Epithelial-mesenchymal transition (EMT) is a crucial
process that promotes cell motility, wound healing, tissue
regeneration, fibrogenesis and tumor metastasis (1). EMT has been reported to be implicated in
multiple steps of several developmental processes involved in tumor
progression (2). It can lead to a
loss of cell-cell junctions in tumor cells and a decrease in the
expression of E-cadherin in the epithelium (3). EMT can also lead to an increase in
expression of vimentin (4), a marker
of mesenchymal-derived cells. Notably, EMT facilitates cancer cells
to initiate distant metastasis and is able to increase motility of
cancer cells at the leading tumor edge and cell invasion (5). EMT is an important process in cancer
cell migration and invasion. Thus, manipulating the EMT process
in vivo may be a useful strategy to prevent tumor
metastasis. Notably, the cytokine interleukin (IL-8) is important
for EMT, and it is highly expressed in the cancer microenvironment
(6). Although IL-8 has a
pro-inflammatory role, cancer cells are able to evade host immune
defense mechanisms (7). The chemokine
IL-8 is secreted by fibroblasts, endothelial and immune cells. IL-8
expression is closely associated with cancer (6). Furthermore, the involvement of IL-8 in
angiogenesis (8), and cancer cell
invasion and metastasis has been previously reported (9). A previous study has indicated that in
cancer patients, increased expression of IL-8 in tumor tissues may
be associated with EMT (10).
It has been previously demonstrated that EMT in
tumors can be induced by the secretion of specific factors,
including IL-6, IL-8, vascular endothelial growth factor (VEGF),
transforming growth factor β (TGFβ), SNAIL, matrix metalloprotease
(MMP), tumor necrosis factor α (TNFα) and TWIST (2,11–14). The secretion of pro-inflammatory
cytokines (TNFα and IL-6), chemokine IL-8 and growth factors (TGFβ
and VEGF) has also been reported in A549 cells, and may have
important associations with cancer (2). IL-8 is able to promote cell motility,
cancer metastasis and cell invasion (15) following EMT. Tumor cells are able to
secrete IL-8 via an autocrine mechanism, which can promote EMT. A
previous study revealed that knockdown of IL-8 suppressed the level
of phosphorylated AKT in S18 cells (4). IL-8 knockdown may lead to upregulation
of the epithelial marker E-cadherin as well as downregulation of
the mesenchymal markers vimentin and fibronectin (4). Additionally, IL-8 has been closely
associated with EMT (16) and may
promote tumor metastasis and cell invasion.
Cancer cells undergo a reversal of EMT, termed
mesenchymal-epithelial transition (MET) (17), to invade multiple organs until they
migrate to their final destination for colonization. In contrast to
EMT, MET is associated with colonization at the metastatic site
(18). Once mesenchymal cells reach
their destination, the cell phenotype changes to an epithelial
phenotype via MET to colonize the organs (19). These changes involve the loss of
cell-cell junctions, and cells acquire motility and invasive
capabilities (3). Cancer cells must
undergo the MET process to migrate to their destination.
Furthermore, MET is closely associated with cancer cells, which may
acquire a second colonization (otherwise termed metastasis).
A vital characteristic of cancer metastasis is
induction of EMT, which is associated with interaction with the
extracellular matrix (20). In the
extracellular matrix, the role of the cellular factors is to
communicate with the intracellular matrix and a number of these
factors have been reported to be associated with EMT, including
IL-6, IL-8, VEGF, TGFβ, SNAIL, MMP, TNFα and TWIST. The mechanism
underlying the interaction between these factors and EMT is
complicated. Notably, the dramatic phenotypic change in EMT is
coupled with motility and metastasis (21). Understanding the underlying mechanisms
involved in normal morphogenesis and designing treatment strategies
to reduce EMT is vital (17).
During the EMT process, cancer cells lose cell
polarity and adhesion. The cancer cells acquire increased migratory
and invasive capabilities. The EMT process is regulated by several
signaling pathways (22,23), which lead to cancer cell migration and
invasion. In breast cancer, cancer cells penetrate and transmigrate
into the basement membrane barriers, causing angiogenesis and
invasion (24) into the circulation.
It has been observed that when EMT was activated by epithelial
cells in the epithelium constituent of carcinosarcomas, the cells
exhibited epithelial and mesenchymal phenotype (25). Furthermore, the study was able to
directly assess epithelial plasticity and EMT reversal.
The EMT in carcinoma allows cancer cells to gain
increased motility and invasiveness. EMT involves a change in
phenotype from epithelial to mesenchymal, thereby allowing cells to
invade and colonize nearby tissues. Disseminated cancer cells need
to transmigrate the epithelial status during the period of
metastatic colonization. These cancer cells have high proliferative
potential, allowing the formation of secondary tumors. The normal
cellular junctions consist of specific epithelial splicing and
epigenetic mechanisms to maintain epithelial homeostasis (25). In summary, EMT is regulated by many
factors in the extracellular and intracellular matrix. It has a
vital role in regulating cancer cell motility, metastasis,
invasion, reverse transition and establishment of a secondary
tumor.
E-cadherin and vimentin are markers of epithelial
and mesenchymal cells, respectively (6). It has been previously demonstrated in
A549 human lung carcinoma cell line that a number of cytokines are
associated with EMT, including IL-8, VEGF, TGFβ and TNFα (26). It has also been reported that TGFβ is
able to induce EMT in multiple cell lines via activation of the
E-cadherin repressor (19). Bone
morphogenetic protein 7 is a member of the transforming growth
factor-β family and serves an important role in kidney development
(27). TGFβ has an important role in
cell migration and is a tumor-promoting factor. It has also been
reported that TGFβ is able to upregulate MMP expression in A549
lung cancer cells. TGFβ is able to induce EMT to promote metastasis
(28). Other studies have also
revealed that TGFβ is able to induce EMT by upregulating the
expression of zinc finger E-box binding homeobox 1 (ZEB1) in renal
tubular epithelial cells (29).
Epstein-Barr virus-induced TGFβ-spleen associated tyrosine kinase
(Syk)/Src AKT/extracellular signal-regulated kinase (ERK) signaling
may also be able to promote malignant and invasive potential in
human corneal epithelial cells by inducing EMT, and thus may be an
effective therapeutic target for the treatment of ocular disease
(30). Therefore, the TGFβ signaling
pathway may have an important role in advancing tumor progression
and metastasis.
