Introduction
Periostin, also known as osteoblast-specific factor
2, is an 811 amino acid long protein that is secreted by
osteoblasts (1). The protein is a
member of the transforming growth factor (TGF)-β-inducible protein
superfamily and is induced in mesenchymal cells following
stimulation by TGF-β, bone morphogenetic protein 2, interleukin
(IL)-4 or IL-13 (2–4). Periostin serves a vital role in the
maintenance and development of tooth and bone tissue, in addition
to cardiac development and healing (2–4).
Potential factors stimulating the secretion of
periostin may also include platelet-derived growth factor-AA and
-BB, fibroblast growth factor (FGF)-A, -B and −1, and angiotensin
II (Ang II). Furthermore, FGF-1 and Ang II enhance periostin
expression following activation of the Ras/MEK1/2/extracellular
signal-regulated kinase (ERK)1/2, phosphatidylinositol 3-kinase
(PI3K)/Akt/p70S6K and Ras/p38 mitogen-activated protein kinase
signaling pathways (5,6). Periostin has become a key protein
investigated in cancer studies due to its vital function in
regulating cell matrix interactions and organization by binding to
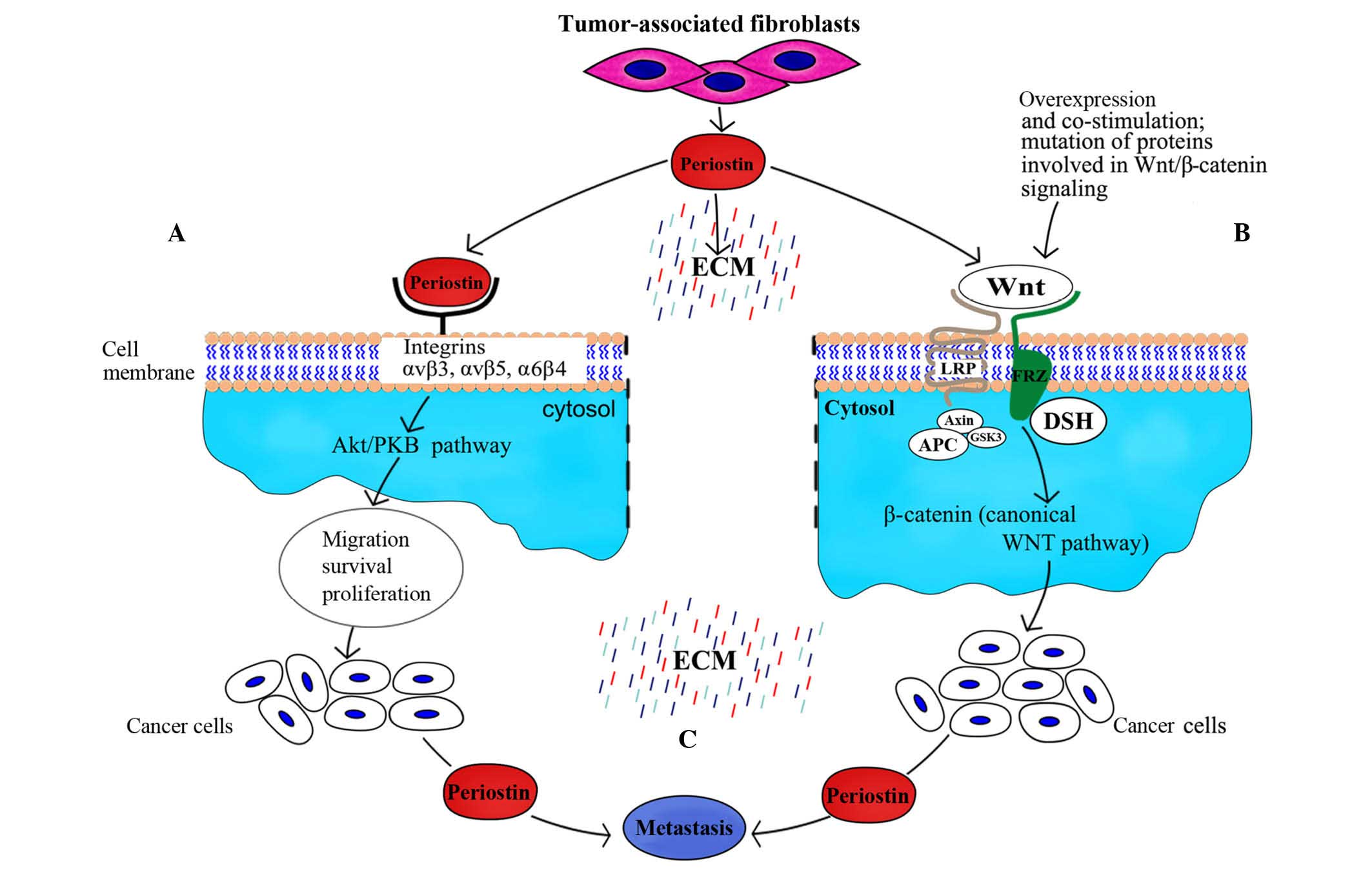
proteins such as collagen I/V, fibronectin and tenascin C (7). The protein also functions as a ligand
for integrins, including αvβ3, αvβ5 and α6β4 (8). Furthermore, periostin interacts with
various molecules to affect a number of pathways, such as Notch 1
signaling (9,10), and thus, a lack of periostin may be
responsible for a suppression of Notch 1 signaling (9,10).
Conversely, it cannot be excluded that the overexpression of
periostin may result in the activation of Notch 1 signaling in
cancer (10). Additionally, periostin
was identified as being involved in gastric cancer cell
epithelial-mesenchymal transition (EMT) (11). This study reviews the current
state-of-the-art role of periostin in gastrointestinal cancers.
Periostin in esophageal cancer
A previous study observed that periostin was present
in the normal esophagus, but its levels were significantly lower in
healthy esophagus tissues than in other areas of the
gastrointestinal tract, including the stomach and colorectum
(12). Meanwhile, periostin
