Introduction
Cervical cancer is one of the most common
malignancies in women worldwide, and remains one of the leading
causes of cancer-associated mortalities globally (1). Epidemiological studies show that human
papillomavirus (HPV) infection, in particular HPV16, is relevant in
the onset of cervical cancer (2).
However, an increasing body of evidence has indicated that the
occurrence of cervical cancer is a result of the combined action of
multiple factors, since the incidence of HPV infection in women of
childbearing age is higher, but the incidence of cervical cancer is
lower among these patients (3).
Therefore, HPV infection is not the only factor involved in the
development of cervical cancer (4,5). Following
infection with high-risk HPV, cervical cancer develops through
precancerous lesions termed cervical intraepithelial neoplasia
(CIN), and eventually progresses to invasive cervical cancer. To
the best of our knowledge, the molecular mechanisms underlying the
progression of HPV infection to cervical cancer largely remain to
be elucidated.
Previous studies have demonstrated that the
deregulation of microRNAs (miRs) contributes to the process of
cervical carcinogenesis (6). miRs are
small non-coding RNA molecules that are able to bind to the
3′-untranslated region of their target messenger (m)RNA, thus
regulating the stability and translation of mRNAs, which results in
the inhibition of translation or degradation of the target mRNA
(7). Due to their ability to alter
the expression of protein-coding oncogenes and tumor suppressor
genes, miRs are proposed to have a significant role in cancer
development (8). Similar to various
growth-suppressive protein-encoding tumor suppressor genes, certain
miR genes harbor CpG islands, which are susceptible to
methylation-mediated silencing (9,10).
Therefore, hypermethylation-mediated silencing of tumor-suppressive
miRs may be an important mechanism of tumorigenesis (11).
Folate is a methyl donor during the methylation
cycle, which ensures the maintenance of adequate cellular levels of
S-adenosylmethionine for its use in biological methylation
reactions, including DNA methylation (12). Thus, folic acid deficiency may prevent
normal methylation of epigenetically regulated genes (13). A number of previous studies have
reported that folate levels are inversely associated with the risk
of developing cervical cancer through the hypermethylation of CpG
islands in certain tumor suppressor genes (14–16).
However, to the best of our knowledge, methylation-mediated
transcriptional repression of miRs regulated by folic acid has not
been previously investigated in cervical cancer. Investigation of
miR methylation provides a novel approach and insight for
evaluating new molecules and mechanisms of progression in cervical
cancer.
To evaluate the potential role of folate-associated
miR methylation during the progression of CIN to cervical cancer,
the present study assessed the expression of miR-203 and miR-375 in
a range of tissues obtained from normal, CIN and cervical cancer
cases. The population study and the experiments in cells
demonstrated that folic acid is involved in the methylation of the
CpG islands of miR-203 and miR-375. The results of the present
study contribute to the understanding of the potential mechanism of
cervical carcinogenesis induced by folate deficiency, which may
lead to novel strategies for the prevention of CIN progression to
cervical cancer.
Materials and methods
Patients and samples
Biopsy tissues were obtained from 90 women, with a
median age of 32 years (range, 22–67 years), who were subjected to
colposcopic examinations between January 2012 and January 2013 in
the Department of Obstetrics and Gynecology of The Second Hospital
of Shanxi Medical University (Taiyuan, China). According to the
results of the pathological diagnosis of the cervical biopsies
under colposcopy, patients in the study group were divided into CIN
or stage IA-IIA cervical cancer (30 cases each). Patients with
negative intraepithelial lesion or malignancy constituted the
control group (30 cases). According to the 2009 International
Federation of Gynecology and Obstetrics staging system (17), 15 cases of cervical squamous cell
carcinoma (SCC) were stage Ia, 6 cases were stage Ib and 9 cases
were stage IIa. According to the pathological classification
(18), 4 cases were
highly-differentiated, 24 cases were moderately-differentiated and
2 cases were poorly-differentiated cervical SCC. Among the patients
with CIN, 5 cases were stage I, 15 cases were stage II and 10 cases
were stage III. The staging of CIN were classified between CIN I
and CIN III, according to Wright et al (19). Normal cervical tissues from the same
period were obtained from the complete uterine hysterectomy
practices on the patients.
All participants were Han Chinese and had been
living in Shanxi for >5 years, and their diagnoses were
confirmed by two pathological examinations. Participants with
nutritional megaloblastic anemia, hemolytic disease, leukemia,
liver disease, other tumors or who had been administered B vitamins
within the 3 months prior to the study, were excluded. None of the
patients received radiotherapy or chemotherapy prior to surgery.
