Introduction
Epithelial ovarian cancer (EOC) is the leading cause
of mortality from gynecological malignancies. There are >21,290
cases annually in the United States, and 14,180 women can be
expected to succumb to the disease in 2015 (1). Due to the lack of effective tools for
early detection, the majority of patients with EOC are in an
advanced disease stage at diagnosis. The prognosis of EOC is
usually poor, with a 5-year survival rate at ~44% (1). Although there have been advancements in
surgery and chemotherapy treatment in previous years, the 5-year
survival rate for patients with EOC has not been improved
significantly in the last two decades (2). Therefore, it is necessary to identify
effective biomarkers that can aid diagnosis and therapy for
patients with EOC.
MicroRNAs (miRNAs/miRs) are a family of short
non-coding RNAs that negatively regulate gene expression at the
post-transcriptional level (3). By
regulating protein translation, miRNAs have emerged as powerful
regulators of a wide range of biological processes. Evidence is
emerging for the role of miRNAs in modulating cancer development.
One particular class of miRNAs has emerged to target survival
signaling pathways or signaling pathways that regulate apoptosis,
sensitivity and the cell cycle. This class includes miR-365 and
miR-29, which target cyclin D1 and apoptosis regulator Bcl-2
(Bcl-2) in colon cancer (4), and
collagen type Iα1 in ovarian cancer (5), respectively. Additionally, miR-19a and
miR-195-5p target Bcl-2 in gastric cancer (6).
miR-18a belongs to the miR-17-92 cluster, which
includes six miRNAs (miR-17, −20a, −18a, −19a, −19b and −92a) and
is located at the chromosome 13q31.1 region (7). Of the six miR-17-92 cluster members,
miR-19a and -b have been demonstrated to be key promoters of cancer
development and cancer cell proliferation (8,9).
Meanwhile, the role of miR-18a in cancer development remains
elusive. Several studies have observed that miR-18a is highly
expressed in several types of cancer, including nasopharyngeal
carcinoma (10), and prostate
(11) and gastric (12,13)
cancer. The miRNA promotes cancer development via inducing
proliferation and inhibiting apoptosis. Notably, studies have also
demonstrated that miR-18a can act as a tumor suppressor in breast
(14) and colorectal (15) cancer through inhibiting the expression
of hypoxia inducible factor 1α subunit and cell division cycle 42
(14,15). Nam et al (16) determined the expression of miR-18a in
advanced stages of ovarian cancer and observed that higher miR-18a
expression was significantly correlated with a poor prognosis.
To the best of our knowledge, studies have yet to
determine the expression and effects of miR-18a in EOC. In the
present study, series studies were performed to investigate the
effects of miR-18a on ovarian cancer growth and its associated
mechanism, and the potential predictive of miR-18a in patients with
EOC.
Materials and methods
Collection of tissue samples
Non-malignant (normal) and EOC tissues were obtained
with informed patient consent and ethical approval from the
Academic Medical Center Institutional Review Board of West China
Second University Hospital, Sichuan University (Chengdu, China).
The clinical tissues were obtained from the West China Second
University Hospital. Experiments described were performed on
samples obtained from the Academic Medical Center.
Cell culture and treatment
The ovarian cancer cell lines (A2780cp, A2780s,
SKOV3 and CAOV3), the human ovarian epithelium cell line (HOEC) and
the HEK293 cells were obtained from the American Type Culture
Collection (Manassas, VA, USA). The cells were cultured in
Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal
bovine serum (both Gibco; Thermo Fisher Scientific, Inc., Waltham,
MA, USA). All the cells were maintained in a humidified incubator
containing 5% CO2 at 37°C.
The lentivirus-based miR-18a expression system
(lenti-miR-18a) and the negative control (lenti-NC) were purchased
from GeneChem Co., Ltd. (Shanghai, China). Cell infection was
performed according to the manufacturer's instructions. Cells were
infected with lenti-miR-18a using a multiplicity of infection of
10. Puromycin (2 µg/ml) was added for selection at 72 h
post-infection. Cell transfection was performed using
FuGENE® HD transfection reagent (Roche Diagnostics,
Indianapolis, IN, USA) according to the manufacturer's
instructions.
Briefly, cells were seeded in 6-well plates at a
density of 2×105 cells/well and cultured for 24 h to
reach 70–80% confluence. Plasmids [2 µg; tumor protein
p53-regulated inhibitor of apoptosis gene 1 (TRIAP1)-3′untranslated
region (UTR), TRIAP1-3′UTR-mutant, IPMK-3′UTR and
IPMK-3′UTR-mutant] were diluted in 100 µl DMEM without serum. A
total of 5 µl FuGENE HD Transfection Reagent was added to the tubes
containing diluted DNA. The suspension was mixed and incubated for
15 min at room temperature, and subsequently added to the 6-well
plates.
Cell cycle assay
Cell cycle assays were performed by flow cytometry.
Briefly, the cells post-treatment were collected by centrifugation
for 3 min at 1,200 × g at 4°C and resuspended in 1 ml cold PBS.
Following fixation with 4 ml absolute ethanol, the cells were
centrifuged for 3 min at 1,200 × g at 4°C and resuspended in 1 ml
PBS. The cells were subsequently stained with 100 µl light
sensitive propidium iodide (1 mg/ml; Beyotime Institute of
Biotechnology, Beijing, China) and incubated for 5 min at room
temperature prior to flow cytometry. A total of 5×104
cells from each group were loaded and counted in each phase.
