Introduction
Breast cancer is the most common malignancy in
females in the United States, and the disease is expected to cause
40,430 mortalities in 2014 according to the National Cancer
Institute's Surveillance, Epidemiology and End Results (SEER)
Program (1). Although breast cancer
is one of the most life-threatening diseases, the molecular
processes involved in its pathogenesis have not yet been fully
elucidated. Various risk factors, including family history, genetic
mutations, reproductive history and environmental alterations,
correspond to cancer development (2,3). Advances
in modern biochemistry provide numerous technical strategies for
illustrating the etiology of breast malignancies. However, the
complexity of cancer evolution, which involves a number of genes
and molecular pathways, prevents further investigation of
underlying mechanisms.
Over the past few decades, developments in early
cancer detection and the combination of multiple treatments have
significantly reduced breast cancer mortality (1). Among these, targeted therapy is one of
the most promising strategies. However, with ever-increasing new
cases and the development of drug resistance, there is an urgent
requirement to identify more effective methods to deliver
biochemical therapies.
Previous studies have demonstrated that
phosphoinositide 3-kinase (PI3K) signaling plays a critical role in
cell proliferation. Within this pathway, key components including
PIK and AKT have emerged as potential targets for drug development
(4,5).
However, a closely related protein, serum- and
glucocorticoid-regulated protein kinase (SGK), which shares 54% of
its identity with the kinase domains of AKT, has received little
attention (6). Vasudevan et al
(7) reported that a number of cancer
cell lines are SGK3-dependent but exhibit only minimal AKT
activation, indicating that SGK3 is an essential independent factor
in the PI3K pathway and providing an alternative explanation for
tumor behavior. SGK is active in various cell signaling pathways
and serves as a significant factor in cell phosphorylation, which
subsequently triggers downstream substrates to stimulate cellular
responses, including cell survival, differentiation and material
transduction. SGK3, first described by Kobayashi et al
(6), differs significantly in terms
of enzyme structure, function, corresponding gene locus and
regulation of protein expression compared with its isoform SGK1,
which has been more frequently investigated. Current animal
experimental data provide limited information with regard to the
function of SGK3, and do not address the application of this enzyme
in humans. However, previous studies have revealed that increased
SGK3 expression in various types of tumor cells contributes to
cancer development as well as tumor cell infiltration and
migration, suggesting that this enzyme may be a potential target in
disease prognosis and drug development (8).
In this study, we investigated SGK3 expression in
human breast tissues and the potential effects of increased SGK3
levels on tumor cell multiplication, invasion, migration and
apoptosis. We also examined the underlying mechanisms of these
phenomena.
Materials and methods
Cells and plasmids
The human breast cancer cell line MDA-MB-231 (Cancer
Institution, Chinese Academy of Medical Science, Beijing, China)
was cultured in RPMI-1640 media containing 10% fetal bovine serum
(FBS) at 37°C with 5% CO2. The pAcG-4T3-SKG3 plasmid,
which contains 1.3 kb of human SGK3 cDNA with
BamHI/XhoI sites, was provided by Dr. Xianming Xia
(Johns Hopkins University School of Medicine, Baltimore, MD, USA).
The pEGFP-N1 plasmid was a gift from Dr. Changzhi Huang (Cancer
Institute and Hospital, Chinese Academy of Medical Sciences and
Peking Union Medical College, Beijing, China).
Reagents and antibodies
The following reagents and antibodies were applied
in this study: breast tissue chips BR725 (Alenabio Company, Xi'an,
China), RPMI-1640 media (Gibco Life Technologies, Carlsbad, CA,
USA), FBS (Invitrogen Life Technologies, Carlsbad, CA, USA),
trypsin (CAS number 9002-07-7, Sigma-Aldrich, St. Louis, MO, USA),
BamHI/XhoI and PrimeScript™ RT-PCR kit (Takara
Biotechnology Co., Ltd., Dalian, China), PCR enzyme mix and
Advantage PCR cloning kit (Clontech Laboratories, Inc., Mountain
View, CA, USA), SV Total RNA isolation system (Promega Corporation,
Madison, WI, USA), radioimmunoprecipitation assay (RIPA) buffer
(Applygen Technologies, Inc., Beijing, China), anti-β-actin,
anti-SGK3, anti-BAD, anti-Bcl-xL, anti-matrix metalloproteinase 2
(MMP2), anti-MMP9, anti-phospho-GSK-3β, anti-phospho-β-catenin
(CTS, Boston, MA, USA), anti-breast cancer metastasis suppressor 1
(BRMS1) and horseradish peroxidase-labeled secondary antibody
(Santa Cruz Biotechnology, Inc., Dallas, Texas, USA). Sangon
Biotech (Shanghai, China) synthesized all primers.
Specimens and tissue microarray
analyses
Breast tissue microarray analyses were performed to
assess SGK3 expression in formalin-fixed, paraffin-embedded
specimens from various breast tissues, including 24 normal breast
samples, 24 breast carcinoma specimens and 24 adjacent noncancerous
tissues. The clinicopathological data are presented in Table I. Visible brown ink in cells using
fluorescence microscopy indicated positive SGK3 expression. The
integral optical density (IOD) of each image was analyzed using
Image-Pro Plus software (Media Cybernetics, Rockville, MD, USA).
The study was approved by the Institutional Review Board of Qiqihar
Medical University, China, and patient consent was waived.
 | Table I.Clinicopathological characteristics of
enrolled breast cancer patients and tissues. |
Table I.
