Introduction
Osteosarcoma (OS) is the most frequent primary
malignant bone tumor in children and young adolescents (1,2). The
survival rates for OS have improved considerably since the
introduction of multi-agent chemotherapy in the 1980s, with a
5-year overall survival rate of 60–65% for patients without
metastasis (3), and have remained
relatively constant for the past two decades (4,5). The
development of conventional OS is a multistep process characterized
by genetic and epigenetic alterations that affect key cellular
pathways involved in cell growth and development (6). Therefore, an improved understanding of
the molecular mechanisms involved in OS initiation and progression
may contribute to the identification of a therapeutic target for
the treatment of OS, which may additionally improve the overall
outcome of patients with OS.
MicroRNAs (miRNAs) are a family of small non-coding
RNAs (19–25 nucleotides long), which are involved in the regulation
of gene expression by inducing messenger (m)RNA degradation or
repressing mRNA translation (7).
miRNAs have been demonstrated to be involved in numerous
physiological processes, including cell growth, apoptosis,
development and tumorigenesis (7–12). miRNAs
may act as oncogenes or tumor suppressors in the regulation of
carcinogenesis, metastasis and drug resistance (10), and are considered to be biomarkers for
the diagnosis of various types of cancer and potential novel
targets for anticancer therapies (13).
miRNA-497 (miR-497), an important member of the
miR-15/16/195/424/497 family of miRNAs (14), is downregulated in various types of
cancer, including ovarian (15),
breast (16), colorectal (17) and cervical cancer (18). Previous studies have demonstrated that
miR-497 exerts a tumor suppressor function in human colorectal
cancer (17), cervical cancer
(18), breast carcinoma (16), neuroblastoma (19), gastric cancer (20) and ovarian cancer (15,21). In
addition, miR-497 has been observed to be downregulated in OS cell
lines, compared with normal bone cells, using miRNA microarray
profiling in an integrative expression profiling approach (22). However, the exact role of miR-497 in
OS remains relatively unknown. Therefore, the aim of the current
study was to investigate the role of miR-497 in OS in vitro
and in vivo in order to determine the potential use of
miR-497 in OS therapy.
Materials and methods
Cell lines and cell culture
Human OS MG63 cells were obtained from the Shanghai
Institute of Biochemistry and Cell Biology of the Chinese Academy
of Sciences (Shanghai, China). The cells were cultured in RPMI-1640
medium (Sigma-Aldrich, St. Louis, MO, USA) supplemented with 10%
Gibco fetal calf serum (FCS; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA), 100 U/ml penicillin (Sigma-Aldrich) and 100
mg/ml streptomycin (Sigma-Aldrich), and maintained at 37°C in a
humidified atmosphere containing 5% CO2. The complete
medium was replaced every 2–3 days.
miRNA transfection
miR-497 mimic and negative control miRNA (Shanghai
GenePharma Co., Ltd., Shanghai, China) were transiently transfected
into MG63 cells in 6-well plates (Sigma-Aldrich) using 100 nM
Invitrogen Oligofectamine™ Transfection Reagent (Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from MG63 cells transfected
with miR-497 mimic or negative control miRNA, or from tumors of
mice inoculated with cells overexpressing miR-497 or negative
control cells, using Invitrogen TRIzol® reagent (Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. For
the detection of miR-497, RT-qPCR was performed using SYBR Green
Master Mix (Takara Biotechnology Co., Ltd., Dalian, China) and ABI
PRISM® 7500 Sequence Detection System (Thermo Fisher Scientific,
Inc.). Amplification of miR-497 and U6 was performed with 1 cycle
at 95°C for 3 min, followed by 30 cycles at 95°C for 10 sec and
60°C for 10 sec. The primers (Takara Biotechnology Co., Ltd.) used
were as follows: miR-497, forward 5′-GTGCAGGGTCCGAGGT-3′ and
reverse 5′-TAGCCTGCAGCACACTGTGGT-3′; and U6 (control), forward
5′-CTCGCTTCGGCAGCACATATACT-3′ and reverse
5′-ACGCTTCACGAATTTGCGTGTC-3′. U6 small nuclear RNA was used as a
normalization control. The relative expression of each gene was
calculated and normalized using the 2−∆∆Cq method
(Cq<35) (23).
Western blot analysis
The transfected cells were lysed 48 h subsequent to
transfection using RIPA Lysis and Extraction Buffer (Thermo Fisher
Scientific, Inc.), and the protein concentration was measured using
Pierce™ BCA Protein Assay Kit (Thermo Fisher Scientific, Inc.).
