Introduction
Dendritic cells (DCs) are antigen-presenting cells
(APCs) that serve a critical function in maintaining tolerance to
self-antigens and in the induction and regulation of immune
responses due to their intrinsic immune-priming abilities. These
properties make the development of DC-based vaccines for cancer
immunotherapy an attractive prospect (1,2). However,
to date, the clinical outcomes of studies investigating DC-based
vaccination for the treatment of cancer have not been promising.
One of the limitations of this approach is the low expression level
of antigenic epitopes on cancer cells, making them poorly
immunogenic to DCs (3–6). In cancer immunotherapy, it is essential
to sufficiently expose tumor-associated antigens (TAAs) to DCs in
order to stimulate DC maturation and facilitate the subsequent
activation of antigen-specific immune responses. Our previous data
revealed that calreticulin (CRT) is a key molecule involved in cell
recognition and that it may act as an ‘immunological adjuvant’ by
translocating type I transmembrane glycoprotein mucin 1 (MUC1), a
breast cancer antigen, to the surface of 4T1 cells (7). The MUC1-CRT-primed 4T1 cells were
recognized by DCs, leading to a specific antitumor immune
response.
Therefore, efforts to develop tumor vaccines have
focused on promoting the maturation of DCs as a means of enhancing
antitumor immunity (8–10). However, the induction of immunity
against cancer cells is also restricted by intrinsic inhibitory
mechanisms; for example, suppressor of cytokine signaling 1 (SOCS1)
functions as a negative regulator of Janus kinases (JAKs), thereby
acting as an inhibitory regulator of antigen presentation by DCs
and the magnitude of adaptive immune responses (11). The present study specifically focused
on the incubation of SOCS1-silenced DCs with MUC1-CRT-primed 4T1
cells in an attempt to enhance specific immunological effects in
vitro and in vivo. RNA interference technology, which is
widely used to silence endogenous molecules involved in DC-mediated
apoptosis and immunosuppressive signaling, was utilized to improve
responses against cancer cells (12,13). In
the present study, small interfering RNAs (siRNAs) were effectively
introduced into DCs with a simple and controllable antigen delivery
system: A novel, human-derived, positively-charged cell-penetrating
peptide, denoted hpp10 (14),
accompanied and shielded the negatively-charged siRNA via a
double-stranded RNA-binding domain (DRBD) (15).
The present study has implications for the
development of more effective DC-based breast cancer vaccines by
combining high TAA expression levels on breast cancer cells with
silencing of the critical inhibitor of antigen presentation on DCs.
Therefore, the results of the present study may lead to an improved
antitumor immunotherapeutic approach for the treatment of breast
cancer.
Materials and methods
Experimental animals
Approval from the Medical Animal Care and Welfare
Committee of China Three Gorges University (Hubei, China) was
obtained prior to any experimentation using animals. A total of 97
female BALB/c mice (18±2 g, 4–6 weeks old) were purchased from the
Laboratory Animal Center of China Three Gorges University. All mice
were housed in specific pathogen-free conditions, with free access
to food and water. The ambient temperature was maintained at 22±2°C
with a humidity of 50–60% and a 12 h light/dark cycle. All
experiments in mice were performed following the relevant
institutional guidelines and regulations, and were subject to a
protocol approved by the China Three Gorges University Animal Care
Committee.
Drugs and chemicals
Lipofectamine 2000 was purchased from Thermo Fisher
Scientific, Inc. (Waltham, MA, USA). Lipopolysaccharide (LPS) and
anti-signal transducer and activator of transcription 1 (STAT1)
antibodies were purchased from Sigma-Aldrich; Merck KGaA
(Darmstadt, Germany; cat. nos. L2630 and SAB4300133, respectively);
the Lactate Dehydrogenase (LDH) Activity Assay kit was also
purchased from Sigma-Aldrich; Merck KGaA. Mitoxantrone (MIT) was
purchased from Jiangsu Aosaikang Pharmaceutical Co., Ltd. (Nanjing,
China). Magnetic beads and anti-SOCS1 antibodies (cat. no. 38-5200)
were purchased from eBioscience; Thermo Fisher Scientific, Inc.
(Waltham, MA, USA). Mouse granulocyte-macrophage colony-stimulating
factor (mGM-CSF) and mouse interleukin-4 (mIL-4) primary antibodies
were purchased from PeproTech, Inc. (Rocky Hill, NJ, USA; cat. nos.
315-03 and 500-p45, respectively), which were used for DC cell
culture as described previously (7).
Anti-cluster of differentiation (CD)11c-allophycocyanin,
anti-CD83-fluorescein isothiocyanate and anti-phosphorylated
(p)-STAT1 antibodies were purchased from Biolegend, Inc. (San
Diego, CA, USA; cat. No. 117305, cat. nos. 121505 and 686402,
respectively). Purified anti-β-actin antibody was purchased from
Biolegend, Inc. (San Diego, CA, USA; cat. no. 643801). Horseradish
peroxidase (HRP)-labeled goat anti-mouse immunoglobulin G (IgG)
[heavy and light (H+L)] and HRP-labeled goat anti-rabbit IgG (H+L)
were purchased from Beyotime Co. (Shanghai, China; cat. nos. A0216
and A0208, respectively). Interferon (IFN)-γ and tumor necrosis
factor (TNF)-α ELISA kits were purchased from Boster Biological
Technology (Pleasanton, CA, USA; cat. nos. EK0375 and EK0527,
respectively). Lymphocyte separation medium was purchased from
Dakewe. Co. (Shenzhen, China; cat. no. DKW33-R0100). RPMI-1640
medium, L-glutamine, streptomycin and penicillin were all purchased
from Biosera, Inc. (Nuaille, France). Bovine serum albumin and
fetal bovine serum (FBS) and beta-actin were purchased from Gibco;
Thermo Fisher Scientific, Inc. All primers were synthesized by
Sangon Biotech Co., Ltd. (Shanghai, China).