The role of MMPs in cell invasion and tumor
metastasis has been well established. MMPs are able to remodel the
cell cytoskeleton in tumor cells to induce EMT. It has been
demonstrated that MMP-2 is able to facilitate tumor metastasis and
cell invasion. MMP-2 may therefore be a sensitive predictor of lung
tumor progression (31). Furthermore,
MMPs released by tumor-associated neutrophils, may facilitate tumor
progression, leading to cytoskeleton remodeling and promotion of
tumor metastasis (32). Therefore,
MMPs may have an important role in tumor metastasis and thus may
provide a novel target for cancer therapeutics.
Previous studies have suggested that SNAIL has an
important role in EMT in cancer cells, particularly in epithelial
tumor cells (33,34). SNAIL is able to reduce the number of
cell-cell junctions in cancer tissue and alter the cell
cytoskeleton (35). SNAIL, vimentin
and TWIST are upregulated in human hepatic cells, and these changes
are associated with EMT (13). It has
also been demonstrated that SNAIL is able to directly activate IL-8
via binding with the IL-8 receptor (36). EMT is promoted by the expression of
Notch, which leads to E-cadherin activation via SNAIL (37–39).
Additionally, a previous study suggested that high expression of
Notch leads to EMT (40). Notably,
SNAIL-induced EMT can be eliminated by anti-IL-8 receptor B
neutralizing antibodies, suggesting that IL-8 has an effective role
in mediating SNAIL-induced EMT and in advancing carcinoma
development (41). Targeting IL-8 may
provide a novel strategy for the treatment of cancer. Previous
studies have also revealed close associations between EMT and SNAIL
in tumor metastasis (42–44). ZEB1, TWIST and SNAIL have also been
implicated in carcinoma via the phosphatidyl inositol 3-kinase
(PI3K) (45,46) and glycogen synthase kinase 3β
signaling pathways (42).
It has been reported that the levels of IL-8 and
IL-8 receptor type 1 (CXCR1) and 2 (CXCR2) increase due to the
inhibition of phosphatase and tensin homolog (PTEN) in prostate
carcinoma (51,52). Overexpression of CXCR1 and CXCR2 has
been detected in prostate cancer tissue and promotes tumor
progression by contributing to cell proliferation and angiogenesis
(52). Furthermore, overexpression of
IL-8 and CXCR2 has been closely associated with tumor progression
and metastasis in esophageal squamous cell carcinoma (53). IL-8 may induce tumor progression,
metastasis and angiogenesis via CXCR2 (54). The role of microRNA-200 in inhibiting
angiogenesis via downregulation of IL-8 and CXCR1 in ovarian cancer
cell lines has also been reported (55). The expression of CXCR1 and CXCR2 was
not affected by chemotherapy in breast cancer where there was
increased expression of IL-8, thus CXCR may be desensitized prior
to and following chemotherapy (56).
IL-8 and CXCL1 can affect angiogenesis via endothelial CXCR2
receptors (57). In summary, IL-8 and
its receptors are closely associated with tumor progression,
angiogenesis and metastasis, which may promote EMT in tumor
cells.
Blocking IL-8 signaling is a potential strategy to
inhibit EMT and thus reduce tumor progression, metastasis and
angiogenesis, which may lead to improvements in 5-year disease-free
survival and overall survival rates (11). There is increased expression of IL-8
in the tumor microenvironment. IL-8 secretion may be mediated via
fibroblasts, endothelial cells and immune cells, which may promote
EMT in cancer (11).
The p38/Jun N-terminal kinase (JNK)-activating
transcription factor-2 (ATF-2) signaling pathway serves a vital
role in cell invasion and EMT, which is mediated by autocrine IL-8
in A549 lung cancer cells (2).
Previous studies have demonstrated that ATF-2 is able to promote
tumorigenesis, and has been observed to be upregulated in various
types of carcinoma, including mouse skin tumors (58), human neuroblastoma (59) and prostatic neoplasia (60). Notably, ATF-2 can be activated by IL-8
transcription (61). IL-8 is able to
induce the JNK/p38-ATF-2 signaling pathway and promote invasion in
A549 lung cancer cells (2).
Furthermore, ATF-2 is a potential therapeutic target for inhibiting
tumor metastasis (2).
The PI3K/AKT signaling pathway has an important role
in promoting cell proliferation and survival. Inhibiting the AKT
signaling pathway can lead to a decrease in cell motility, which is
induced by IL-8 stimulation. AKT is an important signaling pathway
for modulating IL-8-induced cell motility and invasion (62). PTEN can result in dysregulation of the
PI3K/AKT signaling axis in pancreatic ductal adenocarcinoma
(63). Furthermore, loss of PTEN
induces the upregulation of IL-8 signaling in prostate carcinoma
(51). A previous study has
demonstrated that there is high expression of phosphorylated AKT
when cells are treated with recombinant human netrin-1 in a human
hepatocellular carcinoma cell line (64). The role of AKT in activating nuclear
factor (NF)-κB signaling has been well established (65). Additionally, the NF-κB signaling
pathway has been associated with IL-8, which has an important
function in regulating tumor invasion (66). Therefore, cell motility can be
promoted by IL-8 via the AKT signaling pathway. It has also been
reported that AKT signaling can lead to EMT in breast cancer cells
(67).
