Introduction
Triple negative breast cancer (TNBC) accounts for
approximately 15% of all breast cancers. This subset of breast
cancer is defined by the absence of positive staining for estrogen
receptor (ER) and progesterone receptor (PR) and lack amplification
of HER2. Molecular profiling of TNBC has revealed that most TNBC
have a subset of gene expression patterns that are associated to
basal-myoepithelial cells in the breast. TNBC is more likely to
affect younger women, women of African American descent, women who
were exposed to radiation at an early age and BRCA1 mutation
carriers (1–3). TNBCs are considered aggressive tumors
with a high degree of genomic instability and therefore usually
present as high grade, large tumors with a high proliferative index
(4).
Due to the aggressive nature of TNBC tumors and
decreased efficacy of targeted chemotherapy, TNBCs are associated
with poor prognosis and early visceral metastasis (5). Survival rates for women who have a
systemic or local recurrence within 3–5 years of treatment (early
recurrence) are significantly lower survival rates for hormone
receptor positive (ER/PR+) breast cancer, while those who recur
after 5 years have survival rates similar to ER/PR+ cancers
(4). Understanding the molecular
mechanisms that drive growth of TNBC may lead to better treatment,
as well as predictive biomarkers that may identify women at risk
for aggressive disease. TNBCs represent a collection of subtypes,
some of which are associated with rapid progression, while others
are reported as being less aggressive. Therefore, there is a
clinical need to determine the molecular differences between these
subtypes of TNBC patients in order to provide the most effective
treatment; however a reliable prognostic marker has not been
identified.
Molecular pathways crucial in embryonic development,
including the Hedgehog (HH) and Wnt signaling pathways have been
shown to play an important role in breast cancer development and
progression. Abnormal regulation of these pathways can lead to
activation of genes essential for cell proliferation, cell
survival, and therapeutic resistance (6,7). In
addition, activation of HH and Wnt signaling have been implicated
in the growth and resistance of cancer stem cells and maintenance
of the stem cell niche (8–12). These pathways are also key regulators
of genes controlling epithelial-mesenchymal transition (EMT),
contributing to cancer invasion and metastasis (13,14).
The HH signaling pathway is critical for growth and
differentiation during embryonic development. Initiation of the
pathway requires secreted HH molecules [Sonic HH (SHH), Desert and
Indian] to bind to and inhibit the cell surface HH receptor,
Patched (PTCH). This binding relieves the PTCH-mediated suppression
of the transmembrane protein smoothened (SMO), leading to multiple
intracellular events that result in the stabilization, nuclear
translocation and activation of the glioma-associated oncogene
(Gli) family of transcription factors, which initiate transcription
of HH target genes (15). However,
aberrant HH signaling has been shown to be associated with
malignant transformation in many tissues, including breast
(16–19). It has been reported that protein
levels of SHH, PTCH, and Gli-1 in the breast tumor are
significantly elevated compared to the adjacent normal breast ducts
(20,21). Overactivation of HH signaling is
thought to result in increased number of mammary stem cells,
resulting in tumor formation (10).
However, there are a limited number of studies on how the HH
pathway relates to patient prognosis. Inactivation of HH through
high expression of Ptch in patients with invasive ductal breast
carcinoma was associated with a favorable prognosis (22). Additionally, patients with
inflammatory breast cancer had elevated SHH expression, which was
associated with poor prognosis (23).
The Wnt signaling pathway is a complex signaling
network involved in many physiological processes, including tissue
patterning, cell migration, EMT, and maintenance of stem cells
(24–26). Canonical Wnt signaling involves
stabilization of β-catenin and translocation of the protein to the
nucleus resulting in activation of Wnt target genes. In the absence
of Wnt ligand, β-catenin is in a multi-protein, cytoplasmic complex
with axin and adenomatous polyposis coli (APC). Upon
phosphorylation by glycogen synthase kinase-3 (GSK-3β) and casein
kinase-1β, β-catenin is degraded (24,26,27). In
the presence of canonical Wnt proteins, Wnt binds to the receptors,
Frizzled and lipoprotein receptor-related proteins (LRP) 5 and 6,
resulting in activation of Dishevelled. This in turn disrupts the
β-catenin-APC-axin complex, blocking degradation of β-catenin and
allowing accumulation and translocation of β-catenin to the nucleus
where it forms an activation complex with the T-cell
factor/lymphoid enhancing factor (Tcf/LEF) family of transcription
factors, leading to expression of genes critical for development of
cell transformation and cancer, including cyclin D1, c-Myc, and
peroxisome-proliferator-activated receptor-β (PPAR-β) (26–28). Only
canonical Wnt signaling drives β-catenin mediated transcription and
thus nuclear β-catenin is considered an indicator of Wnt activation
(29).
Aberrant canonical Wnt signaling has been shown in
many tumors, including breast (9,30–32). In mouse models, overexpression of a
β-catenin mutant and mutations of the Apc gene have resulted
in mammary tumorigenesis (33). Human
breast cancer cell lines show higher amplification of canonical Wnt
genes (34,35) concomitant with downregulation of
non-canonical Wnt gene s (35) and a
higher expression of Dishevelled (36). Normally in the human breast duct and
lobules, β-catenin is localized at the cell membrane, bound to
E-cadherin and not part of Wnt signaling (25,31,33,37–39).
Studies have reported a complete loss of β-catenin at the cell
membrane in invasive lobular breast carcinomas consistent with a
loss of E-cadherin; however, localization of β-catenin in the cell
was not reported in these studies (37,38). In
addition, in invasive ductal carcinoma, a loss of membranous
β-catenin correlates with increased expression of canonical Wnt
signaling target genes, suggesting activation of the canonical Wnt
pathway and this correlates with poorer outcomes in breast cancer
patients (31). However, others have
shown that a loss of membranous β-catenin staining is not
correlated with tumor stage, grade, or outcome (37). Therefore, how activation of the
Wnt/β-catenin signaling pathway correlates with breast cancer
patient prognosis is not completely clear.