Bioinformatic analysis
The miRWalk database (www.ma.uni-heidelberg.de/apps/zmf/mirwalk) and
other programs, including miRanda (http://www.microrna.org/microrna/getGeneForm.do),
Sanger miRDB (http://www.mirdb.org/miRDB/) and Targetscan
(http://genes.mit.edu/tscan/targetscanS.html), were
used variously for target prediction. The online tool miRWalk
(version 2.0; www.umm.uni-heidelberg.de/apps/zmf/mirwalk/predictedmirnagene)
was used to predict potential target mRNAs of miR-18a. To verify
this bioinformatic analysis, the TRIAP1-3′UTR and the IPMK-3′UTR,
each of which contains a single miR-18a binding site, were cloned
downstream of the luciferase open reading frame. Meanwhile,
TRIAP1-3′UTR mutant and IPMK-3′UTR mutant, which contain mutated
miR-18a binding sites, were also introduced into the luciferase
constructs. The 293 cells were infected with lenti-miR-18a, and
puromycin was used to select the cells with stable expression. qPCR
analysis confirmed that miR-18a was upregulated in the cells (data
not shown). The luciferase constructs (TRIAP1-3′UTR, TRIAP1-3′UTR,
IPMK-3′UTR and IPMK-3′UTR mutant) were transfected separately into
cells that stably expressed miR-18a. Luciferase activity was
determined 4 to 6 h later.
Luciferase assays
The pMiRluc-TRIAP1-3′UTR and pMiRluc-IPMK-3′UTR
constructs containing miR-18a binding site were purchased from
Fulengen Biotechnology Co. (Guangzhou, China). The luciferase
reporter was co-transfected with lenti-NC (negative control) or
lenti-miR-18a using FuGENE HD (Roche Diagnostics). Luciferase
activity was measured 4 h after by luciferase reporter assay
(Promega Corporation, Madison, WI, USA), and the values were
normalized with activity of β-galactosidase.
Reverse transcription-qPCR
(RT-qPCR)
Total RNA was extracted from each experimental group
using TRIzol® reagent (Invitrogen Life Technologies;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
instructions. RNA concentration was assessed spectrophotometrically
at 260 nm (Thermo ND 2000; Thermo Fisher Scientific, Inc.
Wilmington, DE, USA). RT was performed using a RT kit (Takara Bio,
Inc., Otsu, Japan). PCR was subsequently performed using a Real
Time PCR kit (Takara Bio, Inc.) on a Bio-Rad CFX96 thermal cycler
(Bio-Rad Laboratories, Inc., Hercules, CA, USA), with U6 as an
internal control. PCR conditions were as follows: Denaturation at
94°C for 2 min; 30 cycles of amplification at 94°C for 30 sec,
annealing at 58°C for 30 sec, and extension at 72°C for 1 min. This
was followed by a terminal elongation step at 72°C for 10 min. The
Cq value of each PCR product was calculated, and the fold-change
was analyzed, according to the protocols of previous studies
(17). The primers for miR-18a and U6
were purchased from RiboBio Technology (Guangzhou, China; the
sequences were not supplied due to the rules of the company). All
experiments were performed in triplicate.
Cell viability detection assay
The Cell Counting Kit-8 (CCK-8) assay (Beyotime
Institute of Biotechnology) was performed to detect cell viability
according to the manufacturer's protocol. Briefly, 10 µl CCK-8 was
added into the cell culture medium and incubated at 37°C for 4 h.
Subsequently, absorbance was measured at 450 nm and each experiment
was performed three times.
Western blotting
Following treatment, cells were lysed on ice for 20
min with radioimmunoprecipitation assay buffer (Beyotime Institute
of Biotechnology). A total of 20 µg protein was separated by
SDS-PAGE on a 8–12% gel and electronically transferred onto a
polyvinylidene difluoride membranes (EMD Millipore, Billerica, MA,
USA). Following blocking at 4°C overnight with 5% no-fat milk in
TBT Tween 20, the membranes were incubated with the recommended
dilutions of primary antibodies against TRIAP1 (dilution, 1:800;
cat. no. LS-C346398-50; LifeSpan Biosciences, Inc., Seattle, WA,
USA), IPMK (dilution, 1:500; cat. no. TA337886; OriGene
Technologies, Inc., Beijing, China), AMP-activated protein kinase
(AMPK) (dilution, 1:1,000; cat. no. 2603), phosphorylated (p)-AMPK
(dilution: 1:800; cat. no. 2535) p53 (dilution, 1:2,000; cat. no.
2524), cleaved caspase 3 (CC3) (dilution, 1:500; cat. no. 9661),
Akt (dilution, 1:1,000; cat. no. 13038), p-Akt (dilution, 1:1,000;
cat. no. 4060), cyclin D1 (dilution, 1:800; cat. no. 2978; all Cell
Signaling Technology, Inc., Danvers, MA, USA) and
glyceraldehyde-3-phosphate dehydrogenase (dilution, 1:5,000; cat.
no. SC47724; Santa Cruz Biotechnology, Inc. Dallas, TX, USA) for 1
h at 37°C. The membranes were subsequently incubated with the
following secondary antibodies: Horseradish peroxidase
(HRP)-conjugated anti-rabbit Immunoglobulin G (IgG; dilution,
1:10,000; cat. no. ZB2301); and the HRP-conjugated anti-mouse IgG
antibody (dilution, 1:10,000; cat. no. ZB2305) both from OriGene
Technologies, Inc. at room temperature for 2 h. Peroxidase-labeled
bands were visualized using an enhanced chemiluminescence kit (EMD
Millipore), according to the manufacturer's protocol.
Terminal deoxynucleotidyl
transferase-deoxyuridine triphosphate nick end labeling (TUNEL)
assay
TUNEL assay was performed using a DeadEnd™
Fluorometric TUNEL system (Promega Corporation) to detect apoptotic
cells in A2780cp and A2780s cells and A2780cp tumor tissues,
according to the manufacturer's instructions. Cell nuclei with
green fluorescent staining were defined as TUNEL-positive and
visualized using a fluorescence microscope (DTX500; Nikon
Corporation, Tokyo, Japan). For quantification of the
TUNEL-positive cells, the number of green fluorescent cells was
counted in randomly selected fields (magnification, ×200). The cell
nuclei were subsequently counter-stained with
4′,6-diamidino-2-phenylindole (Beyotime Institute of
Biotechnology).