Cell line culture
Bone marrow-derived dendritic cells (BMDCs) were
collected and identified as previously described (7). The DC2.4 cell line was purchased from
Suershengwu Co., Ltd. (Shanghai, China), the breast cancer 4T1 cell
line, originating from BALB/c mice, was purchased from American
Type Culture Collection (Manassas, VA, USA) and all cells were
maintained in the laboratory in the China Three Gorges University.
Cells were cultured (2.0×105/ml) in complete medium
(RPMI-1640 medium supplemented with 10 mmol/l L-glutamine, 10%
heat-inactivated FBS, 100 U/ml penicillin and 100 µg/ml
streptomycin) in a humidified incubator at 37°C containing 5%
CO2.
SOCS1 siRNA selection
Three siRNAs targeting SOCS1 (and a negative
control) were synthesized by Shanghai GenePharma Co., Ltd.
(Shanghai, China), the sequences of which are listed in Table I. The RNAs were de-protected and
annealed according to the manufacturer's protocol. DC2.4 cells were
seeded onto 6-well culture plates (1.0×106/well) and
were cultivated to semi-confluence in RPMI-1640 medium for 24 h at
37°C. Cells were further incubated with 1.0 µg SOCS1 siRNA and 3.0
µl Lipofectamine 2000 reagent in 100 µl serum-free medium for 4–6 h
at 37°C. Subsequently, the cells were lysed in whole-cell
extraction buffer (50 mM Tris/HCl, pH 7.4, 250 mM NaCl, 5 mM EDTA,
50 mM NaF, 1 mM Na3VO4, 1% Nonidet P40, 0.02%
NaN3 and 1 mM phenylmethylsulfonyl fluoride). Following
centrifugation at 10,000 × g for 20 min at 4°C, the supernatant was
collected and the protein concentration was quantified by a BCA
assay. Total protein (40 µg per lane) was used for western blot
analyses and was probed with primary antibodies specific to SOCS1,
pSTAT1 and STAT1 according to a protocol described previously
(16).
 | Table I.SOCS1 siRNAs and hpp10-DRBD
sequences. |
Table I.
SOCS1 siRNAs and hpp10-DRBD
sequences.
| Name | Sequences
(5′-3′) |
|---|
| Scramble control |
FAM-UUCUCCGAAGGUGUCACGU |
| SOCS1 siRNA1 |
FAM-AGACCUUCGACUGCCUUUUTT |
| SOCS1 siRNA2 |
FAM-CUACCUGAGUUCCUUCCCCTT |
| SOCS1 siRNA3 |
FAM-ACACUCACUUCCGCACCUUTT |
| Hpp10-DRBD |
AAAATCCCCCTGCCCCGCTTCAAACTGAAATGTATCTTCTGTAAGAAGCGGAGGAAAAGACTCGAGGGCGATCCGGCTGGTGATCTTTCAGCAGGTTTCTTCATGGAGGAACTT |
|
|
AATACATACCGTCAGAAGCAGGGAGTAGTACTTAAATATCAAGAACTGCCTAATTCAGGACCTCCACATGATAGGAGGTTTACATTTCAAGTTATAATAGATGGAAGAGAATTTCCAGAAGGTGAAGGTAGATCAAAGAAGGAAGCAAAAATGCCGCAGCCAAATTAGCTGTTGAGATACTTAATAAGGAAAAGAAGGCAGGATCC |
To measure cytokine production, splenic lymphocytes
were separated and collected in a lymphocyte separation medium, as
described previously (17), and
successful purification was verified by light field microscopy
(magnification, ×40). DC2.4 cells were incubated with 1.0 µg SOCS1
siRNAs in 100 µl serum-free medium for 1 h at 37°C. To stimulate
DCs, 2×105 cells/ml were cultured in the presence of 100
ng/ml LPS for 24 h and were further incubated with lymphocytes at
37°C in 5% CO2 for 72 h. Following stimulation, IFN-γ
levels in the culture supernatants (100 µl/well) were analyzed
using an ELISA kit in accordance with the manufacturer's
protocol.
Cellular uptake of hpp10-DRBD-SOCS1
siRNA
To efficiently deliver SOCS1 siRNA into BMDCs,
hpp10-DRBD was expressed and purified. Briefly, hpp10-linker-DRBD
was synthesized by Shanghai Shenggong Biology Engineering
Technology Service, Ltd. (Shanghai, China), and was linked to
compatible sites (XhoI/BamHI) of a pET15b vector
(Preclinical Medicine Key Laboratory of China Three Gorges
University) to yield recombinant protein hpp10-DRBD (Table I). hpp10-DRBD was expressed using
isopropyl β-D-1-thiogalactopyranoside induction and was purified
under native conditions in its soluble form, as described
previously (18).
The internalization of hpp10-DRBD-SOCS1 siRNA into
BMDCs was observed using fluorescence microscopy (magnification,
×40). Cells were plated onto6-well-plates (1.0×106/well)
and were cultured to semi-confluence in RPMI-1640 medium for 24 h
at 37°C. Cells were washed three times with PBS, followed by
incubation with 10.0 µg hpp10-DRBD fusion protein and 1.0 µg
selected SOCS1 siRNA in serum-free medium for 1 h at 37°C. The
medium was removed and the cells were washed three times with PBS,
prior to being imaged using a fluorescence microscope
(magnification, ×40). Western blot analysis was performed to detect
SOCS1 expression levels following treatment with hpp10-DRBD-SOCS1
siRNA using an anti-SOCS1 antibody, as aforementioned.
DC preparation
BMDCs were seeded onto 6-well culture plates
(1.0×106/well) in complete medium (RPMI-1640 medium
supplemented with 10 mmol/l L-glutamine, 10% heat-inactivated FBS,
100 U/ml penicillin and 100 µg/ml streptomycin) in a humidified
incubator at 37°C containing 5% CO2 and were divided
into groups A, B, C and D, according to the following treatments.