Upon obtaining written informed consent from all participants,
uterine cervix tissue samples were obtained from the participants
who underwent hysterectomy or biopsy under colposcopy, and
immediately stored at −80°C. The present study was conducted in
accordance with the Declaration of Helsinki, and was conducted with
approval from the Ethics Committee of The Second Hospital of Shanxi
Medical University (Taiyuan, China).
Cell culture
CaSki cervical cancer cells were purchased from the
Cell Bank of Type Culture Collection of the Chinese Academy of
Sciences (Shanghai, China), and preserved in the Department of
Obstetrics and Gynecology of The Second Hospital of Shanxi Medical
University. Cells were cultured in RPMI-1640 medium (Hyclone; GE
Healthcare Life Sciences, Logan, UT, USA) supplemented with 10%
(v/v) fetal bovine serum (Sijiqing; Zhejiang Tianhang Biotechnology
Co., Ltd.; Huzhou, China) in a humidified 5% CO2
incubator at 37°C. Folic acid (Sigma-Aldrich, St. Louis, MO, USA)
was used at final concentrations of 1, 10, 100, 250, 750 and 1,000
µg/ml. The cancer cell line was seeded (3×105
cells/well) into 6-well plates (Corning Incorporated, Corning, NY,
USA), and incubated overnight. Subsequently, cells were treated
with folic acid at the aforementioned concentrations for 48 h.
Following 48 h of incubation, cells were harvested and used in
subsequent experiments.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RNA was isolated from patients' tissues using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) according to the manufacturer's protocol,
and the quantity and concentration of RNA were
spectrophotometrically assessed (NanoDrop™ 2000; Thermo Fisher
Scientific, Inc.) by ethidium bromide visualization (Sigma-Aldrich)
of 28S and 18S ribosomal (r)RNA. The absorbance
(A)260/A280 ratio was determined to be in the
1.9–2.2 range, indicating no contamination (protein contamination,
<1.9; DNA contamination, >1.9; NanoDrop™ 2000; Thermo Fisher
Scientific, Inc.). For miR quantification, the Qiagen OneStep
RT-PCR kit (Qiagen GmbH, Hilden, Germany) was used. Each RT
reaction consisted of 2 µl total RNA, mixed with 1.2 µl primers, 10
µl 2X buffer, 0.2 µl Moloney murine leukemia virus reverse
transcriptase and 6.5 µl diethylpyrocarbonate-treated water in a
final volume of 20 µl, and was incubated at 42°C for 30 min, 85°C
for 10 min and maintained at 4°C. DNase (Sigma-Aldrich) was used
for degrading DNA. RT-qPCR was conducted in a StepOnePlus™
Real-Time PCR system (Thermo Fisher Scientific, Inc.) using SYBR
Premix Ex Taq II (Tli RNase H Plus) (Takara Biotechnology Co.,
Ltd., Dalian, China). PCR was performed using the EpiTect Fast DNA
Bisulfite kit (Qiagen GmbH). Each PCR reaction contained 20 µM
forward primer, 20 µM reverse primer, 10 µl PCR Master Mix, 0.4 µl
Taq DNA polymerase (2.5 U/µl), and double distilled
(dd)H2O to a final volume of 20 µl. The PCR conditions
were as follows: An initial cycle at 95°C for 3 min, followed by 40
cycles at 94°C for 12 sec and 40 sec at 62°C. Primers were designed
using the online bioinformatics tool MethPrimer (www.urogene.org/methprimer). The primers, whose
sequence appears in Table I, were
synthesized by Jima Corp. (Shanghai, China). The 2−ΔΔCq
method (20) was used to determine
the relative quantification of miR expression in the tissue
samples. The levels of miR expression were normalized to the
expression levels of 5S rRNA and represented as fold-change.
 | Table I.Primer sequences for amplification of
miR-375, miR-203 and 5S rRNA. |
Table I.
Primer sequences for amplification of
miR-375, miR-203 and 5S rRNA.