Independently, HH and Wnt signaling pathways play a
role in the progression of breast cancer and therefore the
crosstalk between these developmental pathways is thought to
provide tumor cells with multiple mechanisms to evade chemotherapy.
Current research has shown a potential for crosstalk between HH and
Wnt signaling in cancer. Co-activation of HH and Wnt signaling has
been exhibited in a variety of cancers, including basal cell
carcinoma and pancreatic ductal adenocarcinoma (40). In contrast, it has also been shown
that the activity of the Wnt/β-catenin pathway can be suppressed by
players of the HH pathway (41,42).
However, the impact of HH and Wnt signaling crosstalk in breast
cancer outcomes is poorly understood. Therefore, we wanted to
determine if co-activation of the HH signaling pathway and
canonical Wnt pathway could be used as a prognostic marker in TNBC
patients.
Materials and methods
Patient population
Breast cancer tissue specimens were obtained from
the biorepository at the Helen F. Graham Cancer Center and Research
Institute (HFGCCRI) under protocol approved by the institutional
review board. De-identified samples were obtained from surgical
resection from women diagnosed with TNBC who consented to the use
of their tissues for research. Tissue blocks were prepared by
formalin-fixation and embedded in paraffin for serial sectioning.
Each sample underwent a pathological diagnostic procedure including
staining for the ER, PR and HER2 expression. HER2 status was
further confirmed by FISH. Pathologically confirmed TNBC (ER
expression <1.0%) samples were used in this study. Hematoxylin
and eosin (H&E) stains of tumors sections were reviewed by a
breast cancer pathologist to determine the percentage of tumor
nuclei and necrosis. Only samples containing >60% tumor nuclei
were used for this study.
Immunohistochemical procedure
Immunohistochemical staining was performed on 4 µm
thick tissue sections of each specimen using the LSAB+System-HRP
staining kit (Dako; Agilent Technologies, Inc., Santa Clara, CA,
USA) according to the manufacturer's instructions as previously
described (43). Antibodies to SHH
(cloneEP1190Y; Abcam, Cambridge, UK; and H160-sc9026; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA), Gli-1 (Clone H-300; Santa
Cruz Biotechnology, Inc.), and β-catenin (cloneE247; Abcam) were
used at a dilution of 1:100 for SHH, 1:1,000 for Gli-1 and 1:500
for β-catenin. Both SHH antibodies showed a similar staining
pattern. Specificity of staining was confirmed by omission of the
primary antibody and staining with an isotype matched control
antibody (Jackson Laboratory, Ben Harbor, ME, USA). Slides were
scored by two independent investigators blinded to the sample data.
Slides were scored as having no expression (0), weak (1), moderate (2), or strong (3) tumor cell staining. Nuclear staining was
noted if nuclear stain was observed in more than 10% of cells in 3
fields at ×40 magnification.
Statistical analysis
Spearman correlations were used to test the
relationships between SHH, Gli-1, and β-catenin staining and
clinical characteristics. Due to the small sample size, samples
were grouped into three categories based on activation of HH and
Wnt pathways for survival analysis: i) Those with activation of
neither pathway (no staining for nuclear Gli-1 and nuclear
β-catenin), n=8; ii) only one pathway activated, staining for
either nuclear Gli-1 or nuclear β-catenin, n=8; and iii) those with
staining for nuclear expression of both pathways, n=20. These
groups were then compared using chi-square tests for survival and
recurrence. Lastly, Kaplan-Meier Survival Curves were presented to
show the detrimental effect of dual activation of HH and Wnt
signaling on outcome. Due to the small sample size, multi-variate
analysis was not performed.
Results
Patient cohort
This study comprised a cohort of 36 tumors from
women with histologically confirmed TNBC. A summary of clinical
data is provided in Table I. This
cohort included 21 patients with early stage disease (58%, stage
1-2A) and 15 patients with late stage disease (42%, stage 2B-4).
The majority of patients presented with high grade tumors (grade 3,
86%) as is common in TNBC. Five patients (14%) had grade 2
cancers.
 | Table I.Patient characteristics and
expression of HH and WNT pathway members. |
Table I.
Patient characteristics and
expression of HH and WNT pathway members.
