metastatic effect in human gastric cancer. The effects of COS
on cell invasion and migration ability may be achieved through the
inhibition of MMP-2 expression by decreasing CD147. Overexpression
of CD147 partially protected against COS-mediated inhibition of
MMP-2. Overall, our findings have demonstrated the potential of COS
in suppressing gastric cancer metastasis and that the CD147/MMP-2
pathway may be involved as the key mechanism of its anti-metastatic
effect.
Acknowledgements
This study was supported by grants from the Young
Talent Project of Hubei Provincial Education Department
(Q200724004), Outstanding Youth Scientific Innovation Team of Hubei
University of Medicine (2011CXG03), Scientific Research Foundation
of Hubei University of Medicine (nos. 2010QDJ20 and 2010QDJ21).
References
|
1
|
Zheng L, Qi T, Yang D, et al: microRNA-9
suppresses the proliferation, invasion and metastasis of gastric
cancer cells through targeting cyclin D1 and Ets1. PLoS One.
8:e557192013.
|
|
2
|
Mehlen P and Puisieux A: Metastasis: a
question of life or death. Nat Rev Cancer. 6:449–458. 2006.
|
|
3
|
Nagase H, Visse R and Murphy G: Structure
and function of matrix metalloproteinases and TIMPs. Cardiovasc
Res. 69:562–573. 2006.
|
|
4
|
Murray G, Duncan M, Arbuckle E, Melvin W
and Fothergill J: Matrix metalloproteinases and their inhibitors in
gastric cancer. Gut. 43:791–797. 1998.
|
|
5
|
Partyka R, Gonciarz M, Jalowiecki P,
Kokocinska D and Byrczek T: VEGF and metalloproteinase 2 (MMP 2)
expression in gastric cancer tissue. Med Sci Monit. 18:BR130–BR134.
2012.
|
|
6
|
Zhou J, Zhu P, Jiang JL, et al:
Involvement of CD147 in overexpression of MMP-2 and MMP-9 and
enhancement of invasive potential of PMA-differentiated THP-1. BMC
Cell Biol. 6:252005.
|
|
7
|
Tang W, Chang SB and Hemler ME: Links
between CD147 function, glycosylation, and caveolin-1. Mol Biol
Cell. 15:4043–4050. 2004.
|
|
8
|
Jiang JL, Chan HC, Zhou Q, et al:
HAb18G/CD147-mediated calcium mobilization and hepatoma metastasis
require both C-terminal and N-terminal domains. Cell Mol Life Sci.
61:2083–2091. 2004.
|
|
9
|
Voigt H, Vetter-Kauczok CS, Schrama D,
Hofmann UB, Becker JC and Houben R: CD147 impacts angiogenesis and
metastasis formation. Cancer Invest. 27:329–333. 2009.
|
|
10
|
Zheng HC, Takahashi H, Murai Y, et al:
Upregulated EMMPRIN/CD147 might contribute to growth and
angiogenesis of gastric carcinoma: a good marker for local invasion
and prognosis. Br J Cancer. 95:1371–1378. 2006.
|
|
11
|
Wang B, Xu YF, He BS, et al: RNAi-mediated
silencing of CD147 inhibits tumor cell proliferation, invasion and
increases chemosensitivity to cisplatin in SGC7901 cells in vitro.
J Exp Clin Cancer Res. 29:612010.
|
|
12
|
Li J, He J and Yu C: Chitosan
oligosaccharide inhibits LPS-induced apoptosis of vascular
endothelial cells through the BKCa channel and the p38 signaling
pathway. Int J Mol Med. 30:157–164. 2012.
|
|
13
|
Lee JK, Lim HS and Kim JH: Cytotoxic
activity of aminoderivatized cationic chitosan derivatives. Bioorg
Med Chem Lett. 12:2949–2951. 2002.
|
|
14
|
Chen D, Wang Y, Zhang K, Jiao X, Yan B and
Liang J: Antisense oligonucleotide against clusterin regulates
human hepatocellular carcinoma invasion through transcriptional
regulation of matrix metalloproteinase-2 and e-cadherin. Int J Mol
Sci. 13:10594–10607. 2012.
|
|
15
|
Nam KS and Shon YH: Suppression of
metastasis of human breast cancer cells by chitosan
oligosaccharides. J Microbiol Biotechnol. 19:629–633. 2009.
|
|
16
|
Shen KT, Chen MH, Chan HY, Jeng JH and
Wang YJ: Inhibitory effects of chitooligosaccharides on tumor
growth and metastasis. Food Chem Toxicol. 47:1864–1871. 2009.
|
|
17
|
Karagozlu MZ, Kim JA, Karadeniz F, Kong CS
and Kim SK: Anti-proliferative effect of aminoderivatized
chitooligosaccharides on AGS human gastric cancer cells. Process
Biochem. 45:1523–1528. 2010.
|
|
18
|
Rundhaug JE: Matrix metalloproteinases,
angiogenesis, and cancer: commentary re: A. C. Lockhart et al.,
Reduction of wound angiogenesis in patients treated with
BMS-275291, a broad spectrum matrix metalloproteinase inhibitor
Clin Cancer Res, 9: 00-00, 2003. Clin Cancer Res. 9:551–554.
2003.
|
|
19
|
Kim MM and Kim SK: Chitooligosaccharides
inhibit activation and expression of matrix metalloproteinase-2 in
human dermal fibroblasts. FEBS Lett. 580:2661–2666. 2006.
|
|
20
|
Van Ta Q, Kim MM and Kim SK: Inhibitory
effect of chitooligosaccharides on matrix metalloproteinase-9 in
human fibrosarcoma cells (HT1080). Mar Biotechnol (NY). 8:593–599.
2006.
|
|
21
|
Rajapakse N, Kim MM, Mendis E, Huang R and
Kim SK: Carboxylated chitooligosaccharides (CCOS) inhibit MMP-9
expression in human fibrosarcoma cells via down-regulation of AP-1.
Biochim Biophys Acta. 1760:1780–1788. 2006.
|
|
22
|
Omi Y, Shibata N, Okamoto T, Obara T and
Kobayashi M: The role of CD147 in the invasiveness of follicular
thyroid carcinoma cells. Thyroid. 22:383–394. 2012.
|
|
23
|
Huang W, Luo WJ, Zhu P, et al: Modulation
of CD147-induced matrix metalloproteinase activity: role of CD147
N-glycosylation. Biochem J. 449:437–448. 2013.
|
|
24
|
Li R, Pan Y, He B, et al: Downregulation
of CD147 expression by RNA interference inhibits HT29 cell
proliferation, invasion and tumorigenicity in vitro and in vivo.
Int J Oncol. 43:1885–1894. 2013.
|
|
25
|
Chen L, Pan Y, Gu L, et al: ERK1/2
signalling pathway is involved in CD147-mediated gastric cancer
cell line SGC7901 proliferation and invasion. Exp Biol Med
(Maywood). 238:903–912. 2013.
|