| Gene | Primer sequence,
5′-3′ | Product length,
bp |
|---|
| 5S rRNA | F:
ACGGCCATACCACCCTGAAC | 91 |
|
| R:
GGCGGTCTCCCATCCAAGTA |
|
| hsa-miR-375 | F:
CTTACTATCCGTTTGTTCGTTCG | 81 |
|
| R:
TATGGTTGTTCTCGTCTCTGTGTC |
|
| hsa-miR-203 | F:
AACCTTGCTCGTGAAATGTTTAG | 80 |
|
| R:
TATGCTTGTTCTCGTCTCTGTGTC |
|
Methylation-specific PCR (MSP)
MSP consists of two different PCRs that amplify a
bisulfite-treated DNA sample using primers that are specific for
methylated or unmethylated sequences (21). MSP was performed using the EpiTect MSP
Kit (Qiagen GmbH). The MSP reactions consisted of 0.35 µM primers,
25 µl EpiTect Master Mix, 15 µl target DNA and ddH2O to
a final volume of 50 µl. The primer sequences for MSP were as
follows: miR-375, methylated primers, sense,
5′-AGCGGCGTATAGTTTTTTTTATTC-3′; antisense,
5′-CGAACCTAAACGTTTTATTCGTT-3′; unmethylated primers, sense,
5′-CTTACTATCCGTTTGTTCGTTCG-3′; antisense,
5′-TATGGTTGTTCTCGTCTCTGTGTC-3′; miR-203, methylated primers, sense,
3′-TTGCGGAGAGAGGAGTTTTC-5′; antisense,
3′-CTACAACAAAACAAAAAATACGCG-5′; unmethylated primers, sense,
3′-GAGTTGTGGAGAGAGGAGTTTTT-5′; antisense,
3′-CTACAACAAAACAAAAAATACACAAC-5′. The reaction conditions were as
follows: An initial cycle of 15 min at 95°C, followed by 35 cycles
of 15 sec at 94°C, 30 sec at 55°C and 30 sec at 72°C. The amplicons
were evaluated on 8% agarose electrophoresis gel. The presence or
absence of one of the amplicons was associated with the pattern of
the target gene.
Statistical analysis
Statistical analysis was performed with SPSS version
16.0 (SPSS, Inc., Chicago, IL, USA). GraphPad Prism (GraphPad
Software, Inc., La Jolla, CA, USA) was used to draw graphs in
Fig. 1 and 3. Microsoft Excel 2010 (Microsoft
Corporation, Redmond, WA, USA) was used to draw graphs in Fig. 2 and 4.
Comparison between multiple sets of quantitative data was performed
using single factor (also known as one-way) analysis of variance
(ANOVA), while multiple comparisons between groups were performed
using Fisher's least significant difference t test. Methylation