|
Characteristics | No. | % | P-value stage | P-value RFS | P-value OS |
|---|
| Stage |
|
|
|
|
|
| 1A | 6 | 16.7 |
|
|
|
| 1B | 1 |
2.8 |
|
|
|
| 2A | 14 | 38.9 |
|
|
|
| 2B | 4 | 11.1 |
|
|
|
| 3A | 4 | 11.1 |
|
|
|
| 3B | 0 |
0.0 |
|
|
|
| 3C | 5 | 13.9 |
|
|
|
| 4 | 2 |
5.6 |
| 0.0055 | 0.0036 |
| Grade |
|
|
|
|
|
| 2 | 5 | 13.9 |
|
|
|
| 3 | 31 | 86.1 |
0.3 | 0.732 | 0.985 |
| SHH |
|
|
|
|
|
| 1 | 8 | 22.2 |
|
|
|
| 2 | 20 | 55.6 |
|
|
|
| 3 | 8 | 22.2 | 0.0440 | 0.051 | 0.026 |
| Gli-1 |
|
|
|
|
|
| 0 | 1 |
2.8 |
|
|
|
| 1 | 9 | 25.0 |
|
|
|
| 2 | 14 | 38.9 |
|
|
|
| 3 | 12 | 33.3 | 0.0010 | 0.0213 | 0.017 |
| Nuclear
Gli-1 |
|
|
|
|
|
|
<10% | 11 | 30.6 |
|
|
|
|
>10% | 25 | 69.4 | 0.0080 | 0.003 | 0.0387 |
| β-catenin |
|
|
|
|
|
|
Membranous | 13 | 36.1 |
|
|
|
|
Nuclear/cytoplasmic | 23 | 63.9 | 0.0030 | 0.019 | 0.025 |
Expression of stem cell pathways and
relationship to clinical characteristics
Serial sections were stained for expression of SHH,
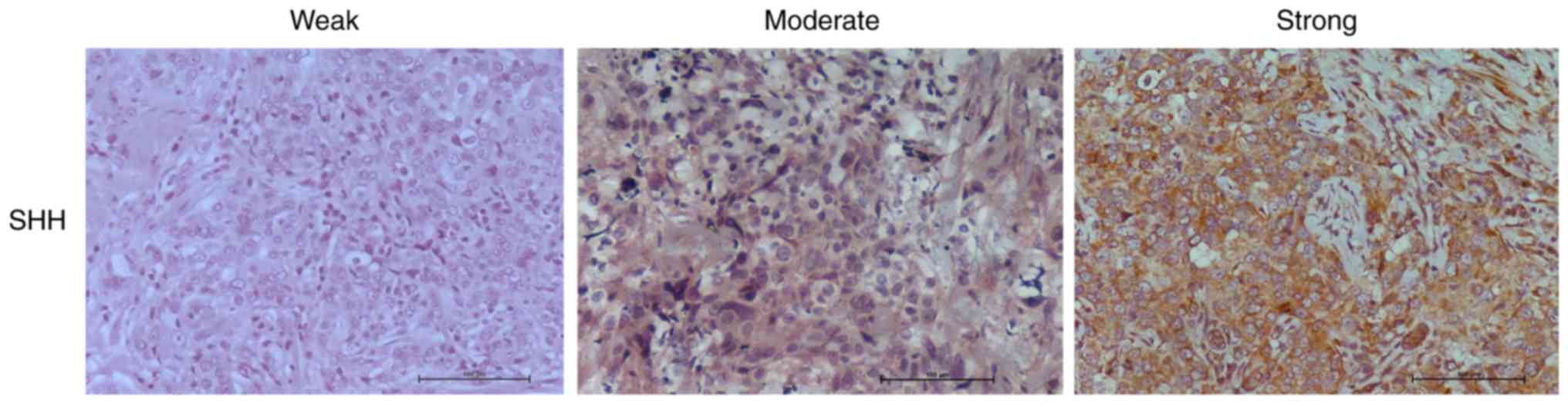
Gli-1 and β-catenin. For SHH and Gli-1, slides were scored as
having no expression (0), weak (1),
moderate (2), or strong (3) tumor cell staining (Fig. 1; Table
I). All samples showed cytoplasmic staining for SHH with 8
(22%) showing weak, 20 (56%) showing moderate, and 8 (22%) showing
strong staining (Fig. 1A; Table I). Expression of SHH was weakly
correlated to tumor stage (P=0.044) (Table I). Cytoplasmic Gli-1 expression was
observed in 35 (97%) of the 36 samples, with 9 samples (25%)
showing weak staining, 14 samples (39%) having moderate staining
and 12 samples (33%) having strong staining (Table I) (Fig.
1B). Nuclear Gli-1 expression was observed in 25 samples (70%).
Both cytoplasmic and nuclear Gli-1 expression correlated to tumor
stage (Table I).
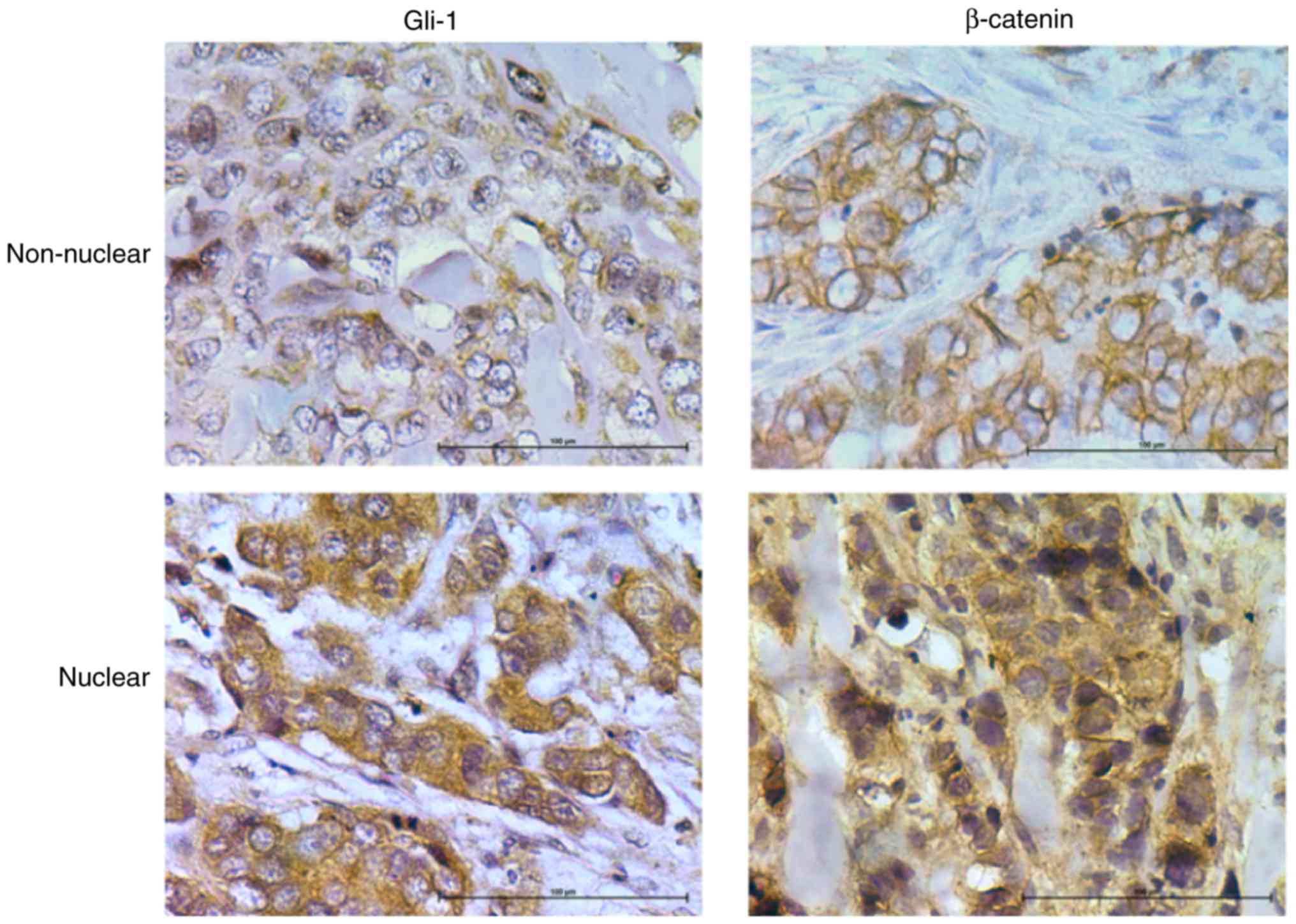
For β-catenin staining, slides were categorized as
having membranous staining or cytoplasmic/nuclear staining
(Fig. 2). Membranous β-catenin
staining was observed in 13 samples (36%). Nuclear/cytoplasmic
staining was observed in 64% of samples Cytoplasmic/nuclear
β-catenin was highly correlated to tumor stage (Table I). There was no significant
association with grade for any proteins examined (Table I). This is most likely due to the fact
that most tumors (n=31, 86%) were grade 3. This is consistent with
the fact that most TNBC are high grade at diagnosis.
Association between HH and Wnt
activation
SHH is significantly correlated to cytoplasmic
expression of Gli-1 (P<.001, Table
II). Interestingly, there was no association between nuclear