expression was identified to be significantly higher in esophageal
squamous cell carcinoma (ESCC) compared with adjacent
non-neoplastic esophageal epithelium (13). mRNA profiling of ESCC tissues
demonstrated an upregulation of periostin as high as 11-fold
(13). Similarly, a quantitative
tissue proteomic study reported a 7-fold upregulation of periostin
in tumor tissues compared with normal tissues (14). However, a further study investigated
whether periostin was secreted by cancer cells or non-epithelial
cells, including cancer-associated fibroblasts, and it was observed
that it was primarily produced by the latter. At the same time,
periostin was reported to bind as a ligand to αvβ3 and αvβ5
integrins (Fig. 1), thus signaling
via the PI3K-Akt pathway within oesophageal adenocarcinoma cells;
this indicated that periostin accumulated to a higher degree in
ESCC tissue than in adjacent tissue outside the tumor (15).
Luo et al (16)
reported that the overexpression of periostin correlated with
adventitia invasion, lymph node metastasis and
tumor-node-metastasis (TNM) stage. In addition, it was also
observed that lymph node metastasis, adventitia invasion and TNM
stage were associated with positive expression of synuclein γ
(breast cancer-specific protein 1) (16), which may be responsible for altering
protease secretion and subsequently affecting the tumor
biology.
The majority of studies investigating ESCC have
focused on the association between periostin and vascular
endothelial growth factor (VEGF) and microvessel density (MVD) in
the development of cancer and prediction of patient prognosis
(17). Periostin-positive tumors
reportedly exhibit higher levels of VEGF and MVD compared with
periostin-negative tumors (17), with
a positive correlation identified between VEGF and periostin; these
results indicate that periostin serves a key role in ESCC
tumorigenesis through the induction and/or promotion of
angiogenesis (17). As high levels of
VEGF in serum is associated with a more advanced stage and poorer
response to chemoradiotherapy, VEGF may be viewed as a predictive
and prognostic factor of further tumor progression and shorter
patient survival (18).