Following heating at 100°C for 10 min in the presence of a loading
buffer, equal amounts of protein lysates (50 µg) were separated
using 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(Bio-West Inc., Logan, UT, USA) at 100 V for 1 h, and transferred
onto Invitrogen nitrocellulose membranes (Thermo Fisher Scientific,
Inc.) at 120 V for 1 h. Following blocking with 5% skimmed milk
[diluted with phosphate-buffered saline (PBS); Sigma-Aldrich], the
membranes were incubated overnight at 4°C with the following
primary antibodies: i) Monoclonal rabbit anti-human matrix
metalloproteinase (MMP)-2 (catalog no., 13132; dilution, 1:2,000;
Cell Signaling Technology, Inc., Danvers, MA, USA); ii) monoclonal
mouse anti-human MMP-9 (catalog no., sc-12759; dilution, 1:1,000;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA); iii) monoclonal
mouse anti-human urokinase plasminogen activator (uPA; catalog no.,
sc-59729; dilution, 1:3,000; Santa Cruz Biotechnology, Inc.); and
iv) anti-human glyceraldehyde-3-phosphate dehydrogenase (GAPDH;
catalog no., sc-59729; dilution, 1:10,000; Santa Cruz
Biotechnology, Inc.). GAPDH was used as a loading control.
Subsequently, the membranes were incubated with secondary goat
anti-mouse horseradish peroxidase-conjugated immunoglobulin G
antibody (catalog no., sc-2005; dilution, 1:10,000; Santa Cruz
Biotechnology, Inc.) at room temperature for 2 h, and proteins were
detected using enhanced chemiluminescence (Pierce™ ECL Western
Blotting Substrate; Thermo Fisher Scientific, Inc.).
Proliferation analysis
In vitro cell proliferation was analyzed using Cell
Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan). Briefly, MG63 cells were transfected with a
miR-497 mimic or negative control miRNA in 6-well plates
(5×105 cells/well). Following 24 h, the cells were
trypsinized (Sigma-Aldrich) and re-seeded in 96-well plates
(Sigma-Aldrich) at a density of 1×103 cells/well. CCK-8
reagent (10 µl/well) was added at 0, 24, 48 and 72 h, and cells
were incubated for 2.5 h at 37°C. Optical density (OD) was measured
at 450 nm using a microplate reader (Multiskan Spectrum; Thermo
Labsystems, Helsinki, Finland). The experiment was performed ≥3
times, and similar results were obtained.
Colony formation assay
For the colony formation assay, 24 h subsequent to
transfection with miR-497 mimic or negative control miRNA, the
cells were seeded in a 6-well plate at a low cell density (1,000
cells/well), and maintained in RPMI-1640 medium containing 10% FCS
for 2 weeks. The colonies were fixed with methanol (Sigma-Aldrich),
and stained with 0.1% crystal violet (Sigma-Aldrich) diluted in 20%
methanol for 15 min. Colony numbers were quantified using AlphaView
software version 2.0 (ProteinSimple, San Jose, CA, USA), and the
percentage of colony formation was calculated by adjusting the
number of negative control cells to 100%.
Cell cycle and cell apoptosis
assay
The effects of miR-497 on the cell cycle and
apoptosis of OS cells were examined using flow cytometry. Briefly,
MG63 cells were transfected with a miR-497 mimic or negative
control miRNA for 48 h. Cell cycle progression was monitored using
Cell Cycle Assay Kit (Fluorometric - Green) (catalogue number,
ab112116; Abcam, Cambridge, MA, USA). The cells were acquired using
BD™ LSR II flow cytometer (BD Biosciences, San Jose, CA, USA), and
analyzed using Weasel version 3.1 software (available from
http://www.frankbattye.com.au/Weasel/WeaselDownload.html).
The percentage of cells undergoing apoptosis was determined with
Annexin V staining and propidium iodide exclusion using BD
Pharmingen™ FITC Annexin V Apoptosis Detection Kit I (BD
Biosciences). The cells were acquired using BD™ LSR II flow
cytometer, and analyzed using FACSDiva™ version 4.0 software (BD
Biosciences).
Furthermore, the activity of caspase-3 and caspase-9
was investigated as an additional indicator of apoptosis using
Caspase 3 Colorimetric Activity Assay Kit, DEVD and Caspase 9
Colorimetric Activity Assay Kit, LEHD, respectively (EMD Millipore,
Billerica, MA, USA), according to the manufacturer's protocol. The
relative caspase activity was calculated by adjusting the value
obtained for the negative control cells to 100%.
Transwell® migration and invasion
assays
For the Transwell® migration assays,
1×105 transfected cells were plated in the top chamber
of a Transwell® plate onto a non-coated membrane (24-well insert;
pore size, 8 µm; Corning Inc., Corning, NY, USA). For the invasion
assay, 1×105 transfected cells were plated in the top
chamber of a Transwell® plate onto a Matrigel™-coated membrane.
Each well was coated with 60 µg BD Matrigel™ Basement Membrane
Matrix (BD Biosciences) prior to the invasion assay. The cells were
incubated for 24 h (migration assay) or 48 h (invasion assay). The
cells that had migrated to the lower chamber of the Transwell®
plate were fixed in 70% ethanol (Sigma-Aldrich) for 30 min, and
stained with 0.2% crystal violet for 10 min. The number of cells
migrating or invading through the membrane were counted in five
randomly selected fields under a light microscope (BX51;
magnification, ×200; Olympus Corp., Tokyo, Japan).