Gli-1 and cytoplasmic SHH expression (P=0.291). Expression of both
cytoplasmic and nuclear Gli-1 were correlated with tumor stage
(P<.001) and each other (P<.001). Likewise, there was an
association between cytoplasmic and nuclear β-catenin (P<.001)
and both correlated to tumor stage (P<.003, cytoplasmic,
P<.001 nuclear). SHH expression was significantly correlated to
cytoplasmic/nuclear β-catenin (P<.001). Likewise, a significant
correlation was observed between nuclear Gli-1 and nuclear
β-catenin (P<.002) (Table II).
This association remained when adjusting for tumor stage and
grade.
| Table II.Correlation of HH and WNT pathways in
TNBC. |
Table II.
Correlation of HH and WNT pathways in
TNBC.
| Variables |
| SHH | Gli-1 | Nuclear Gli | Nuclear Bcat |
|---|
| SHH | Correlation
coefficient | 1.000 | 0.598a | 0.181 | 0.520a |
|
| P-value |
| 0.000 | 0.291 | 0.001 |
| Gli-1 | Correlation
coefficient | 0.598a | 1.000 | 0.585a | 0.649a |
|
| P-value | 0.000 | | 0.000 | 0.000 |
| Nuclear Gli | Correlation
coefficient | 0.181 | 0.585a | 1.000 | 0.506a |
|
| P-value | 0.291 | 0.000 | | 0.002 |
|
Nuclear/cytoplasmic | Correlation
coefficient | 0.520a | 0.649a | 0.506a | 1.000 |
| β-catenin | P-value | 0.001 | 0.000 | 0.002 |
|
Co-activation of HH and Wnt predict
recurrence-free and overall survival
Independently, overexpression of SHH, Gli-1 and
β-catenin, as well as nuclear localization of Gli-1 and β-catenin
were associated with recurrence free and overall survival (Table I). To determine if dual activation of
HH and Wnt pathways lead to a worse prognosis, patients were
divided into three groups: those without activation of either
pathway, those with activation of only HH or Wnt, and those with
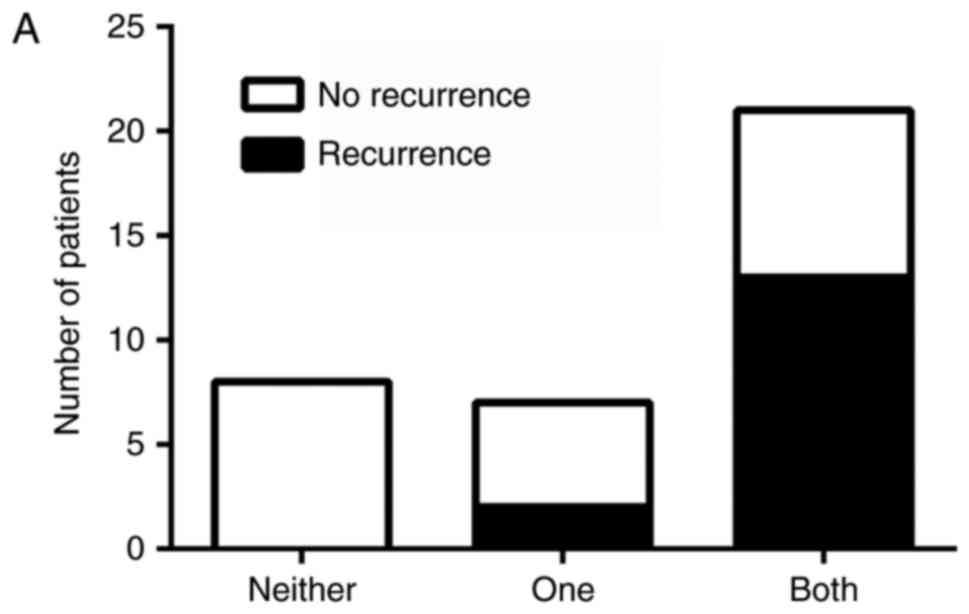
activation of both pathways. There was an association between
pathway activation and recurrence, χ2 (2)=11.11, P=.004, with those having
activation of both HH and Wnt pathways being more likely to be in
the later stage at diagnosis and at greater risk for recurrence
(Fig. 3A). Likewise, there was an
association between pathway activation and survival, χ2
(2)=6.75, P=.034, with longer
survival observed in patients who lacked activation of both
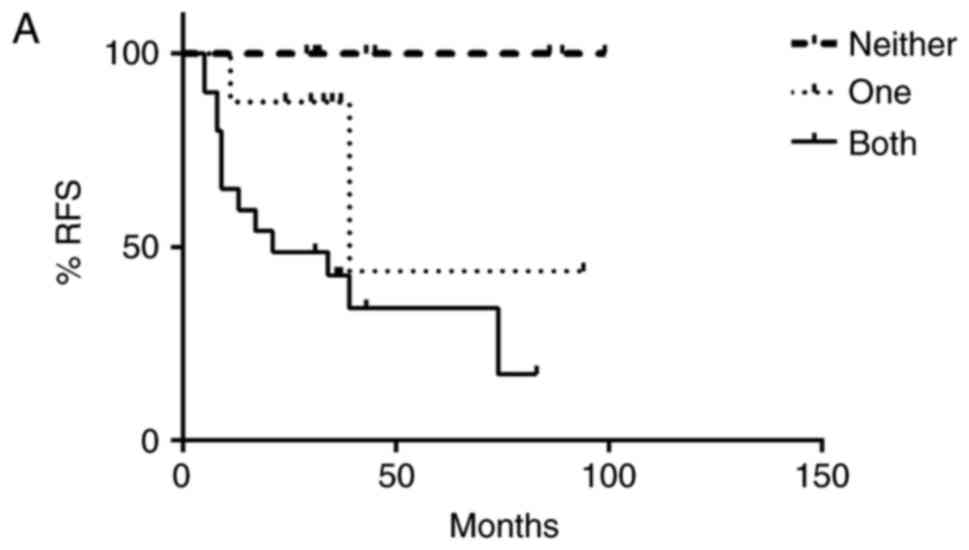
pathways (Fig. 3B). Progression free
and overall survival time of the patients with dual activation of
HH and Wnt pathways was significantly decreased (Fig. 4A and B). Patients who did not exhibit
activation of HH or Wnt pathways had a 100% survival rate and no
evidence of recurrence as of the end of this study. However,
patients with both HH and Wnt pathways activated had a greater
