Introduction
The S100 calcium binding proteins are a multi-gene
family composed of ≥20 members in humans, each encoded by a
separate gene (1). The deregulated
expression of several members of the S100 protein family, including
S100 calcium binding protein (S100) B, S100A2, S100A4, S100A6,
S100A11 and S100P has been reported to be associated with the
progression and metastasis of various types of human cancer
(2). Head and neck cancer is eighth
leading cause of cancer-associated mortality worldwide (3). Previous studies have indicated the
potential association between S100 proteins and head and neck
cancer (4,5), and the altered expression of various
S100 proteins has been reported in primary and metastatic laryngeal
carcinoma (6). In a previous study,
S100A11 was revealed to be overexpressed in laryngeal tumor
tissues, as compared with the corresponding noncancerous tissues,
which was determined by reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) and western blot analysis
(7). In addition, S100A11 expression
was correlated with human epithelial type 2 cell migration
(7). The expression of S100A4 has
also been correlated with invasion and metastasis in oral squamous
cell carcinoma (8).
Nasopharyngeal carcinoma (NPC) is the most prevalent
head and neck cancer in Southern China and Southeast Asia (9). NPC is distinct from other types of head
and neck cancer due to its sensitivity to radiotherapy and
chemotherapy (10). Although
radiotherapy enables a high cure rate for early-stage NPC, the
treatment outcome for the advanced stage disease remains poor
(11). Local recurrences and the high
frequency of distant metastasis are two major causes of treatment
failure in NPC following radiotherapy (12).
S100P is a 95 amino acid protein and a member of the
S100 family of EF-hand motif calcium-binding proteins, which was
initially purified from the human placenta and exhibits a
restricted cellular distribution (13). Its deregulated expression has
previously been observed in association with the progression and
metastasis of various types of human cancer, including breast
(14), lung (15), gastric (16) and prostate cancer (17). Despite these observations, the
functional role and the underlying mechanism of action of S100P in
nasopharyngeal carcinoma requires further investigation.
In the present study, the differential expression of
S100P in NPC tissues was examined using immunohistochemistry (IHC),
and the changes of proliferation and migration of C666-1 cells were
analyzed following S100P silencing with the aim of investigating
the role of S100P in NPC.
Patients and methods
Patients and tissue samples
Pathological tissue specimens were obtained
retrospectively from 78 patients with nasopharyngeal carcinoma and
30 patients with benign inflammation at The Department of
Otolaryngology-Head and Neck Surgery, Changzheng Hospital, Second
Military Medical University (Shanghai, China) between January 2005
and June 2008. Patient characteristics were obtained from the
medical records (Table I). The median
age of the 78 patients with nasopharyngeal carcinoma was 51.78
years old (range, 21–89 years), and 22 (28.21%) of the patients
were >60 years of age. The median age of 30 patients with benign
inflammation was 52.09 years (range, 19–78 years), and 8 (26.67%)
of the patients were >60 years of age. There were 53 males
(67.95%) and 25 females (32.05%) with nasopharyngeal carcinoma,
compared with 22 males (73.33%) and 8 females (26.67%) with benign
inflammation.
 | Table I.Clinical characteristics of the
patients with nasopharyngeal carcinoma. |
Table I.
Clinical characteristics of the
patients with nasopharyngeal carcinoma.
|
| S100Ps |
|
|---|
|
|
|
|
|---|
| Patient
characteristics | Negative | Positive | P-value |
|---|
| Sex |
|
| 0.835 |
|
Male | 22 | 31 |
|
|
Female | 11 | 14 |
|
| Age, years |
|
| 0.234 |
|
<60 | 20 | 33 |
|
|
≥60 | 13 | 12 |
|
| Primary tumor |
|
| 0.066 |
| T1 | 22 | 38 |
|
|
T2-T4 | 11 | 7 |
|
| Nodal status |
|
| 0.656 |
|
N0-N1 | 20 | 25 |
|
|
N2-N3 | 13 | 20 |
|
| Stage |
|
| 0.929 |
|
I–II | 18 | 25 |
|
|
III–IV | 15 | 20 |
|
| Histological
grade |
|
| 0.392 |
|
Keratinized | 5 | 3 |
|
|
Non-keratinized | 8 | 15 |
|
|
Undifferentiated | 20 | 27 |
|
The pathological tissue specimens of all the
patients who had undergone neither chemotherapy nor radiotherapy
underwent biopsy for immunohistochemical examination of S100P
expression levels. The patients follow-up data was available for
>5 years. All the tissue specimens were obtained for the present
study following the receipt of written informed patient consent.
The collection of these tissue samples was undertaken with the
approval of The Second Military Medical University Ethics
Committee.
IHC
IHC staining was performed using a Dako EnVision
Peroxidase/DAB system (Dako North America, Inc., Carpinteria, CA,
USA) according to the manufacturer's protocol. The tissue samples
from each patient were fixed in formalin and embedded in paraffin.
Subsequently, the tissue sections (4 µm thick) were deparaffinized
using xylene, rehydrated with graded ethanol and heated for 1 h at
65°C. The sections were submerged in EDTA antigenic retrieval
buffer (pH 8.0, K800221-2, EnVision FLEX+, Agilent Tech) and were
heated with high heat for 3 min, then heated with low heat for 15
min using a microwave (Galanz microwave oven P70D20P-N9). Following
treatment with 0.3% H2O2 for 15 min at room temperature to block
endogenous peroxidase activity, the tissue sections were treated
with 1% bovine serum albumin (ab192603; Abcam, Cambridge, UK) for
30 min in the 37°C incubator to reduce non-specific binding, and
subsequently incubated with a primary rabbit polyclonal anti-S100P
antibody (dilution, 1:500; ab124743; Abcam) overnight at 4°C,
followed by incubation with horseradish peroxidase-labeled
secondary antibodies (dilution, 1:300; ab181875; Abcam) at 4°C for
30 min. The slides were developed using diaminobenzidine
tetrahydrochloride and counterstained with hematoxylin.
Immunohistochemically stained tissue sections were scored
independently by two pathologists (Department of Pathology,
Changzheng Hospital, Second Military Medical University) who were
blinded to the clinical parameters. The tissue specimens were
defined as positive for S100P expression if the tumor cells were
distinctly stained by the anti-S100P antibody. Entire tissue
sections were imaged under a microscope to enable the scoring of
S100P staining. In each section, 5 fields were selected randomly
under a high power microscope (magnification, ×400), within each
field 200 tumor cells were counted. The staining intensity was
scored as follows: 0 (negative), 1 (weak) and 2 (intense). The
extent of staining was scored as 0 (0–10%), 1 (11–25%), 2 (26–50%)
and 3 (51–100%), according to the proportion of positive-staining
areas in the entire carcinoma area, or the entire section for the
benign tissue samples. The product of the intensity and the extent
of staining scores was used as the final staining score (0–6) for
S100P (score=intensity of staining score × extent of staining
score). For the purpose of statistical evaluation, tumors
(nasopharyngeal carcinoma tissue sections) with a final staining
score of >2 were considered to be positive.
Cell culture and maintenance of cell
lines
The C666-1 cell line was obtained from the cell bank
of The Central Laboratory of Xiangya Hospital (Central South
University, Changsha, China) (18).
C666-1 cells were cultured in RPMI-1640 (GE Healthcare Life
Sciences; Hyclone, Logan, UT, USA) supplemented with 12% fetal
bovine serum (FBS) (Biowest, Maine et Loire, France) and 1%
penicillin and streptomycin (P/S) (Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) and were maintained in a humidified 37°C
incubator containing 5% CO2.