Clinicopathological characteristics of
enrolled breast cancer patients and tissues.
| Clinicopathological
factors | Mean or number |
|---|
| Age (years) | 47.22±8.90 |
| Tumor type |
|
| Invasive
ductal carcinoma | 20 |
| Atypical
medullary carcinoma | 2 |
| Mucinous
adenocarcinoma | 2 |
| Histological
grade |
|
| 1 | 1 |
| 2 | 19 |
| 3 | 1 |
| N/A | 3 |
Reconstruction of pEGFP-N1-SGK3
A full-length clone encoding human SGK3 was
obtained as follows: XhoI and BamHI sites were
introduced via the forward and reverse primers, which were designed
according to the SGK3 cDNA using Primer Premier 5.0 software
(Premier Biosoft, Palo Alto, CA, USA). SGK3 was amplified by
polymerase chain reaction (PCR) using pAcG-4T3-SKG3 as a template
and the specific primers 5′-CCGCTCGAGACCATGGCCCTGAAGATTC-3′
(forward) and 5′-CGCGGATCCAAAAATAAGTCTTCTG-3′ (reverse). Purified
PCR products and the pEGFP-N1 vector were digested with XhoI
and BamHI, and subsequently ligated to construct
pEGFP-N1-SGK3. Positive clones were analyzed by sequencing
(Invitrogen, Shanghai, China) and cross-comparisons were performed
using the results and data from GenBank [ClustalW software,
European Molecular Biology Laboratory - European Bioinformatics
Institute (EMBL-EBI)].
Plasmid transfection
Logarithmic phase MDA-MB-231 cells were seeded into
six-well plates (Nunc, Roskilde, Denmark) at 24 h before
transfection to achieve a density of 2×105 cells/well.
Each well was transfected with 3 µg pEGFP-N1-SGK3 or pEGFP-N1 using
GenEscort™ I (Isegen, Nanjing, China) according to the
manufacturer's instructions. Forty-eight hours after transfection,
the treated cells were harvested and SGK fusion protein analyses
were performed. Cells successfully transfected with pEGFP-N1-SGK3
or pEGFP-N1 were termed group SGK3 and group N1 (negative control),
respectively. MDA-MB-231 cells served as a blank control (group
231). All in vitro experiments were carried out for each
group.
Cell growth assays
Logarithmic phase tumor cells were placed separately
in four wells in a 96-well plate (500 cells/well in five plates).
As described by Shi et al (9)
one plate was used for MTT assays for five consecutive days.
Briefly, 10 µl MTT reagent was added to each well and cultured for
2 h at 37°C. Media were discarded and cells were incubated with 150
µl dimethyl sulfoxide for 15 min with mild agitation. Cells were
counted every day by measuring the absorbance at 5700 nm. Cell
growth curves were subsequently generated.
Colony formation assays
Cells were seeded in six-well plates (500
cells/well) and cultured for 2 weeks at 37°C with 5%
CO2. Each sample was performed in triplicate, and
culture media were changed every 3 days. Cells were fixed in
methanol for 30 min, stained with crystal violet, and air-dried.
The colony formation rate = number of colonies / number of seeded
cells.
Cell cycle analyses
Tumor cells from the three groups were seeded in
six-well plates (1×105 cells per well) and incubated for
48 h before digestion with 0.25% trypsin. All cells were washed
twice with pre-cooled phosphate-buffered saline (PBS) and incubated
for 30 min at room temperature with propidium iodide (PI;
Sigma-Aldrich). Cells were immediately examined by flow cytometry
using a FACSCalibur (Becton-Dickinson, New York, NY, USA). Data
were analyzed using ModFit LT (Verity Software House, Topsham, ME,
USA).
Horizontal migration tests (scratch
tests)
Tumor cells were placed in six-well plates
(1×105 cells per well). Once a single layer of cells had
formed, two scratches were made by swiping a small sterilized tip
across the inner surface of the bottom of the plate. The scratch
area was washed with PBS until the cells were removed. RPMI-1640
was added and cells were cultured for 24 h. The distance between
cells at 0, 12 and 24 h in the scratch area was measured using
microscopy. Horizontal cell migration rates were automatically
analyzed using Image-Pro Plus software.
Vertical migration tests (Transwell
tests)
Logarithmic growth phase cells were cultured in
serum-free media for 12–24 h, and then collected and suspended at a
density of 2×105 cells/ml in media containing 0.2% FBS.
Cell suspensions (200 µl) were added to the upper Transwell
chamber, and 600 µl RPMI-1640 media containing 10% FBS was added to
the lower chamber. After 24 h, cells that remained in the upper
membrane were gently removed with cotton and washed three times
with PBS. Cells that migrated to the lower membrane were fixed in
methanol for 30 min and stained for 20 min with 0.1% crystal
violet. Cells were counted in five random fields of the lower
membrane to identify the number of vertically migrating cells.
mRNA expression of apoptosis- and
invasion-related genes
We isolated total RNA from tumor cells using the SV
Total RNA isolation kit. cDNA was prepared using the RT-PCR kit as
per the manufacturer's instructions. The sequences of primers are
listed in Table II. Reactions were
subjected to 30 cycles of 94°C denaturation for 4 min, 94°C melting
for 30 sec, annealing for 30 sec (specific annealing temperatures
are listed in Table II), 72°C
extension for 60 sec, with a final elongation step at 72°C for 10
min. PCR products were separated by electrophoresis in 1% agarose
gels, visualized using ethidium bromide, and analyzed using the
AlphaImager HP system (ProteinSimple, San Jose, CA, USA).
| Table II.Primer sequences used for apoptosis-
and invasion-related mRNA analyses. |
Table II.
