Introduction
Cell culture is important for the evaluation of cell
physiology and cellular responses to pharmaceutical compounds
(1,2).
Many cell lines have been established and cultured in appropriate
media such as minimum essential medium (MEM), Dulbecco's Modified
Eagle Medium (DMEM), and RPMI-1640. Culture media may include
glucose, amino acids, vitamins, inorganic salts, and serum; each
medium comprises different kinds and quantities of components. To
perform precise evaluations, researchers must select the medium
appropriate for the cells in their research.
Vitamin B6 comprises pyridoxine (PN),
pyridoxal (PL), pyridoxamine, and phosphorylated forms, such as
pyridoxine-5′-phosphate, pyridoxal-5′-phosphate (PLP) and
pyridoxamine-5′-phsphate (3). It acts
as a coenzyme for amino acid metabolism. In general, DMEM is used
with 20 µM PN or PL. Although it is suggested that the difference
between these vitamin B6 compounds does not affect cell
proliferation, high concentration of vitamin B6 did
inhibit cell growth in several cancer cells, and the effect of PL
was stronger than that of PN (4–7).
Conversely, the influence of optimal concentrations on other cell
physiological effects is poorly understood. In this study, we
evaluated the effects of PL and PN on cell growth and melanogenesis
in B16F10 murine melanoma cells.
Materials and methods
Materials
PL hydrochloride (P6155) and PN hydrochloride
(P9755) were purchased from Sigma-Aldrich (St. Louis, MO, USA).
DMEM without vitamin B6 was manufactured by Funakoshi
(Tokyo, Japan). Hoechst 33342 and propidium iodide (PI) were
purchased from Dojindo Molecular Technologies, Inc., (Kumamoto,
Japan) and Sigma-Aldrich; Merck KGaA, (Darmstadt, Germany),
respectively. 3-isobutyl-1-methylxanthine (IBMX) was obtained from
Sigma-Aldrich; Merck KGaA. Block Ace was purchased from Dainippon
Sumimoto Pharma Co., Ltd., (Osaka, Japan). Antibodies to tyrosinase
(sc-7834), PARP (no. 9542), and β-actin (AC-15, A-5441) were
obtained from Santa Cruz Biotechnology, Inc., (Dallas, TX, USA),
Cell Signaling Technology, Inc., (Danvers, MA, USA), and
Sigma-Aldrich, respectively. ECL Prime Western Blotting Detection
Reagent was purchased from GE Healthcare (Chicago, IL, USA).
Cell culture
B16F10 cells were kindly gifted by Prof. Naoto Oku
(School of Pharmaceutical Sciences, University of Shizuoka, Japan).
The cells were maintained in DMEM without vitamin B6 and
supplemented with 10% heat-inactivated fetal bovine serum (FBS)
under 5% CO2 at 37°C. They were cultured in DMEM without
vitamin B6 for more than 1-week before being subjected
to analysis.
Cell proliferation and viability
assay
Cell proliferation and viability assays examined the
effect of vitamin B6 on B16F10 cells. To assess the
effect of vitamin B6, the cells were seeded at
1×105 cells/ml medium into 96-well plates in the
presence of PL or PN at 20–500 µM for 72 h. The cells were then
counted using trypan blue staining. To analyze the cell survival
rate, both attached and detached cells were counted; the ratio of
attached cell numbers was calculated as viable cells. To examine
cell survival in detail, Hoechst-PI staining was performed. Hoechst
and PI were used at 2 µg/ml.
To analyze the effect of hydrogen peroxide
(H2O2) on cell proliferation, B16F10 cells
were seeded at 1×105 cells/ml medium into 96-well plates
in the presence of PL or PN at 20 µM for 24 h. The cells were then
added with H2O2 at concentrations of 1–10 µM.
After 24 h treatment, survival cells were counted by trypan blue
staining.
Western blot analysis
Western blot analysis was performed as previously
described (8,9). The cells were treated with 100 µM IBMX
for 24 h. The proteins were separated by SDS-PAGE and transferred
onto nitrocellulose membranes. The membranes were blocked with 4%
Block Ace solution. Anti-β-actin, anti-PARP, and anti-tyrosinase
antibody were used at 1:10,000, 1:1,500, and 1:250, respectively.