Transfection of small interfering RNA
(siRNA)
S100P knockdown experiments were performed by
transfecting S100P siRNA into C666-1 cells. siRNA against human
S100P was chemically synthesized by GenePharma Co., Ltd.,
(Shanghai, China) according to the sequence published previously by
Arumugam et al (19). The
siRNA sequences were as follows: Forward,
5′-AAUGGAGAUGCCCAGGUGGACTT-3′ and reverse,
5′-GUCCACCUGGGCAUCUCCAUUTT-3′. The negative control siRNA sequences
were as follows: Forward, 5′-UUCUCCGAACGUGUCACGUTT-3′ and reverse,
5′-ACGUGACACGUUCGGAGAATT-3′. S100P siRNA transfection was performed
using Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) according to the manufacturer's
protocol. C666-1 cells were plated at a density of 2×105
cells/well in 6-well plates and were incubated at 37°C overnight in
2 ml RPMI-1640 (GE Healthcare Life Sciences; Hyclone) supplemented
with 12% fetal bovine serum (FBS; Biowest) without antibiotics.
Upon reaching 30–50% confluency, the culture medium was aspirated.
A total of 5 µl of 20 µM siRNA and 5 µl Lipofectamine®
2000 were combined for 20 min and subsequently added to a final
volume of 2 ml of serum-free Gibco™ Opti-MEM™
medium (Thermo Fisher Scientific, Inc.). The medium was replaced 5
h following transfection, and RNA and protein were harvested from
the cells at 24 or 48 h post-transfection for the evaluation of
S100P knockdown. The effects of S100P gene knockdown were examined
using RT-qPCR and western blotting.
RNA isolation and RT-qPCR
analysis
Total RNA (2 µg) was extracted from the S100P
siRNA-transfected C666-1 cells using TRIzol® reagent
(Ambion; Thermo Fisher Scientific, Inc.), and reverse transcription
was performed using the RevertAid First Strand cDNA Synthesis kit
(Fermentas; Thermo Fisher Scientific, Inc.). The primers were
chemically synthesized by GenePharma. The primer sequences were as
follows: Forward, 5′-ATGACGGAACTAGAGACAGCCATGGGC-3′ and reverse,
5′-GGAATCTGTGACATCTCCAGCGCATCA-3′ (19). The primer sequences for the internal
control GAPDH were as follows: Forward,
5′-GGTGGTCTCCTCTGACTTCAACA-3′ and reverse,
5′-GTTGCTGTAGCCAAATTCGTTGT-3′. The reaction mixtures were incubated
for 60 min at 42°C followed by heating at 72°C for 5 min for
reverse transcription. The resulting cDNA (2 µg) was added to a 20
µl reaction mix containing 0.8 µl primers and 10 µl
SYBR®-Green Real Time PCR Master mix (Toyobo Co., Ltd.,
Osaka, Japan). RT-qPCR was performed using the ABI 7300 system
(Applied Biosystems; Thermo Fisher Scientific, Inc.). The
thermocycling conditions were set as follows: 40 cycles at 95°C for
3 min, 95°C for 15 sec and 60°C for 31 sec. The expression value of
S100P compared with that of GAPDH was determined using the
2−ΔΔCq method (20).
Western blotting
The cell lysates were prepared using
radioimmunoprecipitation assay buffer supplemented with
phenylmethylsulfonyl fluoride (Beyotime Institute of Biotechnology,
Shanghai, China), followed by centrifugation at 12,000 × g for 5
min at 4°C. Protein concentrations were estimated using the
bicinchoninic acid protein assay kit (Beyotime Institute of
Biotechnology), according to the manufacturer's protocol; 50 µg
protein was denatured in 1X loading buffer (Beyotime Institute of
Biotechnology) at 98°C for 5 min and separated by 12% SDS-PAGE, and
then electrotransferred onto nitrocellulose membranes with an
electroblotting apparatus (Bio-Rad Laboratories Inc., Hercules, CA,
USA). Western blot analysis was conducted using the primary
anti-human S100P antibodies (dilution, 1:1,000; Abcam; ab124743),
receptor for advanced glycation end products (RAGE; dilution,
1:1,000; Abcam; ab3611), epidermal growth factor receptor (EGFR;
dilution, 1:1,000; 18986-1-AP; ProteinTech Group Inc., Chicago, IL,
USA), cluster of differentiation (CD)44 (dilution, 1:1,000;
15675-1-AP; ProteinTech Group Inc.), matrix metalloproteinase
(MMP)2 (dilution, 1:1,000; 10373-2-AP; ProteinTech Group Inc.) and
MMP9 (dilution, 1:1,000; 10375-2-AP; ProteinTech Group Inc.). The
membranes were incubated overnight at 4°C with primary antibodies.
The secondary antibodies [Anti-rabbit IgG (heavy chain or light
chain binding)] goat; dilution, 1:10,000; 611-1302) conjugated with
horseradish peroxidase were incubated with the nitrocellulose
membrane at room temperature for 1 h. Subsequently, the membranes
were washed with Tris-buffered saline and Tween-20 and treated with
an enhanced chemiluminescence reagent (EMD Millipore, Billerica,
MA, USA). Immunoreactive bands were detected by exposure to X-ray
film. For quantification, the target protein was normalized to the
internal standard protein GAPDH through comparison of the gray
scale values. This analysis was performed using Gel-Pro Analyzer
software, version 4.0 (Media Cybernetics, Inc., Rockville, MD,
USA).
Cell proliferation assay
The proliferation of C666-1 cells transfected with
S100P siRNA or negative control siRNA was determined using a Cell
Counting kit-8 (CCK-8) assay (Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan). Triplicates of cell samples were seeded into
96-well plates at a density of 5×103 cells/well. At the
indicated time points (24, 48 and 72 h), 10 µl CCK-8 solution was
added to each well. Following incubation for 2 h at 37°C, the
absorbance was measured at 450 nm using a spectrophotometer (Thermo
Fisher Scientific, Inc.).
Colony formation assay
The C666-1 cells transfected with S100P siRNA or
negative control siRNA were seeded at a density of 1,000 cells/well
in a 6-well plate. The cells were cultured in 2 ml RPMI-1640
supplemented with 12% FBS and 1% P/S, and were incubated at 37°C
and 5% CO2 for 2 weeks. The culture medium was replaced every three
days. Upon completion of the incubation, the culture medium was
aspirated and the cells were washed with PBS, fixed in 1 ml
methanol for 30 min at room temperature and stained with 0.4%
crystal violet for 20 min at room temperature. The number of
colonies containing >50 cells was counted manually using an
inverted microscope (Olympus Corp., Tokyo, Japan) and averaged from
the duplicate wells.
Migration assay
The migration of the C666-1 cells transfected with
S100 siRNA or negative control siRNA was assessed using Transwell
cell culture chamber inserts (Corning Incorporated, Corning, NY,
USA) with an 8-µm pore size. A total of 1×104 cells that
had been treated with S100P siRNA or negative control siRNA for 24
h were seeded into Transwell filter membrane chambers with 100 µl
RPMI-1640 supplemented with 1% FBS. A total of 600 µl RPMI-1640
supplemented with 20% FBS was added to the lower compartment as a
chemoattractant. Following incubation for 20 h at 37°C in a
humidified 5% CO2 atmosphere, the cells on the upper surfaces of
the wells were removed with cotton swabs and the cells on the upper
surfaces that had migrated to the lower chamber, were fixed in cold
methanol at 4°C for 10 min, and stained with 0.4% crystal violet
for 10 min at room temperature. Excess stain was removed using
physiological saline and the chambers were air-dried at room
temperature for ~20 min. For each experiment, five fields on the
undersides of the membranes were randomly selected for imaging, and
the transmigrated cells in the five random fields were counted and
the mean was determined from the duplicate wells.