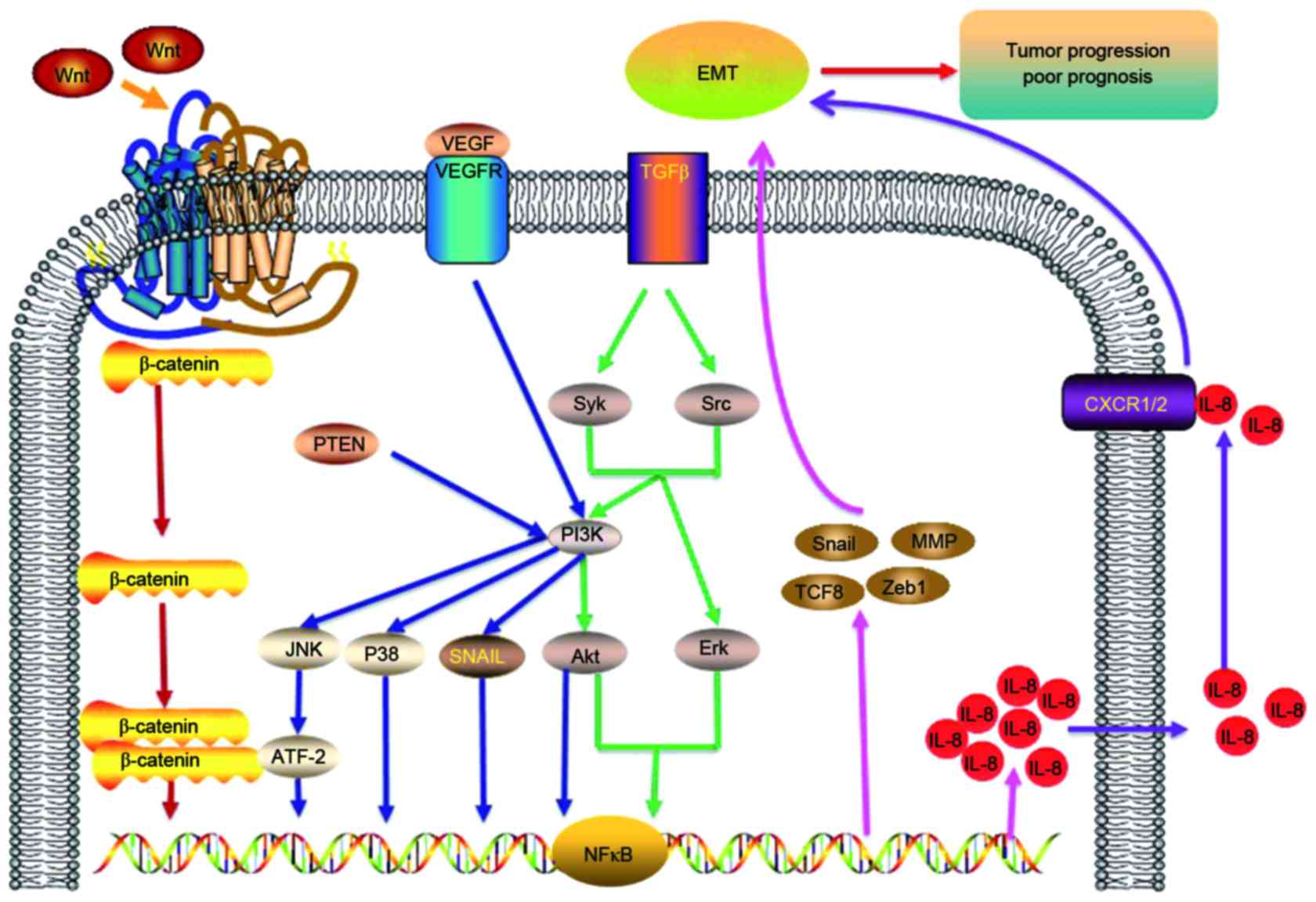
Several signaling pathways have been associated with
EMT, including the PI3K and Wnt signaling pathways (68). The Wnt signaling pathway can be
activated via overexpression of IL-8 (69). Previous studies have also indicated
that the Wnt signaling pathway serves an important role in
mediating cell-cell adhesion and beta-catenin self-phosphorylation
in tumor cells (70,71). Therefore, the PI3K/AKT, NF-κB and Wnt
signaling pathways are closely associated with IL-8 and EMT
(62,72). The signaling pathways and factors
associated with EMT in tumor cell proliferation, metastasis,
invasion and angiogenesis are displayed in Table I (11,12,14,73–83).
The cytokine IL-8 is a potential therapeutic target
for treating inflammatory diseases (84,85) and
inhibiting carcinoma angiogenesis (86). A previous study has demonstrated that
IL-8 mRNA expression has a role in EMT and tumor progression
(87). In prostate carcinoma cells,
increased expression of IL-8 promotes cancer progression, but the
expression of E-cadherin is reduced. In addition, IL-8 has an
important role in cancer cell proliferation, invasion and
metastasis (62). Cancer cells
secrete IL-8, and promote angiogenesis, cell proliferation,
metastasis and invasion (62). Under
hypoxic conditions, cancer cells are able to secrete IL-8 (78). Additionally, the expression of IL-8 in
various types of carcinoma tissues, including breast, colon,
gastric, lung and ovarian cancer has also been reported (88). IL-8 secretion is induced by TGFβ
stimulation (79) and SNAIL
overexpression (36), which can lead
to EMT in colorectal cancer cells.
IL-8 can induce and maintain the mesenchymal
phenotype to facilitate metastatic carcinoma progression. Previous
studies have reported that IL-8 secreted by the tumor stroma is
able to induce cell proliferation (89), migration, invasion and EMT (11,79,90). These
factors may enable cancer cells to evade apoptosis, and thus
promote cell survival (91). IL-8 is
also able to induce angiogenesis (92–94), and
facilitate cancer progression and metastasis in melanoma and
ovarian cancer (95,96). Increased serum levels of IL-8 are
associated with the risk of lung cancer, which precedes diagnosis
(77). Furthermore, a previous study
has demonstrated that the blockade of CXCR1 with a CXCR1-specific
blocking antibody or repertaxin, the small-molecule inhibitor of
IL-8, was able to inhibit angiogenesis, invasion, metastasis and
tumor progression in xenograft tumor models (97,98). IL-8
is able to promote cell migration, invasion, and metastasis
(72). IL-8 also serves a vital role
in EMT and can regulate the tumor microenvironment (6). In summary, there are close associations
between IL-8 and EMT in cancer. A list of the signaling pathways
and factors associated with IL-8, including PI3K/AKT, NF-κB,
p38/ATF-2, JNK, MMP and Wnt signaling pathways are displayed in
Fig. 1.
Increased expression of IL-8 in the tumor
microenvironment is associated with cell invasion and metastasis.
IL-8 may have an important role in ovarian cancer metastasis and
induces EMT (99). Another study also
reported the important role of a number of cytokines including IL-8
in tumor metastasis (100).
Furthermore, invasive tumor cells have increased expression of IL-8
compared with non-cancerous tissue cells (101). Tumor cells can promote cell motility
when there is an increased expression of IL-8 in the tumor
microenvironment, thus promoting cancer cell migration and
metastasis (102). The expression of
the brachyury gene is positively associated with IL-8 and
negatively associated with E-cadherin. This may lead to induction
of EMT and tumor metastasis in primary lung cancer (103). Studies generally have reported that
EMT can be induced by IL-8, which promotes cancer metastasis and
angiogenesis (6,11). However, IL-8 secretion in the tumor
microenvironment can also be induced by EMT.
Researchers have observed that IL-8 serves an
important role in the prognosis of several types of carcinoma,
including breast, colorectal, lung, gastric and prostate cancer
(77,104–108).
Increased levels of IL-8 and MMP-3 are indicators of poor prognosis
in triple-negative breast carcinomas (109). A study demonstrated that NF-κB
upregulates IL-8, which led to tumor progression and poorer
outcomes in a pancreatic cancer model (100). Brachyury mRNA is able to induce the
secretion of IL-8 and reduce the 5-year disease-free survival and
overall survival rates (103).