rates were compared with χ2 test, and Pearson
correlation test was used for correlation analysis. The experiments
were all repeated three times. P<0.05 was considered to indicate
a statistically significant difference.
Results
Expression of miR-375 and miR-203
To identify whether miR-375 and miR-203 were
potentially involved in the severity of cervical lesions, total RNA
was isolated from normal tissues [which served as the negative
control (NC)], CIN and SCC tissues. Using RT-qPCR, the expression
of miR-375 and miR-203 was detected in NC, CIN and SCC tissue
samples. Notably, the relative expression levels of miR-375 and
miR-203 significantly differed between the NC, CIN and SCC groups
(P<0.001; Table II). Furthermore,
miR-375 and miR-203 demonstrated lower expression in CIN and SCC,
compared with NC.
| Table II.Expression levels of miR-375 and
miR-203 in tissues. |
Table II.
Expression levels of miR-375 and
miR-203 in tissues.
| A, miR-375 |
|---|
|
|---|
| Groups | Expression of
miR-375 | t | P-value |
|---|
| NC | 1.02±0.13 | 42.732 | <0.001 |
| CIN | 0.04±0.02 | −8.800 | <0.001 |
| SCC | 0.08±0.02 | 40.869 | <0.001 |
|
| B, miR-203 |
|
| Groups | Expression of
miR-203 | t | P-value |
|
| NC | 1.00±0.17 | 27.278 | <0.001 |
| CIN | 0.13±0.04 | −13.789 | <0.001 |
| SCC | 0.27±0.03 | 23.184 | <0.001 |
Effects of folate on miR-375 and
miR-203 expression
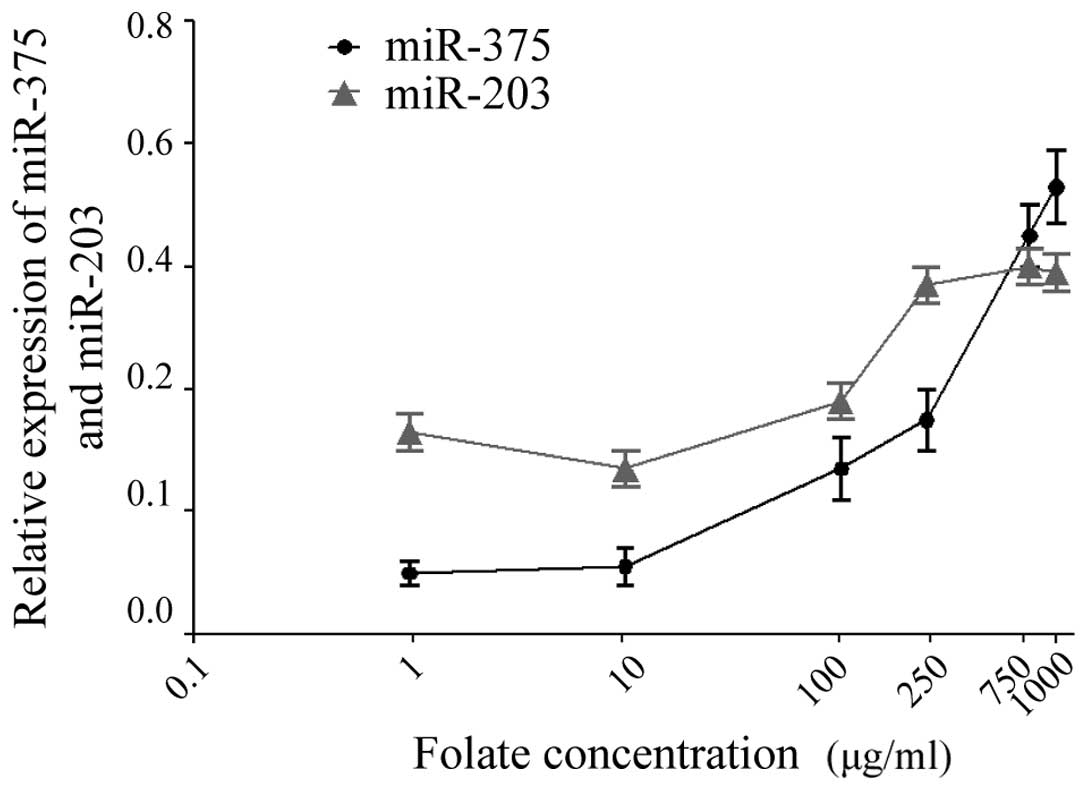
CaSki cells were treated with increasing
concentrations of folate (1, 10, 100, 250, 750 and 1,000 µg/ml).
The expression levels of miR-375 and miR-203 in CaSki cells were
detected by RT-qPCR following treatmen t with folate. The results
revealed that there was a significant difference in the expression
levels of miR-375 and miR-203 in CaSki cells at various folate
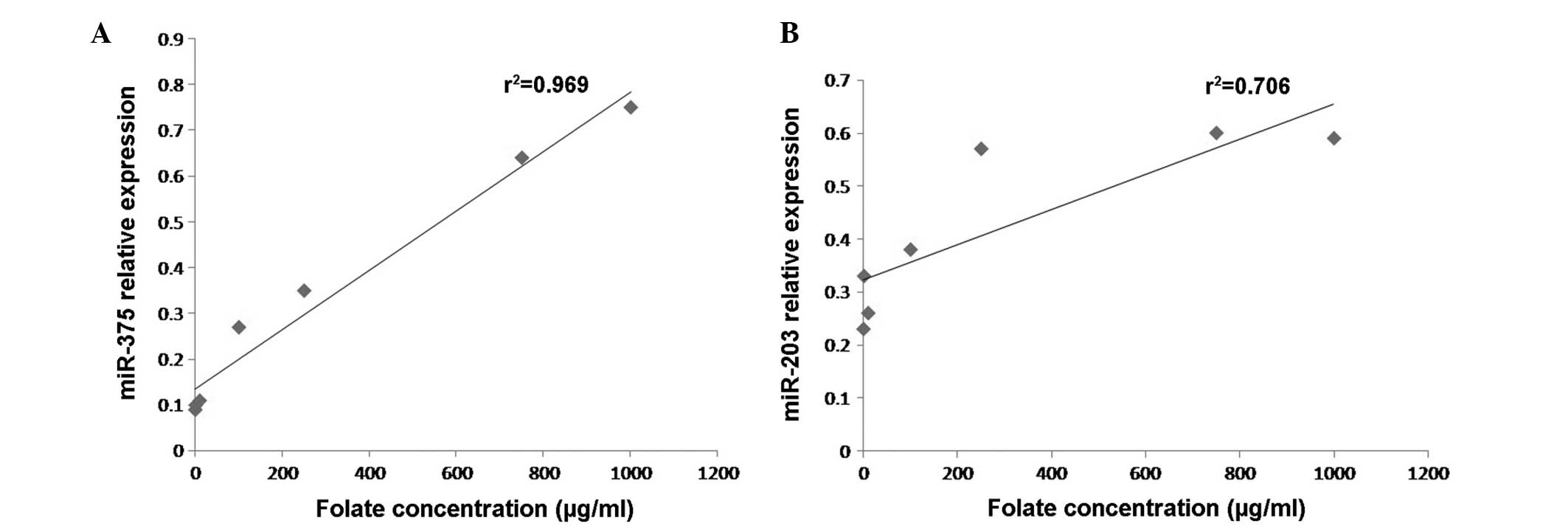
concentrations (one-way ANOVA; P<0.001) (Fig. 1). As indicated in Fig. 2, statistical analysis revealed that
miR-375 and miR-203 were positively correlated with folate
concentration (Pearson correlation test; r2=0.963 and
0.708, respectively; P<0.05).