Animal study
To establish the A2780cp intraperitoneal cancer
model, A2780cp cells (8×106) infected with lenti-miR-18a
and lenti-NC (negative control), were injected into the
intraperitoneal space of BALB/c nude female mice (6–8 weeks old;
~20 g). A total of 6 mice per group were used. The mice were
obtained from the Animal Center of Sichuan University and housed at
26°C, with a 12-h light/dark cycle and ad libitum access to food
and water. At 30 days after tumor cell injection, the mice were
anesthetized using diethyl ether (100 mg/kg; Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) and sacrificed. The dissected tumors were
weighed. The tumors and viscera were examined grossly and
microscopically stained with hematoxylin and eosin. Animal studies
were performed with approval from with the Institutional Animal
Care and Treatment Committee of Sichuan University.
Immunostaining of proliferating cell
nuclear antigen (PCNA)
Expression of PCNA was analyzed using mouse
anti-human PCNA antibody (cat. no. I0710; Santa Cruz Biotechnology,
Inc.). Paraffin-embedded tumor sections (3–5 µm) were mounted on
3-aminopropyl triethoxysilane-coated glass slides. Sections were
deparaffinized in xylene, treated with a graded series of alcohol
[100, 95 and 80% ethanol/double-distilled H2O (v/v)] and
rehydrated in PBS (pH 7.4). Antigen retrieval was performed by
heating for 3 min in a pressure cooker with 0.1 mol/l citrate
buffer (pH 6.0). Endogenous peroxide was blocked with 3%
H2O2 for 10 min. Following PBS washes, slides
were blocked with 5% normal goat serum (Beyotime Institute of
Biotechnology) in PBS for 15 min at room temperature and incubated
with primary anti-PCNA antibody (1:200) in blocking solution
overnight at 4°C. All slides were subsequently incubated with
biotin-conjugated goat anti-mouse or goat anti-rat secondary
antibody (both dilution 1:200; cat. nos. SP9001 and SP9002,
respectively; OriGene Technologies, Inc.) for 15 min at 37°C and
with streptavidin-biotin complex at 37°C for 15 min. The
immunoreaction was visualized using diaminobenzidine peroxide
solution and the nuclei were counterstained with hematoxylin. All
specimens were observed using Olympus BX600 microscopes and SPOT
Fiex camera (both Olympus Corporation, Tokyo, Japan). The total
number of cells and the number of PCNA-positive cells were counted,
and the percentage of PCNA-positive cells was calculated.
Statistical analysis
All data were analyzed using one-way analysis of
variance. Statistical analyses were performed using SPSS Statistics
version 19.0 (IBM SPSS, Armonk, NY, USA). Values are expressed as
mean ± standard error. P<0.05 was considered to indicate a
statistically significant difference.
Results
miR-18a is downregulated in malignant
ovarian tissues and ovarian cancer cells
To determine the expression level of miR-18a in
ovarian malignant and normal tissues, malignant ovarian and
non-malignant (normal) tissues were collected and used for miR-18a
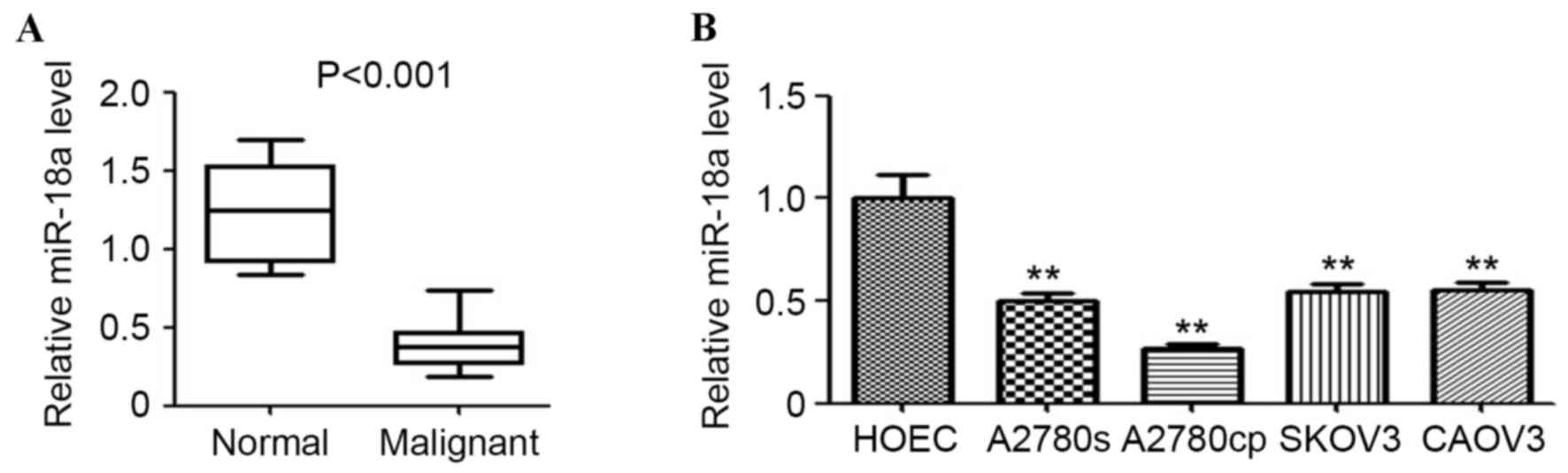
detection by qPCR. As shown in Fig.