Additionally, the levels of IL-8 and its receptor may be employed
as an indictor to predict prognosis and survival rate. Increased
levels of IL-8 in ovarian, lung, renal and breast cancer have been
reported, and this was associated with poor prognosis (55). Therefore, inhibiting IL-8 may be a
potential strategy to control cancer cell migration, invasion and
metastasis. Furthermore, IL-8 is able to induce EMT and this leads
to a poor outcome in hepatocellular cancer patients (10). In summary, IL-8 has an important
association with EMT and prognosis in cancer patients.
EMT has an important role in the progression of
cancer metastasis. Induction of EMT is closely associated with
distant metastasis and cell invasion in tumor progression, and
indicates a poor prognosis. A number of studies have reported that
multiple factors can affect EMT in cancer, including IL-6, IL-8,
VEGF, TGFβ, SNAIL, MMP, TNFα and TWIST (30,110,111),
which can enhance cell motility and promote tumor metastasis. EMT
in cancer cells involves loss of cell-cell junctions and the
acquisition of cell motility and invasion factors. Multiple
signaling pathways are closely associated with EMT in tumors,
including TGFβ-Syk/Src-AKT/ERK, p38/JNK-ATF-2, PI3K/AKT, NF-κB and
Wnt signaling pathways (2,30,112–114).
Additionally, IL-8 and its receptors have associations with EMT in
cancer patients, thus blocking the IL-8/IL-8R axis may have be a
potential strategy to improve prognosis for cancer patients.
The present study was supported by the National
Natural Science Foundation of China (grant no. 11262020) and the
Foundation of Sichuan Provincial Science and Technology Program
(grant nos. 2013JY0021, 2014SZ0002-15 and 2016SZ0020).
|
1
|
Lee K and Nelson CM: New insights into the
regulation of epithelial-mesenchymal transition and tissue
fibrosis. Int Rev Cell Mol Biol. 294:171–221. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Desai S, Laskar S and Pandey BN: Autocrine
IL-8 and VEGF mediate epithelial-mesenchymal transition and
invasiveness via p38/JNK-ATF-2 signalling in A549 lung cancer
cells. Cell Signal. 25:1780–1791. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nieto MA: The ins and outs of the
epithelial to mesenchymal transition in health and disease. Annu
Rev Cell Dev Biol. 27:347–376. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li XJ, Peng LX, Shao JY, Lu WH, Zhang JX,
Chen S, Chen ZY, Xiang YQ, Bao YN, Zheng FJ, et al: As an
independent unfavorable prognostic factor, IL-8 promotes metastasis
of nasopharyngeal carcinoma through induction of
epithelial-mesenchymal transition and activation of AKT signaling.
Carcinogenesis. 33:1302–1309. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mathias RA, Gopal SK and Simpson RJ:
Contribution of cells undergoing epithelial-mesenchymal transition
to the tumour microenvironment. J Proteomics. 78:545–57. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Palena C, Hamilton DH and Fernando RI:
Influence of IL-8 on the epithelial-mesenchymal transition and the
tumor microenvironment. Future Oncol. 8:713–722. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Malmberg KJ and Ljunggren HG: Escape from
immune- and nonimmune-mediated tumor surveillance. Semin Cancer
Biol. 16:16–31. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Koch AE, Polverini PJ, Kunkel SL, Harlow
LA, DiPietro LA, Elner VM, Elner SG and Strieter RM: Interleukin-8
as a macrophage-derived mediator of angiogenesis. Science.
258:1798–1801. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bhusari PA and Khairnar KB: Greater
omental pancake tumour due to metastasis of ovarian cancer-a
cadaveric study. J Clin Diagn Res. 8:142–143. 2014.PubMed/NCBI
|
|
10
|
Yu J, Ren X, Chen Y, Liu P, Wei X, Li H,
Ying G, Chen K, Winkler H and Hao X: Dysfunctional activation of
neurotensin/IL-8 pathway in hepatocellular carcinoma is associated
with increased inflammatory response in microenvironment, more
epithelial mesenchymal transition in cancer and worse prognosis in
patients. PLoS One. 8:e560692013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fernando RI, Castillo MD, Litzinger M,
Hamilton DH and Palena C: IL-8 signaling plays a critical role in
the epithelial-mesenchymal transition of human carcinoma cells.
Cancer Res. 71:5296–5306. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Islam SS, Mokhtari RB, El Hout Y, Azadi
MA, Alauddin M, Yeger H and Farhat WA: TGF-β1 induces EMT
reprogramming of porcine bladder urothelial cells into collagen
producing fibroblasts-like cells in a Smad2/Smad3-dependent manner.
J Cell Commun Signal. 8:39–58. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bose SK, Meyer K, Di Bisceglie AM, Ray RB
and Ray R: Hepatitis C virus induces epithelial-mesenchymal
transition in primary human hepatocytes. J Virol. 86:13621–13628.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Radisky DC, Levy DD, Littlepage LE, Liu H,
Nelson CM, Fata JE, Leake D, Godden EL, Albertson DG, Nieto MA, et
al: Rac1b and reactive oxygen species mediate MMP-3-induced EMT and
genomic instability. Nature. 436:123–127. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ye Y, Liu P, Wang Y, Li H, Wei F, Cheng Y,
Han L and Yu J: Neurotensin, a novel messenger to cross-link
inflammation and tumor invasion via epithelial-mesenchymal
transition pathway. Int Rev Immunol. 35:340–350. 2016.PubMed/NCBI
|
|
16
|
Zhou N, Lu F, Liu C, Xu K, Huang J, Yu D
and Bi L: IL-8 induces the epithelial-mesenchymal transition of
renal cell carcinoma cells through the activation of AKT signaling.
Oncol Lett. 12:1915–1920. 2016.PubMed/NCBI
|
|
17
|
Nieto MA: Epithelial plasticity: A common
theme in embryonic and cancer cells. Science. 342:12348502013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hugo H, Ackland ML, Blick T, Lawrence MG,
Clements JA, Williams ED and Thompson EW: Epithelial-mesenchymal
and mesenchymal-epithelial transitions in carcinoma progression. J
Cell Physiol. 213:374–383. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kim MJ, Doh II, Bae GY, Cha HJ and Cho YH:
Cell-matrix adhesion characterization using multiple shear stress
zones in single stepwise microchannel. Appl Phys Lett.
105:0837012016. View Article : Google Scholar
|
|
21
|
Rokavec M, Öner MG, Li H, Jackstadt R,
Jiang L, Lodygin D, Kaller M, Horst D, Ziegler PK, Schwitalla S, et
al: IL-6R/STAT3/miR-34a feedback loop promotes EMT-mediated
colorectal cancer invasion and metastasis. J Clin Invest.