Paramount genetic alterations associated with the
initiation and progression of ESCC include mutations underlying the
overexpression of p53 and epidermal growth factor receptor (EGFR)
(19). It was previously demonstrated
that culturing EPC2-hTERT-EGFR-p53R175H cells, which
overexpress EGFR and contain a p53 missense mutation, in a
three-dimensional organotypic culture induced the invasion of these
cells into the extracellular matrix (20). Michaylira et al (21) reported that the induction of
periostin, which was preferentially expressed in invading cells,
was dependent upon EGFR signaling and p53 mutation. The restoration
of wild-type p53 function was identified as a contributing factor
to the attenuation of periostin function in an organotypic culture
model of engineered ESCC (22).
Esophageal cells overexpressing periostin and mutant
p53R175H exhibited an upregulation in the signal
transducer and activator of transcription 1 (STAT1) network and
STAT-1 dependent genes (22). The
knockdown of STAT1 in invasive EPC-hTERT-p53R175H-POSTN
and EPC-hTERT-EFGR-P53 R175H cells resulted in reduced
invasiveness, while periostin-knockdown in ESCC tumor xenografts
led to a decrease in p53 expression, STAT1 activation and a
significant decrease in cluster of differentiation
(CD)44hiCD24low tumor-initiating cells
(22).
As periostin levels vary significantly between
healthy and diseased tissues, the protein may be used as a
distinguishing marker in the diagnosis and therapy of cancer to a
certain extent as confirmed by Kwon et al (23) in ESCC. The study focused on the
simultaneous involvement of aurora kinase B (AURKB) and periostin
and heat shock protein 47 (HSP47), which each exhibited a
significant increase at the mRNA and protein level (23). Notably, AURKB and HSP47 levels were
increased during early stages of cancer, whereas periostin levels
increased during cell-cell interactions between cancer and adjacent
stromal cells (23).
Periostin in gastric cancer
Normal and benign gastric tissues express periostin
at the highest level among the non-neoplastic tissues of other
organs of the gastrointestinal tract, with eventual impact on
normal epithelium physiology (12).
The effect of periostin is considerably more marked in gastric
cancer, and is reportedly overexpressed at the mRNA and protein
levels in comparison to adjacent notmal tissues (24). Kikuchi et al (25) identified that periostin expression was
increased in stage II–IV stomach cancer tissues. By contrast to
benign gastric tissues, with cancer progression, the linear
deposition of periostin disappeared in intestinal type gastric
tubular adenocarcinoma. Furthermore, in situ hybridization
documented the presence of periostin expression in fibroblasts, but
not in cancer cells (25). Periostin
promoted in vitro growth of OCUM-2MLN and OCUM-12 cancer
cell lines; this process was associated with ERK activation
(25). Finally, the study
demonstrated that in vivo mouse fibroblasts with positive
periostin expression were susceptible to tumor formation (25).
The varying deposition patterns of periostin in
cancer tissue bears testimony to its diverse biological role
depending on the source of its origin (26). A recent study by Lv et al
(26) demonstrated that the
restoration of periostin in gastric cancer cells inhibited their
growth and invasiveness. Furthermore, epithelial cell-derived
periostin had a positive effect on retinoblastoma protein (Rb)
phosphorylation, which induced the activation of the E2F1 target
gene p14ARF (26). This
gene was responsible for mouse double minute 2 homolog (Mdm2)
inactivation, which led to the stabilization of p53 and E-cadherin
proteins (26). Notably, under these
circumstances, Rb deletion resulted in the abolishment of the
aforementioned effects (26).
A further key role of periostin in gastric cancer is
the induction of angiogenesis (27).
MKN-45 cells under hypoxic conditions exhibited increased periostin
expression, which subsequently activated ERK1/2 and resulted in
VEGF-mediated angiogenesis (27).
When periostin was knocked down in these cells by specific small
interfering RNA, this effect was abolished (27).
Nicotine has been identified as an important factor
responsible for the induction of gastric cancer, with periostin
reported to increase cell invasion, proliferation and EMT in
nicotine-induced gastric cancer (11). Nicotine has also been demonstrated to
upregulate the expression of periostin in gastric cancer cells via
a cyclooxygenase-2-dependent pathway, and thus promotes the mitosis
and proliferation of gastric cells (11).