For group A, BMDCs were incubated with 10.0 µg hpp10-DRBD and 1.0
µg SOCS1 siRNA in serum-free medium for 1 h at 37°C. For group B,
pEGFP-type I transmembrane glycoprotein mucin 1 (MUC1)-CRT was
constructed as previously described (7) and the primer pairs are presented in
Table II. The pEGFP-MUC1-CRT plasmid
(1.0 µg) was transfected into 4T1 cells to construct
MUC1-CRT-primed 4T1 cells by adding 3.0 µl Lipofectamine 2000 for
4–6 h at 37°C, and the efficiency was observed using fluorescence
microscopy (magnification, ×40). Subsequently, 1.0×106
MUC1-CRT-primed 4T1 cells were treated with 8 µg/ml MIT for 12 h to
induce CRT and MUC1 translocation to the surface of pre-apoptotic
4T1 cells, prior to being further incubated
(1.0×106/well at 37°C) with BMDCs. Cells in group C were
double-treated; BMDCs were incubated with hpp10-DRBD-SOCS1 siRNA
followed by incubation with MUC1-CRT-primed 4T1 cells (as described
for groups A and B). Finally, group D BMDCs were untreated. All
cells were washed three times with PBS and collected.
| Table II.Primer pairs. |
Table II.
Primer pairs.
| Primer name | Sequence (5′-3′) | Restriction
enzyme |
|---|
| CRT_F | GGTTCTGTCGACGACCCTGCCATCTATTTC | SalI |
| CRT_R | TACGGATCCCTACAGCTCATCCTTGGC | BamHI |
| MUC1_F | AATAGTCGACCCGGACACCAGGCCGGCCCC | SalI |
| MUC1_R | ATATGGATCCGGCCGAGGTGACACCATGGG | BamHI |
| MUC1-CRT_F | GGTCGTCGACAGAACCGCCGGCCGAGG | SalI |
| MUC1-CRT_R | GGTTCTGGATCCGACCCTGCCATCTATTTC | BamHI |
For the LDH leakage assay and animal experiments,
BMDCs from each of the 4 groups were collected separately using
magnetic beads according to the manufacturer's protocol. The beads
provided a strong magnetic attraction for efficient separation in
standard microcentrifuge tubes, and were adjusted to a
concentration of 1.0×107/ml for further use.
In vitro functional assays
Cytokine production assays
BMDCs were seeded onto 6-well culture plates
(1.0×106/well), and splenocytes were harvested and
prepared as single cell suspensions (7). Pre-treated BMDCs (as described under the
‘DC preparation’ subheading) were incubated with splenocytes at
37°C for 72 h. Following centrifugation at 800 × g for 5 min at
room temperature, the supernatant was collected and culture
supernatants (100 µl/well) were used to analyze production of the
cytokines IFN-γ and TNF-α by ELISA, according to the manufacturer's
protocol.
T cell proliferation assay
The MTT assay was used to evaluate T cell
proliferation. BMDCs and splenocytes were prepared as
aforementioned. Pre-treated BMDCs (5×105 cells/well) (as
described under the ‘DC preparation’ subheading) were incubated
with splenocytes (5×105 cells/well) at 37°C for 24, 48
or 72 h, and were washed three times with PBS. Cells were incubated
in fresh medium containing MTT solution (5 mg/ml with PBS) for 4 h
at 37°C and absorbance was measured at 540/570 nm using a Multiskan
Spectrum plate reader (Thermo Fisher Scientific, Inc.).
LDH leakage assays
Membrane integrity was assessed by LDH leakage into
the culture medium. Pre-treated BMDCs (5×105 cells/well)
were incubated with splenocytes (5×105 cells/well) at
37°C for 48 h, and the splenocytes were separated by magnetic
beads, according to the manufacturer's protocol. 4T1 cells were
harvested and single-cell suspensions were prepared as previously
described (7) and subsequently
incubated with separated splenocytes at 37°C for 16 h. The specific
killing activity of cytotoxic T lymphocytes induced by DC was
evaluated using an LDH assay. Cell-free supernatant was obtained
and transferred onto 96-well plates, and the LDH substrate reaction
buffer, taken from the LDH Activity Assay kit, was added to each
well for a 10-min enzymatic reaction according to the
manufacturer's protocol Absorbance was measured at 560/590 nm using
a Multiskan Spectrum plate reader.
In vivo functional assays
Immunization with DCs
A total of 50 mice were randomly divided into 5
groups (A, B, C, D and E). To generate a breast cancer-bearing
mouse model, the BALB/c mouse breast cancer 4T1 cell line was
sub-cultured twice and intradermally injected into the dorsal skin
of BALB/c mice (2.0×105 cells per injection). After 5
days of growth, a palpable tumor had formed. The mice were then
intradermally injected with one of the following: 100 µl normal
saline (group A), 100 µl 1.0×107/ml untreated DCs (group
B), 100 µl 1.0×107/ml SOCS1 siRNA-treated DCs (group C),
100 µl 1.0×107/ml MUC1-CRT-primed 4T1-treated DCs (group
D) or 100 µl 1.0×107/ml SOCS1 siRNA-treated and
MUC1-CRT-primed 4T1-treated (double-treated) DCs (group E). One day
post-injection, 50 µg TNF-α was intraperitoneally administered to
each mouse. Tumor size (mm3) was assessed every other
day. After 16 days, mice were sacrificed, tumors were isolated and
the size of the tumors was measured.
To assess the production of cytokines from BALB/c
mice bearing 4T1 tumors, tumors were collected and depleted of red
blood cells, and IFN-γ and TNF-α production was detected as
aforementioned.
Statistical analysis
Comparisons between two groups were assessed with a
Mann-Whitney U test. Comparisons among multiple groups were
performed using one-way analysis of variance followed by Tukey's
post hoc analysis. Kaplan-Meier survival curves were analyzed using
log-rank tests. GraphPad Prism software (version 5.0; GraphPad
Software, Inc., La Jolla, CA, USA) was used to create graphs and to
perform statistical analysis. P<0.05 was considered to indicate
a statistically significant difference.