124:1853–1867. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Berx G, Raspé E, Christofori G, Thiery JP
and Sleeman JP: Pre-EMTing metastasis? Recapitulation of
morphogenetic processes in cancer. Clin Exp Metastasis. 24:587–597.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang J and Weinberg RA:
Epithelial-mesenchymal transition: At the crossroads of development
and tumor metastasis. Dev Cell. 14:818–829. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ota I, Li XY, Hu Y and Weiss SJ: Induction
of a MT1-MMP and MT2-MMP-dependent basement membrane transmigration
program in cancer cells by Snail1. Proc Natl Acad Sci USA. 106:pp.
20318–20323. 2009; View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sarrio D, Rodriguez-Pinilla SM, Hardisson
D, Cano A, Moreno-Bueno G and Palacios J: Epithelial-mesenchymal
transition in breast cancer relates to the basal-like phenotype.
Cancer Res. 68:989–997. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Desai S, Kumar A, Laskar S and Pandey BN:
Cytokine profile of conditioned medium from human tumor cell lines
after acute and fractionated doses of gamma radiation and its
effect on survival of bystander tumor cells. Cytokine. 61:54–62.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Dudley AT, Lyons KM and Robertson EJ: A
requirement for bone morphogenetic protein-7 during development of
the mammalian kidney and eye. Genes Dev. 9:2795–2807. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Illman SA, Lehti K, Keski-Oja J and Lohi
J: Epilysin (MMP-28) induces TGF-beta mediated epithelial to
mesenchymal transition in lung carcinoma cells. J Cell Science.
119:3856–3865. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xiong M, Jiang L, Zhou Y, Qiu W, Fang L,
Tan R, Wen P and Yang J: The miR-200 family regulates
TGF-β1-induced renal tubular epithelial to mesenchymal transition
through Smad pathway by targeting ZEB1 and ZEB2 expression. Am J
Physiol Renal Physiol. 302:F369–F379. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Park GB, Kim D, Kim YS, Kim S, Lee HK,
Yang JW and Hur DY: The Epstein-Barr virus causes
epithelial-mesenchymal transition in human corneal epithelial cells
via Syk/src and Akt/Erk signaling pathways. Invest Ophthalmol Vis
Sci. 55:1770–1779. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Guo CB, Wang S, Deng C, Zhang DL, Wang FL
and Jin XQ: Relationship between matrix metalloproteinase 2 and
lung cancer progression. Mol Diagn Ther. 11:183–192. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gregory AD and Houghton AM:
Tumor-associated neutrophils: New targets for cancer therapy.
Cancer Res. 71:2411–2416. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Fang S, Yu L, Mei H, Yang J, Gao T, Cheng
A, Guo W, Xia K and Liu G: Cisplatin promotes mesenchymal-like
characteristics in osteosarcoma through Snail. Oncol Lett.
12:5007–5014. 2016.PubMed/NCBI
|
|
34
|
Alba-Castellon L, Olivera-Salguero R,
Mestre-Farrera A, Pena R, Herrera M, Bonilla F, Casal JI, Baulida
J, Peña C and García de Herreros A: Snail1-dependent activation of
cancer-associated fibroblast controls epithelial tumor cell
invasion and metastasis. Cancer Res. 76:6205–6217. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Haraguchi M, Sato M and Ozawa M:
CRISPR/Cas9n-mediated deletion of the Snail 1Gene (SNAI1) reveals
its role in regulating cell morphology, cell-cell interactions and
gene expression in ovarian cancer (RMG-1) cells. PLoS One.
10:e01322602015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hwang WL, Yang MH, Tsai ML, Lan HY, Su SH,
Chang SC, Teng HW, Yang SH, Lan YT, Chiou SH and Wang HW: SNAIL
regulates interleukin-8 expression, stem cell-like activity and
tumorigenicity of human colorectal carcinoma cells.
Gastroenterology. 141:279–291, 291.e1-e5. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Leong KG, Niessen K, Kulic I, Raouf A,
Eaves C, Pollet I and Karsan A: Jagged1-mediated Notch activation
induces epithelial-to-mesenchymal transition through Slug-induced
repression of E-cadherin. J Exp Med. 204:2935–2948. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Niessen K, Fu Y, Chang L, Hoodless PA,
McFadden D and Karsan A: Slug is a direct Notch target required for
initiation of cardiac cushion cellularization. J Cell Biol.
182:315–325. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sahlgren C, Gustafsson MV, Jin S,
Poellinger L and Lendahl UL: Notch signaling mediates
hypoxia-induced tumor cell migration and invasion. Proc Natl Acad
Sci USA. 105:pp. 6392–6397. 2008; View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Timmerman LA, Grego-Bessa J, Raya A,
Bertrén E, Pérez-Pomares JM, Díez J, Aranda S, Palomo S, McCormick
F, Izpisúa-Belmonte JC and de la Pompa JL: Notch promotes
epithelial-mesenchymal transition during cardiac development and
oncogenic transformation. Genes Dev. 18:99–115. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yanagawa J, Walser TC, Zhu LX, Hong L,
Fishbein MC, Mah V, Chia D, Goodglick L, Elashoff DA, Luo J, et al:
Snail promotes CXCR2 ligand-dependent tumor progression in
non-small cell lung carcinoma. Clin Cancer Res. 15:6820–6829. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liu ZC, Chen XH, Song HX, Wang HS, Zhang
G, Wang H, Chen DY, Fang R, Liu H, Cai SH and Du J: Snail regulated
by PKC/GSK-3β pathway is crucial for EGF-induced
epithelial-mesenchymal transition (EMT) of cancer cells. Cell
Tissue Res. 358:491–502. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Gras B, Jacqueroud L, Wierinckx A, Lamblot
C, Fauvet F, Lachuer J, Puisieux A and Ansieau S: Snail family
members unequally trigger EMT and thereby differ in their ability
to promote the neoplastic transformation of mammary epithelial
cells. PLoS One. 9:e922542014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wettstein G, Bellaye PS, Kolb M, Hammann
A, Crestani B, Soler P, Marchal-Somme J, Hazoume A, Gauldie J,
Gunther A, et al: Inhibition of HSP27 blocks fibrosis development
and EMT features by promoting Snail degradation. FASEB J.