A key issue in chemotherapy is the sensitivity of
the cancer cells to the applied drugs (28). SGC-7901 human gastric cancer cells
proved susceptible to the apoptosis caused by cisplatin, but
overexpression of periostin made these cells more resistant to
cisplatin treatment. The same cells treated with 5-fluorouracil
(5-FU) behaved in a similar manner (28). Overexpression of periostin in the
SGC-7901 gastric cancer cell line increased resistance to 5-FU or
cisplatin-induced apoptosis and reduced the release of cytochrome
c from the mitochondria, resulting in diminished cleavage of
poly(ADP-ribose) polymerase-1 and caspase-3, and decreased
expression of p53 and Bax, in addition to increased Akt
phosphorylation and B-cell lymphoma 2 expression (28). It is important to note that the
interaction between the p53 and PI3K/Akt pathways is crucial for
apoptosis and survival of cancer cells. Furthermore, activation of
Akt may result in destabilization of p53 by Mdm2 (29).
Periostin in colorectal cancer
A relatively high level of periostin was present in
normal gastric and colon tissues in comparison to other regions of
the alimentary tract, which bears testimony to the role that it
plays in the normal physiology of the epithelium (12). Preoperative serum concentrations of
periostin were observed to be significantly higher in colorectal
cancer (CRC) compared with benign colorectal polyps or adenoma and
healthy controls (30). This, in
turn, functioned as a marker associated with the oncoming
metastasis and generally poor prognosis for the patients (30). Additionally, more advanced stages of
the disease were characterized by greater concentrations of
periostin (30). CRC tissues
exhibited upregulated periostin expression compared with normal
tissues, while no periostin mRNA expression was observed in 4 CRC
cell lines, including SW480, HT-29, LS174T and SW620 (30). Generally, lower postoperative
concentrations of periostin suggest that the increased levels were
produced by tumors. Ben et al (30) suggested that periostin may be produced
by the stromal cells surrounding the tumor, but not by the CRC
cells themselves.
In the normal colon, the highest level of periostin
expression is observed in the cells lining the colonic crypts,
whilst in CRC, periostin accumulates diffusely in the malignant
epithelium. In healthy tissues, mouse and human colonic depositions
of periostin are in close proximity to pericryptal fibroblasts, and
are also observed in numerous neoplastic tissues (31). Kikuchi et al (31) identified the connection between
periostin expression and colorectal crypt fibroblasts and
infiltrating fibroblasts of the tumor tissue. Periostin-mediated
induction of fibroblasts resulted in an increase in the number of
HCT 116 colon cancer cells in co-culture (31).
Regarding studies on periostin, a particularly
important observation was the expression and function of the
protein in CD133+ tumor cells (32). CD133 is a 120-kD transmembrane
glycoprotein, which has been recently employed as a cancer stem
cell marker in various types of carcinoma (32). Wu et al (32) observed that CD133+ CRC
tumor cells, which possess potent tumorigenicity responsible for
tumor initiation and maintenance, were characterized by
significantly higher periostin expression compared with
CD133− CRC tumor cells. Statistical analysis
demonstrated a linear correlation between the expression of
periostin and clinicopathological factors, including lymph node
metastasis, tumor size, histological type, postoperative liver
metastasis and TNM stage (32).
Periostin enhances cancer cell survival and promotes
their metastatic growth via the Akt/protein kinase B (PKB) pathway
(33). As a result, suppression of
the Akt/PKB pathway promotes apoptosis through Bad and caspase-9
activation (33). In addition, CRC
cell exposure to anti-periostin antibodies results in the
activation of apoptosis and potentiation of the effects of 5-FU
chemotherapy (33).
Xiao et al (34) reported that HT-29 and SW480 CRC cells
treated with 5-FU or oxaliplatin demonstrated greater levels of
periostin at the protein and mRNA levels. Notably, periostin was
responsible for inhibiting cell apoptosis, and periostin silencing
subsequently led to increased apoptosis and enhanced cleavage of
caspase-3 and poly(ADP-ribose) polymerase-1 in the CRC cells
following drug treatment (34).