Results
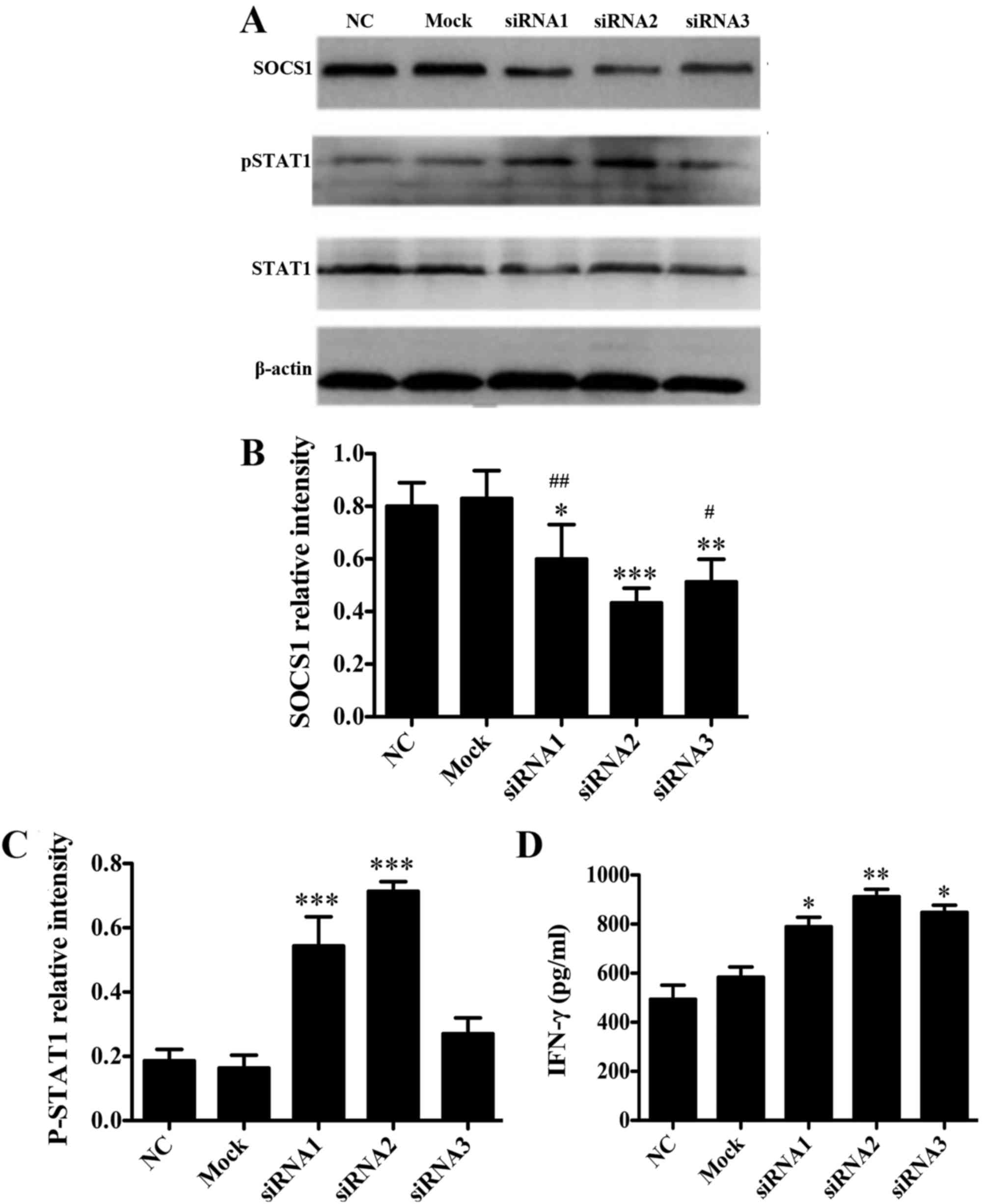
SOCS1 negatively regulates DCs in
vitro
To investigate whether SOCS1 negatively regulates
DCs in vitro, three synthetic siRNA oligo duplexes were
efficiently transfected into the DC2.4 cell line using
Lipofectamine 2000. As verified by western blot analysis, when
compared with NC group the protein expression levels of SOCS1 in
DC2.4 cells transfected with SOCS1 siRNA2 were significantly
reduced (P<0.001 vs. NC; Fig. 1A and
B), which were significantly lower than those in the cells
transfected with SOCS1 siRNA1 and siRNA3, respectively (P<0.05
vs. SOCS1 siRNA2; Fig. 1A and B). In
addition, when compared with NC group the cells transfected with
SOCS1 siRNA2 pSTAT1 expression levels were significantly increased
(P<0.001 vs. NC; Fig. 1A and C),
and the secretion of cytokine IFN-γ following LPS stimulation was
significantly increased (P<0.01 vs. NC; Fig. 1D). These data suggested that SOCS1
siRNA2 negatively regulated the JAK/STAT pathway and activated
pro-inflammatory cytokine production. Therefore, SOCS1 siRNA2 was
used for subsequent experiments.
Cellular uptake of hpp10-DRBD-SOCS1
siRNA
To validate the penetration of BMDCs and the
delivery of SOCS1 siRNA by the recombinant protein hpp10-DRBD
(Fig. 2A), a cellular uptake
experiment was performed. Fluorescence microscopy was used to
demonstrate transfection efficiency and revealed that >95% of
the DCs were transfected successfully with SOCS1 siRNA (data not
shown). Cells were subsequently collected and lysed for western
blot analysis. The results demonstrated that hpp10-DRBD
successfully delivered siRNA into BMDCs, and that this silenced the
expression of SOCS1 (Fig. 2B).