Primer sequences used for apoptosis-
and invasion-related mRNA analyses.
| mRNA | Sequence | Annealing temperature
(°C) | Length (bp) |
|---|
| bad | F
5′-CCATCCCTTCGTCGTCCTC-3′ | 51.0 | 162 |
|
| R
5′-GCTCCGGCAAGCATCATC-3′ |
|
|
| bcl-xl | F
5′-CGGGCATTCAGTGACCTGAC-3′ | 54.3 | 151 |
|
| R
5′-TCAGGAACCAGCGGTTGAAG-3′ |
|
|
| mmp2 | F
5′-GTATTTGATGGCATCGCTCA-3′ | 52.0 | 196 |
|
| R
5′-CATTCCCTGCAAAGAACACA-3′ |
|
|
| mmp9 | F
5′-GGAGCCGCTCTCCAAGAAGCTT-3′ | 58.6 | 522 |
|
| R
5′-CTCCTCCCTTTCCTCCAGAACAGAA-3′ |
|
|
| brms1 | F
5′-ACTGAGTCAGCTGCGGTTGCGG-3′ | 61.7 | 335 |
|
| R
5′-AAGACCTGGAGCTGCCTCTGGCGTGC-3′ |
|
|
| β-actin | F
5′-CATCTCTTGCTCGAAGTCCA-3′ | 52.8 | 263 |
|
| R
5′-ATCATGTTTGAGACCTTCAACA-3′ |
|
|
Western blot analysis
Cells were collected by centrifugation for 5 min at
800 rpm, and proteins were extracted following lysis in RIPA
buffer. Protein concentrations were determined using protein assays
(Amersham Biosciences, London, England). Purified proteins (50 µl)
were separated by sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) and transferred to nitrocellulose
membranes. Membranes were blocked for 2 h at room temperature in 5%
non-fat milk and incubated overnight at 4°C with primary antibodies
against BAD, Bcl-xL, MMP2, MMP9 and BRMS1 (1:1,000 dilution).
Following incubation with the horseradish peroxidase-labeled
secondary antibodies (1:5,000 dilution), membranes were washed and
proteins were visualized using chemiluminescence reagents. Western
blotting results were quantified using ImageJ software (National
Institutes of Health, Bethesda, MD, USA).
Statistical analyses
SPSS version 17.0 (SPSS Inc., Chicago, IL, USA) was
used for data analyses. All results are presented as the means ±
standard deviation. One-way analysis of variance and Student's
t-tests were used to compare the difference in means within groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
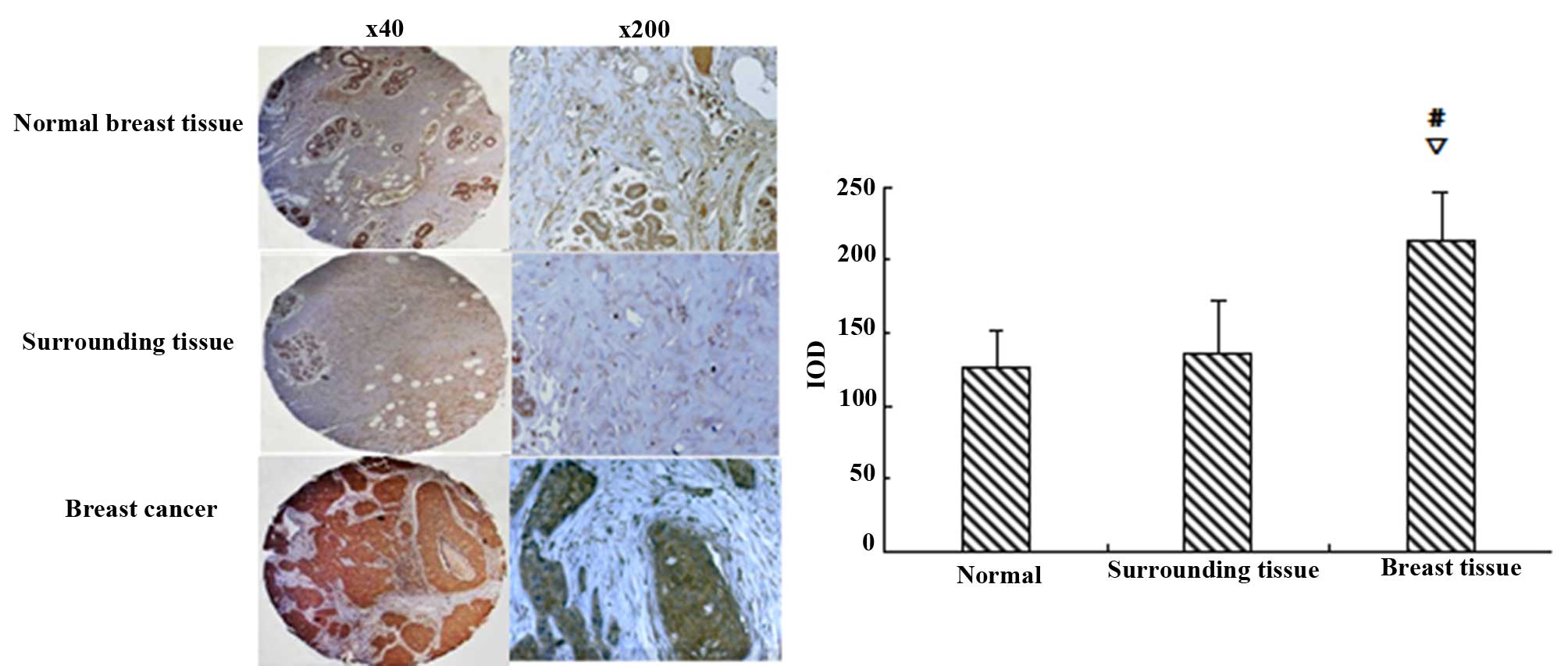
SGK3 expression in breast tissues
The IOD of each enrolled breast tissue revealed no
significant differences in SGK3 expression between normal and
adjacent noncancerous tissues (P>0.05). However, SGK3 levels
were significantly higher in breast carcinoma samples compared with
normal and adjacent noncancerous tissues (P<0.01, Fig. 1).