Tumor growth in vivo
A total of 20 male BALB/c mice (5–6 weeks old) were
purchased from the Jilin Institute of Experimental Animals
(Changchun, China), and were maintained under specific
pathogen-free conditions at room temperature with a 12 h light/dark
cycle and humidity of 60–70%, and provided with food (Jilin
Institute of Experimental Animals) and water ad libitum.
Subsequently, 2×106 cells that were stably
overexpressing miR-497 or negative control miRNA were suspended in
100 µl PBS and injected into the flanks of the mice (overexpressing
miR-497 group, n=10; negative control group, n=10). The length and
width of the resulting tumors were measured every 7 days using a
digital caliper (Sigma-Aldrich). Tumor volume was calculated using
the following formula: Tumor volume (mm3) = 0.5236 ×
width2 × length. The mice were sacrificed 4 weeks
subsequent to inoculation, and the tumors were resected and
weighed. The tumors were used to measure the miR-497 expression
levels using the aforementioned RT-qPCR method. All animal
experiments were performed following the standards of animal care,
as outlined in the Guide for the Care and Use of Laboratory Animals
of Jilin University (Changchun, China), following a protocol
approved by the Ethics Committee of Jilin University.
Statistical analysis
The data were presented as the mean ± standard
deviation of ≥3 independent experiments. The differences between
the groups were analyzed using Student's t test when there were 2
groups, or one-way analysis of variance when there were ≥2 groups.
The analyses were performed using GraphPad Prism version 5 software
(GraphPad Software, Inc., La Jolla, CA, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
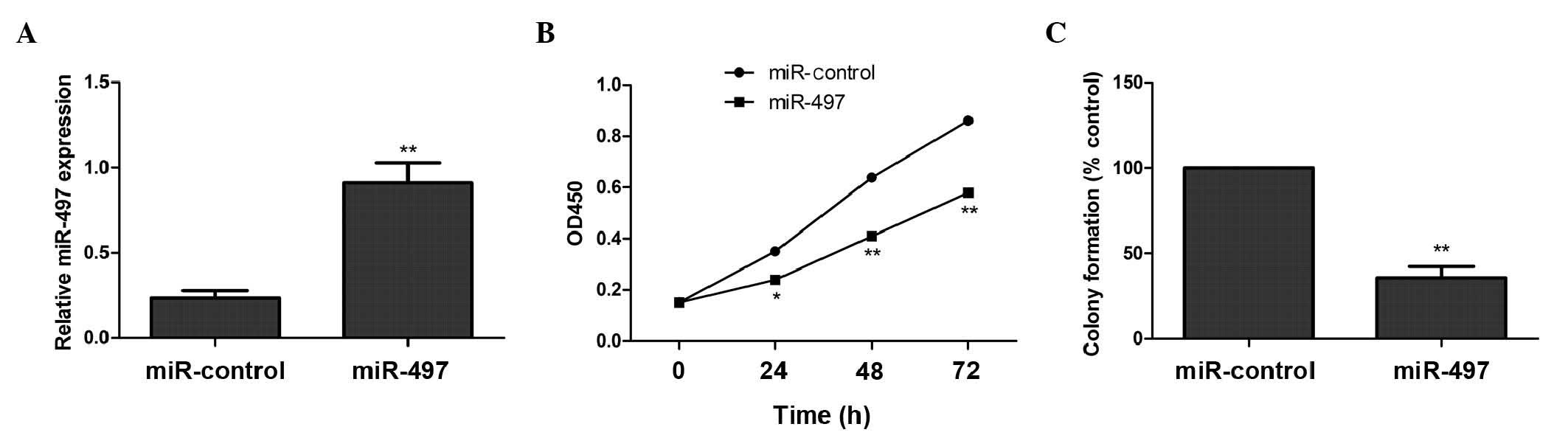
miR-497 inhibits cell proliferation
and colony formation in human OS MG63 cells
To examine the role of miR-497 in the growth of OS
cells, MG63 cells were transfected with a miR-497 mimic or a
negative control miRNA. Increased expression of miR-497 in cells
transfected with the miR-497 mimic was confirmed using RT-qPCR
(Fig. 1A). Overexpression of miR-497
significantly decreased the proliferation of cells, compared with
negative control cells (Fig. 1B). In
addition, miR-497 overexpression significantly inhibited colony
formation, compared with the corresponding negative control
(Fig. 1C). These results indicate
that miR-497 exhibits a growth-inhibitory role in OS cells.