Wound healing assay
C666-1 cells transfected with S100P siRNA or
negative control siRNA for 24 h were seeded at a density of
2×106 single cells per well in a 6-well plate at 90–95%
confluency. The cell monolayer was scratched using a sterile 200 µl
pipette tip. The cells were rinsed with fresh medium [(RPMI-1640;
GE Healthcare Life Sciences; Hyclone) supplemented with 5% FBS
(Biowest)] to remove any free-floating cells and debris. Images
were captured immediately using an inverted microscope equipped
with a digital camera (Olympus) and wound healing was observed at
various time points (0, 12, 24 and 48 h) around the scrape line.
Images of representative scrape lines were captured. The wound area
was measured using Adobe Photoshop software version 7.0 (Adobe
Systems, Inc., San Jose, CA, USA).
Statistical analysis
The data were analyzed using SPSS version 18.0
(SPSS, Inc., Chicago, IL, USA). χ2 test and Yates'
correction were used to analyze the correlation between S100P
expression levels and the histological type, stage, age and gender
distribution. Kaplan-Meier analysis was used to analyze the
correlation between S100P immunoreactivity and overall and
disease-free survival. All the experiments were performed ≥3 times
and the results are presented as the mean ± standard deviation.
Statistical comparisons between two groups of data were performed
using the two-tailed Student's t-test. Multiple group comparisons
were performed using one-way analysis of variance. P<0.05 was
considered to indicate a statistically significant difference.
Results
The clinical and pathological
significance of S100P expression levels in nasopharyngeal
cancer
To investigate the clinical and pathological
significance of S100P expression levels in nasopharyngeal cancer,
the tissues of 78 patients with NPC and 30 patients with benign
inflammation were analyzed using IHC. Intense positive nuclear
staining was observed in all the neoplastic cells in the positive
controls. The tissue specimens from the patients with NPC exhibited
diffuse or focal positive nuclear staining. Fig. 1A and B present the typical
nasopharyngeal carcinoma cells. Immunoreactivity was more
pronounced in the basal layer of the normal and dysplastic
epithelium (Fig. 1C, D, E and F). Of
a total of 78 NPC samples, positive staining for the S100P protein
was observed in 45 (57.7%) tissue samples, whereas no staining was
detected in 33 (42.3%) tissue samples. No S100P staining was
observed in the 30 benign inflammation tissues (Fig. 1G and H). The correlation between S100P
expression levels and the histological type, stage, age and sex
distribution was examined (Table I),
and no significant differences were observed according to the
χ2 test and Yates' correction (P>0.05). The median
follow-up time was 63.7 months (range, 13–93 months), and the
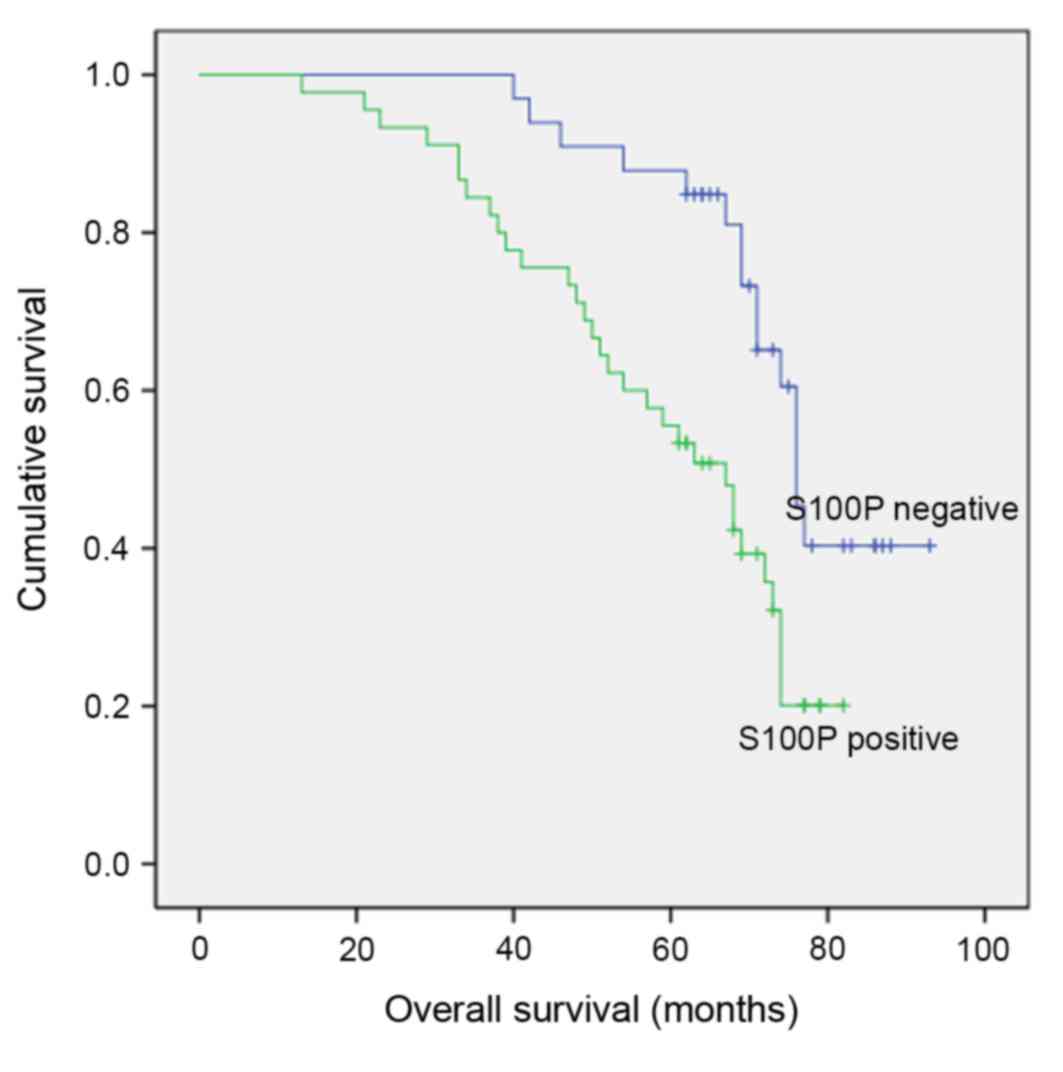
5-year cumulative survival rate was 68.3±2.5% standard error.
According to Kaplan-Meier analysis, a statistically significant
correlation was observed between S100P immunoreactivity and overall
and disease-free survival (Fig. 2;
P<0.01).