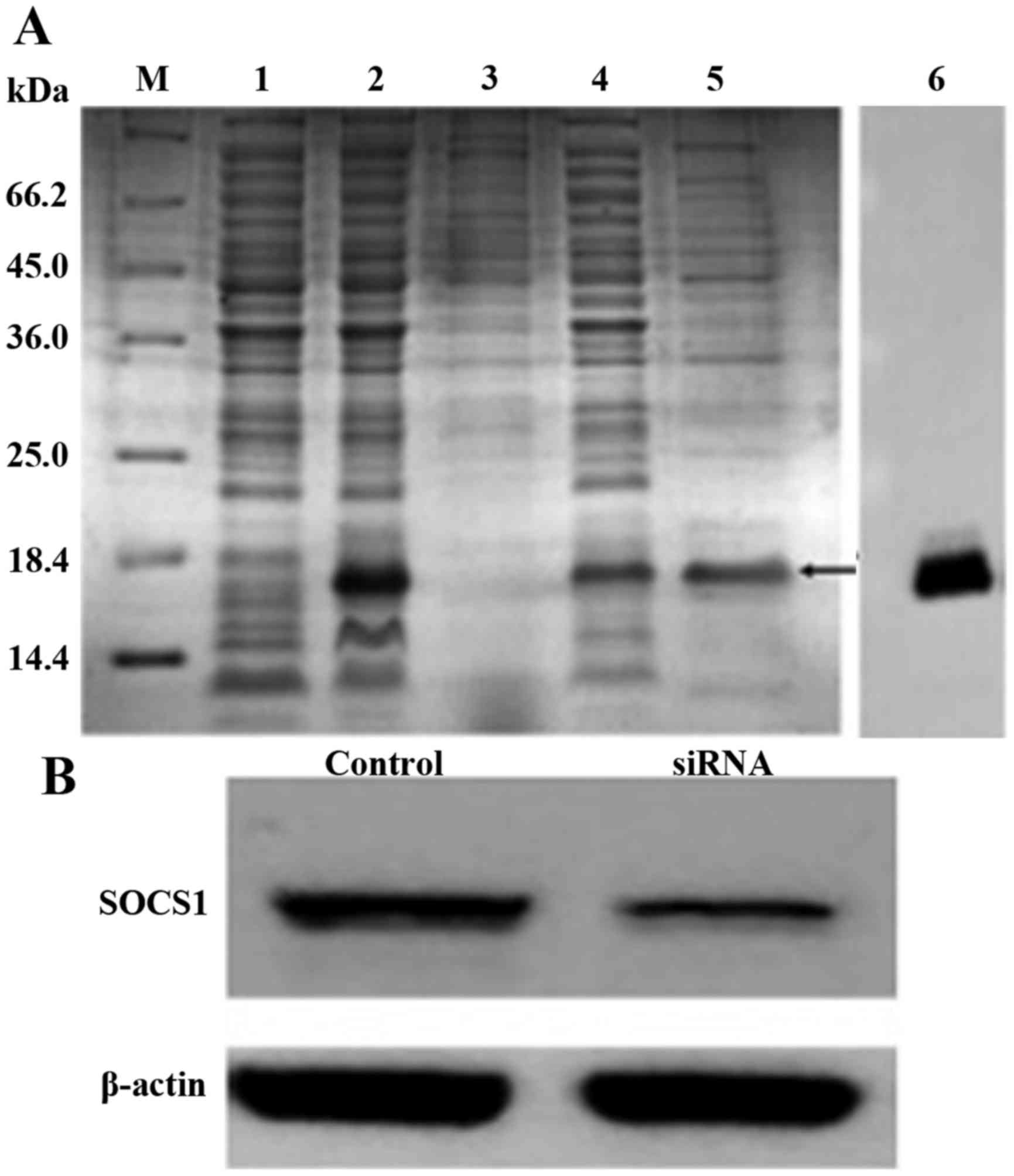 | Figure 2.Hpp10-DRBD construction and SOCS1
siRNA delivery. (A) SDS-PAGE and Coomassie Blue staining of
purified recombinant hpp10-DRBD. M, protein marker 431; 1,
non-induced protein; 2, induced protein; 3, precipitate; 4,
supernatant; 5, eluted protein; and 6, western blotting to detect
the purified protein. (B) Western blot analysis was performed to
detect SOCS1 expression levels in BMDCs following transfection with
hpp10-DRBD-SOCS1 siRNA for 48 h. DRBD, double-stranded RNA
binding-domain; SOCS1, suppressor of cytokine signaling 1; siRNA,
small interfering RNA; BMDC, bone marrow-derived dendritic
cell. |
Silencing of SOCS1 enhances the
immunological effect of MUC1-CRT-primed 4T1-treated DCs in
vitro
To validate whether SOCS1 siRNA enhances cytokine
production in MUC1-CRT-primed 4T1-treated BMDCs, BMDCs were
incubated with SOCS1 siRNA followed by MUC1-CRT-primed 4T1 cells
prior to being incubated with splenocytes. BMDCs were transfected
successfully with MUC1-CRT (data not shown). Compared with NC
group, the double-treated BMDCs exhibited significantly increased
production of IFN-γ and TNF-α (P<0.001 vs. NC; Fig. 3A and B), and production of these
molecules was also significantly increased compared with
MUC1-CRT-primed 4T1-treated BMDCs (P<0.05 vs. SOCS1 siRNA2;
Fig. 3A and B). These results
demonstrated that SOCS1-silencing, combined with MUC1-CRT-primed
4T1 cell treatment, may induce increased cytokine production by
DCs.