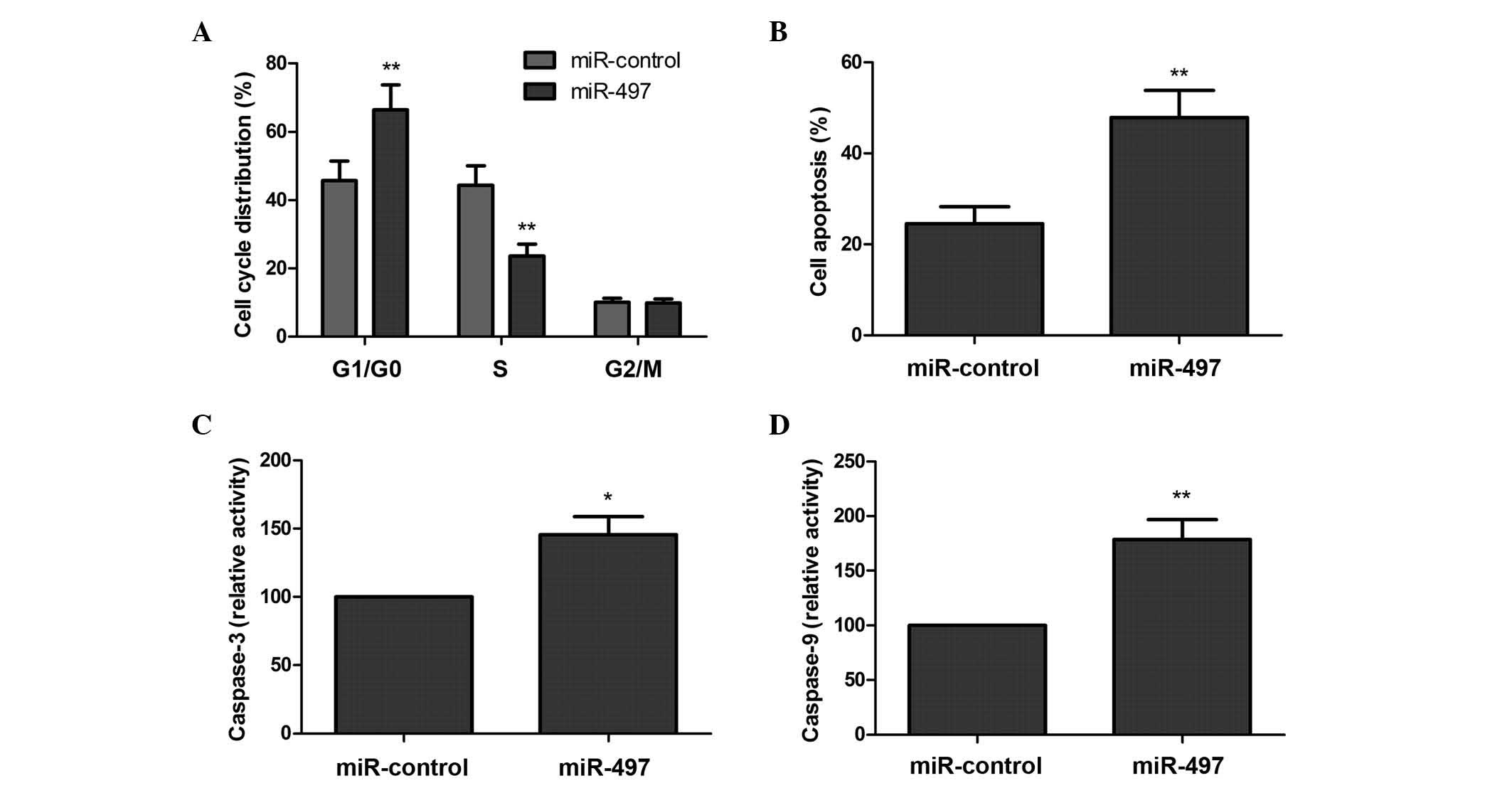
miR-497 induces G1 arrest and
apoptosis in MG63 cells
In order to determine the effects of miR-497 on the
cell cycle in MG63 cells, flow cytometry assays were performed. As
revealed by Fig. 2A, overexpression
of miR-497 increased the percentage of cells in the G1/G0 phase of
the cell cycle, and decreased the percentage of cells in the S
phase, compared with the negative control (P<0.05).
Subsequently, the role of miR-497 in the apoptosis
of MG63 cells was assessed. There was a significantly increased
percentage of apoptotic cells in the group that was overexpressing
miR-497, compared with the corresponding negative control group
(P<0.05; Fig. 2B). In addition,
the effects of miR-497 on caspase-3 and caspase-9 activity were
analyzed. As revealed by Fig. 2C and
D, caspase-3 and caspase-9 activity was significantly increased
in cells transfected with the miR-497 mimic, compared with cells
transfected with the negative control (P<0.05). These results
suggest that overexpression of miR-497 induces OS cells to arrest
at the G0/G1 phase of the cell cycle and undergo apoptosis.