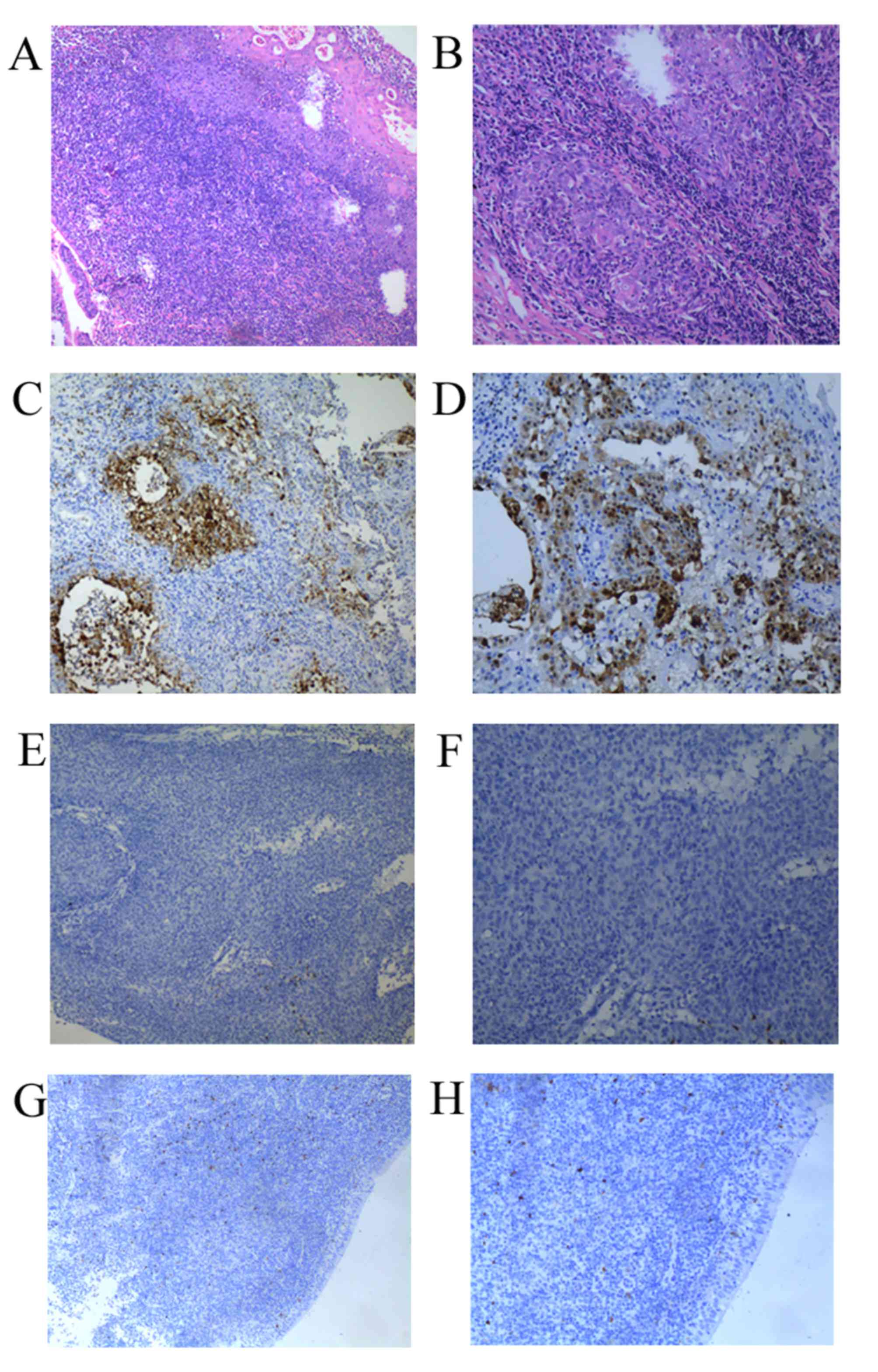 | Figure 1.Hematoxylin and eosin staining in NPC
tissues. (A) S100P positive staining in NPC (magnification, ×100).
(B) S100P positive staining in NPC (magnification, ×200). (C) S100P
negative staining in NPC (magnification, ×100). (D) S100P negative
staining in NPC (magnification, ×200). (E) S100P negative staining
in benign inflammation (magnification, ×100). (F) S100P negative
staining in benign inflammation (magnification, ×200). (G)
magnification, ×100; (H) magnification, ×200). NPC, nasopharyngeal
carcinoma; S100P, S100 calcium binding protein P. |
Knockdown of S100P decreases cell
proliferation and invasion in C666-1 cells
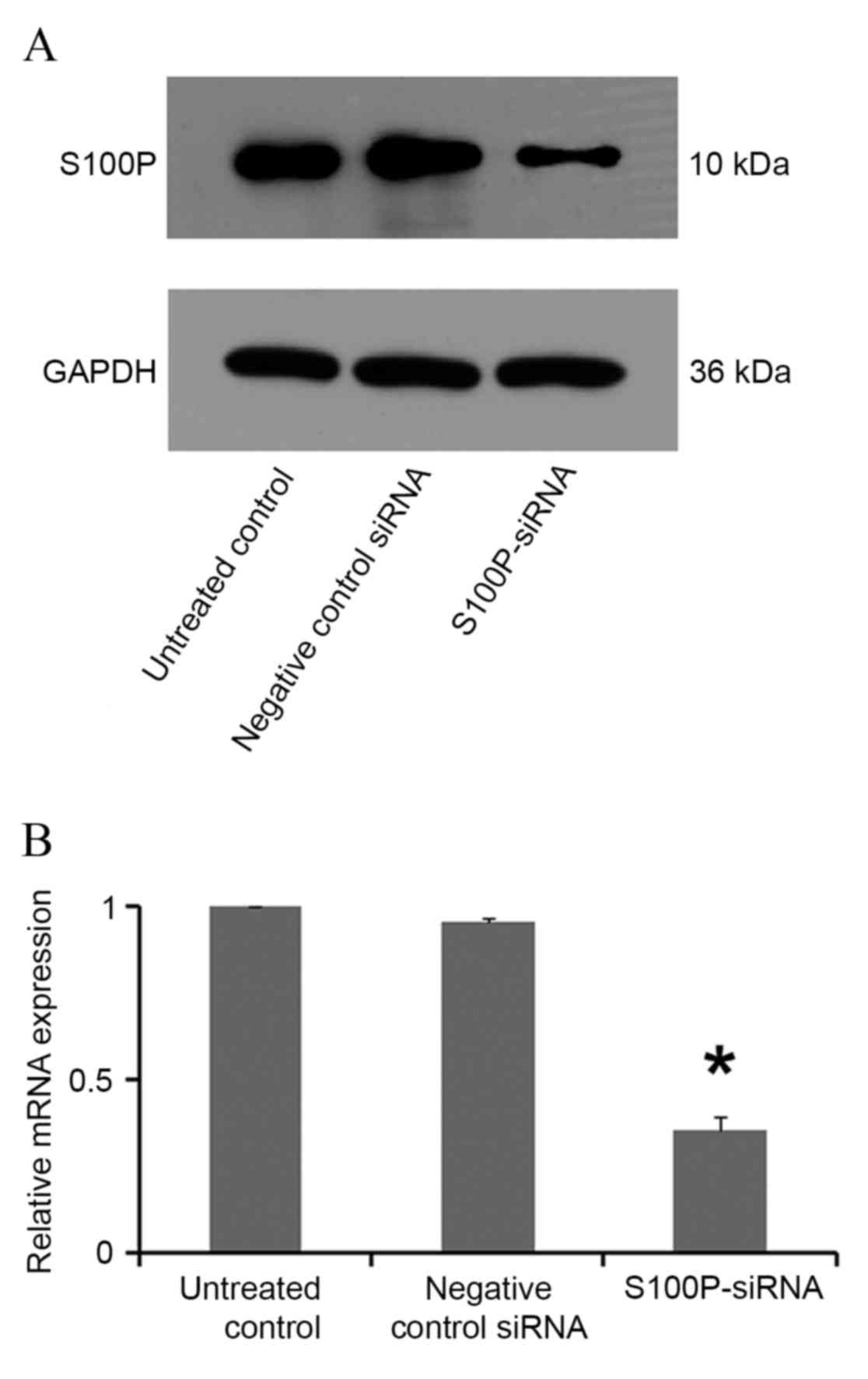
In siRNA S100P transfected C666-1 cells, decreased
levels of S100P mRNA and protein were observed using RT-qPCR and
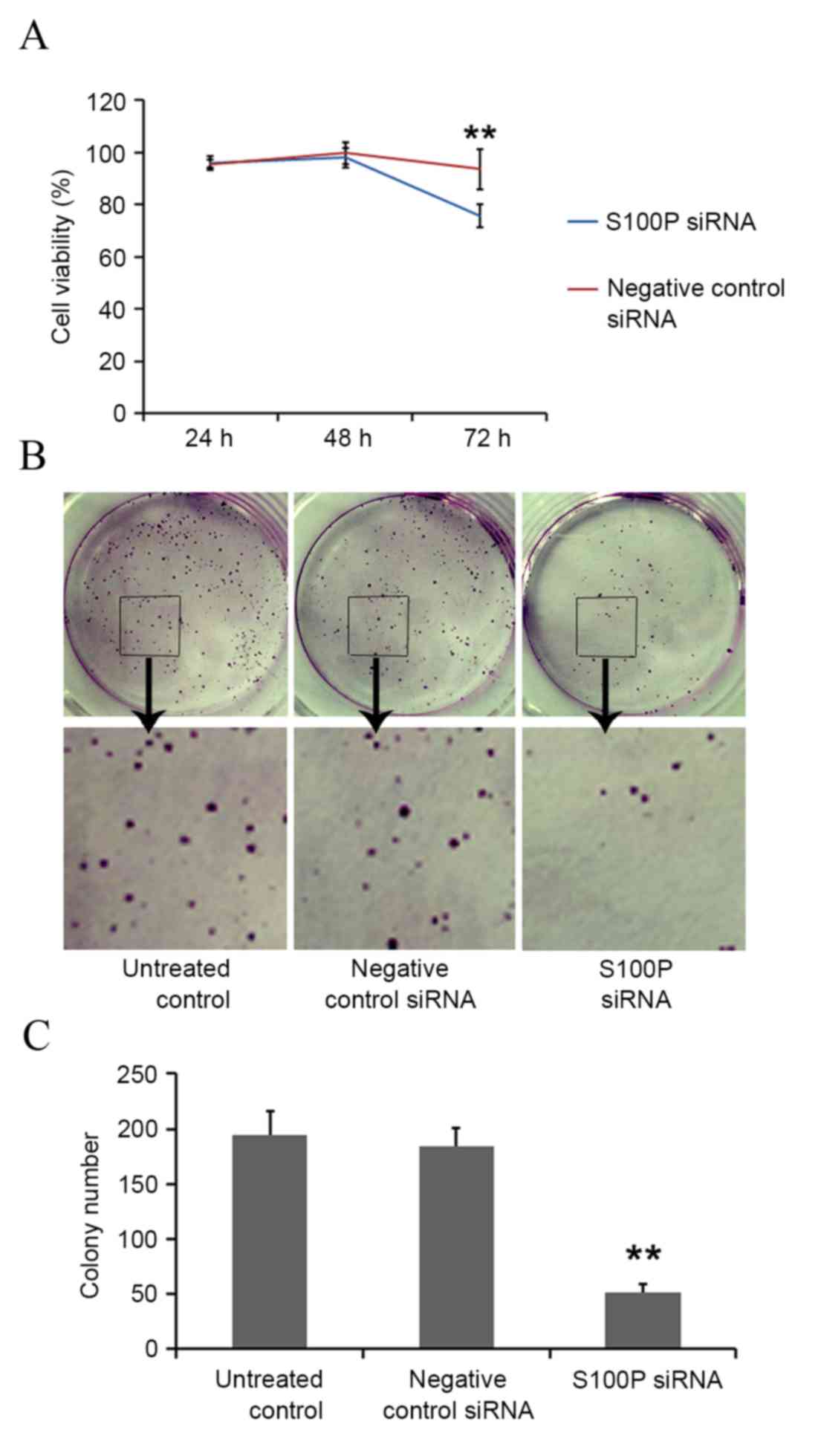
western blot analyses, respectively (P<0.001; Fig. 3). Cell growth, colony formation and
migration assays were performed. S100P knockdown produced no
significant effects on cell proliferation at 24 (P<0.622) and 48