 | Figure 3.Silencing of SOCS1 enhances the
immunological effects of MUC1-CRT-primed 4T1-treated DCs in
vitro. Pre-treated BMDCs were incubated with splenocytes, and
culture supernatants (100 µl/well) were used to analyze production
of the cytokines, (A) TNF-α and (B) IFN-γ, by ELISA assay. (C)
Pre-treated BMDCs were incubated with splenocytes at 37°C for 24,
48 or 72 h, and an MTT assay was used to evaluate T cell
proliferation. (D) Pre-treated BMDCs were incubated with
splenocytes at 37°C for 48 h and splenocytes were separated by
magnetic beads, followed by incubation with 4T1 cells at 37°C for
30 min. The specific killing activity of cytotoxic T lymphocytes
induced by DC was evaluated using an LDH assay. *P<0.05,
**P<0.01 and ***P<0.001 vs. NC; #P<0.05.
Cell-free supernatant was used to detect membrane integrity by
lactate dehydrogenase assay. SOCS1, suppressor of cytokine
signaling 1; MUC1, type I transmembrane glycoprotein mucin 1; CRT,
calreticulin; DCs, dendritic cells; BMDCs, bone marrow-derived DCs;
TNF-α, tumor necrosis factor-α; IFN-γ, interferon-γ; LDH, lactate
dehydrogenase. |
The induction of T cell proliferation by BMDCs is an
important antitumor function. As presented in Fig. 3C, compared with NC group the
double-treatment of BMDCs significantly induced T cell production
(P<0.001 vs. NC) and this induction was significantly increased
compared with the MUC1-CRT-primed 4T1-treated BMDCs (P<0.05;
Fig. 3C), suggesting that
SOCS1-silencing, followed by MUC1-CRT-primed 4T1 treatment of DCs,
increased T cell proliferation.
Subsequently, the cytotoxicity of pre-treated BMDCs
on the breast cancer 4T1 cell line was evaluated via an LDH release
assay. As presented in Fig. 3D,
compared with NC group the double-treatment of BMDCs significantly
induced LDH release (P<0.001 vs. NC), and the cytotoxic effect
of the double treatment was increased compared with MUC1-CRT-primed
4T1-treated BMDCs (P<0.05 vs. SOCS1 siRNA2; Fig. 3D). These results suggested that
SOCS1-silencing, followed by MUC1-CRT-primed 4T1 treatment of DCs,
is associated with a higher level of membrane disturbance,
indicating cytotoxicity. Collectively, the experiments indicated
that silencing of SOCS1 in MUC1-CRT-primed 4T1-treated BMDCs
induced immunological effects in vitro.
Silencing of SOCS1 enhances the
immunological effects of MUC1-CRT-primed 4T1-treated DCs in
vivo
The in vitro experimental data demonstrated
that SOCS1-silencing combined with MUC1-CRT-primed 4T1 treatment of
BMDCs may serve as a novel therapy for targeting cancer cells. To
assess this in vivo, a breast cancer-bearing BALB/c mouse
model was generated and subjected to pre-treated BMDCs that were
developed in different ways, as before mentioned (groups A-E).
Growth curves of tumors were produced from daily caliper
measurements (Fig. 4A and B). The
mice were sacrificed on the final day and the volume and weight of
the excised tumors were measured (Fig.
4C-E). Compared with NC group the double-treated BMDCs
significantly inhibited the growth of 4T1 cells (P<0.01 vs. NC;
Fig. 4D and E) and the double-treated
BMDCs had a greater inhibitory effect compared
withtheMUC1-CRT-primed 4T1-treated BMDCs (P<0.05 vs. SOCS1
siRNA2; Fig. 4D and E).
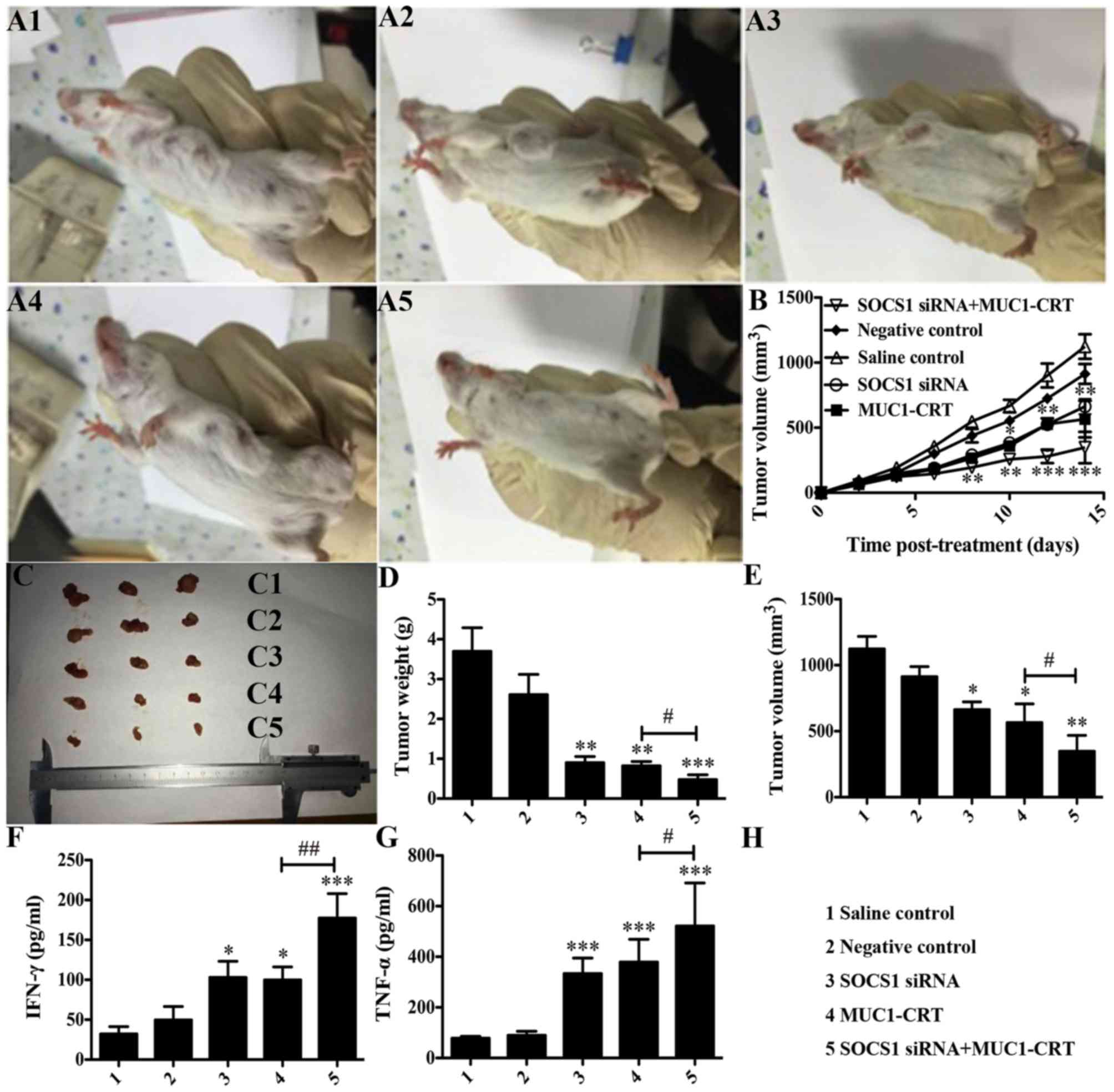 | Figure 4.Silencing of SOCS1 enhances the
immunological effect of MUC1-CRT-primed 4T1-treated DCs in
vivo. Mice were randomly divided into 5 groups (n=10 per
group). Mice in each group were administered with an intradermal
injection of 100 µl normal saline, 100 µl untreated DCs, 100 µl
SOCS1 siRNA-treated DCs, 100 µl MUC1-CRT-primed 4T1-treated DCs or
100 µl SOCS1 siRNA- and MUC1-CRT-primed 4T1-treated
(double-treated) DCs. (A) Example images demonstrate tumor sizes.