27:1549–1560. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zheng H and Kang Y: Multilayer control of
the EMT master regulators. Oncogene. 33:1755–1763. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhang J, Zhou Y and Yang Y: CCR7 pathway
induces epithelial-mesenchymal transition through up-regulation of
Snail signaling in gastric cancer. Med Oncol. 32:4672015.PubMed/NCBI
|
|
47
|
Sullivan NJ, Sasser AK, Axel AE, Vesuna F,
Raman V, Ramirez N, Oberyszyn TM and Hall BM: Interleukin-6 induces
an epithelial-mesenchymal transition phenotype in human breast
cancer cells. Oncogene. 28:2940–2947. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Gonzalez-Moreno O, Lecanda J, Green JE,
Segura V, Catena R, Serrano D and Calvo A: VEGF elicits
epithelial-mesenchymal transition (EMT) in prostate intraepithelial
neoplasia (PIN)-like cells via an autocrine loop. Exp Cell Res.
316:554–567. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Mironchik Y, Winnard PT Jr, Vesuna F, Kato
Y, Wildes F, Pathak AP, Kominsky S, Artemov D, Bhujwalla Z, van
Diest P, et al: Twist overexpression induces in vivo angiogenesis
and correlates with chromosomal instability in breast cancer.
Cancer Res. 65:10801–10809. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wang Y, Yao X, Ge J, Hu F and Zhao Y: Can
vascular endothelial growth factor and microvessel density be used
as prognostic biomarkers for colorectal cancer? A systematic review
and meta-analysis. ScientificWorldJournal.
2014:1027362014.PubMed/NCBI
|
|
51
|
Maxwell PJ, Coulter J, Walker SM,
McKechnie M, Neisen J, McCabe N, Kennedy RD, Salto-Tellez M,
Albanese C and Waugh DJ: Potentiation of inflammatory CXCL8
signalling sustains cell survival in PTEN-deficient prostate
carcinoma. Eur Urol. 64:177–88. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Murphy C, McGurk M, Pettigrew J,
Santinelli A, Mazzucchelli R, Johnston PG, Montironi R and Waugh
DJ: Nonapical and cytoplasmic expression of interleukin-8, CXCR1,
and CXCR2 correlates with cell proliferation and microvessel
density in prostate cancer. Clin Cancer Res. 11:4117–4127. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ogura M, Takeuchi H, Kawakubo H, Nishi T,
Fukuda K, Nakamura R, Takahashi T, Wada N, Saikawa Y, Omori T, et
al: Clinical significance of CXCL-8/CXCR-2 network in esophageal
squamous cell carcinoma. Surgery. 154:512–520. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Uzunoglu FG, Yavari N, Bohn BA, Nentwich
MF, Reeh M, Pantel K, Perez D, Tsui TY, Bockhorn M, Mann O, et al:
C-X-C motif receptor 2, endostatin and proteinase-activated
receptor 1 polymorphisms as prognostic factors in NSCLC. Lung
Cancer. 81:123–129. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Pecot CV, Rupaimoole R, Yang D, Akbani R,
Ivan C, Lu C, Wu S, Han HD, Shah MY, Rodriguez-Aguayo C, et al:
Tumour angiogenesis regulation by the miR-200 family. Nat Commun.
4:24272013. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Mendonca MA, Souto FO, Micheli DC,
Alves-Filho JC, Cunha FQ, Murta EF and Tavares-Murta BM: Mechanisms
affecting neutrophil migration capacity in breast cancer patients
before and after chemotherapy. Cancer Chemother Pharmacol.
73:317–324. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Merritt WM, Lin YG, Spannuth WA, Fletcher
MS, Kamat AA, Han LY, Landen CN, Jennings N, de Geest K, Langley
RR, et al: Effect of interleukin-8 gene silencing with
liposome-encapsulated small interfering RNA on ovarian cancer cell
growth. J Natl Cancer Inst. 100:359–372. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Papassava P, Gorgoulis VG, Papaevangeliou
D, Vlahopoulos S, van Dam H and Zoumpourlis V: Overexpression of
activating transcription factor-2 is required for tumor growth and
progression in mouse skin tumors. Cancer Res. 64:8573–8584. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Tindberg N, Porsmyr-Palmertz M and Simi A:
Contribution of MAP kinase pathways to the activation of ATF-2 in
human neuroblastoma cells. Neurochem Res. 25:527–531. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Ricote M, Garcia-Tunon I, Bethencourt F,
Fraile B, Onsurbe P, Paniagua R and Royuela M: The p38 transduction
pathway in prostatic neoplasia. J Pathol. 208:401–407. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Eliopoulos AG, Gallagher NJ, Blake SM,
Dawson CW and Young LS: Activation of the p38 mitogen-activated
protein kinase pathway by Epstein-Barr virus-encoded latent
membrane protein 1 coregulates interleukin-6 and interleukin-8
production. J Biol Chem. 274:16085–16096. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Wang L, Tang C, Cao H, Li K, Pang X, Zhong
L, Dang W, Tang H, Huang Y, Wei L, et al: Activation of IL-8 via
PI3K/Akt-dependent pathway is involved in leptin-mediated
epithelial-mesenchymal transition in human breast cancer cells.
Cancer Biol Ther. 16:1220–1230. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Hill R, Calvopina JH, Kim C, Wang Y,
Dawson DW, Donahue TR, Dry S and Wu H: PTEN loss accelerates
KrasG12D-induced pancreatic cancer development. Cancer Res.
70:7114–7124. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Yan W, Han P, Zhou Z, Tu W, Liao J, Li P,
Liu M, Tian D and Fu Y: Netrin-1 induces epithelial-mesenchymal
transition and promotes hepatocellular carcinoma invasiveness. Dig
Dis Sci. 59:1213–1221. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Song J, Feng L, Zhong R, Xia Z, Zhang L,
Cui L, Yan H, Jia X and Zhang Z: Icariside II inhibits the EMT of
NSCLC cells in inflammatory microenvironment via down-regulation of
Akt/NF-κB signaling pathway. Mol Carcinog. 56:36–48. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Voorzanger N, Touitou R, Garcia E,
Delecluse HJ, Rousset F, Joab I, Favrot MC and Blay JY: Interleukin
(IL)-10 and IL-6 are produced in vivo by non-Hodgkin's lymphoma
cells and act as cooperative growth factors. Cancer Res.
56:5499–5505. 1996.PubMed/NCBI
|
|
67
|
Carpenter RL, Paw I, Dewhirst MW and Lo
HW: Akt phosphorylates and activates HSF-1 independent of heat
shock, leading to Slug overexpression and epithelial-mesenchymal
transition (EMT) of HER2-overexpressing breast cancer cells.
Oncogene. 34:546–557. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Baranwal S and Alahari SK: Molecular
mechanisms controlling E-cadherin expression in breast cancer.