The membrane was next incubated with HRP-conjugated secondary
antibody. ECL Prime Western Blotting Detection Reagent and LAS-3000
(Fuji-Film, Tokyo, Japan) were used for detection. Finally, the
expression levels of tyrosinase were assessed by the Scion Image
Software (Scion, Frederick, MD, USA) (n=3).
Measurement of melanin contents
B16F10 cells were seeded at 1×105
cells/ml medium into 6-well plates in the presence of 20 µM PL or
PN. Following 24 h incubation, cells were treated with 100 µM IBMX
and cultured further for 48 h. To analyze melanin contents,
1.5×106 cells were dissolved in 2N NaOH for 2 h at 80°C.
The absorbance at 450 nm was measured to determine melanin
content.
Statistical analysis
To assess statistical significance, the cell growth
rate was determined by Student's t-test and the results were
confirmed by one-way analysis of variance (ANOVA). The dead cell
rate was analyzed by one-way ANOVA with Dunnett's post hoc test for
the comparison of three groups. The melanin content and tyrosinase
expression were evaluated by Student's t-test. P<0.05 was
considered to indicate a statistically significant difference.
Results
The effect of vitamin B6
compounds on cell growth in B16F10 melanoma cells
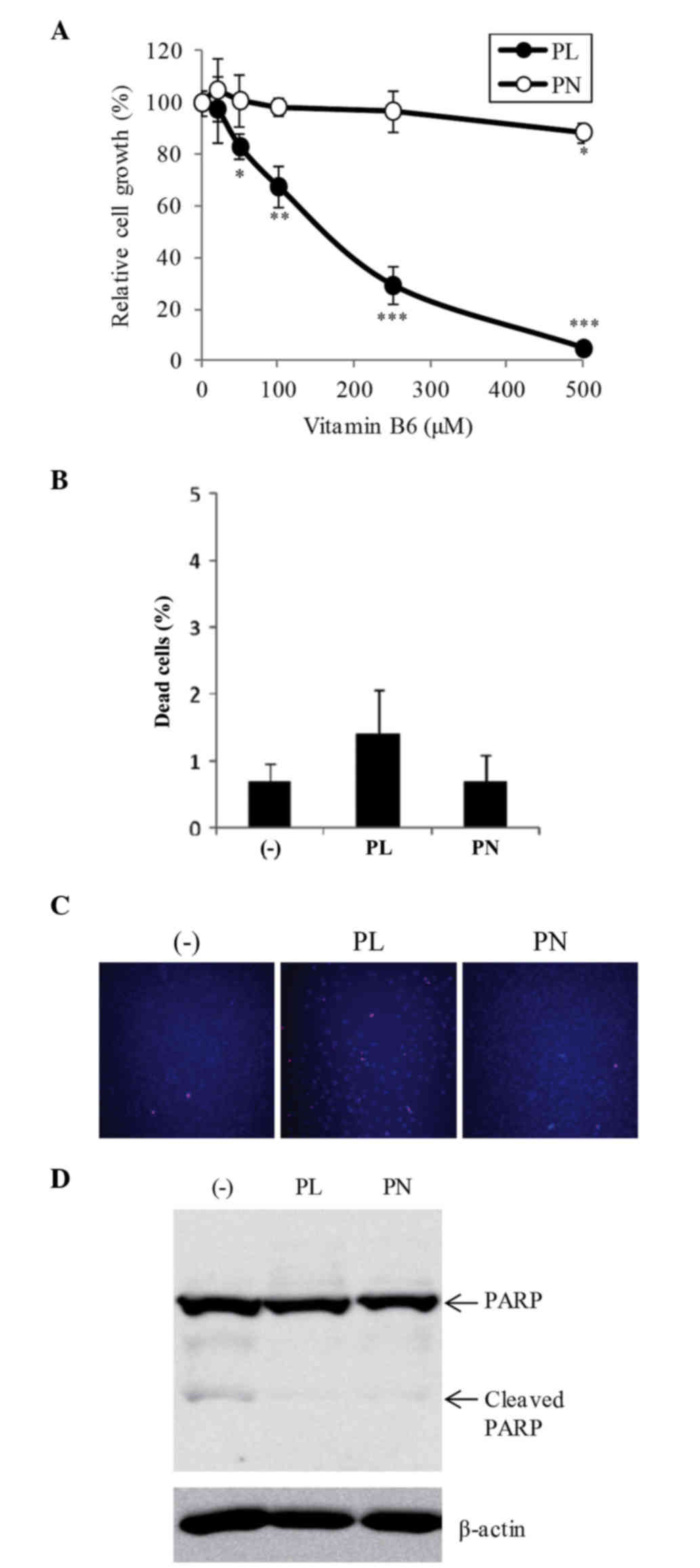
To investigate the effect of PL and PN on cell
proliferation in B16F10 cells, the cells were treated with
different concentrations (20–500 µM) of both PL and PN. PL at
50–500 µM significantly suppressed cell growth compared with DMEM
without vitamin B6 (Fig.