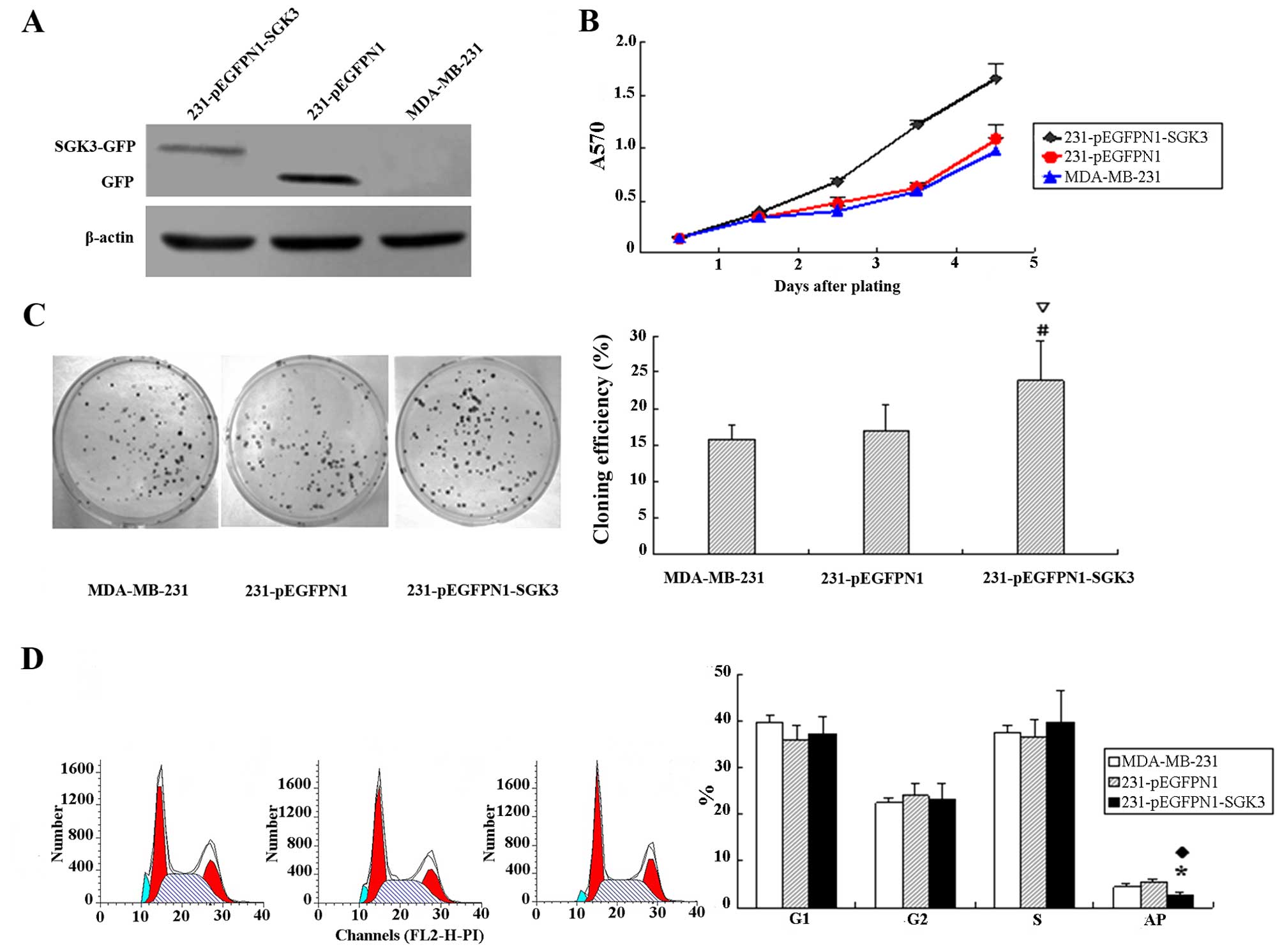
Verification of SGK3 expression in
transfected tumor cells
pEGFP-N1-SGK3 and pEGFP-N1 were successfully
transfected into MDA-MB-231 cells. Western blotting results
revealed SGK3 expression in corresponding host cells; however, the
level of SGK3-green fluorescent protein (GFP) fusion protein in
cells from the SGK3 group was slightly lower than that of GFP in N1
group negative control cells (Fig.
2A).
SGK3 contributes to tumor cell
proliferation
The cell growth curves of each group are presented
in Fig. 2B. We observed accelerating
tumor multiplication in cells from the SGK3 group, whereas the
proliferation rate of N1 and 231 group cells remained low.
Additionally, the doubling times were 27.6, 34.5 and 35.1 h,
respectively, for SCK3, N1 and 231 group cells. These data
demonstrated that SGK3 induced cell proliferation.
Increased colony formation in
SGK3-expressing tumor cells
Significantly higher colony formation rates were
detected in cells from the SGK3 group compared with those from
groups N1 and 231 (16.3±1.96%, 17.1±2.12% and 22.5±1.6%,
respectively; P<0.01). The colony formation rates in N1 and 231
cells were not significantly different (P>0.05, Fig. 2C).
SGK3 inhibits apoptosis
Figure 2D reveals a
lower proportion of apoptotic cells in SGK3-overexpressing cells
compared with cells in group N1 and 231 (P<0.05). No significant
differences were observed with regard to the percentage of G1, G2
and S phase cells (P>0.05).