chance of early recurrence within the first three years compared to
those with only one pathway activated, with greater than half of
the patients exhibiting dual pathway activation relapsing. Patients
with only one pathway activated had a higher risk of later
recurrence than those without activation of either pathway, with
greater than half of these patients relapsing after three years.
Similarly, patients with both pathways activated had a lower
survival rate (50%) compared to patients with only one pathway
activated (87.5%) as of the end of this study (Fig. 4B).
Discussion
In this study we report activation of the HH and Wnt
pathways in TNBC. Expression of SHH, GLI-1, β-catenin were
independently associated to tumor stage. A significant correlation
was observed between nuclear Gli-1 and nuclear β-catenin and our
results indicate that co-activation of both HH and Wnt pathways is
associated with shorter recurrence-free and overall survival
times.
Our findings are consistent with previous studies
that indicate an association between HH signaling and clinical
outcome in breast cancer. Over expression of SMO and Gli-1 has been
observed in TNBC compared to both normal breast tissue, mammary
hyperplasia and ER+breast cancers (44). In this study, expression of Gli-1 was
significantly correlated to tumor stage and lymphatic involvement.
Alternatively, high expression of PTCH, and thereby inactivation of
HH signaling, is associated with favorable prognosis (22). Moreover, a recent report in HER2+
breast cancer indicated that nuclear activation of Gli-1 was
associated with an incomplete pathological response to neoadjuvant
chemotherapy, and poorer survival in both hormone receptor positive
and negative tumors (45). In
addition, in inflammatory breast cancer, high expression of SHH is
associated with poor prognosis (23).
Interestingly, although expression levels of SHH and Gli-1 were
associated with each other and tumor stage in our study, there was
no significant association between SHH and nuclear Gli-1 in tumor
cells. Although stromal staining was not included in our analysis,
weak to moderate staining of SHH was observed in the stroma of
several samples, and may contribute to paracrine activation of HH
signaling. Additionally, non-canonical activation may be
responsible for HH pathway activity in a subset of TNBC.
Non-canonical activation of Gli-1 has been previously described in
claudin-low subsets of breast cancer, and may be driven through a
NF-κB dependent pathway (46).
Activation of Wnt signaling has also been associated
to prognosis of TNBC (47).