h (P<0.245); however, cell proliferation was significantly
decreased at 72 h (P<0.001) in the S100P siRNA group, compared
with the negative control siRNA group and the untreated control
group (Fig. 4A). S100P knockdown
reduced the colony forming ability of the S100P siRNA transfected
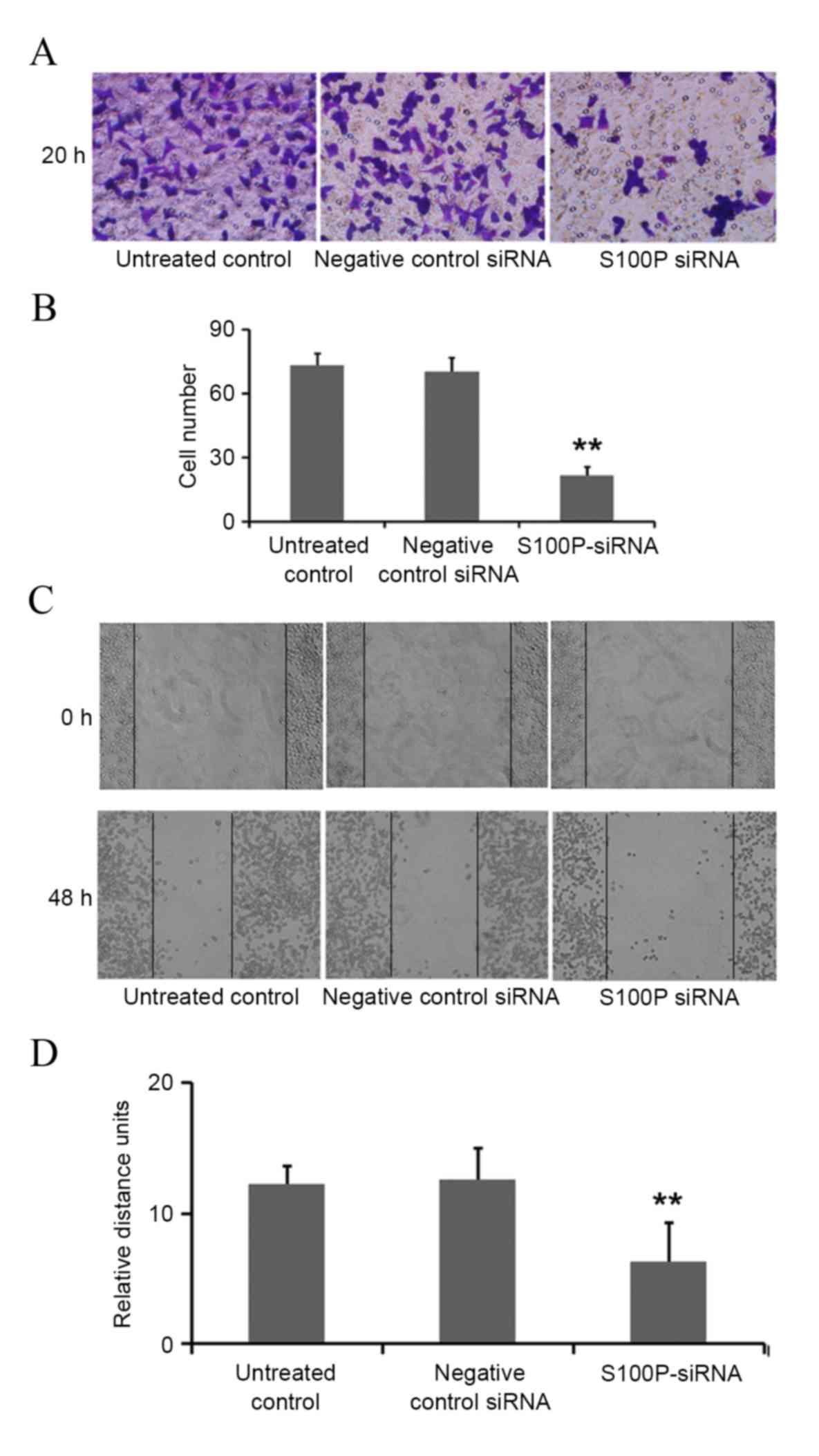
cells (Fig. 4B and C). Additionally,
the effects on C666-1 cell migration were assessed by Transwell and
wound healing assays, revealing that the knockdown of S100P
significantly reduced the invasive ability of cells (P<0.001;
Fig. 5A and B). As presented in
Fig. 5C and D (P<0.001),
photomicrographs captured at 48 h following wounding of the cell
monolayer revealed delayed wound closure in cells transfected with
S100P siRNA, compared with cells transfected with negative control
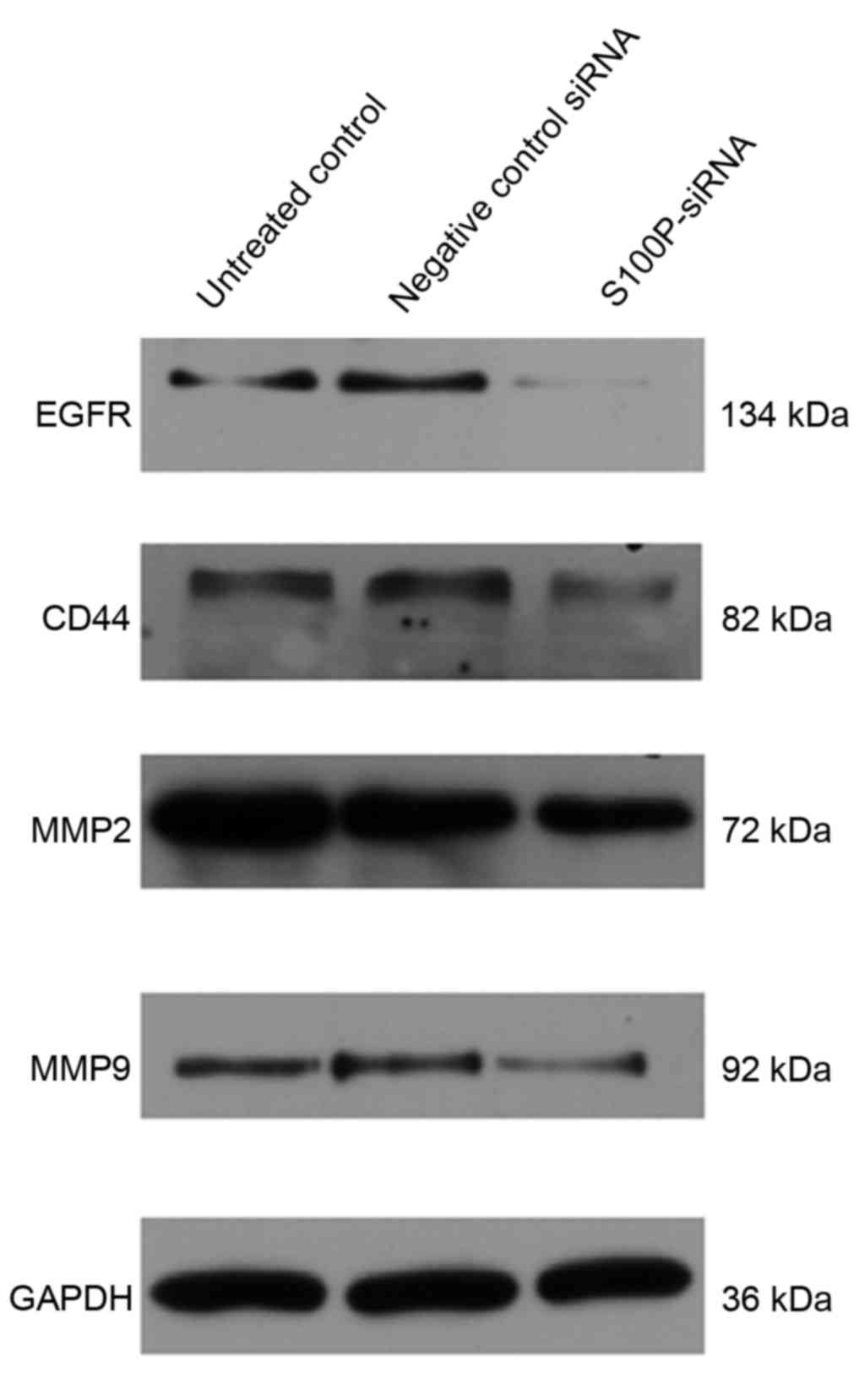
siRNA and the untreated controls. The EGFR, CD44, MMP2 and MMP9
proteins are important during cancer cell invasion and metastasis
(21–23). Therefore, the protein expression
levels of EGFR, CD44, MMP2 and MMP9 in C666-1 cells that had been