1A, miR-18a expression in ovarian malignant tissues was
significantly downregulated compared with the normal ovarian
tissues (P<0.001). Furthermore, the expression of miR-18a in
several ovarian cancer cell lines and the normal ovarian cell line
was determined. The results indicated that miR-18a expression was
significantly lower in the ovarian cancer cell lines (A2780cp,
A2780s, SKOV3 and CAOV3) compared with the non-malignant HOEC cell
line (all P<0.01; Fig. 1B).
miR-18a significantly inhibits
proliferation and induces apoptosis of ovarian cancer cells in
vitro
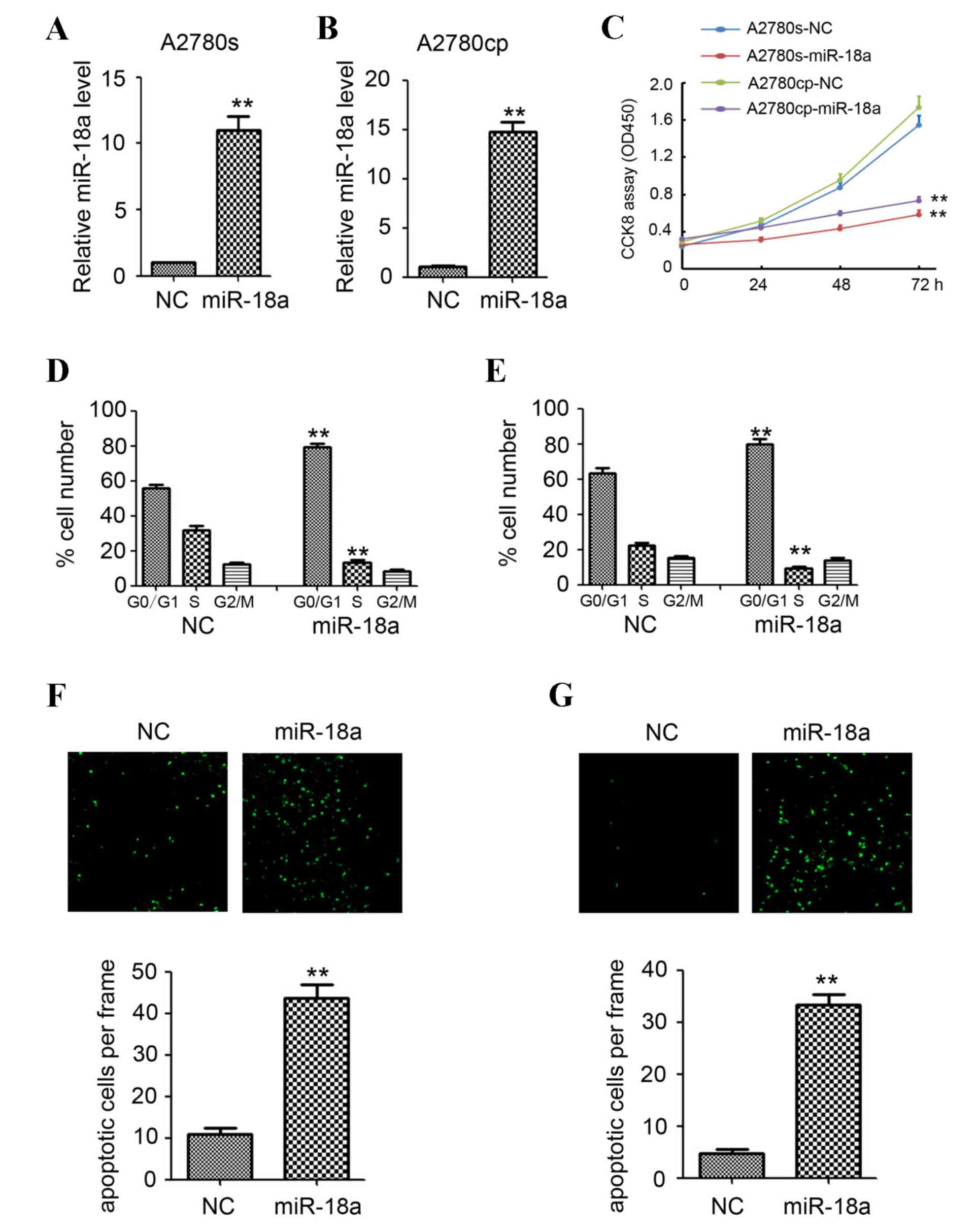
A lentivirus-based miR-18a expression system was
established to overexpress miR-18a in two ovarian cancer cell lines
(A2780cp and A2780s) to investigate the potential involvement of
miR-18a in inhibiting the development of ovarian cancer. As shown
in Fig. 2A and B, qPCR confirmed that
miR-18a expression was significantly upregulated when miR-18a was
overexpressed in A2780cp and A2780s cells compared with the NC
(P<0.01). Analysis of cell viability by CCK-8 assay indicated
that overexpression of miR-18a significantly inhibited cell growth
in the ovarian cancer cell lines (A2780s and A2780cp) compared with
the NC (P<0.01; Fig. 2C).
Flow cytometric analysis of the cell cycle indicated
a significant reduction in the number of cells in the S-phase and a
significant increase in the G0/G1 phase when miR-18a was
overexpressed in the A2780cp and A2780s cells compared with the NC
(P<0.01; Fig. 2D and E). However,
no significant differences were observed in the G2/M phase between
the two groups (Fig. 2D and E).
Analysis of cell apoptosis by TUNEL assay indicated significant
increases in the number of apoptotic cells when miR-18a was
overexpressed in the A2780cp and A2780s cells (P<0.01; Fig. 2F and G).
miR-18a directly targets TRIAP1 and
IPMK
In order to investigate the direct targets of
miR-18a involved in cancer cell proliferation and apoptosis in
human ovarian cells, a large number of potential target proteins in
a database library were screened to identify potential microRNA
binding seed sequences within the 3′-UTR. Bioinformatic analysis
(Targetscan micro, RNA.org, microRNASeq) identified TRIAP1
and IPMK as candidate targets (Fig. 3A
and B).