Periostin silencing also resulted in the suppressed expression of
survivin (34). However, survivin, an
antiapoptotic protein present in CRC cells, repressed apoptosis
when expressed at low levels in SW480 and HT-29 cells with depleted
periostin levels (34). Additionally,
overexpression of periostin increased survivin expression and
phosphorylation of Akt, which was able to be reversed by
pretreatment with the specific inhibitor, LY294002 (34). Periostin interacts with numerous
factors at the onset of colon carcinoma, for example, periostin
contributes to the increase of Wnt signaling in cancer stem cells
by recruiting Wnt ligands, as shown in the experiments by Malanchi
et al (35). Furthermore, it
was demonstrated that periostin is required for cancer stem cell
maintenance and metastasis (35), and
participates in the recruitment of Wnt ligands, subsequently
increasing Wnt signaling in cancer stem cells (35).
Conclusion
Periostin promotes cancer growth, not only in
primary carcinomas of the esophagus, stomach and large bowel
(11,17,35), but
also in cancer of other organs, including prostate cancer (36). As neutralization of periostin
signaling induces cell apoptosis and enhances the cytotoxic effects
of anti-tumor agents in CRC, this protein has emerged as a marker
of poor prognosis and a target for anticancer therapy. Construction
of novel periostin signaling pathway-targeted drugs could affect
the aforemmentioned multiple molecular messengers. These molecular
targets should be selected with such modifications that this type
of treatment would not disturb the correct functions of healthy
cells. However, it is a great challenge to focus the therapeutic
property of the targeted drug to exclusively limit cancer growth in
the pharmacological regulation of periostin expression, as
periostin is a factor that is normally expressed in benign cells
and is involved in multiple interactions with numerous other
molecules.
References
|
1
|
Takeshita S, Kikuno R, Tezuka K and Amann
E: Osteoblast-specific factor 2: Cloning of a putative bone
adhesion protein with homology with the insect protein fasciclin I.
Biochem J. 294:271–278. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Horiuchi K, Amizuka N, Takeshita S,
Takamatsu H, Katsuura M, Ozawa H, Toyama Y, Bonewald LF and Kudo A:
Identification and characterization of a novel protein, periostin,
with restricted expression to periosteum and periodontal ligament
and increased expression by transforming growth factor beta. J Bone
Miner Res. 14:1239–1249. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Takayama G, Arima K, Kanaji T, Toda S,
Tanaka H, Shoji S, McKenzie AN, Nagai H, Hotokebuchi T and Izuhara
K: Periostin: A novel component of subepithelial fibrosis of
bronchial asthma downstream of IL-4 and IL-13 signals. J Allergy
Clin Immunol. 118:98–104. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ji X, Chen D, Xu C, Harris SE, Mundy GR
and Yoneda T: Patterns of gene expression associated with
BMP-2-induced osteoblast and adipocyte differentiation of
mesenchymal progenitor cell 3T3-F442A. J Bone Miner Metab.
18:132–139. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Erkan M, Kleeff J, Gorbachevski A, Reiser
C, Mitkus T, Esposito I, Giese T, Büchler MW, Giese NA and Friess
H: Periostin creates a tumor-supportive microenvironment in the
pancreas by sustaining fibrogenic stellate cell activity.
Gastroenterology. 132:1447–1464. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li P, Oparil S, Feng W and Chen YF:
Hypoxia-responsive growth factors upregulate periostin and
osteopontin expression via distinct signaling pathways in rat
pulmonary arterial smooth muscle cells. J Appl Physiol (1985).
97:1549–1558. 2004. View Article : Google Scholar
|
|
7
|
Norris RA, Damon B, Mironov V, Kasyanov V,
Ramamurthi A, Moreno-Rodriguez R, Trusk T, Potts JD, Goodwin RL,
Davis J, et al: Periostin regulates collagen fibrillogenesis and
the biomechanical properties of connective tissues. J Cell Biochem.
101:695–711. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gillan L, Matei D, Fishman DA, Gerbin CS,
Karlan BY and Chang DD: Periostin secreted by epithelial ovarian
carcinoma is a ligand for alpha(V)beta(3) and alpha(V)beta(5)
integrins and promotes cell motility. Cancer Res. 62:5358–5364.