Biochem Biophys Res Commun. 384:6–11. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Moon HG, Zheng Y, An CH, Kim YK and Jin Y:
CCN1 secretion induced by cigarette smoking extracts augments IL-8
release from bronchial epithelial cells. PLoS One. 8:e681992013.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Cai J, Guan H, Fang L, Yang Y, Zhu X, Yuan
J, Wu J and Li M: MicroRNA-374a activates Wnt/β-catenin signaling
to promote breast cancer metastasis. J Clin Invest. 123:566–579.
2013.PubMed/NCBI
|
|
71
|
Zappulli V, de Cecco S, Trez D, Caliari D,
Aresu L and Castagnaro M: Immunohistochemical expression of
E-cadherin and β-catenin in feline mammary tumours. J Comp Pathol.
147:161–170. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Ahn SH, Park H, Ahn YH, Kim S, Cho MS,
Kang JL and Choi YH: Necrotic cells influence migration and
invasion of glioblastoma via NF-κB/AP-1-mediated IL-8 regulation.
Sci Rep. 6:245522016. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Cheng XS, Li YF, Tan J, Sun B, Xiao YC,
Fang XB, Zhang XF, Li Q, Dong JH, Li M, et al: CCL20 and CXCL8
synergize to promote progression and poor survival outcome in
patients with colorectal cancer by collaborative induction of the
epithelial-mesenchymal transition. Cancer Lett. 348:77–87. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Zhao ZW, Wang YX, Lv X, Nie YM and Wu J:
Shear stress promotes epithelial-mesenchymal transition of
laryngeal cancer cells by inducing IL-8/CXCR1-NF-Kappa B axis. J
Invest Med. 62:S802014.
|
|
75
|
Choi SH, Kwon OJ, Park JY, Kim DY, Ahn SH,
Kim SU, Ro SW, Kim KS, Park JH, Kim S, et al: Inhibition of tumour
angiogenesis and growth by small hairpin HIF-1α and IL-8 in
hepatocellular carcinoma. Liver Int. 34:632–642. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Wu S, Shang H, Cui L, Zhang Z, Zhang Y, Li
Y, Wu J, Li RK and Xie J: Targeted blockade of interleukin-8
abrogates its promotion of cervical cancer growth and metastasis.
Mol Cell Biochem. 375:69–79. 2013.PubMed/NCBI
|
|
77
|
Pine SR, Mechanic LE, Enewold L,
Chaturvedi AK, Katki HA, Zheng YL, Bowman ED, Engels EA, Caporaso
NE and Harris CC: Increased levels of circulating interleukin 6,
interleukin 8, C-reactive protein, and risk of lung cancer. J Natl
Cancer Inst. 103:1112–1122. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Xie K: Interleukin-8 and human cancer
biology. Cytokine Growth Factor Rev. 12:375–391. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Bates RC, DeLeo MJ III and Mercurio AM:
The epithelial-mesenchymal transition of colon carcinoma involves
expression of IL-8 and CXCR-1-mediated chemotaxis. Exp Cell Res.
299:315–324. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Yang JR, Pan TJ, Yang H, Wang T, Liu W,
Liu B and Qian WH: Kindlin-2 promotes invasiveness of prostate
cancer cells via NF-κB-dependent upregulation of matrix
metalloproteinases. Gene. 576:571–576. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Zucchini-Pascal N, Peyre L and Rahmani R:
Crosstalk between beta-catenin and snail in the induction of
epithelial to mesenchymal transition in hepatocarcinoma: Role of
the ERK1/2 pathway. Int J Mol Sci. 14:20768–20792. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Zhao J, Zhang ZR, Zhao N, Ma BA and Fan
QY: VEGF silencing inhibits human osteosarcoma angiogenesis and
promotes cell apoptosis via PI3K/AKT signaling pathway. Int J Clin
Exp Med. 8:12411–12417. 2015.PubMed/NCBI
|
|
83
|
Matsumoto G, Hirohata R, Hayashi K,
Sugimoto Y, Kotani E, Shimabukuro J, Hirano T, Nakajima Y, Kawamata
S and Mori H: Control of angiogenesis by VEGF and
endostatin-encapsulated protein microcrystals and inhibition of
tumor angiogenesis. Biomaterials. 35:1326–1333. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Shen Y, Wang D and Wang X: Role of CCR2
and IL-8 in acute lung injury: A new mechanism and therapeutic
target. Expert Rev Respir Med. 5:107–114. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Skov L, Beurskens FJ, Zachariae CO,
Reitamo S, Teeling J, Satijn D, Knudsen KM, Boot EP, Hudson D,
Baadsgaard O, et al: IL-8 as antibody therapeutic target in
inflammatory diseases: Reduction of clinical activity in
palmoplantar pustulosis. J Immunol. 181:669–679. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Hatfield KJ, Olsnes AM, Gjertsen BT and
Bruserud Ø: Antiangiogenic therapy in acute myelogenous leukemia:
Targeting of vascular endothelial growth factor and interleukin 8
as possible antileukemic strategies. Curr Cancer Drug Targets.
5:229–248. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Li WH, Qiu Y, Zhang HQ, Liu Y, You JF,
Tian XX and Fang WG: P2Y2 receptor promotes cell invasion and
metastasis in prostate cancer cells. Br J Cancer. 109:1666–1675.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Waugh DJ and Wilson C: The interleukin-8
pathway in cancer. Clin Cancer Res. 14:6735–6741. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Schadendorf D, Möller A, Algermissen B,
Worm M, Sticherling M and Czarnetzki BM: IL-8 produced by human
malignant melanoma cells in vitro is an essential autocrine growth
factor. J Immunol. 151:2667–2675. 1993.PubMed/NCBI
|
|
90
|
de Larco JE, Wuertz BR, Rosner KA,
Erickson SA, Gamache DE, Manivel JC and Furcht LT: A potential role
for interleukin-8 in the metastatic phenotype of breast carcinoma
cells. Am J Pathol. 158:639–646. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Maxwell PJ, Gallagher R, Seaton A, Wilson
C, Scullin P, Pettigrew J, Stratford IJ, Williams KJ, Johnston PG
and Waugh DJ: HIF-1 and NF-kappaB-mediated upregulation of CXCR1
and CXCR2 expression promotes cell survival in hypoxic prostate
cancer cells. Oncogene. 26:7333–7345. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Lattanzio L, Tonissi F, Torta I, Gianello
L, Russi E, Milano G, Merlano M and Lo Nigro C: Role of IL-8
induced angiogenesis in uveal melanoma. Invest New Drugs.