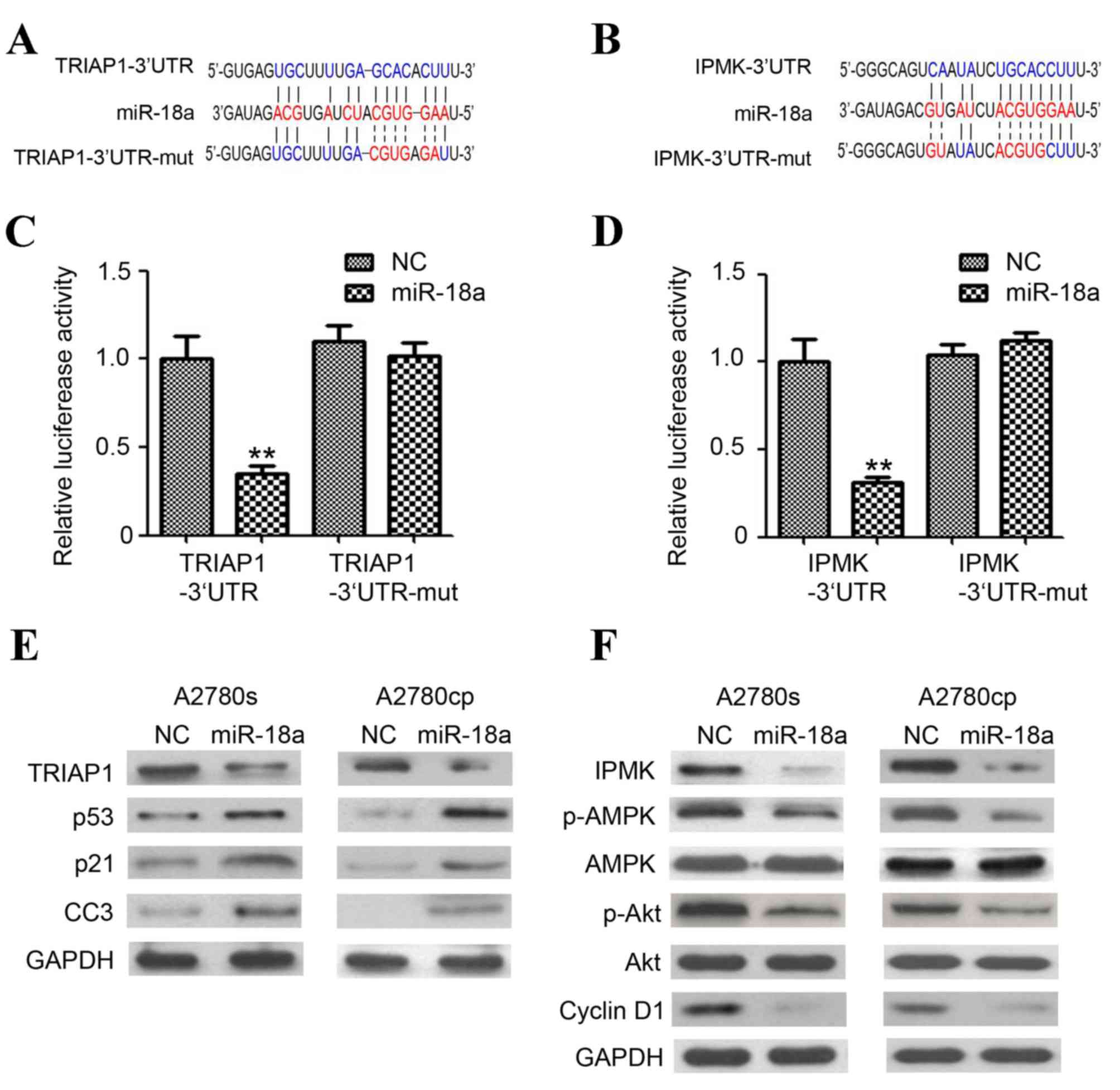 | Figure 3.TRIAP1 and IPMK are the targets of
miR-18a. (A) Predicted duplex formations between TRIAP1-3′UTR and
miR-18a. TRIAP1-3′UTR contains a single predicted miR-18a binding
site. (B) Predicted duplex formations between IPMK-3′UTR and
miR-18a. IPMK-3′UTR contains one single predicted miR-18a binding
site. Sites of target mutagenesis are indicated in red. (C and D)
Relative luciferase reporter expression was normalized to NC. Data
are depicted as mean ± standard error. **P<0.01 vs. lenti-NC
group. (E) Western blot analysis of TRIAP1, p53, p21 and CC3 and
GAPDH in A2780s and A780cp cells following infection with
lenti-miR-18a and lenti-NC (F) Western blot analysis of IPMK,
(p)-adenosine monophosphate (AMP)-activated protein kinase, p-Akt,
Akt and cyclin D1 and GAPDH in A2780s and A780cp cells following
infection with lenti-miR-18a and lenti-NC. CCS, cleaved caspase 3;
GAPDH, glyceraldehyde-3-phosphate dehydrogenase; IPMK, inositol
phosphate multikinase; miR/miRNA, microRNA; NC, negative control;
p-AMPK, phosphorylated-adenosine monophosphate-activated protein
kinase; TRIAP1, tumor protein p53 (TP53)-regulated inhibitor of
apoptosis gene 1; UTR, untranslated region. |
Luciferase expression in the cells overexpressing
miR-18a and also expressing the TRIAP1-3′UTR construct was
significantly reduced. The same finding was observed for the cells
overexpressing miR-18a and also expressing the IPMK-3′UTR construct
(P<0.01; Fig. 3C and D). No
significant reduction in luciferase expression of cells transfected
with miR binding site mutant plasmids was observed (Fig. 3C and D).
Western blotting was subsequently employed to
determine the expression of TRIAP1 and IPMK proteins, and their
downstream targets. Overexpression of miR-18a in A2780cp and A2780s
cells markedly inhibited TRIAP1 and IPMK expression (Fig. 3E and F). The expression of cell
cycle-associated proteins p-Akt, p-AMPK and cyclin D1 was also
downregulated by miR-18a, whereas no significant reduction of Akt
and AMPK expression was observed. Furthermore, miR-18a also
significantly induced the expression of the apoptosis-associated
protein p53, p21 and cleaved caspase 3 in the A2780cp and A2780s
cells (Fig. 3E). Collectively, the
results indicated that miR-18a directly targets TRIAP1 and IPMK,
and hence regulates the expression of the downstream targets.