(B) Tumor volume (mm3) was assessed every other day.
Tumor sizes are presented as (C) an image, (D) tumor weight and (E)
tumor volume following sacrifice and tumor isolation 16 days after
treatment. Secretion of the cytokines, (F) IFN-γ and (G) TNF-α.
*P<0.05, **P<0.01 and ***P<0.001 vs. NC;
#P<0.05, ##P<0.01. SOCS1, suppressor of
cytokine signaling 1; siRNA, small interfering RNA; MUC1, type I
transmembrane glycoprotein mucin 1; CRT, calreticulin; DCs,
dendritic cells; IFN-γ, interferon-γ; TNF-α, tumor necrosis
factor-α. |
Furthermore, tumors were collected and depleted of
red blood cells, and IFN-γ and TNF-α production was detected. As
presented in Fig. 4F and G, compared
with NC group the double-treated BMDCs exhibited significantly
increased production of IFN-γ and TNF-α (P<0.001 vs. NC;
Fig. 4F and G), and this was also
significantly increased compared with that in the MUC1-CRT-primed
4T1-treated BMDCs (P<0.05 vs. SOCS1 siRNA2; Fig. 4F and G). These experiments indicated
that the silencing of SOCS1 combined with MUC1-CRT-primed 4T1
treatment of BMDCs may induce enhanced immunological effects in
vivo.
Discussion
DCs are considered to be the most effective APCs
with regard to their ability to prime tumor antigen-specific T
cells and initiate immune responses against tumors (6). DC-based tumor vaccines represent a
promising therapeutic approach, but have achieved limited success
in clinical settings thus far. In order to improve the efficiency
of DC maturation, our previous studies used CRT to translocate MUC1
to the 4T1 cell surface, where they worked together to induce a
strong apoptotic reaction. The results demonstrated that the
exposure of MUC1-CRT on the surface of breast cancer 4T1 cells
facilitates their uptake by DCs and the subsequent presentation of
TAAs to T cells (10).
DC maturation serves as the critical switch from the
maintenance of self-tolerance to the induction of immunity.
However, the identification of a means of overcoming inhibitory
immune regulatory mechanisms and eliciting effective T-cell
responses to antigens preferentially expressed by tumor cells
remains a major challenge. SOCS1 acts as a critical ‘brake’ in DCs
that disables their potency as a tumor vaccine. SOCS1 functions as
a negative regulator of signaling by various cytokines; for
example, Hanada et al (17)
observed that SOCS1−/− DCs exhibited a more mature
phenotype, were hyper-responsive to LPS and induced auto-reactive
antibody production. Furthermore, Shen et al (11) demonstrated that SOCS1 serves a
critical function in regulating the extent of DC antigen
presentation and hence the magnitude of adaptive immunity.
Additionally, a previous study revealed that silencing of SOCS1 in
antigen-presenting DCs enhances antigen-specific antitumor immunity
(11).
The present study attempted to further increase the
potency of MUC1-CRT-primed 4T1-treated DCs by using siRNA to
silence the expression of an endogenous molecule in DCs. To begin
with, a SOCS1 siRNA with high knockdown efficiency in the DC2.4
cell line was selected (Fig. 1) and,
subsequently, hpp10-DRBD was utilized to deliver this siRNA into
BMDCs. The use of other materials for delivery, including cationic
lipids and polymers, often fails to successfully introduce siRNAs
into the entire population of cells, particularly primary cells and
non-adherent cells, and such materials may also produce cytotoxic
effects when used at high concentrations. By contrast, the hpp10
delivery peptide has been revealed to rapidly transport various
types of biomolecules into the entire population of cells in our
previous study (14). In addition,
DRBD is able to mask the negative charge of siRNA, allowing hpp10
to efficiently deliver the siRNA into the DCs (15). In the present study, hpp10-DRBD was
demonstrated to be highly efficient in facilitating the
internalization of SOCS1 siRNA by BMDCs (Fig. 2).
The results of the present study demonstrated that
SOCS1 regulated the extent of antigen presentation by mature DCs,
thereby providing a regulatory mechanism that allows DCs to control
the magnitude and duration of adaptive immunity (Fig. 3). Vaccination with SOCS1-silenced and
MUC1-CRT-primed 4T1-treated BMDCs enhances antigen-specific
antitumor immunity (Fig. 4), and
SOCS1 silencing may permit antigen-presenting immunogenic DCs to
persistently stimulate antigen-specific T cells in vivo
(14).