31:1107–1114. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Li KC, Huang YH, Ho CY, Chu CY, Cha ST,
Tsai HH, Ko JY, Chang CC and Tan CT: The role of IL-8 in the
SDF-1α/CXCR4-induced angiogenesis of laryngeal and hypopharyngeal
squamous cell carcinoma. Oral Oncol. 48:507–515. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Shen XH, Xu SJ, Jin CY, Ding F, Zhou YC
and Fu GS: Interleukin-8 prevents oxidative stress-induced human
endothelial cell senescence via telomerase activation. Int
Immunopharmacol. 16:261–267. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Rofstad EK and Halsør EF: Vascular
endothelial growth factor, interleukin 8, platelet-derived
endothelial cell growth factor, and basic fibroblast growth factor
promote angiogenesis and metastasis in human melanoma xenografts.
Cancer Res. 60:4932–4938. 2000.PubMed/NCBI
|
|
96
|
Shahzad MM, Arevalo JM, Armaiz-Pena GN, Lu
C, Stone RL, Moreno-Smith M, Nishimura M, Lee JW, Jennings NB,
Bottsford-Miller J, et al: Stress effects on FosB- and
interleukin-8 (IL8)-driven ovarian cancer growth and metastasis. J
Biol Chem. 285:35462–3547070. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Ginestier C, Liu S, Diebel ME, Korkaya H,
Luo M, Brown M, Wicinski J, Cabaud O, Charafe-Jauffret E, Birnbaum
D, et al: CXCR1 blockade selectively targets human breast cancer
stem cells in vitro and in xenografts. J Clin Invest. 120:485–497.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Sparmann A and Bar-Sagi D: Ras-induced
interleukin-8 expression plays a critical role in tumor growth and
angiogenesis. Cancer Cell. 6:447–458. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Yin J, Zeng F, Wu N, Kang K, Yang Z and
Yang H: Interleukin-8 promotes human ovarian cancer cell migration
by epithelial-mesenchymal transition induction in vitro. Clin
Transl Oncol. 17:365–370. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Kim SW, Hayashi M, Lo JF, Fearns C, Xiang
R, Lazennec G, Yang Y and Lee JD: Tid1 negatively regulates the
migratory potential of cancer cells by inhibiting the production of
interleukin-8. Cancer Res. 65:8784–8791. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Luca M, Huang S, Gershenwald JE, Singh RK,
Reich R and Bar-Eli M: Expression of interleukin-8 by human
melanoma cells up-regulates MMP-2 activity and increases tumor
growth and metastasis. Am J Pathol. 151:1105–1113. 1997.PubMed/NCBI
|
|
102
|
Sheridan C, Kishimoto H, Fuchs RK,
Mehrotra S, Bhat-Nakshatri P, Turner CH, Badve R Jr, Goulet S and
Nakshatri H: CD44+/CD24- breast cancer cells exhibit enhanced
invasive properties: An early step necessary for metastasis. Breast
Cancer Res. 8:R592006. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Haro A, Yano T, Kohno M, Yoshida T, Koga
T, Okamoto T, Takenoyama M and Maehara Y: Expression of Brachyury
gene is a significant prognostic factor for primary lung carcinoma.
Ann Surg Oncol. 20 Suppl 3:S509–S516. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Choi J, Song N, Han S, Chung S, Sung H,
Lee JY, Jung S, Park SK, Yoo KY, Han W, et al: The associations
between immunity-related genes and breast cancer prognosis in
Korean women. PLoS One. 9:e1035932014. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Hanker LC, Rody A, Holtrich U, Pusztai L,
Ruckhaeberle E, Liedtke C, Ahr A, Heinrich TM, Sänger N, Becker S
and Karn T: Prognostic evaluation of the B cell/IL-8 metagene in
different intrinsic breast cancer subtypes. Breast Cancer Res
Treat. 137:407–16. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Reitter EM, Ay C, Kaider A, Pirker R,
Zielinski C, Zlabinger G and Pabinger I: Interleukin levels and
their potential association with venous thromboembolism and
survival in cancer patients. Clin Exp Immunol. 177:253–260. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Wang S, Wu X, Zhang J, Chen Y, Xu J, Xia
X, He S, Qiang F, Li A, Shu Y, et al: CHIP functions as a novel
suppressor of tumour angiogenesis with prognostic significance in
human gastric cancer. Gut. 62:496–508. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Manna S, Singha B, Phyo SA, Gatla HR,
Chang TP, Sanacora S, Ramaswami S and Vancurova I: Proteasome
inhibition by bortezomib increases IL-8 expression in
androgen-independent prostate cancer cells: The role of IKKα. J
Immunol. 191:2837–2846. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Han J, Bae SY, Oh SJ, Lee J, Lee JH, Lee
HC, Lee SK, Kil WH, Kim SW, Nam SJ, et al: Zerumbone suppresses
IL-1β-induced cell migration and invasion by inhibiting IL-8 and
MMP-3 expression in human triple-negative breast cancer cells.
Phytother Res. 28:1654–1660. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Epanchintsev A, Shyamsunder P, Verma RS
and Lyakhovich A: IL-6, IL-8, MMP-2, MMP-9 are overexpressed in
Fanconi anemia cells through a NF-κB/TNF-α dependent mechanism. Mol
Carcinog. 54:1686–1699. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Dong P, Xiong Y, Watari H, Hanley SJ,
Konno Y, Ihira K, Yamada T, Kudo M, Yue J and Sakuragi N: MiR-137
and miR-34a directly target Snail and inhibit EMT, invasion and
sphere-forming ability of ovarian cancer cells. J Exp Clin Cancer
Res. 35:1322016. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Meng J, Zhang XT, Liu XL, Fan L, Li C, Sun
Y, Liang XH, Wang JB, Mei QB, Zhang F and Zhang T: WSTF promotes
proliferation and invasion of lung cancer cells by inducing EMT via
PI3K/Akt and IL-6/STAT3 signaling pathways. Cell Signal.
28:1673–1682. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Huang T, Chen Z and Fang L: Curcumin
inhibits LPS-induced EMT through downregulation of NF-κB-snail
signaling in breast cancer cells. Oncol Rep. 29:117–124.
2013.PubMed/NCBI
|
|
114
|
Zhang Z, Chen H, Xu C, Song L, Huang L,
Lai Y, Wang Y, Chen H, Gu D, Ren L and Yao Q: Curcumin inhibits
tumor epithelial-mesenchymal transition by downregulating the Wnt
signaling pathway and upregulating NKD2 expression in colon cancer
cells. Oncol Rep. 35:2615–2623. 2016.PubMed/NCBI
|