miR-18a inhibits ovarian tumor growth
in vivo
To further elucidate the effects of miR-18a on
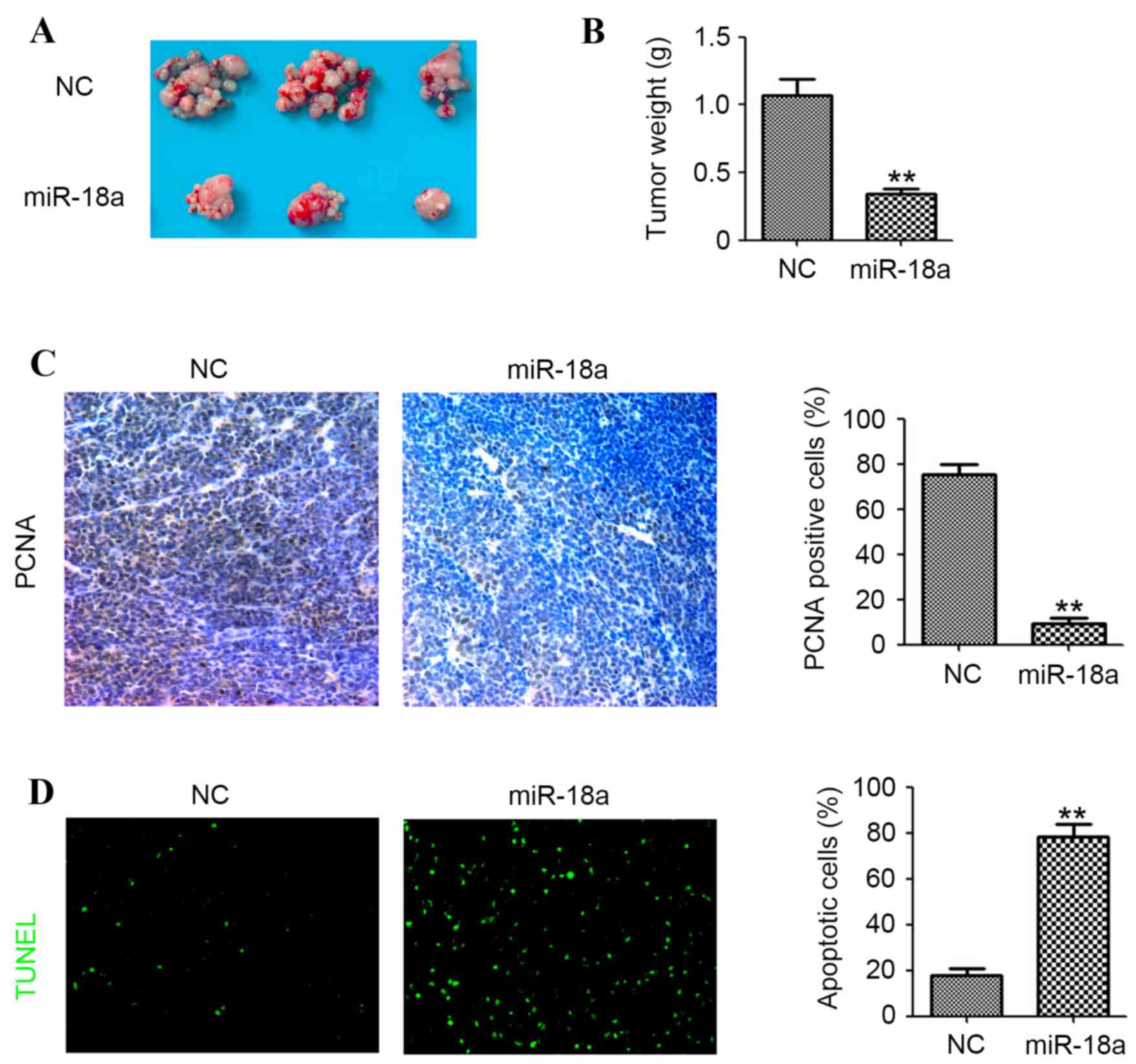
ovarian tumor growth in vivo, the A2780cp intraperitoneal
mouse model was employed. The tumor was collected (Fig. 4A) and the weight was measured 30 days
after the A2780cp injection. The mean tumor weight was 1.05±0.11
and 0.37±0.04 g in the NC and miR-18a group (P<0.01; Fig. 4B). Furthermore, the proliferative and
apoptotic activity of the A2780cp tumors were determined by PCNA
staining and TUNEL assay. When compared with the negative controls,
lower numbers of PCNA-positive cells and higher numbers of
apoptotic cells were found in the miR-18a group (P<0.01;
Fig. 4C and D). Together, the results
in the present study indicated that miR-18a may inhibit A2780cp
intraperitoneal tumor growth in vivo by inhibiting
proliferation and inducing apoptosis.
Discussion
An initial analysis of the expression and effects of
miR-18a in EOC indicated that the expression of miR-18a was lower
in EOC tissue and all ovarian cancer cell lines investigated
compared with non-malignant (normal) ovarian tissues and the HOEC
cell line. Thus, we hypothesize that miR-18a may act as a tumor
suppressor in the development of EOC. The lentivirus-based miR-18a
expression system was employed and used to infect ovarian cancer
cell lines (A2780s and A2780cp). Overexpression of miR-18a
significantly induced cell cycle arrest and apoptosis.
Bioinformatic and luciferase reporter gene assay demonstrated that
miR-18a directly targeted TRIAP1 and IPMK, and hence regulated the
expression of downstream targets. Furthermore, in vivo study
and staining indicated that miR-18a may significantly inhibit
A2780cp intraperitoneal tumor growth by inhibiting proliferation
and inducing apoptosis.
miR-18a is encoded by the miR-17-92a-1 cluster host
gene (MIR17HG) gene, which is one of the most frequently
deregulated genes in human cancer (7). MIR17HG was originally identified as a
tumor promoter in the heart, thyroid and lungs (18). However, emerging evidence suggests
that the loss of MIR17HG may contribute to the development and
progression of other types of cancer, thereby implicating a tumor
suppressor function (19).