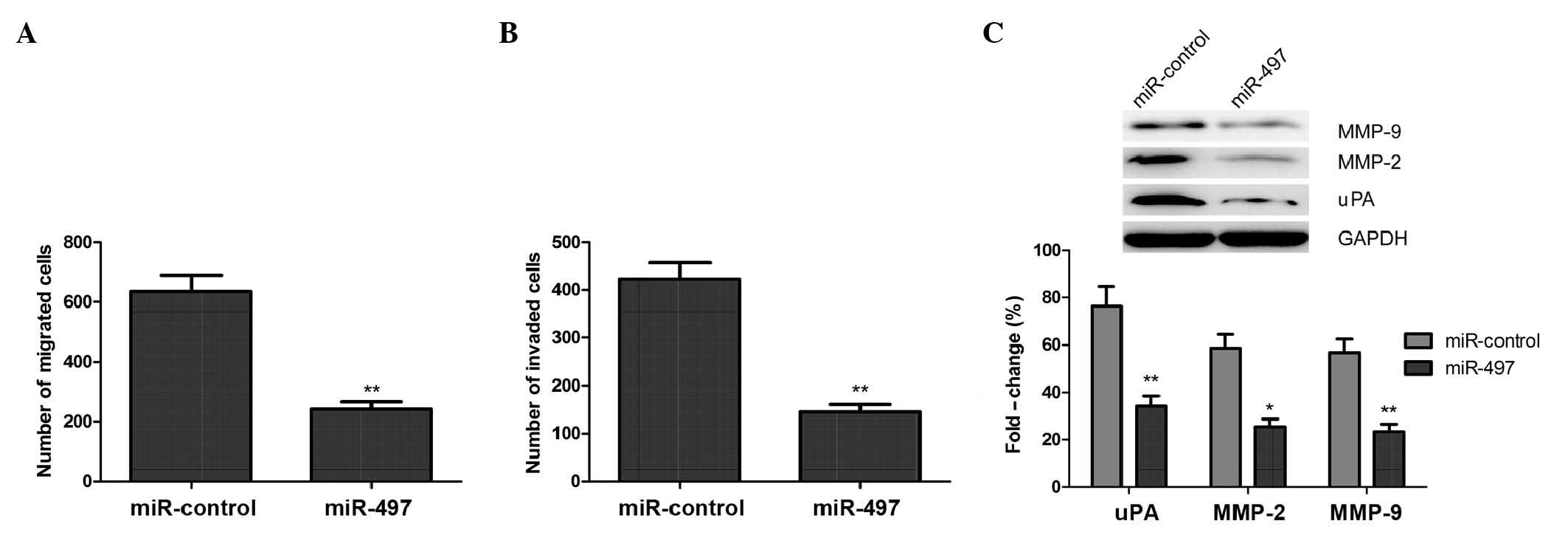
miR-497 inhibits cell migration and
invasion in MG63 cells
Since miR-497 may act as a tumor suppressor, the
present study investigated if restoration of miR-497 expression
would affect the migration and invasion abilities of OS cells. A
Transwell® assay without Matrigel™ demonstrated that the migratory
ability of MG63 cells was markedly decreased in cells transfected
with a miR-497 mimic, compared with cells transfected with negative
control miRNA (Fig. 3A).
Subsequently, a Transwell® assay with Matrigel™ demonstrated that
miR-497-transfected MG63 cells displayed significantly decreased
invasive abilities, compared with the negative control group
(Fig. 3B).
Furthermore, the present study analyzed the effects
of miR-497 on the expression of uPA, MMP-2 and MMP-9 using western
blot analysis. The results demonstrated that overexpression of
miR-497 resulted in a significant decrease in the protein
expression levels of uPA, MMP-2 and MMP-9, compared with cells
transfected with a negative control (P<0.05; Fig. 3C). These findings suggest that miR-497
inhibits the migratory and invasive abilities of OS cells in
vitro by inhibiting the expression of uPA, MMP-2 and MMP-9.