Overexpression of WNT ligands was found to predict recurrence in
late stage TNBC (48). Wnt pathway
activity is also associated to increased metastasis in TNBC. Using
a classifier trained on β-catenin transfected mammary cells, in a
meta-analysis of 11 studies and 1,878 breast cancer patients found
Wnt activity associated primarily to TNBC compared to other
subtypes of breast cancer (49). In
addition, patients with high WNT activity were more likely to have
increased risk of brain and lung metastasis.
Our data show increased nuclear activity of Gli-1
and β-catenin correlates with increasing tumor stage and dual
activation of both is associated to shorter recurrence times and
overall survival, suggesting a role in invasion and disease
progression. Both pathways are involved in the regulation of genes
critical for drug resistance (50,51), EMT
(13,52) and differentiation (53,54).
Moreover, co-expression of HH and β-catenin was observed in poorly
differentiated breast cancers compared to low grade and benign
lesions (55). This expression
correlated with a mesenchymal-like phenotype defined by expression
of CD44 and vimentin, suggesting that simultaneous activation of HH
and Wnt pathways may promote enhanced EMT and de-differentiation of
breast cancer cells.
Although we observed significant expression of
nuclear Gli-1 and nuclear β-catenin in TNBC samples, it is unknown
if there is cross-talk between these pathways that results in
increased activation of one or both pathways. Direct interactions
of the HH and Wnt pathways have previously been reported.
Transcriptome analysis of WNT3a responsive TNBC cell lines revealed
Wnt target genes that are involved in the HH pathway signaling
(56). Additionally, exogenous
expression of constitutively nuclear β-catenin has been shown to
increase activity of a Gli-1 reporter, suggesting that active Wnt
signaling enhances Gli transcriptional activity (57). Likewise, inhibition of HH signaling
with the SMO inhibitor cyclopamine has been reported to decrease
β-catenin-TCF transcriptional activity in colon cancer lines
(58). Further research is needed to
determine the consequences of HH-Wnt interactions in breast cancer
in relation to drug resistance and metastasis.
Our study is the first to report that co-activation
of HH and Wnt pathways in clinical TNBC samples is associated to
shorter recurrence times and decreased survival when compared to
activation of only one of these pathways. Although this study was
performed in a limited cohort of patients, our findings suggest
that activation status of HH and Wnt pathways may provide a
prognostic biomarker for patients at risk for treatment resistance
and early recurrence and may be more valuable than independent
analysis of either pathway. However, our limited sample size
prevented multi-variant analysis to determine if co-activation of
HH and WNT pathways occurs independent of tumor stage. These
findings should be confirmed in subsequent studies with larger
patient numbers.
Acknowledgements
This project was supported by the Delaware INBRE
program, with a grant from the National Institute of General
Medical Sciences-NIGMS (P20 GM103446) from the National Institutes
of Health and the state of Delaware.
Glossary
Abbreviations
Abbreviations:
|
TNBC
|
triple negative breast cancer
|
|
SHH
|
sonic Hedgehog
|
|
HH
|
Hedgehog
|
References
|
1
|
Carey LA, Perou CM, Livasy CA, Dressler
LG, Cowan D, Conway K, Karaca G, Troester MA, Tse CK, Edmiston S,
et al: Race, breast cancer subtypes, and survival in the carolina
breast cancer study. JAMA. 295:2492–2502. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Foulkes WD, Stefansson IM, Chappuis PO,
Bégin LR, Goffin JR, Wong N, Trudel M and Akslen LA: Germline BRCA1
mutations and a basal epithelial phenotype in breast cancer. J Natl
Cancer Inst. 95:1482–1485. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Castiglioni F, Terenziani M, Carcangiu ML,
Miliano R, Aiello P, Bertola L, Triulzi T, Gasparini P, Camerini T,
Sozzi G, et al: Radiation effects on development of HER2-positive
breast carcinomas. Clin Cancer Res. 13:46–51. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dent R, Trudeau M, Pritchard KI, Hanna WM,
Kahn HK, Sawka CA, Lickley LA, Rawlinson E, Sun P and Narod SA:
Triple-negative breast cancer: Clinical features and patterns of
recurrence. Clin Cancer Res. 13:4429–4434. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hudis CA and Gianni L: Triple-negative
breast cancer: An unmet medical need. Oncologist. 16 Suppl
1:S1–S11. 2011. View Article : Google Scholar
|
|
6
|
Jia Y and Xie J: Promising molecular
mechanisms responsible for gemctiabine resistance in cancer. Genes
Dis. 2:299–306. 2015. View Article : Google Scholar
|
|
7
|
Angeloni V, Tiberio P, Appierto V and
Daidone MG: Implications of stemness-related signaling pathways in
breast cancer response to therapy. Semin Cancer Biol. 31:43–51.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Taipale J and Beachy PA: The hedgehog and
Wnt signalling pathways in cancer. Nature. 411:349–354. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Reya T and Clevers H: Wnt signalling in
stem cells and cancer. Nature. 434:843–850. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu S, Dontu G, Mantle ID, Patel S, Ahn
NS, Jackson KW, Suri P and Wicha MS: Hedgehog signaling and Bmi-1
regulate self-renewal of normal and malignant human mammary stem
cells. Cancer Res. 66:6063–6071. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Takebe N, Harris PJ, Warren RQ and Ivy SP:
Targeting cancer stem cells by inhibiting Wnt, notch, and hedgehog
pathways. Nat Rev Clin Oncol. 8:97–106. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Clevers H, Loh KM and Nusse R: Stem cell
signaling. An integral program for tissue renewal and regeneration:
Wnt signaling and stem cell control. Science. 346:12480122014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Takebe N, Warren RQ and Ivy SP: Breast
cancer growth and metastasis: Interplay between cancer stem cells,
embryonic signaling pathways and epithelial-to-mesenchymal
transition. Breast Cancer Res. 13:2112011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Flemban A and Qualtrough D: The potential
role of hedgehog signaling in the luminal/basal phenotype of breast
epithelia and in breast cancer invasion and metastasis. Cancers
(Basel). 7:1863–1884. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ingham PW and McMahon AP: Hedgehog
signaling in animal development: Paradigms and principles. Genes
Dev. 15:3059–3087. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lewis MT, Ross S, Strickland PA, Sugnet
CW, Jimenez E, Scott MP and Daniel CW: Defects in mouse mammary
gland development caused by conditional haploinsufficiency of
Patched-1. Development. 126:5181–5193. 1999.PubMed/NCBI
|
|
17
|
Kubo M, Nakamura M, Tasaki A, Yamanaka N,
Nakashima H, Nomura M, Kuroki S and Katano M: Hedgehog signaling
pathway is a new therapeutic target for patients with breast
cancer. Cancer Res. 64:6071–6074. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Moraes RC, Zhang X, Harrington N, Fung JY,
Wu MF, Hilsenbeck SG, Allred DC and Lewis MT: Constitutive
activation of smoothened (SMO) in mammary glands of transgenic mice
leads to increased proliferation, altered differentiation and
ductal dysplasia. Development. 134:1231–1242. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hui M, Cazet A, Nair R, Watkins DN,
O'Toole SA and Swarbrick A: The hedgehog signalling pathway in
breast development, carcinogenesis and cancer therapy. Breast
Cancer Res. 15:2032013. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Im S, Choi HJ, Yoo C, Jung JH, Jeon YW,
Suh YJ and Kang CS: Hedgehog related protein expression in breast
cancer: Gli-2 is associated with poor overall survival. Korean J
Pathol. 47:116–123. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Noman AS, Uddin M, Rahman MZ, Nayeem MJ,
Alam SS, Khatun Z, Wahiduzzaman M, Sultana A, Rahman ML, Ali MY, et
al: Overexpression of sonic hedgehog in the triple negative breast
cancer: Clinicopathological characteristics of high burden breast
cancer patients from Bangladesh. Sci Rep. 6:188302016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wolf I, Bose S, Desmond JC, Lin BT,
Williamson EA, Karlan BY and Koeffler HP: Unmasking of
epigenetically silenced genes reveals DNA promoter methylation and
reduced expression of PTCH in breast cancer. Breast Cancer Res
Treat. 105:139–155. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bièche I, Lerebours F, Tozlu S, Espie M,
Marty M and Lidereau R: Molecular profiling of inflammatory breast
cancer: Identification of a poor-prognosis gene expression
signature. Clin Cancer Res. 10:6789–6795. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Polakis P: Wnt signaling and cancer. Genes
Dev. 14:1837–1851. 2000.PubMed/NCBI
|
|
25
|
Nelson WJ and Nusse R: Convergence of Wnt,
beta-catenin, and cadherin pathways. Science. 303:1483–1487. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Duchartre Y, Kim YM and Kahn M: The Wnt
signaling pathway in cancer. Crit Rev Oncol Hematol. 99:141–149.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Katoh M and Katoh M: WNT signaling pathway
and stem cell signaling network. Clin Cancer Res. 13:4042–4045.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jamieson C, Sharma M and Henderson BR:
Targeting the β-catenin nuclear transport pathway in cancer. Semin
Cancer Biol. 27:20–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Clevers H: Wnt/beta-catenin signaling in
development and disease. Cell. 127:469–480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jönsson M, Borg A, Nilbert M and Andersson
T: Involvement of adenomatous polyposis coli (APC)/beta-catenin
signalling in human breast cancer. Eur J Cancer. 36:242–248. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lin SY, Xia W, Wang JC, Kwong KY, Spohn B,
Wen Y, Pestell RG and Hung MC: Beta-catenin, a novel prognostic
marker for breast cancer: Its roles in cyclin D1 expression and
cancer progression. Proc Natl Acad Sci USA. 97:4262–4266. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
King TD, Suto MJ and Li Y: The
Wnt/β-catenin signaling pathway: A potential therapeutic target in
the treatment of triple negative breast cancer. J Cell Biochem.
113:13–18. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hatsell S, Rowlands T, Hiremath M and
Cowin P: Beta-catenin and Tcfs in mammary development and cancer. J
Mammary Gland Biol Neoplasia. 8:145–158. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Huguet EL, McMahon JA, McMahon AP,
Bicknell R and Harris AL: Differential expression of human Wnt
genes 2, 3, 4, and 7B in human breast cell lines and normal and
disease states of human breast tissue. Cancer Res. 54:2615–2621.
1994.PubMed/NCBI
|
|
35
|
Benhaj K, Akcali KC and Ozturk M:
Redundant expression of canonical Wnt ligands in human breast
cancer cell lines. Oncol Rep. 15:701–707. 2006.PubMed/NCBI
|
|
36
|
Nagahata T, Shimada T, Harada A, Nagai H,
Onda M, Yokoyama S, Shiba T, Jin E, Kawanami O and Emi M:
Amplification, up-regulation and over-expression of DVL-1, the
human counterpart of the Drosophila disheveled gene, in primary
breast cancers. Cancer Sci. 94:515–518. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bànkfalvi A, Terpe HJ, Breukelmann D, Bier
B, Rempe D, Pschadka G, Krech R, Lellè RJ and Boecker W:
Immunophenotypic and prognostic analysis of E-cadherin and
beta-catenin expression during breast carcinogenesis and tumour
progression: A comparative study with CD44. Histopathology.