MIR17HG is able to inhibit the proliferation of
luminal breast cancer cells by targeting a steroid receptor
co-activator (nuclear receptor coactivator 3), cyclin D1 and
estrogen receptor 1 (20–23). Previous studies have demonstrated that
the tertiary structure of the folded primary-miR-17-92 transcript
may contribute to less efficient processing of miR-18a (24,25). The
miR-17-92 cluster has been reported to correlate with the poor
prognosis of patients with a genetic etiology of ovarian cancer
(26). Therefore, miR-18a may serve a
homeostatic role in regulating the oncogenic effect of the entire
miR-17-92 cluster. The present study demonstrated a lower
expression of miR-18a in EOC patient tissues and human ovarian
cancer cell lines. Functional study indicated that miR-18a may
inhibit A2780s and A2780cp tumor growth in vitro and in
vivo by inhibiting proliferation and inducing apoptosis.
p53 cell survival factor (TRIAP1; p53CSV) contains a
p53 binding site within its coding sequence and is upregulated in
multiple myeloma (27). TRIAP1 has
been demonstrated to suppress apoptosis through interacting with
cytoplasmic heat shock protein family A (Hsp70) member 1A and
apoptotic peptidase activating factor 1 (18), and inhibiting the expression of p21
(28). Furthermore, TRIAP1 interact
with PRELI domain-containing protein 1, mitochondrial (PRELI) to
form the TRIAP1/PRELI complex, which inhibits apoptosis by
promoting cardiolipin accumulation and link p53-mediated cell
survival to mitochondrial cardiolipin (29). In the present study, it was
demonstrated that TRIAP1 was the direct target of miR-18a by
luciferase assay. Additionally, western blotting results indicated
that TRIAP1 expression was inhibited by miR-18a. The downstream
targets of TRIAP1, including p53, p21 and CC3 also increased
following the overexpression of miR-18a. These results provide
evidence that miR-18a induces ovarian cancer apoptosis through
directly targeting TRIAP1 and hence regulating apoptosis-related
protein expression.
IPMK is a key regulatory enzyme in inositol
phosphate disposition and is also a member of the inositol
phosphokinase 6-kinase family of enzymes, but is not primarily
associated with the formation of inositol pyrophosphates. IPMK has
been shown to act as an activator of Akt, AMPK and the mechanistic
target of rapamycin pathway (30–33).
Several studies have suggested that IPMK inhibitors may be
beneficial in treating obesity, diabetes mellitus and cell
proliferation-associated diseases (31,33,34). In
the present study, it was demonstrated that IPMK was the direct
target of miR-18a, and the expression of IPMK was significantly
inhibited by miR-18a in A2780cp and A2780s ovarian cancer cells.
The downstream targets of IMPK, including p-Akt, p-AMPK and cyclin
D1, were also regulated by miR-18a. These results demonstrated that
miR-18a inhibits ovarian cancer cell proliferation by directly
targeting IPMK and hence is able to regulate cell
proliferation-associated protein expression.
Collectively, the present study examined the
expression of miR-18a in EOC and the potential role of miR-18a in
modulating ovarian cancer growth. miR-18a detection by qPCR
indicated that miR-18a was reduced in EOC tissues and all ovarian
cancer cell lines investigated. Functional study demonstrated the
tumor suppressor role of miR-18a in ovarian cancer development.
Further study of the mechanism indicated that miR-18a directly
targets TRIAP1 and IPMK, hence inducing cell cycle arrest and
apoptosis. The present study may provide a novel diagnostic method
and be a therapeutic target for EOC.
Acknowledgements
The present study was supported by the National Key
Basic Research Program (973 Program) of China (grant nos.
2010CB529905 and 2011CB910703).
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Coleman MP, Forman D, Bryant H, Butler J,
Rachet B, Maringe C, Nur U, Tracey E, Coory M, Hatcher J, et al
Cancer survival in Australia, Canada, Denmark, Norway, Sweden, and
the UK, 1995–2007 (the International Cancer Benchmarking
Partnership), : An analysis of population-based cancer registry
data. Lancet. 377:127–138. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nie J, Liu L, Zheng W, Chen L, Wu X, Xu Y,
Du X and Han W: microRNA-365, down-regulated in colon cancer,
inhibits cell cycle progression and promotes apoptosis of colon
cancer cells by probably targeting Cyclin D1 and Bcl-2.
Carcinogenesis. 33:220–225. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yu PN, Yan MD, Lai HC, Huang RL, Chou YC,
Lin WC, Yeh LT and Lin YW: Downregulation of miR-29 contributes to
cisplatin resistance of ovarian cancer cells. Int J Cancer.
134:542–551. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xia T, Liao Q, Jiang X, Shao Y, Xiao B, Xi
Y and Guo J: Long noncoding RNA associated-competing endogenous
RNAs in gastric cancer. Sci Rep. 4:60882014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Volinia S, Calin GA, Liu CG, Ambs S,
Cimmino A, Petrocca F, Visone R, Iorio M, Roldo C, Ferracin M, et
al: A microRNA expression signature of human solid tumors defines
cancer gene targets. Proc Natl Acad Sci USA. 103:2257–2261. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mu P, Han YC, Betel D, Yao E, Squatrito M,
Ogrodowski P, de Stanchina E, D'Andrea A, Sander C and Ventura A:
Genetic dissection of the miR-17~92 cluster of microRNAs in
Myc-induced B-cell lymphomas. Genes Dev. 23:2806–2811. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Olive V, Bennett MJ, Walker JC, Ma C,
Jiang I, Cordon-Cardo C, Li QJ, Lowe SW, Hannon GJ and He L: miR-19
is a key oncogenic component of mir-17-92. Genes Dev. 23:2839–2849.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Luo Z, Dai Y, Zhang L, Jiang C, Li Z, Yang
J, McCarthy JB, She X, Zhang W, Ma J, et al: miR-18a promotes
malignant progression by impairing microRNA biogenesis in
nasopharyngeal carcinoma. Carcinogenesis. 34:415–425. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hsu TI, Hsu CH, Lee KH, Lin JT, Chen CS,
Chang KC, Su CY, Hsiao M and Lu PJ: MicroRNA-18a is elevated in
prostate cancer and promotes tumorigenesis through suppressing STK4
in vitro and in vivo. Oncogenesis. 3:e992014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Su ZX, Zhao J, Rong ZH, Wu YG, Geng WM and
Qin CK: Diagnostic and prognostic value of circulating miR-18a in
the plasma of patients with gastric cancer. Tumour Biol.