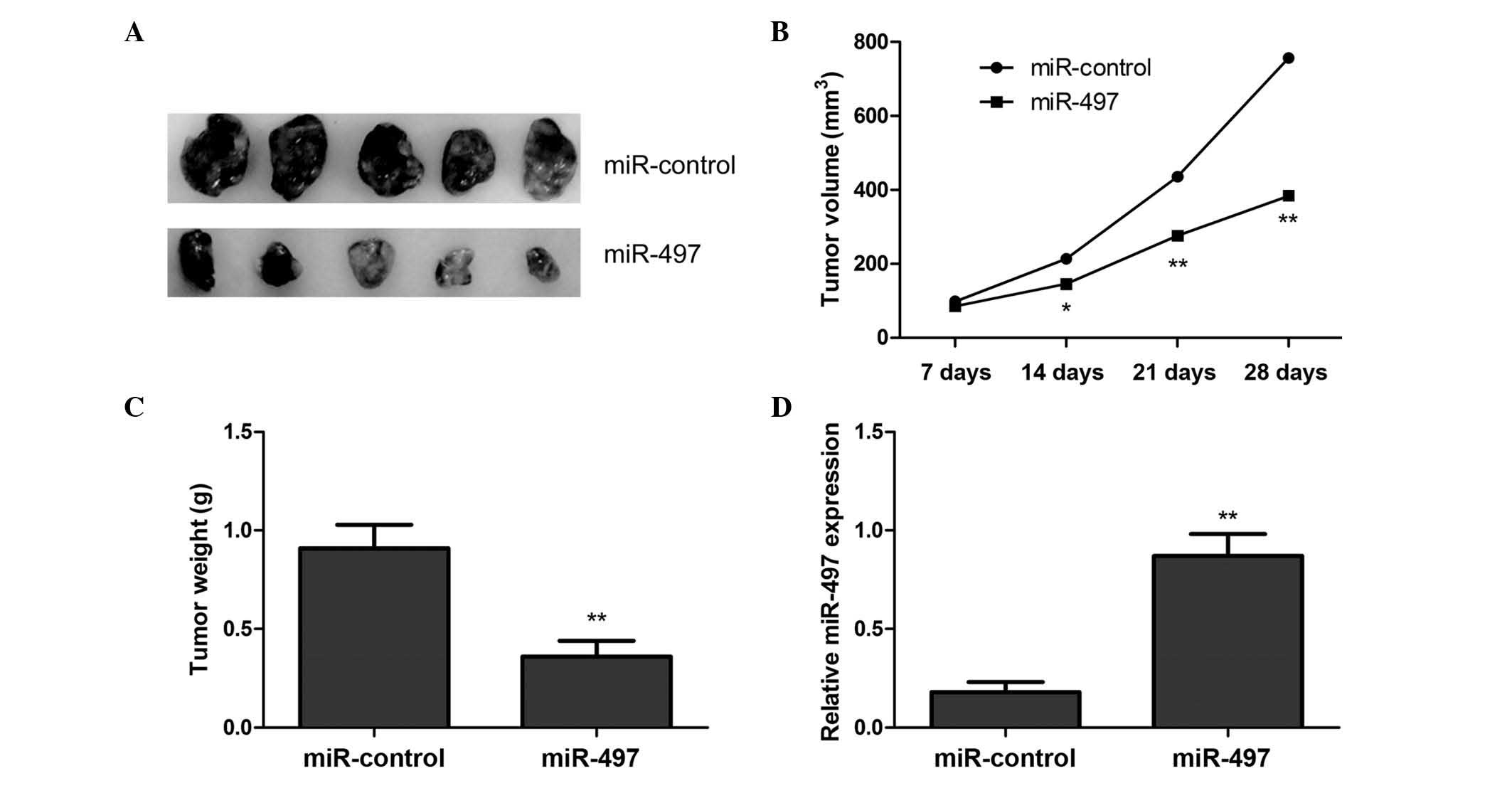
miR-497 inhibits tumor growth in
vivo
To examine the role of miR-497 in OS tumor
development, the present study used a BALB/c xenograft mouse model.
Mice were transplanted with MG63 cells overexpressing miR-497 or
negative control miRNA. The mice were sacrificed, and the tumor
tissues were excised 28 days subsequent to transplantation of the
transfected MG63 cells. The results demonstrated that
miR-497-overexpressing tumors were significantly smaller than
tumors of mice transfected with negative control miRNA (Fig. 4A). In addition, overexpression of
miR-497 significantly reduced xenograft tumor volume (Fig. 4B) and tumor weight (Fig. 4C), compared with the negative control
group. To determine miR-497 transfection efficiency in the nude
mouse model, the present study examined the expression levels of
miR-497 in the xenograft tumors using RT-qPCR. The results revealed
that miR-497 expression was upregulated in the xenograft tumors of
mice injected with cells transfected with the miR-497 mimic,
compared with tumors of mice injected with cells transfected with
negative control miRNA (Fig. 4D).
These results imply that overexpression of miR-497 may inhibit OS
tumor growth in vivo.
Discussion
Increased expression of miR-497 has been
demonstrated to suppress cancer cell proliferation, promote
apoptosis, decrease migration and invasion, and increase
chemosensitivity in cancer cells (16–22).
However, the roles and mechanisms of miR-497 in OS remain unclear.
To the best of our knowledge, the current study demonstrated for
the first time that overexpression of miR-497 inhibited the
proliferation, colony formation, migration and invasion abilities
of OS cells, and increased the percentage of apoptotic cells in
vitro. In addition, the present study demonstrated that
restoration of miR-497 expression inhibited OS tumor growth in a
nude mouse model. These results suggest that miR-497 inhibits tumor
growth in vitro and in vivo, as previously reported
(16–21).