treated with S100P or negative control siRNA were assessed using
western blotting. The results revealed that the expression levels
of these proteins were decreased in the S100P siRNA group, compared
with the negative control siRNA and untreated control groups
(Fig. 6; EGFR, P<0.01; CD44,
P<0.01; MMP2, P<0.001; MMP9, P<0.01). Taken together,
these results indicate that a reduction in the level of S100P
expression is able to suppress the growth and invasion of C666-1
cells. These functions support our hypothesis that the regulation
of S100P expression is an important step by which cancer cells are
able to promote their growth and migration during
carcinogenesis.
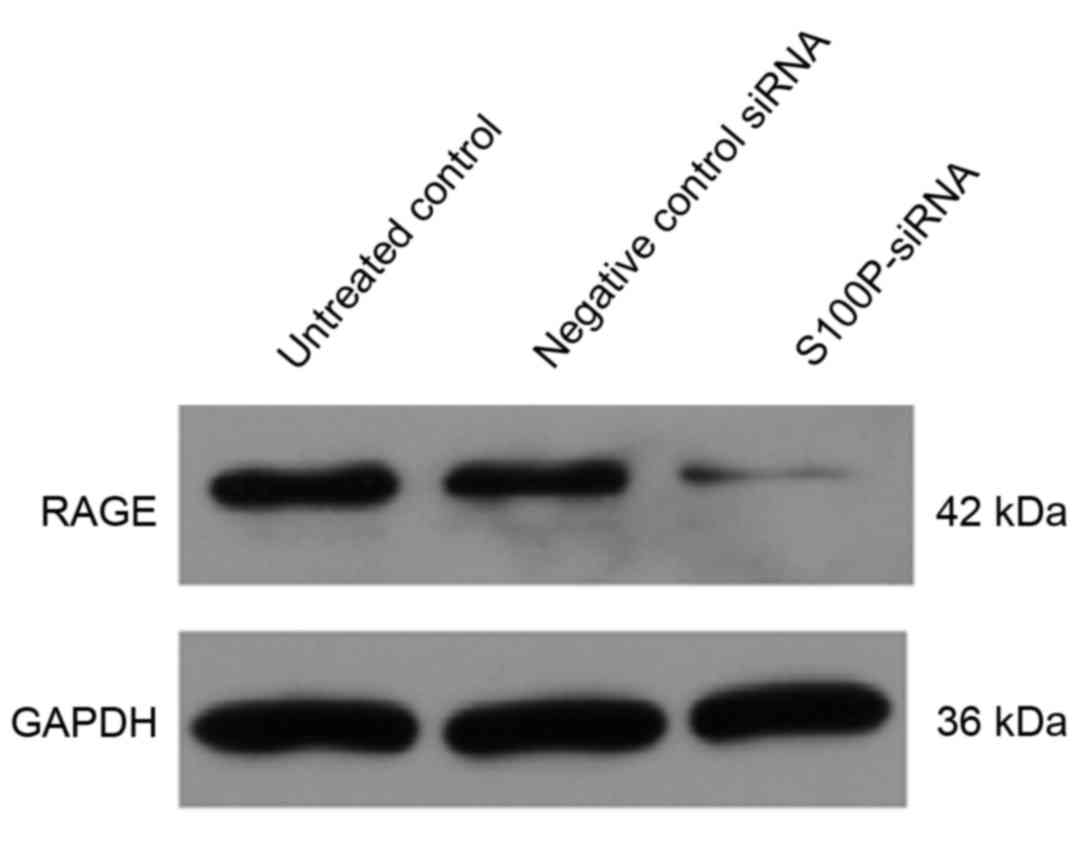
Extracellular S100 proteins act via a variety of
cellular receptors (24,25) and RAGE has been suggested to be a
general receptor for the S100 family of proteins (26). Using western blot analysis, the
present study observed that RAGE expression was downregulated in
the S100P siRNA group, compared with the negative siRNA control and
untreated control groups (Fig. 7;
P<0.001).