2002.PubMed/NCBI
|
|
9
|
Tkatchenko TV, Moreno-Rodriguez RA, Conway
SJ, Molkentin JD, Markwald RR and Tkatchenko AV: Lack of periostin
leads to suppression of Notch1 signaling and calcific aortic valve
disease. Physiol Genomics. 39:160–168. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sriram R, Lo V, Pryce B, Antonova L, Mears
AJ, Daneshmand M, McKay B, Conway SJ, Muller WJ and Sabourin LA:
Loss of periostin/OSF-2 in ErbB2/Neu-driven tumors results in
androgen receptor-positive molecular apocrine-like tumors with
reduced Notch1 activity. Breast Cancer Res. 17:72015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu Y and Liu BA: Enhanced proliferation,
invasion, and epithelial-mesenchymal transition of
nicotine-promoted gastric cancer by periostin. World J
Gastroenterol. 17:2674–2680. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tai IT, Dai M and Chen LB: Periostin
induction in tumor cell line explants and inhibition of in vitro
cell growth by anti-periostin antibodies. Carcinogenesis.
26:908–915. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pawar H, Kashyap MK, Sahasrabuddhe NA,
Renuse S, Harsha HC, Kumar P, Sharma J, Kandasamy K, Marimuthu A,
Nair B, et al: Quantitative tissue proteomics of esophageal
squamous cell carcinoma for novel biomarker discovery. Cancer Biol
Ther. 12:510–522. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kashyap MK, Marimuthu A, Kishore CJ, Peri
S, Keerthikumar S, Prasad TS, Mahmood R, Rao S, Ranganathan P,
Sanjeeviah RC, et al: Genomewide mRNA profiling of esophageal
squamous cell carcinoma for identification of cancer biomarkers.
Cancer Biol Ther. 8:36–46. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Underwood TJ, Hayden AL, Derouet M, Garcia
E, Noble F, White MJ, Thirdborough S, Mead A, Clemons N, Mellone M,
et al: Cancer-associated fibroblasts predict poor outcome and
promote periostin-dependent invasion in oesophageal adenocarcinoma.
J Pathol. 235:466–477. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Luo JH, Zhou J and Gao Y: Correlation
between periostin and SNCG and esophageal cancer invasion,
infiltration and apoptosis. Asian Pac J Trop Med. 6:516–519. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang W, Sun QK, He YF, Ma DC, Xie MR, Ji
CS and Hu B: Overexpression of periostin is significantly
correlated to the tumor angiogenesis and poor prognosis in patients
with esophageal squamous cell carcinoma. Int J Clin Exp Pathol.
7:593–601. 2014.PubMed/NCBI
|
|
18
|
Cheng JC, Graber MS, Hsu FM, Tsai CL,
Castaneda L, Lee JM, Chang DT and Koong AC: High serum levels of
vascular endothelial growth factor-A and transforming growth
factor-β1 before neoadjuvant chemoradiotherapy predict poor
outcomes in patients with esophageal squamous cell carcinoma
receiving combined modality therapy. Ann Surg Oncol. 21:2361–2368.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shang L, Liu HJ, Hao JJ, Jiang YY, Shi F,
Zhang Y, Cai Y, Xu X, Jia XM, Zhan QM and Wang MR: A panel of
overexpressed proteins for prognosis in esophageal squamous cell
carcinoma. PLoS One. 9:e1110452014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Okawa T, Michaylira CZ, Kalabis J, Stairs
DB, Nakagawa H, Andl CD, Johnstone CN, Klein-Szanto AJ, El-Deiry
WS, Cukierman E, et al: The functional interplay between EGFR
overexpression, hTERT activation, and p53 mutation in esophageal
epithelial cells with activation of stromal fibroblasts induces
tumor development, invasion, and differentiation. Genes Dev.
21:2788–2803. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Michaylira CZ, Wong GS, Miller CG,
Gutierrez CM, Nakagawa H, Hammond R, Klein-Szanto AJ, Lee JS, Kim
SB, Herlyn M, et al: Periostin, a cell adhesion molecule,
facilitates invasion in the tumor microenvironment and annotates a
novel tumor-invasive signature in esophageal cancer. Cancer Res.