Previous studies have revealed that tumor suppressor
miRNAs are involved in the carcinogenesis and tumor progression of
OS, and exert tumor suppressor effects in OS. Li et al
(24) reported that miRNA-145 acts as
a tumor suppressor by inhibiting OS cell growth through suppression
of rho-associated, coiled-coil-containing protein kinase 1. Sun
et al (25) reported that
upregulation of miRNA-646 expression inhibited OS cell
proliferation, migration and invasion by downregulating fibroblast
growth factor-2. In addition, Zhao et al (26) demonstrated that transfection of a
miRNA-410 expression plasmid into OS cells inhibited cell
proliferation and contributed to cell apoptosis by regulating
vascular endothelial growth factor (VEGF). Zhang et al
(27) revealed that miRNA-101 may act
as a tumor suppressor in OS, since it has a suppressive role in OS
cell migration and invasion by targeting enhancer of zeste homolog
2. miR-497, another potential tumor suppressor, has been
demonstrated to be downregulated in OS cells (21). The present study identified that
overexpression of miR-497 inhibited the tumor growth of OS cells
in vitro and in vivo, which suggests that miR-497 may
be a promising therapeutic target for the treatment of patients
with OS.
miRNAs are key in the regulation of cancer cell
proliferation, cell cycle distribution, migration and invasion by
regulating multiple target genes (28). Several targets have been identified
that are essential in cell cycle and survival pathways, including
insulin-like growth factor 1 receptor (17,18), WEE1
G2 checkpoint kinase (19), B-cell
lymphoma-2 (29), cyclin D2 (30) and VEGF (31). Previously, checkpoint kinase 1 and
hepatoma-derived growth factor were also identified as target genes
for miR-497 (32,33). Therefore, the present study
hypothesizes that miR-497 inhibits OS cell proliferation,
clonogenicity, migration and invasion, and induces OS cell
apoptosis and cell arrest at the G0/G1 phase of the cell cycle by
regulating multiple target genes.
In summary, the results of the present study
indicate that overexpression of miR-497 in OS cells is able to
inhibit OS cell proliferation, colony formation, migration and
invasion, and induce G1 phase arrest and apoptosis of OS cells
in vitro. In addition, the present study revealed that
overexpression of miR-497 suppresses OS tumor growth in a xenograft
mouse model. Therefore, these findings suggest that miR-497 may be
a potential molecular target for OS therapy.
References
|
1
|
Ottaviani G, Robert RS, Huh WW, Palla S
and Jaffe N: Sociooccupational and physical outcomes more than 20
years after the diagnosis of osteosarcoma in children and
adolescents: Limb salvage versus amputation. Cancer. 119:3727–3736.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ottaviani G and Jaffe N: The etiology of
osteosarcoma. Cancer Treat Res. 152:15–32. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
PosthumaDeBoer J, Witlox MA, Kaspers GJ
and van Royen BJ: Molecular alterations as target for therapy in
metastatic osteosarcoma: A review of literature. Clin Exp
Metastasis. 28:493–503. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chou AJ, Kleinerman ES, Krailo MD, Chen Z,
Betcher DL, Healey JH, Conrad EU III, Nieder ML, Weiner MA, Wells
RJ, et al: Children's Oncology Group: Addition of muramyl
tripeptide to chemotherapy for patients with newly diagnosed
metastatic osteosarcoma: A report from the Children's Oncology
Group. Cancer. 115:5339–5348. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Grignani G, Palmerini E, Dileo P, Asaftei
SD, D'Ambrosio L, Pignochino Y, Mercuri M, Picci P, Fagioli F,
Casali PG, et al: A phase II trial of sorafenib in relapsed and
unresectable high-grade osteosarcoma after failure of standard
multimodal therapy: An Italian Sarcoma Group study. Ann Oncol.
23:508–516. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Luetke A, Meyers PA, Lewis I and Juergens
H: Osteosarcoma treatment - where do we stand? A state of the art
review. Cancer Treat Rev. 40:523–532. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kato M and Slack FJ: MicroRNAs: Small
molecules with big roles - C. elegans to human cancer. Biol
Cell. 100:71–81. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kloosterman WP and Plasterk RH: The
diverse functions of microRNAs in animal development and disease.
Dev Cell. 11:441–450. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Iorio MV and Croce CM: MicroRNA
dysregulation in cancer: Diagnostics, monitoring and therapeutics.
A comprehensive review. EMBO Mol Med. 4:143–159. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nana-Sinkam SP and Croce CM: MicroRNA
dysregulation in cancer: Opportunities for the development of
microRNA-based drugs. IDrugs. 13:843–846. 2010.PubMed/NCBI
|
|
12
|
Suzuki H, Maruyama R, Yamamoto E and Kai
M: Epigenetic alteration and microRNA dysregulation in cancer.