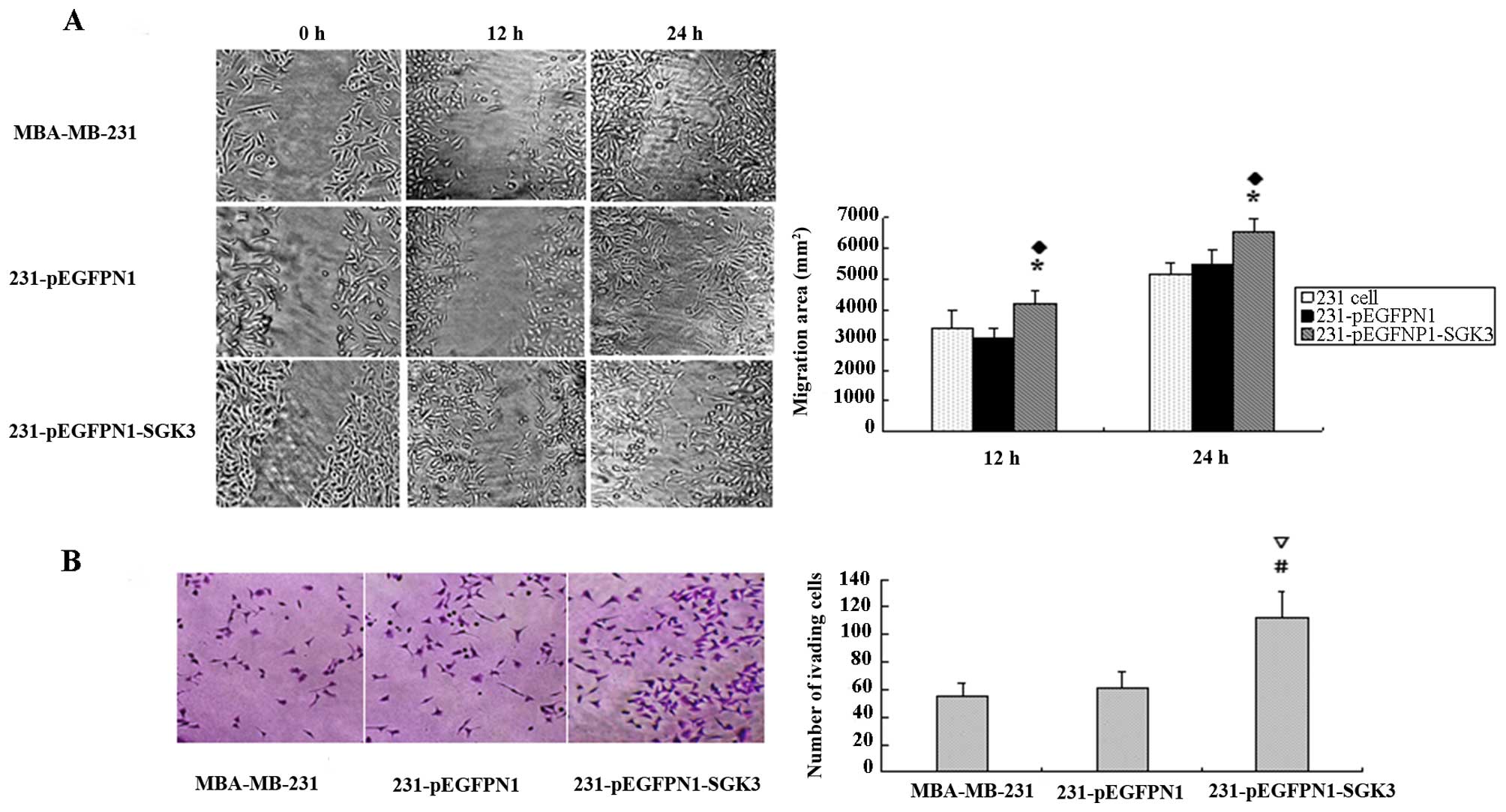
Cell migration in SGK3-overexpressing
cells
Horizontal migration tests revealed that scratches
in the blank and negative control groups remained virtually
unchanged after 12 h, whereas SGK3-overexpressing cells revealed
higher migration rates (P<0.05). After 24 h, scratches were
hardly visible in the SGK3 group cells; however, N1 and 231 cells
had not yet recovered from the damage (P<0.05, Fig. 3A). We also observed similar results in
vertical migration tests. Larger numbers of SGK3 group cells
migrated to the lower membrane compared with N1 and 231 cells
(P<0.01), but no differences were observed between the latter
two groups (P>0.05, Fig. 3B).
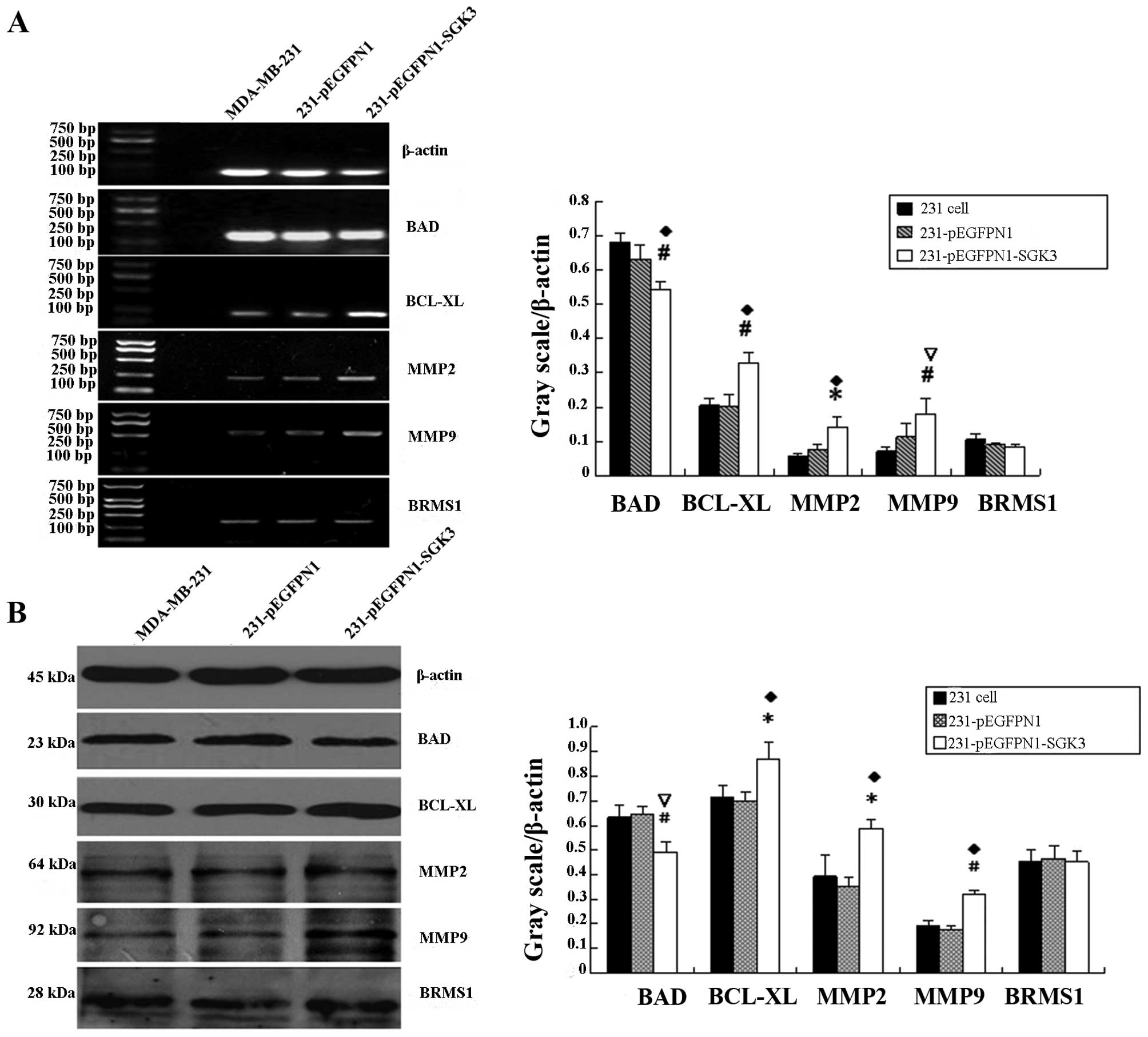
mRNA expression of apoptosis- and
invasion-related genes
RT-PCR product analyses using the AlphaImager HP
system revealed that the grayscale values of electrophoretic bands
for bcl-xl, mmp2 and mmp9 in the SGK3 group