Discussion
The overexpression of S100P, which was first
purified from the human placenta, has been detected in a variety of
types of cancer, and correlates with tumor growth and metastasis
(19,27). Several studies have linked S100P
expression levels to cell proliferation, invasion and migration in
various forms of cancer (28).
To investigate the clinical and pathological
significance of S100P expression in nasopharyngeal cancer, the
expression levels of the S100P protein in human nasopharyngeal
tissues were evaluated using IHC. The results revealed
significantly increased S100P expression levels in 45/78 (57.7%)
patients with NPC. In addition, a significant correlation between
S100P immunoreactivity and overall disease-free survival was
observed.
A number of in vitro and in vivo
studies have evaluated the effects of the silencing of genes
involved in certain signaling pathways that promote cancer
progression, including oncogenesis, apoptosis, cell cycle
regulation, cell senescence, tumor-host interactions and resistance
to conventional therapies (29,30). In
the present study, the potential effects of S100P on the
characteristics of nasopharyngeal cancer were investigated, and
siRNA was used to silence S100P expression in C666-1 cells. The
results revealed that S100P protein and mRNA expression levels were
significantly reduced in the S100P siRNA-transfected group. Cell
growth assays were performed to evaluate the association between
S100P expression levels and C666-1 cell proliferation. The results
from the CCK-8 and colony formation assays revealed that S100P
silencing decreased the proliferative ability of C666-1 cells,
which suggests that S100P may be a tumor growth-associated gene in
NPC.
Cancer metastasis is a multistep process involving
migration and invasion through the tumor stroma, intravasation,
tumor cell dissemination, extravasation and cell growth at the
metastatic sites (31). The present
study investigated the migratory characteristics of C666-1 cells,
following S100P silencing, using Transwell and wound-healing
assays. The migration of C666-1 cells was observed to be
significantly reduced when S100P expression was inhibited by siRNA.
Furthermore, the expression levels of EGFR, CD44, MMP2 and MMP9
were decreased significantly when S100P was silenced, suggesting
that S100P is associated with cell migration in NPC. EGFR is a type
I receptor tyrosine kinase that is overexpressed in a number of
solid tumors, including types of head and neck carcinoma, and is
associated with a poor prognosis following treatment (32). CD44 is a cell-surface molecule that
has been implicated in a diverse range of cell-cell and cell-matrix
interactions (33). MMPs, a family of
zinc-binding endopeptidases, possess an established association
with cancer-cell invasion and metastasis (34). EGFR, CD44 and MMPs are therefore key
factors during cancer development, progression and metastasis, and
downregulation of the expression of these factors, as a result of
decreased S100P expression levels, suggests a correlation between
S100P expression levels and C666-1 cell proliferation and migration
(35).
RAGE, a member of the immunoglobulin protein family
of cell surface molecules (36),
shares structural homology with other immunoglobulin-like receptors
(37) and is important in certain
human pathologies, including cancer (38). The present study analyzed the
expression levels of RAGE protein in S100P siRNA transfected C666-1
cells using western blot analysis. The results revealed that RAGE
expression was downregulated in the S100P siRNA group, as compared
with the negative siRNA control and untreated control groups. A
number of S100 proteins interact with RAGE in vitro and
trigger RAGE-dependent signaling in cell-based assays. For
instance, Arumugam et al (39)
demonstrated that S100P is able to trigger the activation of
nuclear factor-κB via the mitogen activated protein kinase
signaling pathway in a RAGE-dependent manner in BxPC3 and SW480
adenocarcinoma cells. The present study revealed that the
siRNA-mediated knockdown of S100P expression in C666-1 cells
significantly inhibits cancer cell growth, migration and invasion
in vitro. Further studies may investigate the role of S100P
by examining its effects on tumor growth and metastasis in
vivo.
In conclusion, the results of the present study
suggested that S100P may be an important regulatory protein during
the promotion of NPC cell proliferation and migration.
Additionally, the current study provided information about the
function of proliferation and migration of S100P in C666-1 cells
that may aid the elucidation of the molecular mechanisms underlying
tumor metastasis and the identification of clinically relevant
biomarkers for metastasis prevention in NPC.
Acknowledgements
The present study was supported by the Basic
Research Project of The Shanghai Technology Commission (grant no.
12JC1411100), The Key Disciplines Group Construction Project of the
Pudong Health Bureau of Shanghai (grant no. PWZxq 2014–09) and The
Project of The Shanghai Technology Commission (grant nos.
14DZ1940103 and 14411960400).
References
|
1
|
Marenholz I, Heizmann CW and Fritz G: S100
proteins in mouse and man: From evolution to function and pathology
(including an update of thenomenclature). Biochem Biophys Res
Commun. 322:1111–1122. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Heizmann CW, Fritz G and Schäfer BW: S100
proteins: Structure, functions and pathology. Front Biosci.
7:d1356–d1368. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ragin CC, Modugno F and Gollin SM: The
epidemiology and risk factors of head and neck cancer: A focus on
human papillomavirus. J Dent Res. 86:104–114. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zha C, Jiang XH and Peng SF: iTRAQ-based
quantitative proteomic analysis on S100 calcium binding protein A2
in metastasis of laryngeal cancer. PLoS One. 10:e01223222015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Moriyama-Kita M, Endo Y, Yonemura Y,
Heizmann CW, Schäfer BW, Sasaki T and Yamamoto E: Correlation of
S100A4 expression with invasion and metastasis in oral squamous
cell carcinoma. Oral Oncol. 40:496–500. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nakashima T, Yano G, Hayashi I and Katsuta
Y: Epithelial membrane antigen and S-100 protein-labeled cells in
primary and metastatic laryngeal carcinomas. Head Neck. 14:445–451.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang C, Zhang Z, Li L, Zhang J, Wang J,
Fan J, Jiao B and Zhao S: S100A11 is a migration-related protein in
laryngeal squamous cell carcinoma. Int J Med Sci. 10:1552–1559.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Moriyama-Kita M, Endo Y, Yonemura Y,
Heizmann CW, Schäfer BW, Sasaki T and Yamamoto E: Correlation of
S100A4 expression with invasion and metastasis in oral squamous
cell carcinoma. Oral Oncol. 401:496–500. 2004. View Article : Google Scholar
|
|
9
|
Sung FL, Poon TC, Hui EP, Ma BB, Liong E,
To KF, Huang DP and Chan AT: Antitumor effect and enhancement of
cytotoxic drug activity by cetuximab in nasopharyngeal carcinoma
cells. In Vivo. 19:237–245. 2005.PubMed/NCBI
|
|
10
|
Paiar F, Di Cataldo V, Zei G, Pasquetti
EM, Cecchini S, Meattini I, Mangoni M, Agresti B, Iermano C, Bonomo
P and Biti G: Role of chemotherapy in nasopharyngeal carcinoma.