34:25–34. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Karayiannakis AJ, Nakopoulou L,
Gakiopoulou H, Keramopoulos A, Davaris PS and Pignatelli M:
Expression patterns of beta-catenin in in situ and invasive breast
cancer. Eur J Surg Oncol. 27:31–36. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wong SC, Lo SF, Lee KC, Yam JW, Chan JK
and Hsiao Wendy WL: Expression of frizzled-related protein and
Wnt-signalling molecules in invasive human breast tumours. J
Pathol. 196:145–153. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yang SH, Andl T, Grachtchouk V, Wang A,
Liu J, Syu LJ, Ferris J, Wang TS, Glick AB, Millar SE and Dlugosz
AA: Pathological responses to oncogenic hedgehog signaling in skin
are dependent on canonical Wnt/beta3-catenin signaling. Nat Genet.
40:1130–1135. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
41
|
He J, Sheng T, Stelter AA, Li C, Zhang X,
Sinha M, Luxon BA and Xie J: Suppressing Wnt signaling by the
hedgehog pathway through sFRP-1. J Biol Chem. 281:35598–35602.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Schneider FT, Schänzer A, Czupalla CJ,
Thom S, Engels K, Schmidt MH, Plate KH and Liebner S: Sonic
hedgehog acts as a negative regulator of {beta}-catenin signaling
in the adult tongue epithelium. Am J Pathol. 177:404–414. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Opdenaker LM, Arnold KM, Pohlig RT,
Padmanabhan JS, Flynn DC and Sims-Mourtada J: Immunohistochemical
analysis of aldehyde dehydrogenase isoforms and their association
with estrogen-receptor status and disease progression in breast
cancer. Breast Cancer (Dove Med Press). 6:205–209. 2014.PubMed/NCBI
|
|
44
|
Tao Y, Mao J, Zhang Q and Li L:
Overexpression of hedgehog signaling molecules and its involvement
in triple-negative breast cancer. Oncol Lett. 2:995–1001.
2011.PubMed/NCBI
|
|
45
|
Liu S, Duan X, Xu L, Ye J, Cheng Y, Liu Q,
Zhang H, Zhang S, Zhu S, Li T and Liu Y: Nuclear Gli1 expression is
associated with pathological complete response and event-free
survival in HER2-positive breast cancer treated with
trastuzumab-based neoadjuvant therapy. Tumour Biol. 37:4873–4881.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Colavito SA, Zou MR, Yan Q, Nguyen DX and
Stern DF: Significance of glioma-associated oncogene homolog 1
(GLI1) expression in claudin-low breast cancer and crosstalk with
the nuclear factor kappa-light-chain-enhancer of activated B cells
(NFκB) pathway. Breast Cancer Res. 16:4442014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xu WH, Liu ZB, Yang C, Qin W and Shao ZM:
Expression of dickkopf-1 and beta-catenin related to the prognosis
of breast cancer patients with triple negative phenotype. PLoS One.
7:e376242012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Tsai CH, Chiu JH, Yang CW, Wang JY, Tsai
YF, Tseng LM, Chen WS and Shyr YM: Molecular characteristics of
recurrent triple-negative breast cancer. Mol Med Rep. 12:7326–7334.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Dey N, Barwick BG, Moreno CS,
Ordanic-Kodani M, Chen Z, Oprea-Ilies G, Tang W, Catzavelos C,
Kerstann KF, Sledge GW Jr, et al: Wnt signaling in triple negative
breast cancer is associated with metastasis. BMC Cancer.
13:5372013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Sims-Mourtada J, Izzo JG, Ajani J and Chao
KS: Sonic hedgehog promotes multiple drug resistance by regulation
of drug transport. Oncogene. 26:5674–5679. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Xia Z, Guo M, Liu H, Jiang L, Li Q, Peng
J, Li JD, Shan B, Feng P and Ma H: CBP-dependent Wnt/β-catenin
signaling is crucial in regulation of MDR1 transcription. Curr
Cancer Drug Targets. 15:519–532. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Micalizzi DS, Farabaugh SM and Ford HL:
Epithelial-mesenchymal transition in cancer: Parallels between
normal development and tumor progression. J Mammary Gland Biol
Neoplasia. 15:117–134. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Alexander CM, Goel S, Fakhraldeen SA and
Kim S: Wnt signaling in mammary glands: Plastic cell fates and
combinatorial signaling. Cold Spring Harb Perspect Biol. 4:pii:
a008037. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kakarala M and Wicha MS: Implications of
the cancer stem-cell hypothesis for breast cancer prevention and
therapy. J Clin Oncol. 26:2813–2820. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Scimeca M, Antonacci C, Colombo D,
Bonfiglio R, Buonomo OC and Bonanno E: Emerging prognostic markers
related to mesenchymal characteristics of poorly differentiated
breast cancers. Tumour Biol. 37:5427–5435. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Maubant S, Tesson B, Maire V, Ye M,
Rigaill G, Gentien D, Cruzalegui F, Tucker GC, Roman-Roman S and
Dubois T: Transcriptome analysis of Wnt3a-treated triple-negative
breast cancer cells. PLoS One. 10:e01223332015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Maeda O, Kondo M, Fujita T, Usami N, Fukui
T, Shimokata K, Ando T, Goto H and Sekido Y: Enhancement of
GLI1-transcriptional activity by beta-catenin in human cancer
cells. Oncol Rep. 16:91–96. 2006.PubMed/NCBI
|
|
58
|
Qualtrough D, Rees P, Speight B, Williams
AC and Paraskeva C: The hedgehog inhibitor cyclopamine reduces
β-catenin-Tcf transcriptional activity, induces E-cadherin
expression, and reduces invasion in colorectal cancer cells.
Cancers (Basel). 7:1885–1899. 2015. View Article : Google Scholar : PubMed/NCBI
|