were significantly higher compared with the other two groups
(P<0.05, P<0.01). However, bad mRNA expression
decreased in SGK3-overexpressing cells (P<0.05, Fig. 4A). brms1 mRNA levels were
comparable among the three groups (P>0.05). No significant
differences in these genes were noted between the N1 and 231 group
cells (P>0.05).
Western blot analysis of apoptosis-
and migration-related proteins
Bcl-xL, MMP2 and MMP9 expression was significantly
higher in SGK3-overexpressing cells compared with the other two
groups (P<0.05, P<0.01), whereas BAD expression was decreased
(P<0.01, Fig. 4B). There was no
difference in BRMS1 expression among the three groups (P>0.05).
In the blank and negative control groups, no significant
differences were detected with regard to these five proteins
(P>0.05).
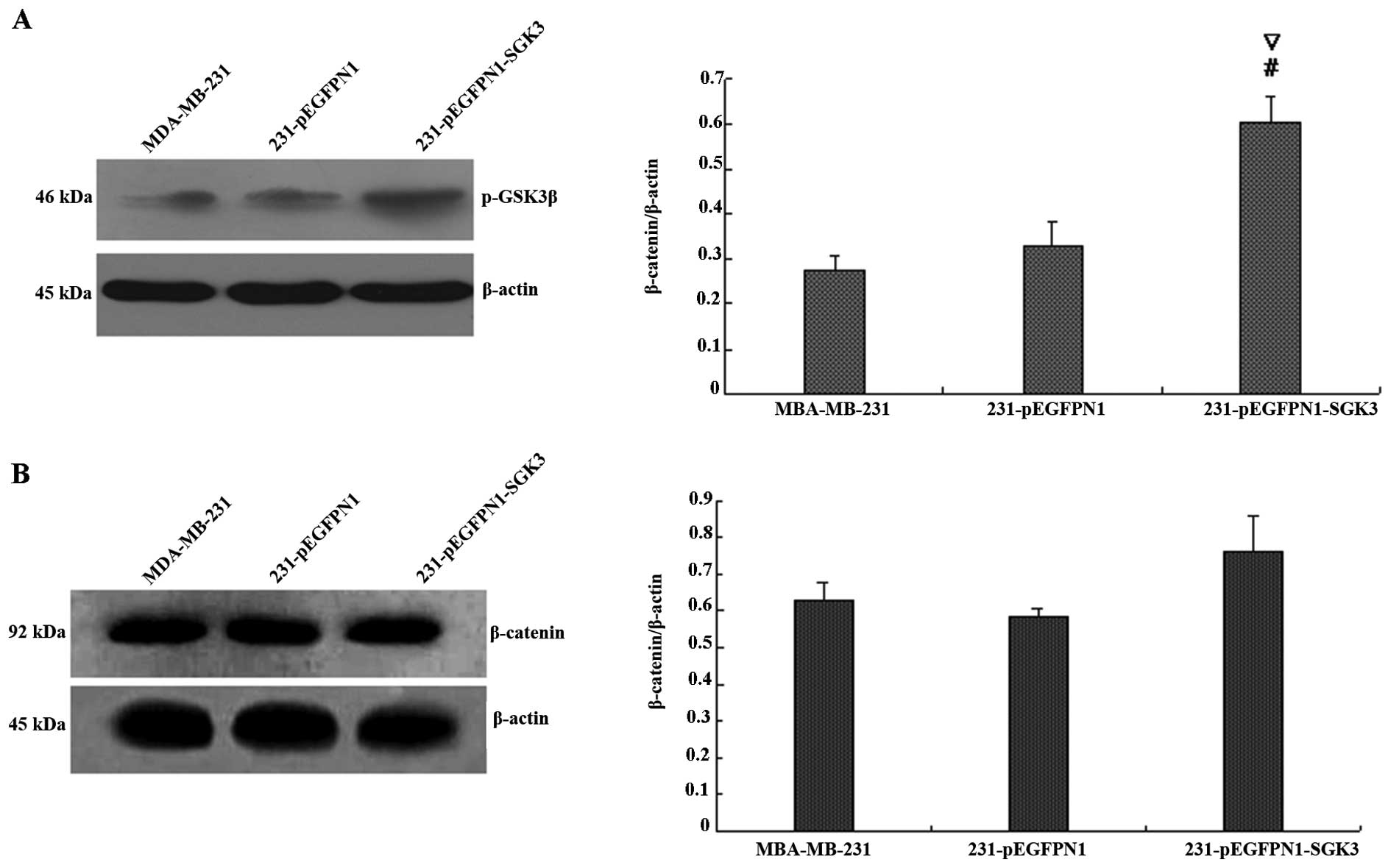
Effect of SGK3 overexpression on PI3K
signaling
We next examined the expression of phospho-GSK3β and
phospho-β-catenin (Fig. 5A and B).