Effects of folate on miR-375 and
miR-203 methylation
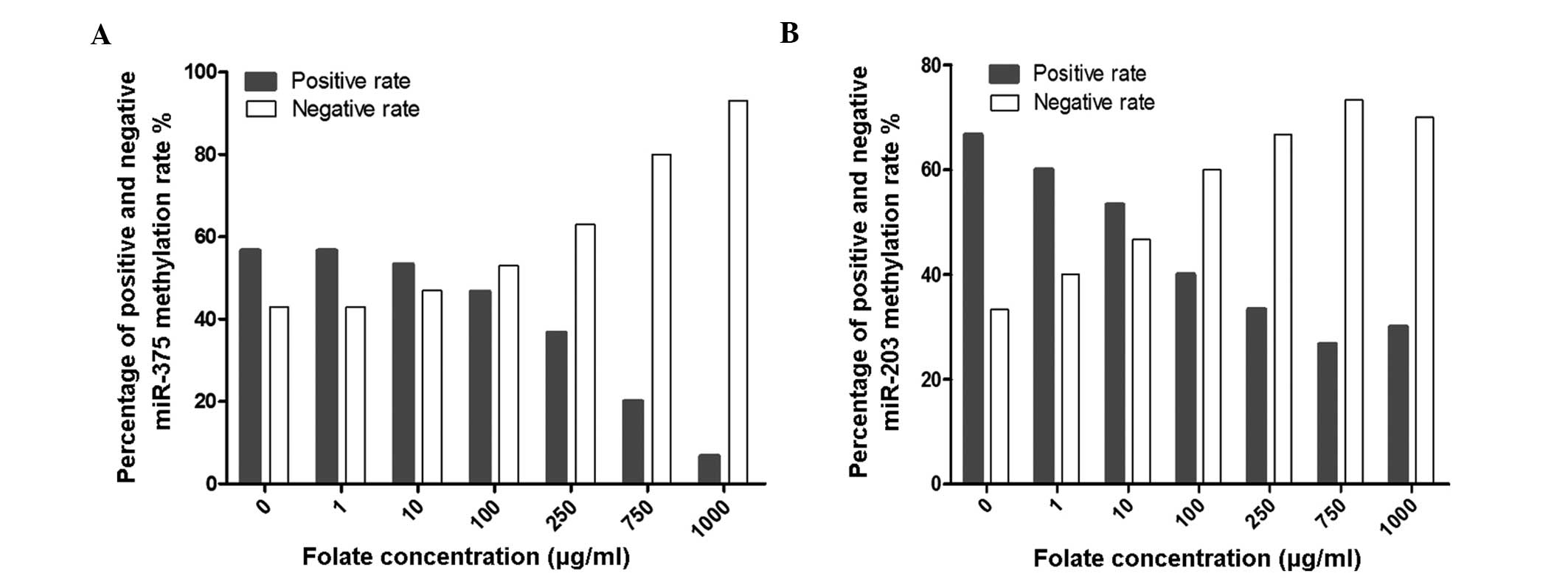
MSP was used to detect the methylation status of
miR-375 and miR-203 following treatment with various concentrations
of folate. The results revealed that the positive rates of miR-375
and miR-203 methylation decreased gradually with increasing
concentrations of folic acid (χ2=25.696, P<0.001 and
χ2=16.716, P=0.006; Fig. 3A
and B, respectively).
Correlation analysis
Statistical analysis revealed that the expression
levels and the methylation status of miR-375 and miR-203 were
negatively correlated (r2=0.788, P<0.001;
r2=0.875, P<0.001; Fig. 4A
and B, respectively).
Discussion
Cervical cancer is the second most aggressive type
of malignant tumor, and causes serious damage to women's health
(22). Each year, there are ~150,000
newly diagnosed cases of cervical cancer in China, which accounts
for almost 1/3 of the total cases worldwide (12). Previous studies have identified that
the occurrence of cervical cancer is associated with HPV infection,
but is additionally influenced by other biological events (23). miR alterations have been demonstrated
to have a significant role in various steps of tumor formation and
progression (13). Similar to certain
growth-suppressive protein-encoding tumor suppressor genes, certain
miR genes harbor CpG islands, which are susceptible to
methylation-mediated silencing (24).