1A). At 500 µM, PL showed 95.1% inhibition of cell
proliferation, whereas PN showed only 11.4% inhibition (Fig. 1A). The dead cell ratio among
VB6 free, PL, and PN was no significant difference
(Fig. 1B). We further confirmed the
effects of vitamin B6 by Hoechst 33342 and PI staining.
Hoechst 33342 stains both viable and dead cells, and the condensed
chromatin of apoptotic cells are observed more brightly than the
nuclei of viable cells. PI stains dead cells such as apoptotic
cells and necrotic cells. At 500 µM PL, although the number of
Hoechst-stained cells was significantly decreased, PI-stained cells
were scarcely increased (Fig. 1C).
The slightly dead cells were detected the condensed chromatin of
apoptotic cells (data not shown). The cleavage of PARP, a hallmark
of apoptosis, was hardly observed at 500 µM PL or PN (Fig. 1D). On the other hand, cell morphology
by PL or PN did not change (data not shown). These results showed
that 500 µM PL strongly suppressed cell growth, but did not induce
cell death. Otherwise, 20 µM of both vitamin B6
compounds, which are the normal concentration of DMEM, did not
affect cell proliferation (Fig.
1A).
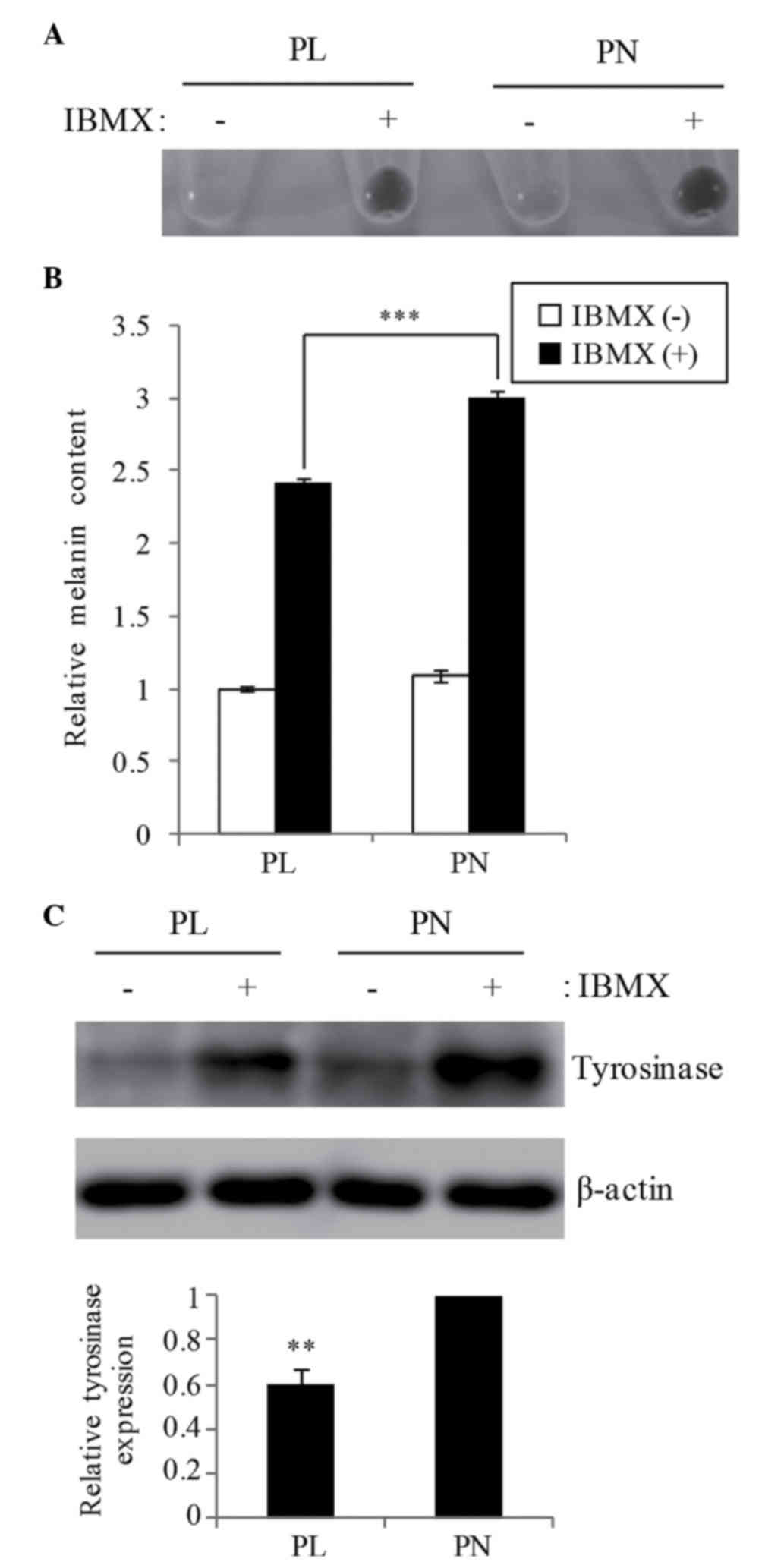
Melanogenesis in B16F10 cells
The effect of 20 µM PL and PN on IBMX-stimulating
melanogenesis in B16F10 cells was examined. The total melanin
content was increased in both PL and PN by IBMX stimulation,
whereas the melanin content in the presence of PL was 19.9% lower
than that in PN (Fig. 2A and B). The
melanin content in PN was similar to that in the IBMX-only cells
without the addition of PL/PN (data not shown). Western blot
analysis showed that tyrosinase expression in PL is lower when
compared with PN (Fig. 2C). The
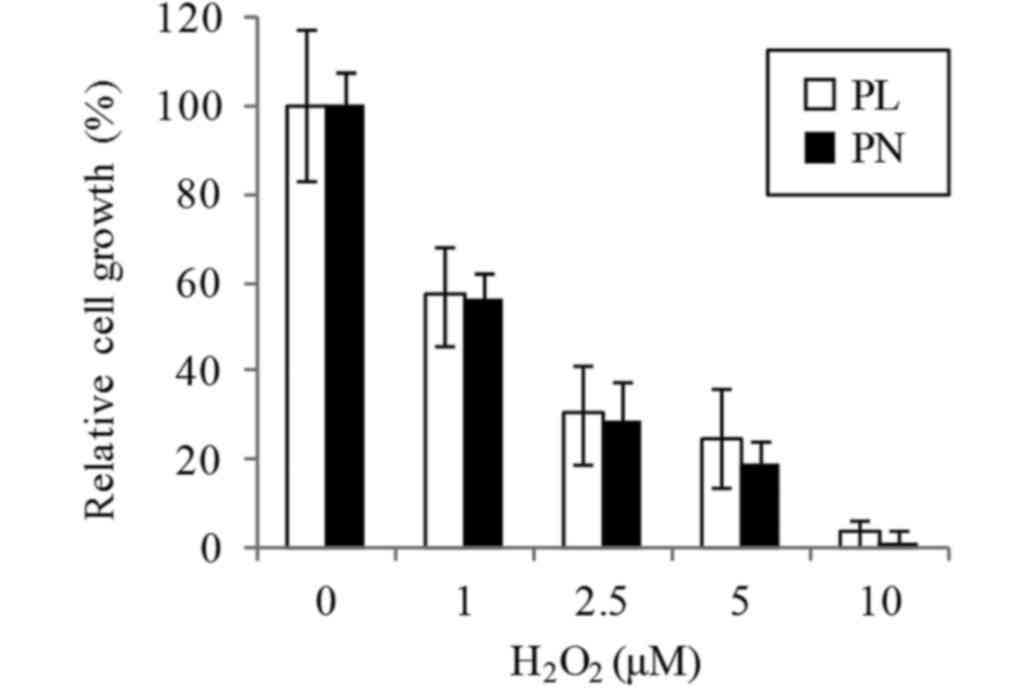
oxidation also involved melanogenesis. We confirmed that the cell
resistance to oxidative stress by H2O2 is not
different media containing PL or PN (Fig.