The grayscale value of phospho-GSK3β demonstrated that expression
was higher in the SGK3 group cells (P<0.01), whereas no
differences were detected between N1 and 231 group cells
(P>0.05). In contrast, western blot analyses revealed that
SGK3-overexpressing cells did not differ in phospho-β-catenin
levels compared with the other two groups (P>0.05). Thus, SGK3
overexpression did not contribute to β-catenin phosphorylation.
Discussion
In this study, we observed higher expression levels
of SGK3 in breast cancer tissues compared with adjacent
noncancerous and normal breast tissues using microarray assay.
These data are similar to previous publications evaluating SGK3
expression in prostate, breast and liver cancers (8,10–12). Our findings confirm that SGK3 may be a
specific target for drug development and delivery, since this
protein is strictly expressed in tumor cells.
The results of gene analyses in our study indicated
successful reconstruction of the eukaryotic expression vector
pEGFP-N1-SGK3 using the GenEscort I system. This expression
construct may be used for future investigations of breast cancer
invasion and migration, along with their corresponding mechanisms.
GFP labeling was also employed to achieve better visualization of
the protein. The transfection system had an efficiency of greater
than 70% when delivering the plasmid to MDA-MB-231 cells, and our
transfection results were verified by western blot analysis in the
different groups of cells.
Cell cycle analyses revealed that cells in the SGK3
group had lower rates of apoptosis compared with group N1 and 231
cells, indicating that this protein may alter the behavior of tumor
cells. Additionally, as demonstrated by the cell growth curves,
SGK3-overexpressing cells presented more robust proliferation rates
in the same logarithmic phase as the blank and negative control
groups. Moreover, migration tests verified that SGK3 contributed to
both tumor cell recovery and invasion. Scratch tests illustrated
that SGK3 cells were more resistant to external damage and
regenerated within 24 h. Transwell assays revealed that higher
numbers of SGK3-overexpressing cells migrated to the lower chamber
compared with N1 and 231 group cells. These data demonstrate that
expression of exogenous SGK3 cDNA significantly improved breast
cancer cell multiplication and invasion.
Gene dysregulation is a critical biological process
contributing to tumor development. This theory postulates that
uncontrolled cell proliferation and decreased apoptosis are caused
by gene dysregulation and an upset in the balance between cell
growth and death, thereby leading to tumor formation. Apoptosis has
become a major target for ameliorating drug sensitivity and
enhancing drug delivery (13).
Molecules in the BCL-2 family, including BAD and Bcl-xL, are
notable factors in cell apoptosis and contribute to a so-called
‘apoptosis checkpoint’. Our study demonstrated that SGK3
upregulates bcl-xl mRNA and protein levels, which inhibits
the apoptosis process. Simultaneously, the expression of bad
mRNA and protein, which facilitates apoptosis, significantly
decreased. Another pivotal malignant behavior is the ability of
invasion and migration, which are considered together as the most
critical cause of patient mortality in clinical practice. MMPs have
great potential for improving tumor invasion and migration
(14–18), and also serve as biomarkers for
predicting cancer development (15,16).
RT-PCR and western blot assays indicated that SGK3 enhanced the
mRNA and protein levels of MMP2 and MMP9, which further advanced
tumor progression. Previous studies have reported that BRMS1 plays
a critical role in metastasis in various tumors, including breast,
ovarian, melanoma and hepatocellular carcinoma (19–22).
Notably, BRMS1 mRNA and protein levels in the SGK3 group cells were
not significantly different compared with cells in groups N1 and
231. One possible explanation for this discrepancy might be that
other critical molecules involved in the process between matrix
invasion and distant metastasis were not fully examined, and they
may not respond to altered SGK3 levels.
GSK-3β is a downstream component of SGK3 in the PI3K
pathway, and as expected, SGK3 levels significantly affected GSK-3β
phosphorylation. However, β-catenin phosphorylation did not change
with changes in SGK3 expression. Phosphorylation of β-catenin at
specific Ser/Thr residues targets it for degradation via the
ubiquitin proteasome pathway. However, mutations in β-catenin that
cause dephosphorylation increase accumulation of the non-degradable
protein, ultimately leading to activation of cancerous genes.
β-catenin is crucial for Wnt-Ca2+ signaling, and the
absence of a response to SGK3 suggests that SGK3 is not a
significant component in this pathway. We propose that the
SGK3-mediated aggressive behavior of breast cancer cells is
conducted via the PI3K pathway.
This study has certain limitations. First, we
presented preliminary data of SGK3 function in a small group of
breast tissues. Additionally, our samples were primarily limited to
invasive ductal carcinoma. The behavior of different breast cancer
subtypes and different histology grades when SGK levels are altered
has not yet been examined. Finally, the aggressiveness of cancer
cells following SGK gene silencing should be evaluated.
In conclusion, SGK3 overexpression induced
phosphorylation of GSK-3β, which is downstream in PI3K signaling,
and consequently enhanced the level of apoptosis- and
invasion-related proteins, thereby leading to tumor development and
aggression of breast cancer cells.