Acknowledgements
The authors would like to thank the Nature Science
Foundation of China (grant nos. 81201766 and 81550028), the Hubei
Office of Education Foundation (grant no. Q20151204), the Hubei
Office of Education Foundation (grant nos. Q20151204 and B2016022)
and the Yi Chang Office of Education Foundation (grant no.
A16-301-14) for providing financial support.
References
|
1
|
Obeid M, Tesniere A, Ghiringhelli F, Fimia
GM, Apetoh L, Perfettini JL, Castedo M, Mignot G, Panaretakis T,
Casares N, et al: Calreticulin exposure dictates the immunogenicity
of cancer cell death. Nat Med. 13:54–61. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Obeid M, Tesniere A, Panaretakis T, Tufi
R, Joza N, van Endert P, Ghiringhelli F, Apetoh L, Chaput N,
Flament C, et al: Ecto-calreticulin in immunogenic chemotherapy.
Immunol Rev. 220:22–34. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ahn YH, Hong SO, Kim JH, Noh KH, Song KH,
Lee YH, Jeon JH, Kim DW, Seo JH and Kim TW: The siRNA cocktail
targeting interleukin 10 receptor and transforming growth factor-β
receptor on dendritic cells potentiates tumour antigen-specific
CD8(+) T cell immunity. Clin Exp Immunol. 181:164–178. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Steinman RM, Hawiger D and Nussenzweig MC:
Tolerogenic dendritic cells. Annu Rev Immunol. 21:685–711. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen L: Co-inhibitory molecules of the
B7-CD28 family in the control of T-cell immunity. Nat Rev Immunol.
4:336–347. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hodi FS, Mihm MC, Soiffer RJ, Haluska FG,
Butler M, Seiden MV, Davis T, Henry-Spires R, MacRae S, Willman A,
et al: Biologic activity of cytotoxic T lymphocyte-associated
antigen 4 antibody blockade in previously vaccinated metastatic
melanoma and ovarian carcinoma patients. Pro Natl Acad Sci USA.
100:4712–4717. 2003. View Article : Google Scholar
|
|
7
|
Wang J, Gao ZP, Qin S, Liu CB and Zou LL:
Calreticulin is an effective immunologic adjuvant to tumor
associate antigen. Exp Ther Med. 14:3399–3406. 2017.PubMed/NCBI
|
|
8
|
Demaria O, Gassart A, Coso S, Gestermann
N, Di Domizio J, Flatz L, Gaide O, Michielin O, Hwu P, Petrova TV,
et al: STING activation of tumor endothelial cells initiates
spontaneous and therapeutic antitumor immunity. Proc Natl Acad Sci
USA. 112:pp. 15408–15413. 2015; View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ghosh S, Sarkar M, Ghosh T, Guha I,
Bhuniya A, Saha A, Dasgupta S, Batik S, Bose A and Baral R: Neem
leaf glycoprotein promotes dual generation of central and effector
memory CD8(+) T cells against sarcoma antigen vaccine to induce
protective anti-tumor immunity. Mol Immunol. 71:42–53. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gardai SJ, McPhilips KA, Frasch SC,
Janssen WJ, Starefeldt A, Murphy-Ullrich JE, Bratton DL, Oldenborg
PA, Michalak M and Henson PM: Cell-surface calreticulin initiates
clearance of viable or apoptotic cells through trans-activation of
LRP on the phagocyte. Cell. 123:321–334. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shen L, Evel-Kabler K, Strube R and Chen
SY: Silencing of SOCS1 enhances antigen presentation by dendritic
cells and antigen-specific anti-tumor immunity. Nat Biotechnol.
22:1546–1553. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kim JH, Kang TH, Noh KH, Kim SH, Lee YH,
Kim KW, Bae HC, Ahn YH, Choi EY, Kim JS, et al: Enhancement of DC
vaccine potency by activating the PI3K/AKT pathway with a small
interfering RNA targeting PTEN. Immunol Lett. 134:47–54. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim JH, Kang TH, Noh KH, Bae HC, Ahn YH,
Lee YH, Choi EY, Chun KH, Lee SJ and Kim TW: Blocking the
immunosuppressive axis with small interfering RNA targeting
interleukin (IL)-10 receptor enhances dendritic cell-based vaccine
potency. Clin Exp Immunol. 165:180–189. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang H, Ma JL, Yang YG, Song Y, Wu J, Qin
YY, Zhao XL, Wang J, Zou LL, Wu JF, et al: Efficient therapeutic
delivery by a novel cell-permeant peptide derived from KDM44
protein for antitumor and antifibrosis. Oncotarget. 7:49075–49090.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Eguchi A, Meade BR, Chang YC, Fredrickson
CT, Willert K, Puri N and Dowdy SF: Efficient siRNA delivery into
primary cells by a peptide transduction domain-ds RNA binding
domain fusion protein. Nat Biotechnol. 27:567–571. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yang H, Li S, Li F, Yu K, Yang F and Xiang
J: Recombinant expression of a modified shrimp
anti-lipopolysaccharide factor gene in Pichia pastoris GS115 and
its characteristic analysis. Mar Drugs. 14:E1522016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hanada T, Yoshida H, Kato S, Tanaka K,
Masutani K, Tsukada J, Nomura Y, Mimata H, Kubo M and Yoshimura A:
Suppressor of cytokine signaling-1 is essential for suppressing
dendritic cell activation and system autoimmunity. Immunity.
19:437–450. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kumar A, Kundu S and Debnath DM:
Expression, purification and evaluation of recombinant lipoprotein
of Salmonella typhi as a vaccine candidate. Biologicals.
46:108–113. 2017. View Article : Google Scholar : PubMed/NCBI
|