Therefore, hypermethylation-mediated silencing of tumor-suppressive
miRs may be an important mechanism of tumorigenesis. To the best of
our knowledge, only a small number of studies have investigated the
methylation status of selected miRs, including miR-203 and miR-375,
in cervical cancer (25). The present
study evaluated the expression of miR-203 and miR-375 in a range of
tissues obtained from normal, CIN and cervical cancer cases, with
the aim of determining their expression levels in uterine cervix
premalignant and malignant lesions.
miR-203 has been reported to suppress cancer cell
proliferation and invasion (26).
miR-203 is downregulated in several types of cancer, including
hematopoietic tumors (27). In the
cervix, Cheung et al (24)
conducted the first comprehensive miR expression profiling study in
tissues from normal cervical epithelium (n=9) and high-grade CIN
(stage II, n=12 and stage III, n=12) tissues, in order to identify
miR candidates with potential future clinical applications from a
pool of 202 miRs. The results demonstrated that miR-203 may be
useful to distinguish high-grade CIN specimens from normal cervical
epithelium (lower-bound of fold-change >2; P≤0.05) (28). However, Gocze et al (29) used PCR to detect the expression of 8
different miRs in cervical cancer tissues, and observed that the
expression levels of miR-203 were significantly higher in cervical
cancer tissue samples, compared with normal tissues. Therefore, the
expression of miR-203 in cervical cancer remains under debate. In
the present study, it was observed that the expression levels of
miR-203 in CIN and cervical cancer tissues were lower, compared
with that of normal cervical tissue. The results of the present
study were consistent with previous studies with regard to the
global expression pattern of miRs in cervical dysplasia and cancer
(30). Melar-New and Laimins
(31) demonstrated that the E7
protein of high-risk HPV was able to block differentiation-induced
miR-203 upregulation in human keratinocytes via the
mitogen-activated protein kinase/protein kinase C signaling
pathway. Greco et al (32)
revealed that the E5 protein of high-risk HPV downregulated the
expression of miR-203, and therefore may have a significant role in
HPV infection and subsequent transformation through complex
regulatory patterns of ΔNp63 expression in the host cells. These
previous observations may aid to explain the downregulation of
miR-203 observed in the present study.
Alhough miR-375 has primarily been studied in the
context of diabetes, as it influences β-cell mass and insulin
levels, its expression has also been demonstrated to be decreased
in several malignancies (33). Wang
et al (34) used stem-loop
RT-qPCR to detect the expression levels of miR-375 in cervical
cancer tissues (170 cases) and normal cervical tissues (68 cases).
The authors observed that, compared with normal cervical tissues,
the expression levels of miR-375 in cervical cancer tissues were
4.45 times lower, and closely associated with poor prognostic
factors of cervical cancer (34).
Additional studies revealed that miR-375 expression was upregulated
in cervical cancer cells following paclitaxel treatment in
vitro, and in tissues following paclitaxel combination
chemotherapy (35), which may be
induced directly by targeting E-cadherin (36). However, to the best of our knowledge,
the expression of miR-375 in cervical precancerous lesions has not
been reported thus far. The results of the present study indicated
that, compared with the control group, the expression levels of
miR-375 in the CIN group were markedly decreased, suggesting that
low expression levels of miR-375 may be involved in the process of
carcinogenesis.
miR expression may be reduced by several factors,
including transcriptional factors, mutations, deletions and
methylation (37). However, the
mechanism of downregulation of miR-203 and miR-375 in cervical
cancer remains to be elucidated. Previously, Wilting et al
(38) used MSP to analyze a cell line
panel representing various stages of HPV-induced transformation,
and revealed an increase in methylation of miR-203 and miR-375 with
progression to malignancy. Furthermore, the expression of these
miRs was restored following treatment with a demethylating agent
(38). Additional experiments
confirmed that the methylation levels of miR-203 and miR-375 were
also significantly increased in high-grade CIN and cervical cancer
samples, compared with normal tissues (38).
To the best of our knowledge, methylation-mediated
transcriptional repression of miR-203 and miR-375 regulated by
folic acid has not been previously reported for cervical cancer. In
mammals, folate is directly involved in DNA methylation by
providing methyl groups (39).
Therefore, the biological association between folate and
methylation suggests that the influence of folate in cervical
carcinogenesis may be associated with the alteration of miR
expression via methylation of miR's CpG islands (40). To verify this hypothesis, the present
study additionally examined the effect of elevated folate levels on
the methylation rates of miR-203 and miR-375, and observed that
with increasing folate levels in cervical cancer cells, miR-203 and
miR-375 methylation decreased, while the expression of miR-203 and
miR-375 increased. In addition, the methylation rate of the CpG
island in miR-203 and miR-375 and the expression levels of miR-203
and miR-375 were highly negatively correlated, further confirming
that the downregulation of miR-203 and miR-375 in the process of
cervical carcinogenesis is associated with the high methylation
rate of miR-203 and miR-375, which may be caused by a lack of folic
acid.