35:12119–12125. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu W, Takanashi M, Borjigin N, Ohno SI,
Fujita K, Hoshino S, Osaka Y, Tsuchida A and Kuroda M: MicroRNA-18a
modulates STAT3 activity through negative regulation of PIAS3
during gastric adenocarcinogenesis. Br J Cancer. 108:653–661. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Krutilina R, Sun W, Sethuraman A, Brown M,
Seagroves TN, Pfeffer LM, Ignatova T and Fan M: MicroRNA-18a
inhibits hypoxia-inducible factor 1α activity and lung metastasis
in basal breast cancers. Breast Cancer Res. 16:R782014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Humphreys KJ, McKinnon RA and Michael MZ:
miR-18a inhibits CDC42 and plays a tumour suppressor role in
colorectal cancer cells. PLoS One. 9:e1122882014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nam EJ, Yoon H, Kim SW, Kim H, Kim YT, Kim
JH, Kim JW and Kim S: MicroRNA expression profiles in serous
ovarian carcinoma. Clin Cancer Res. 14:2690–2695. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dai L, Cui X, Zhang X, Cheng L, Liu Y,
Yang Y, Fan P, Wang Q, Lin Y, Zhang J, et al: SARI inhibits
angiogenesis and tumour growth of human colon cancer through
directly targeting ceruloplasmin. Nat Commun. 7:119962016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mendell JT: miRiad roles for the miR-17-92
cluster in development and disease. Cell. 133:217–222. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Eiriksdottir G, Johannesdottir G,
Ingvarsson S, Björnsdottir IB, Jonasson JG, Agnarsson BA,
Hallgrimsson J, Gudmundsson J, Egilsson V, Sigurdsson H and
Barkardottir RB: Mapping loss of heterozygosity at chromosome 13q:
loss at 13q12-q13 is associated with breast tumour progression and
poor prognosis. Eur J Cancer. 34:2076–2081. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yu Z, Wang C, Wang M, Li Z, Casimiro MC,
Liu M, Wu K, Whittle J, Ju X, Hyslop T, et al: A cyclin D1/microRNA
17/20 regulatory feedback loop in control of breast cancer cell
proliferation. J Cell Biol. 182:509–517. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hossain A, Kuo MT and Saunders GF:
Mir-17-5p regulates breast cancer cell proliferation by inhibiting
translation of AIB1 mRNA. Mol Cell Biol. 26:8191–8201. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Castellano L, Giamas G, Jacob J, Coombes
RC, Lucchesi W, Thiruchelvam P, Barton G, Jiao LR, Wait R, Waxman
J, et al: The estrogen receptor-alpha-induced microRNA signature
regulates itself and its transcriptional response. Proc Natl Acad
Sci USA. 106:15732–15737. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yu Z, Willmarth NE, Zhou J, Katiyar S,
Wang M, Liu Y, McCue PA, Quong AA, Lisanti MP and Pestell RG:
microRNA 17/20 inhibits cellular invasion and tumor metastasis in
breast cancer by heterotypic signaling. Proc Natl Acad Sci USA.
107:8231–8236. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chakraborty S, Mehtab S, Patwardhan A and
Krishnan Y: Pri-miR-17-92a transcript folds into a tertiary
structure and autoregulates its processing. RNA. 18:1014–1028.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chaulk SG, Thede GL, Kent OA, Xu Z, Gesner
EM, Veldhoen RA, Khanna SK, Goping IS, MacMillan AM, Mendell JT, et
al: Role of pri-miRNA tertiary structure in miR-17~92 miRNA
biogenesis. RNA Biol. 8:1105–1114. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shen J, Wang D, Gregory SR, Medico L, Hu
Q, Yan L, Odunsi K, Lele SB, Ambrosone CB, Liu S and Zhao H:
Evaluation of microRNA expression profiles and their associations
with risk alleles in lymphoblastoid cell lines of familial ovarian
cancer. Carcinogenesis. 33:604–612. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Park WR and Nakamura Y: p53CSV, a novel
p53-inducible gene involved in the p53-dependent cell-survival
pathway. Cancer Res. 65:1197–1206. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Andrysik Z, Kim J, Tan AC and Espinosa JM:
A genetic screen identifies TCF3/E2A and TRIAP1 as pathway-specific
regulators of the cellular response to p53 activation. Cell Rep.
3:1346–1354. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Potting C, Tatsuta T, König T, Haag M, Wai
T, Aaltonen MJ and Langer T: TRIAP1/PRELI complexes prevent
apoptosis by mediating intramitochondrial transport of phosphatidic
acid. Cell Metab. 18:287–295. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dailey MJ and Kim S: Inositol
polyphosphate multikinase: An emerging player for the central
action of AMP-activated protein kinase. Biochem Biophys Res Commun.
421:1–3. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kim S, Kim SF, Maag D, Maxwell MJ, Resnick
AC, Juluri KR, Chakraborty A, Koldobskiy MA, Cha SH, Barrow R, et
al: Amino acid signaling to mTOR mediated by inositol polyphosphate
multikinase. Cell Metab. 13:215–221. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Roppenser B, Kwon H, Canadien V, Xu R,
Devreotes PN, Grinstein S and Brumell JH: Multiple host kinases
contribute to Akt activation during Salmonella infection. PLoS One.
8:e710152013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Maag D, Maxwell MJ, Hardesty DA, Boucher
KL, Choudhari N, Hanno AG, Ma JF, Snowman AS, Pietropaoli JW, Xu R,
et al: Inositol polyphosphate multikinase is a physiologic
PI3-kinase that activates Akt/PKB. Proc Natl Acad Sci USA.
108:1391–1396. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bang S, Kim S, Dailey MJ, Chen Y, Moran
TH, Snyder SH and Kim SF: AMP-activated protein kinase is
physiologically regulated by inositol polyphosphate multikinase.
Proc Natl Acad Sci USA. 109:616–620. 2012. View Article : Google Scholar : PubMed/NCBI
|