3).
Discussion
In DMEM, either PL or PN is used as a vitamin
B6 compound. Although the difference between these
vitamin B6 compounds does not affect cell growth, the
influence on other physiological effects is poor understood. In the
present study, we compared the effects of PL and PN in DMEM on cell
proliferation and IBMX-stimulating melanogenesis in B16F10 cells.
We clarified 500 µM PL inhibits cell proliferation of B16F10 cells.
The cleavage of PARP was hardly observed, presumably because few
apoptotic cells were observed. The purity of PL is ensured more
than 99%, and the other 1% components include less than 0.5% water.
In addition, high concentration of PL is also reported to inhibit
cell growth through the reduction of nuclear acid precursor uptake
in B16 and human melanoma cells (5,6). We could
think the effect by PL is not for other impurities. In an animal
model, vitamin B6 suppressed colon tumorigenesis by
reducing cell growth (10).
Epidemiologic research reported that vitamin B6 can
protect colon against cancer (11).
B16F10 cells are highly malignant compared to B16 cells. Thus, PL
might be effective in melanoma therapy. Conversely, the inhibitory
effect of PN on cell growth in B16F10 cells is less than that in
PL. In MCF-7 cells, growth suppression by PN was weak compared with
PL (6,7). In general, vitamin B6
functions after its conversation to an active form, PLP. Although
PL is converted to PLP by pyridoxal kinase, conversion of PN to PLP
requires two enzymes, pyridoxal kinase and pyridoxamine oxidase
(3). It has been reported that
intracellular levels of PLP increased in cells exposed to
pharmacologic PL concentrations (0.05–0.5 mM), but not PN in
M21-HPB human melanoma cells (5).
Therefore, the antitumor effect of PL appears to be strong.
However, in RAW264.7 cells, the addition of PL could not increase
intracellular PLP concentration (12). The cell inhibitory effect of PL might
not be dependent on PLP. Further studies are required to clarify
this mechanism.
The synthesis of melanin initiates the conversion of
L-tyrosine to L-dopa and then to dopaquinone by tyrosinase, which
is the rate-limiting enzyme (13).
Dopaquinone spontaneously proceeds to melanin by auto-oxidation. We
found that the inhibition of melanogenesis by PL is stronger than
that by PN. Although anti-oxidative effect of between PL and PN is
not different, tyrosinase expression in the presence of PL was
lower than that in PN by correlation with melanin contents. PN did
not suppress the melanogenesis compared with vitamin B6
free condition. A previous study showed that PL suppressed
lipopolysaccharide-stimulating COX-2 expression more strongly than
PN in RAW264.7 murine macrophage cells (12). PL might inhibit IBMX-stimulating
tyrosinase expression compared with PN. However, melanogenesis is
affected by the media type, pH, and glucose concentration (14–16).
Therefore, we speculated that the difference of cellular
metabolism, such as amino acids by PL and PN, might change the
cellular environment, and therefore, alter melanogenesis and
tyrosinase expression. Additionally, the present study lacks the
melanin quantification in normal cells. To understand the
inhibitory mechanism of melanogenesis by vitamin B6 in
detail, we need to analyze using not only other melanoma cells but
also normal melanocytes.
In conclusion, we showed the different effects of PL
and PN on B16F10 cells. PL might help melanoma therapy and
suppression of skin pigmentation compared to PN. In addition, we
expect that our results offer the researchers an opportunity to
consider the selection of an effective medium for cell culture.
Acknowledgements
The authors thank Prof. Naoto Oku (School of
Pharmaceutical Sciences, University of Shizuoka, Japan) for the
gift of B16F10 cells.