In summary, to the best of our knowledge, the
present study reports for the first time that folate deficiency may
induce cervical carcinogenesis. Folate deficiency downregulates
miR-203 and miR-375 expression via methylation of their CpG
islands. However, the association between folate, methylation of
miRs and cervical cancer remains to be fully elucidated, and the
function and mechanism of these various components requires
validation in future studies.
Acknowledgements
The present study was supported by grants from the
Special Public Welfare Industry Research of the National Health and
Family Planning Commission (Beijing, China; grant no. 201402010)
and the Shanxi Health and Family Planning Commission Youth Project
(Taiyuan, China; grant no. 201301015).
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Schiffman MH and Castle P: Epidemiologic
studies of a necessary causal risk factor: Human papillomavirus
infection and cervical neoplasia. J Natl Cancer Inst. 95:E22003.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Choi YJ and Park JS: Clinical significance
of human papillomavirus genotyping. J Gynecol Oncol. 27:e212016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ho GY, Bierman R, Beardsley L, Chang CJ
and Burk RD: Natural history of cervicovaginal papillomavirus
infection in young women. N Engl J Med. 338:423–428. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhao WH, Hao M, Cheng XT, Yang X, Wang ZL,
Cheng KY, Liu FL and Bai YX: c-myc gene copy number variation in
cervical exfoliated cells detected on fluorescence in situ
hybridization for cervical cancer screening. Gynecol Obstet Invest.
Jan 26–2016.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wilting SM, Snijders PJ, Verlaat W,
Jaspers A, van de Wiel MA, van Wieringen WN, Meijer GA, Kenter GG,
Yi Y, le Sage C, et al: Altered microRNA expression associated with
chromosomal changes contributes to cervical carcinogenesis.
Oncogene. 32:106–116. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang L, Yue Y, Wang X and Jin H: Function
and clinical potential of microRNAs in hepatocellular carcinoma.
Oncol Lett. 10:3345–3353. 2015.PubMed/NCBI
|
|
9
|
Lo R and Weksberg R: Biological and
biochemical modulation of DNA methylation. Epigenomics. 6:593–602.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sarkar D, Leung EY, Baguley BC, Finlay GJ
and Askarian-Amiri ME: Epigenetic regulation in human melanoma:
Past and future. Epigenetics. 10:103–121. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang LQ and Chim CS: DNA methylation of
tumor-suppressor miRNA genes in chronic lymphocytic leukemia.
Epigenomics. 7:461–473. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang JT, Ding L, Jiang SW, Hao J, Zhao WM,
Zhou Q, Yang ZK and Zhang L: Folate deficiency and aberrant
expression of DNA methyltransferase 1 were associated with cervical
cancerization. Curr Pharm Des. 20:1639–1646. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Flatley JE, McNeir K, Balasubramani L,
Tidy J, Stuart EL, Young TA and Powers HJ: Folate status and
aberrant DNA methylation are associated with HPV infection and
cervical pathogenesis. Cancer Epidemiol Biomarkers Prev.
18:2782–2789. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Flatley JE, Sargent A, Kitchener HC,
Russell JM and Powers HJ: Tumour suppressor gene methylation and
cervical cell folate concentration are determinants of high-risk
human papillomavirus persistence: A nested case control study. BMC
Cancer. 14:8032014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bai LX, Wang JT, Ding L, Jiang SW, Kang
HJ, Gao CF, Chen X, Chen C and Zhou Q: Folate deficiency and FHIT
hypermethylation and HPV 16 infection promote cervical
cancerization. Asian Pac J Cancer Prev. 15:9313–9317. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lewin SN, Herzog TJ, Barrena Medel NI,
Deutsch I, Burke WM, Sun X and Wright JD: Comparative performance
of the 2009 International Federation of Gynecology and Obstetrics'
staging system for uterine corpus cancer. Obstet Gynecol.
116:1141–1149. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu HQ, Wang YH, Wang LL and Hao M:
P16INK4A and survivin: Diagnostic and prognostic markers in
cervical intraepithelial neoplasia and cervical squamous cell
carcinoma. Exp Mol Pathol. 99:44–49. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wright TC, Ronnett BM, Kurman RJ and
Ferenczy A: Precancerous lesions of the cervix. Kurman RJ, Ellenson
LH and Ronnett BM: Blaustein's Pathology of the Female Genital
Tract (6th). Springer US. (New York, NY). 193–252. 2011. View Article : Google Scholar
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xiao W, Zhao H, Dong W, Li Q, Zhu J, Li G,
Zhang S and Ye M: Quantitative detection of methylated NDRG4 gene
as a candidate biomarker for diagnosis of colorectal cancer. Oncol
Lett. 9:1383–1387. 2015.PubMed/NCBI
|
|
22
|
Shi JF, Canfell K, Lew JB and Qiao YL: The
burden of cervical cancer in China: Synthesis of the evidence. Int
J Cancer. 130:641–652. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang JT, Ma XC, Cheng YY, Ding L and Zhou
Q: A case-control study on the association between folate and
cervical cancer. Zhonghua Liu Xing Bing Xue Za Zhi. 27:424–427.