70:5281–5292. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wong GS, Lee JS, Park YY, Klein-Szanto AJ,
Waldron TJ, Cukierman E, Herlyn M, Gimotty P, Nakagawa H and Rustgi
AK: Periostin cooperates with mutant p53 to mediate invasion
through the induction of STAT1 signaling in the esophageal tumor
microenvironment. Oncogenesis. 2:e592013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kwon YJ, Lee SJ, Koh JS, Kim SH, Kim YJ
and Park JH: Expression patterns of aurora kinase B, heat shock
protein 47, and periostin in esophageal squamous cell carcinoma.
Oncol Res. 18:141–151. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li JS, Sun GW, Wei XY and Tang WH:
Expression of periostin and its clinicopathological relevance in
gastric cancer. World J Gastroenterol. 13:5261–5266. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kikuchi Y, Kunita A, Iwata C, Komura D,
Nishiyama T, Shimazu K, Takeshita K, Shibahara J, Kii I, Morishita
Y, et al: The niche component periostin is produced by
cancer-associated fibroblasts, supporting growth of gastric cancer
through ERK activation. Am J Pathol. 184:859–870. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lv H, Liu R, Fu J, Yang Q, Shi J, Chen P,
Ji M, Shi B and Hou P: Epithelial cell-derived periostin functions
as a tumor suppressor in gastric cancer through stabilizing p53 and
E-cadherin proteins via the Rb/E2F1/p14ARF/Mdm2 signaling pathway.
Cell Cycle. 13:2962–2974. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Qiu F, Shi CH, Zheng J and Liu YB:
Periostin mediates the increased pro-angiogenic activity of gastric
cancer cells under hypoxic conditions. J Biochem Mol Toxicol.
27:364–369. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li B, Wang L and Chi B: Upregulation of
periostin prevents P53-mediated apoptosis in SGC-7901 gastric
cancer cells. Mol Biol Rep. 40:1677–1683. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Haupt S, Berger M, Goldberg Z and Haupt Y:
Apoptosis - the p53 network. J Cell Sci. 116:4077–4085. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ben QW, Zhao Z, Ge SF, Zhou J, Yuan F and
Yuan YZ: Circulating levels of periostin may help identify patients
with more aggressive colorectal cancer. Int J Oncol. 34:821–828.
2009.PubMed/NCBI
|
|
31
|
Kikuchi Y, Kashima TG, Nishiyama T,
Shimazu K, Morishita Y, Shimazaki M, Kii I, Horie H, Nagai H, Kudo
A and Fukayama M: Periostin is expressed in pericryptal fibroblasts
and cancer-associated fibroblasts in the colon. J Histochem
Cytochem. 56:753–764. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wu G, Wang X and Zhang X: Clinical
implications of periostin in the liver metastasis of colorectal
cancer. Cancer Biother Radiopharm. 28:298–302. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bao S, Ouyang G, Bai X, Huang Z, Ma C, Liu
M, Shao R, Anderson RM, Rich JN and Wang XF: Periostin potently
promotes metastatic growth of colon cancer by augmenting cell
survival via the Akt/PKB pathway. Cancer Cell. 5:329–339. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xiao ZM, Wang XY and Wang AM: Periostin
induces chemoresistance in colon cancer cells through activation of
the PI3K/Akt/survivin pathway. Biotechnol Appl Biochem. 62:401–406.
2015. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Malanchi I, Santamaria-Martínez A, Susanto
E, Peng H, Lehr HA, Delaloye JF and Huelsken J: Interactions
between cancer stem cells and their niche govern metastatic
colonization. Nature. 481:85–89. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Argellati F, Nuzzo PV, Ricci F, Mangerini
R, Rubagotti A and Boccardo F: Dihydrotestosterone and bicalutamide
do not affect periostin expression in androgen-dependent LNCaP
prostate cancer cell lines. Anticancer Res. 33:815–820.
2013.PubMed/NCBI
|