References
|
1
|
Nema R and Khare S: An animal cell
culture: Advance technology for modern research. Adv Biosci
Biotechnol. 3:219–226. 2012. View Article : Google Scholar
|
|
2
|
Galluzzi L, Vitale I, Senovilla L,
Olaussen KA, Pinna G, Eisenberg T, Goubar A, Martins I, Michels J,
Kratassiouk G, et al: Prognostic impact of vitamin B6 metabolism in
lung cancer. Cell Rep. 2:257–269. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Galluzzi L, Vacchelli E, Michels J, Garcia
P, Kepp O, Senovilla L, Vitale I and Kroemer G: Effects of vitamin
B6 metabolism on oncogenesis, tumor progression and therapeutic
responses. Oncogene. 32:4995–5004. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
DiSorbo DM and Nathanson L: High-dose
pyridoxal supplemented culture medium inhibits the growth of a
human malignant melanoma cell line. Nutr Cancer. 5:10–15. 1983.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shultz TD, Santamaria AG, Gridley DS,
Stickney DR and Slater JM: Effect of pyridoxine and pyridoxal on
the in vitro growth of human malignant melanoma. Anticancer Res.
8:1313–1318. 1988.PubMed/NCBI
|
|
6
|
Minamino M, Oka T and Kanouchi H: Growth
suppression and cell death by pyridoxal is dependent on p53 in the
human breast cancer cell line MCF-7. Biosci Biotechnol Biochem.
79:124–129. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nakari M, Kanouchi H and Oka T: High dose
of pyridoxine induces IGFBP-3 mRNA expression in MCF-7 cells and
its induction is inhibited by the p53-specific inhibitor
pifithrin-α. J Nutr Sci Vitaminol (Tokyo). 57:280–284. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Matsuo T, Yamamoto T, Katsuda C, Niiyama
K, Yamamoto A, Yamazaki N, Ohkura K, Kataoka M and Shinohara Y:
Substitution of certain amino acids in a short peptide causes a
significant difference in their immunoreactivities with antibodies
against different epitopes: Evidence for possible folding of the
peptide on a nitrocellulose or PVDF membrane. Biologicals.
37:44–47. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Matsuo T, Komatsu M, Yoshimaru T, Kiyotani
K, Miyoshi Y, Sasa M and Katagiri T: Involvement of B3GALNT2
overexpression in the cell growth of breast cancer. Int J Oncol.
44:427–434. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Komatsu SI, Watanabe H, Oka T, Tsuge H,
Nii H and Kato N: Vitamin B-6-supplemented diets compared with a
low vitamin B-6 diet suppress azoxymethane-induced colon
tumorigenesis in mice by reducing cell proliferation. J Nutr.
131:2204–2207. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ishihara J, Otani T, Inoue M, Iwasaki M,
Sasazuki S and Tsugane S; Japan Public Health Center-based
Prospective Study Group, : Low intake of vitamin B-6 is associated
with increased risk of colorectal cancer in Japanese men. J Nutr.
137:1808–1814. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kanouchi H, Shibuya M, Tsukamoto S,
Fujimura Y, Tachibana H, Yamada K and Oka T: Comparisons of uptake
and cell surface binding among pyridoxal, pyridoxine, and
pyridoxamine in RAW264.7 cells. Nutrition. 26:648–652. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Parvez S, Kang M, Chung HS, Cho C, Hong
MC, Shin MK and Bae H: Survey and mechanism of skin depigmenting
and lightening agents. Phytother Res. 20:921–934. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wolnicka-Glubisz A, Nogal K, Żądło A and
Płonka PM: Curcumin does not switch melanin synthesis towards
pheomelanin in B16F10 cells. Arch Dermatol Res. 307:89–98. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Laskin JD, Mufson RA, Weinstein IB and
Engelhardt DL: Identification of a distinct phase during
melanogenesis that is sensitive to extracellular pH and ionic
strength. J Cell Physio. 103:467–474. 1980. View Article : Google Scholar
|
|
16
|
Nakayasu M, Saeki H, Tohda H and Oikawa A:
Effects of sugars on melanogenesis in cultured melanoma cells. J
Cell Physiol. 92:49–55. 1977. View Article : Google Scholar : PubMed/NCBI
|