2006.(In Chinese). PubMed/NCBI
|
|
24
|
Cheung TH, Man KN, Yu MY, Yim SF, Siu NS,
Lo KW, Doran G, Wong RR, Wang VW, Smith DI, et al: Dysregulated
microRNAs in the pathogenesis and progression of cervical neoplasm.
Cell Cycle. 11:2876–2884. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jiménez-Wences H, Peralta-Zaragoza O and
Fernández-Tilapa G: Human papilloma virus, DNA methylation and
microRNA expression in cervical cancer (Review). Oncol Rep.
31:2467–2476. 2014.PubMed/NCBI
|
|
26
|
Yao T, Rao Q, Liu L, Zheng C, Xie Q, Liang
J and Lin Z: Exploration of tumor-suppressive microRNAs silenced by
DNA hypermethylation in cervical cancer. Virol J. 10:1752013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Michel CI and Malumbres M: microRNA-203:
Tumor suppression and beyond. Microrna. 2:118–126. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen T, Xu C, Chen J, Ding C, Xu Z, Li C
and Zhao J: MicroRNA-203 inhibits cellular proliferation and
invasion by targeting Bmi1 in non-small cell lung cancer. Oncol
Lett. 9:2639–2646. 2015.PubMed/NCBI
|
|
29
|
Gocze K, Gombos K, Juhasz K, Kovacs K,
Kajtar B, Benczik M, Gocze P, Patczai B, Arany I and Ember I:
Unique microRNA expression profiles in cervical cancer. Anticancer
Res. 33:2561–2567. 2013.PubMed/NCBI
|
|
30
|
Gocze K, Gombos K, Kovacs K, Juhasz K,
Gocze P and Kiss I: MicroRNA expressions in HPV-induced cervical
dysplasia and cancer. Anticancer Res. 35:523–530. 2015.PubMed/NCBI
|
|
31
|
Melar-New M and Laimins LA: Human
papillomaviruses modulate expression of microRNA 203 upon
epithelial differentiation to control levels of p63 proteins. J
Virol. 84:5212–5221. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Greco D, Kivi N, Qian K, Leivonen SK,
Auvinen P and Auvinen E: Human papillomavirus 16 E5 modulates the
expression of host microRNAs. PLoS One. 6:e216462011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Costa-Pinheiro P, Ramalho-Carvalho J,
Vieira FQ, Torres-Ferreira J, Oliveira J, Gonçalves CS, Costa BM,
Henrique R and Jerónimo C: MicroRNA-375 plays a dual role in
prostate carcinogenesis. Clin Epigenetics. 7:422015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang F, Li Y, Zhou J, Xu J, Peng C, Ye F,
Shen Y, Lu W, Wan X and Xie X: miR-375 is down-regulated in
squamous cervical cancer and inhibits cell migration and invasion
via targeting transcription factor SP1. Am J Pathol. 179:2580–2588.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shen Y, Wang P, Li Y, Ye F, Wang F, Wan X,
Cheng X, Lu W and Xie X: miR-375 is upregulated in acquired
paclitaxel resistance in cervical cancer. Br J Cancer. 109:92–99.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shen Y, Zhou J, Li Y, Ye F, Wan X, Lu W,
Xie X and Cheng X: miR-375 mediated acquired chemo-resistance in
cervical cancer by facilitating EMT. PLoS One. 9:e1092992014.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Adams BD, Kasinski AL and Slack FJ:
Aberrant regulation and function of microRNAs in cancer. Curr Biol.
24:R762–776. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wilting SM, Verlaat W, Jaspers A, Makazaji
NA, Agami R, Meijer CJ, Snijders PJ and Steenbergen RD:
Methylation-mediated transcriptional repression of microRNAs during
cervical carcinogenesis. Epigenetics. 8:220–228. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Geng Y, Gao R, Chen X, Liu X, Liao X, Li
Y, Liu S, Ding Y, Wang Y and He J: Folate deficiency impairs
decidualization and alters methylation patterns of the genome in
mice. Mol Hum Reprod. 21:844–856. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ho E, Beaver LM, Williams DE and Dashwood
RH: Dietary factors and epigenetic regulation for prostate cancer
prevention. Adv Nutr. 2:497–510. 2011. View Article : Google Scholar : PubMed/NCBI
|