Front Genet. 4:2582013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cho WC: OncomiRs: The discovery and
progress of microRNAs in cancers. Mol Cancer. 6:602007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Finnerty JR, Wang WX, Hébert SS, Wilfred
BR, Mao G and Nelson PT: The miR-15/107 group of microRNA genes:
Evolutionary biology, cellular functions, and roles in human
diseases. J Mol Biol. 402:491–509. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang W, Ren F, Wu Q, Jiang D, Li H, Peng
Z, Wang J and Shi H: MicroRNA-497 inhibition of ovarian cancer cell
migration and invasion through targeting of SMAD specific E3
ubiquitin protein ligase 1. Biochem Biophys Res Commun.
449:432–437. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li D, Zhao Y, Liu C, Chen X, Qi Y, Jiang
Y, Zou C, Zhang X, Liu S, Wang X, et al: Analysis of MiR-195 and
MiR-497 expression, regulation and role in breast cancer. Clin
Cancer Res. 17:1722–1730. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Luo M, Shen D, Zhou X, Chen X and Wang W:
MicroRNA-497 is a potential prognostic marker in human cervical
cancer and functions as a tumor suppressor by targeting the
insulin-like growth factor 1 receptor. Surgery. 153:836–847. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Guo ST, Jiang CC, Wang GP, Li YP, Wang CY,
Guo XY, Yang RH, Feng Y, Wang FH, Tseng HY, et al: MicroRNA-497
targets insulin-like growth factor 1 receptor and has a tumour
suppressive role in human colorectal cancer. Oncogene.
32:1910–1920. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Creevey L, Ryan J, Harvey H, Bray IM,
Meehan M, Khan AR and Stallings RL: MicroRNA-497 increases
apoptosis in MYCN amplified neuroblastoma cells by targeting the
key cell cycle regulator WEE1. Mol Cancer. 12:232013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li W, Jin X, Deng X, Zhang G, Zhang B and
Ma L: The putative tumor suppressor microRNA-497 modulates gastric
cancer cell proliferation and invasion by repressing eIF4E. Biochem
Biophys Res Commun. 449:235–240. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang W, Ren F, Wu Q, Jiang D, Li H and Shi
H: MicroRNA-497 suppresses angiogenesis by targeting vascular
endothelial growth factor A through the PI3K/AKT and MAPK/ERK
pathways in ovarian cancer. Oncol Rep. 32:2127–2133.
2014.PubMed/NCBI
|
|
22
|
Namløs HM, Meza-Zepeda LA, Barøy T,
Østensen IH, Kresse SH, Kuijjer ML, Serra M, Bürger H,
Cleton-Jansen AM and Myklebost O: Modulation of the osteosarcoma
expression phenotype by microRNAs. PLoS One. 7:e480862012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li E, Zhang J, Yuan T and Ma B: MiR-145
inhibits osteosarcoma cells proliferation and invasion by targeting
ROCK1. Tumour Biol. 35:7645–7650. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sun XH, Geng XL, Zhang J and Zhang C:
miRNA-646 suppresses osteosarcoma cell metastasis by downregulating
fibroblast growth factor 2 (FGF2). Tumour Biol. 36:2127–2134. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhao D, Jia P, Wang W and Zhang G:
VEGF-mediated suppression of cell proliferation and invasion by
miR-410 in osteosarcoma. Mol Cell Biochem. 400:87–95. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang K, Zhang Y, Ren K, Zhao G, Yan K and
Ma B: MicroRNA-101 inhibits the metastasis of osteosarcoma cells by
downregulation of EZH2 expression. Oncol Rep. 32:2143–2149.
2014.PubMed/NCBI
|
|
28
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhu W, Zhu D, Lu S, Wang T, Wang J, Jiang
B, Shu Y and Liu P: miR-497 modulates multidrug resistance of human
cancer cell lines by targeting BCL2. Med Oncol. 29:384–391. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yadav S, Pandey A, Shukla A, Talwelkar SS,
Kumar A, Pant AB and Parmar D: miR-497 and miR-302b regulate
ethanol-induced neuronal cell death through BCL2 protein and cyclin
D2. J Biol Chem. 286:37347–37357. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhou TB, Jiang ZP, Liu ZS and Zhao ZZ:
Roles of miR-497 and its potential signaling pathway in diseases
and with vascular endothelial growth factor. J Recept Signal
Transduct Res. 35:303–306. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xie Y, Wei RR, Huang GL, Zhang MY, Yuan YF
and Wang HY: Checkpoint kinase 1 is negatively regulated by miR-497
in hepatocellular carcinoma. Med Oncol. 31:8442014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhao WY, Wang Y, An ZJ, Shi CG, Zhu GA,
Wang B, Lu MY, Pan CK and Chen P: Downregulation of miR-497
promotes tumor growth and angiogenesis by targeting HDGF in
non-small cell lung cancer. Biochem Biophys Res Commun.
435:466–471. 2013. View Article : Google Scholar : PubMed/NCBI
|