Acknowledgements
This study was supported by the Natural Science
Foundation of Heilongjiang Province (QC2014C035).
References
|
1
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li Y, Drabsch Y, Pujuguet P, Ren J, van
Laar T, Zhang L, van Dam H, Clément-Lacroix P and Ten Dijke P:
Genetic depletion and pharmacological targeting of αv integrin in
breast cancer cells impairs metastasis in zebrafish and mouse
xenograft models. Breast Cancer Res. 17:282015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lu D, Zhou X, Yao L, Liu C, Jin F and Wu
Y: Clinical implications of the interleukin 27 serum level in
breast cancer. J Investig Med. 62:627–631. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Engelman JA: Targeting PI3K signalling in
cancer: opportunities, challenges and limitations. Nat Rev Cancer.
9:550–562. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu P, Cheng H, Roberts TM and Zhao JJ:
Targeting the phosphoinositide 3-kinase pathway in cancer. Nat Rev
Drug Discov. 8:627–644. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kobayashi T, Deak M, Morrice N and Cohen
P: Characterization of the structure and regulation of two novel
isoforms of serum- and glucocorticoid-induced protein kinase.
Biochem J. 344:189–197. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Vasudevan KM, Barbie DA, Davies MA,
Rabinovsky R, McNear CJ, Kim JJ, Hennessy BT, Tseng H, Pochanard P,
Kim SY, et al: AKT-independent signaling downstream of oncogenic
PIK3CA mutations in human cancer. Cancer Cell. 16:21–32. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang Y, Zhou D, Phung S, Masri S, Smith D
and Chen S: SGK3 is an estrogen-inducible kinase promoting
estrogen-mediated survival of breast cancer cells. Mol Endocrinol.
25:72–82. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shi L, Wang K, Zhao M, Yuan X and Huang C:
Overexpression of PIP5KL1 suppresses the growth of human cervical
cancer cells in vitro and in vivo. Cell Biol Int. 34:309–315. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu J, Wan M, He Q, Bassett RL Jr, Fu X,
Chen AC, Shi F, Creighton CJ, Schiff R, Huo L and Liu D: SGK3 is
associated with estrogen receptor expression in breast cancer.
Breast Cancer Res Treat. 134:531–541. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu M, Chen L, Chan TH, Wang J, Li Y, Li
Y, Zeng TT, Yuan YF and Guan XY: Serum and glucocorticoid kinase 3
at 8q13.1 promotes cell proliferation and survival in
hepatocellular carcinoma. Hepatology. 55:1754–1765. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang Y, Zhou D and Chen S: SGK3 is an
androgen-inducible kinase promoting prostate cancer cell
proliferation through activation of p70 S6 kinase and up-regulation
of cyclin D1. Mol Endocrinol. 28:935–948. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gatti L, Cossa G, Tinelli S, Carenini N,
Arrighetti N, Pennati M, Cominetti D, De Cesare M, Zunino F,
Zaffaroni N and Perego P: Improved apoptotic cell death in
drug-resistant non-small-cell lung cancer cells by tumor necrosis
factor-related apoptosis-inducing ligand-based treatment. J
Pharmacol Exp Ther. 348:360–371. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang G, Miyake M, Lawton A, Goodison S
and Rosser CJ: Matrix metalloproteinase-10 promotes tumor
progression through regulation of angiogenic and apoptotic pathways
in cervical tumors. BMC Cancer. 14:3102014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Virós D, Camacho M, Zarraonandia I, García
J, Quer M, Vila L and León X: Prognostic role of MMP-9 expression
in head and neck carcinoma patients treated with radiotherapy or
chemoradiotherapy. Oral Oncol. 49:322–325. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Thorsen SB, Christensen SL, Würtz SO,
Lundberg M, Nielsen BS, Vinther L, Knowles M, Gee N, Fredriksson S,
Møller S, et al: Plasma levels of the MMP-9: TIMP-1 complex as
prognostic biomarker in breast cancer: a retrospective study. BMC
Cancer. 13:5982013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang C, Ma HX, Jin MS, Zou YB, Teng YL,
Tian Z, Wang HY, Wang YP and Duan XM: Association of matrix
metalloproteinase (MMP)-2 and −9 expression with
extra-gastrointestinal stromal tumor metastasis. Asian Pac J Cancer
Prev. 15:4187–4192. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang B, Tang F, Zhang B, Zhao Y, Feng J
and Rao Z: Matrix metalloproteinase-9 overexpression is closely
related to poor prognosis in patients with colon cancer. World J
Surg Oncol. 12:242014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang YL, Chen CZ, Jin LP, Ji QQ, Chen YZ,
Li Q, Zhang XH and Qu JM: Effect and mechanism of the metastasis
suppressor gene BRMS1 on the migration of breast cancer cells. Int
J Clin Exp Med. 6:908–916. 2013.PubMed/NCBI
|
|
20
|
Wu Y, Jiang W, Wang Y, Wu J, Saiyin H,
Qiao X, Mei X, Guo B, Fang X, Zhang L, et al: Breast cancer
metastasis suppressor 1 regulates hepatocellular carcinoma cell
apoptosis via suppressing osteopontin expression. PLoS One.
7:e429762012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ventura BV, Quezada C, Maloney SC,
Fernandes BF, Antecka E, Martins C, Bakalian S, di Cesare S and
Burnier MN Jr: Expression of the metastasis suppressor BRMS1 in
uveal melanoma. Ecancermedicalscience. 8:4102014.PubMed/NCBI
|
|
22
|
Sheng XJ, Zhou YQ, Song QY, Zhou DM and
Liu QC: Loss of breast cancer metastasis suppressor 1 promotes
ovarian cancer cell metastasis by increasing chemokine receptor 4
expression. Oncol Rep. 27:1011–1018. 2012.PubMed/NCBI
|