Oncol Rev. 6:e12012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lu H, Peng L, Yuan X, Hao Y, Lu Z, Chen J,
Cheng J, Deng S, Gu J, Pang Q and Qin J: Concurrent
chemoradiotherapy in locally advanced nasopharyngeal carcinoma: A
treatment paradigm also applicable to patients in Southeast Asia.
Cancer Treat Rev. 35:345–353. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Baujat B, Audry H, Bourhis J, Chan AT,
Onat H, Chua DT, Kwong DL, Al-Sarraf M, Chi KH, Hareyama M, et al:
Chemotherapy in locally advanced nasopharyngeal carcinoma: An
individual patient data meta-analysis of eight randomized trials
and 1,753 patients. Int J Radiat Oncol Biol Phys. 64:47–56. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Becker T, Gerke V, Kube E and Weber K:
S100P, a novel Ca(2+)-binding protein from human placenta. cDNA
cloning, recombinant protein expression and Ca2+ binding
properties. Eur J Biochem. 207:541–547. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Da Silva ID Guerreiro, Hu YF, Russo IH, Ao
X, Salicioni AM, Yang X and Russo J: S100P calcium-binding protein
overexpression is associated with immortalization of human breast
epithelial cells in vitro and early stages of breast cancer
development in vivo. Int J Oncol. 16:231–240. 2000.PubMed/NCBI
|
|
15
|
Beer DG, Kardia SL, Huang CC, Giordano TJ,
Levin AM, Misek DE, Lin L, Chen G, Gharib TG, Thomas DG, et al:
Gene-expression profiles predict survival of patients with lung
adenocarcinoma. Nat Med. 8:816–824. 2002.PubMed/NCBI
|
|
16
|
Shyu RY, Huang SL and Jiang SY: Retinoic
acid increases expression of the calcium-binding protein S100P in
human gastric cancer cells. J Biomed Sci. 10:313–319. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Basu GD, Azorsa DO, Kiefer JA, Rojas AM,
Tuzmen S, Barrett MT, Trent JM, Kallioniemi O and Mousses S:
Functional evidence implicating S100P in prostate cancer
progression. Int J Cancer. 123:330–339. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cheung ST, Huang DP, Hui AB, Lo KW, Ko CW,
Tsang YS, Wong N, Whitney BM and Lee JC: Nasopharyngeal carcinoma
cell line (C666-1) consistently harbouring Epstein-Barr virus. Int
J Cancer. 83:121–126. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Arumugam T, Simeone DM, Van Golen K and
Logsdon CD: S100P promotes pancreatic cancer growth, survival, and
invasion. Clin Cancer Res. 11:5356–5364. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wobus M, Rangwala R, Sheyn I, Hennigan R,
Coila B, Lower EE, Yassin RS and Sherman LS: CD44 associates with
EGFR and erbB2 in metastasizing mammary carcinoma cells. Appl
Immunohistochem Mol Morphol. 10:34–39. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sun Y, Liu M, Yang B, Li B and Lu J: Role
of siRNA silencing of MMP-2 gene on invasion and growth of
laryngeal squamous cell carcinoma. Eur Arch Otorhinolaryngol.
265:1385–1391. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen X, Qiu J, Yang D, Lu J, Yan C, Zha X
and Yin Y: MDM2 promotes invasion and metastasis in invasive ductal
breast carcinoma by inducing matrix metalloproteinase-9. PLoS One.
8:e787942013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gao H, Zhang X, Zheng Y, Peng L, Hou J and
Meng H: S100A9-induced release of interleukin (IL)-6 and IL-8
through toll-like receptor 4 (TLR4) in human periodontal ligament
cells. Mol Immunol. 67:223–232. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen B, Miller AL, Rebelatto M, Brewah Y,
Rowe DC, Clarke L, Czapiga M, Rosenthal K, Imamichi T, Chen Y, et
al: S100A9 induced inflammatory responses are mediated by distinct
damage associated molecular patterns (DAMP) receptors in vitro and
in vivo. PLoS One. 10:e01158282015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Leclerc E, Fritz G, Vetter SW and Heizmann
CW: Binding of S100 proteins to RAGE: An update. Biochim Biophys
Acta. 1793:993–1007. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Parkkila S, Pan PW, Ward A, Gibadulinova
A, Oveckova I, Pastorekova S, Pastorek J, Martinez AR, Helin HO and
Isola J: The calcium-binding protein S100P in normal and malignant
human tissues. BMC Clin Pathol. 8:22008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jiang L, Lai YK, Zhang J, Wang H, Lin MC,
He ML and Kung HF: Targeting S100P inhibits colon cancer growth and
metastasis by Lentivirus-mediated RNA interference and proteomic
analysis. Mol Med. 17:709–716. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dykxhoorn DM and Lieberman J: Knocking
down disease with siRNAs. Cell. 126:231–235. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sy SM, Wong N, Lee TW, Tse G, Mok TS, Fan
B, Pang E, Johnson PJ and Yim A: Distinct patterns of genetic
alterations in adenocarcinoma and squamous cell carcinoma of the
lung. Eur J Cancer. 40:1082–1094. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bravo-Cordero JJ, Hodgson L and Condeelis
J: Directed cell invasion and migration during metastasis. Curr
Opin Cell Biol. 24:277–283. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ang KK, Berkey BA, Tu X, Zhang HZ, Katz R,
Hammond EH, Fu KK and Milas L: Impact of epidermal growth factor
receptor expression on survival and pattern of relapse in patients
with advanced head and neck carcinoma. Cancer Res. 62:7350–7356.
2002.PubMed/NCBI
|
|
33
|
Wobus M, Rangwala R, Sheyn I, Hennigan R,
Coila B, Lower EE, Yassin RS and Sherman LS: CD44 associates with
EGFR and erbB2 in metastasizing mammary carcinoma cells. Appl
Immunohistochem Mol Morphol. 10:34–39. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Egeblad M and Werb Z: New functions for
the matrix metalloproteinases in cancer progression. Nat Rev
Cancer. 2:161–174. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kivisaari AK, Kallajoki M, Ala-aho R,
McGrath JA, Bauer JW, Königová R, Medvecz M, Beckert W, Grénman R
and Kähäri VM: Matrix metalloproteinase-7 activates heparin-binding
epidermal growth factor-like growth factor in cutaneous squamous
cell carcinoma. Br J Dermatol. 163:726–735. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Neeper M, Schmidt AM, Brett J, Yan SD,
Wang F, Pan YC, Elliston K, Stern D and Shaw A: Cloning and
expression of a cell surface receptor for advanced glycosylation
end products of proteins. J Biol Chem. 267:14998–15004.
1992.PubMed/NCBI
|
|
37
|
Barclay AN: Membrane proteins with
immunoglobulin-like domains-a master superfamily of interaction
molecules. Semin Immunol. 15:215–223. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Schmidt AM, Yan SD, Yan SF and Stern DM:
The biology of the receptor for advanced glycation end products and
its ligands. Biochim Biophys Acta. 1498:99–111. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Arumugam T, Simeone DM, Schmidt AM and
Logsdon CD: S100P stimulates cell proliferation and survival via
receptor for activated glycation end products (RAGE). J Biol Chem.
279:5059–5065. 2004. View Article : Google Scholar : PubMed/NCBI
|