Introduction
Gliomas are the most common primary brain tumor in
adults and are categorized into four World Health Organization
grades on the basis of histopathological characteristics (1). Despite current multidisciplinary
treatments including surgery, radiotherapy and chemotherapy, no
significant prognostic improvement has been obtained in patients
with gliomas over the last decade, because of the aggressive nature
of high-grade gliomas, the survival rates of patients with
high-grade gliomas are <10% at 5 years (2). Previously, efforts have been made to
identify new effective therapeutic agents.
Momordica anti-human immunodeficiency virus
protein of 30 kDa (MAP30), first extracted and purified from
Momordica charantia in 1990, is a type I
ribosome-inactivating protein (RIP) (3). Previous studies indicated that MAP30
exhibits a variety of anti-infection, anti-diabetic, antiviral and
antitumor bioactive effects (4–10). The
antitumor ability of MAP30 has been the subject of previous studies
(11–15). However, certain researchers have
already noted the effects of MAP30 on the inhibition of U87 cells
in screening for anticancer effects (3,7,16,17).
However, the mechanism of MAP30 on glioma cells has not been
elucidated in detail. The aim of the present study was to
investigate the effects and mechanism of MAP30 on U87 and U251
glioblastoma cell lines in vitro. Although some researchers
identified that the DNA profile of U87MG was different from that of
the original cells in 2016, it is likely to be a bona fide human
glioblastoma cell line of unknown origin; therefore, in the present
study the U87 and U251 cell lines were used.
Materials and methods
Materials
High-glucose Dulbecco's modified Eagle's medium
(DMEM), fetal bovine serum (FBS), penicillin G and streptomycin
were all purchased from Gibco; Thermo Fisher Scientific, Inc.
(Waltham, MA, USA). The U87 and U251 human malignant glioblastoma
cell lines were purchased from China Infrastructure of Cell Line
Resources (Beijing, China). MAP30 was kindly provided by Professor
Y.F. Meng (College of Life Sciences, Sichuan University, Chengdu,
China) (18). The primary antibodies
against leucine-rich-repeat-containing G-protein-coupled receptor 5
(LGR5; cat. no. AB75850), β-catenin (cat. no. AB32572), glycogen
synthase kinase 3β phosphorylated at Ser9
(pGSK-3βSer9; cat. no. AB75814), c-Myc (cat. no.
AB32072), cyclin D1 (cat. no. AB134175) and second
mitochondria-derived activator of caspase (Smac; cat. no. AB32023)
used for western blot analysis were purchased from Abcam
(Cambridge, UK).
Cell culture
U87 and U251 cells were cultured in DMEM
supplemented with 10% FBS, 100 U/ml penicillin and 100 U/ml
streptomycin at 37°C in a humidified atmosphere of 5%
CO2 in air. The medium was changed every 3–4 days, and
cultures were split using 0.25% trypsin. All experiments were
carried out on cells with viability >95%.
Cell viability assay
When U87 and U251 cells reached the exponential
growth phase, they were treated with MAP30 (2.7 µM) or PBS (0.01 M,
pH 7.2–7.4; Gibco; Thermo Fisher Scientific, Inc.), and images were
captured under an inverted light microscope (×400 magnification;
Olympus CKX41; Olympus Corporation, Tokyo, Japan) at 0 and 24
h.
A Cell Counting Kit-8 (CCK-8; Dojindo Molecular
Technologies, Inc., Kumamoto, Japan) assay was used to evaluate
cell viability. U87 and U251 cells were resuspended in DMEM with
10% FBS at a density of 5×103 cells/well and cultured in
96-well plates. The cells were treated with various concentrations
(0.5, 1, 2, 4, 8 and 16 µM) of MAP30 for 24, 48 and 72 h, prior to
medium being replaced with 10 µl CCK-8 and 100 µl FBS-free DMEM.
Cells were incubated for 1 h at 37°C and the absorbance of each
well was measured at 450 nm using a microplate reader.
Cell migration and invasion
assays
The effect of MAP30 on glioma cell migration was
assessed using wound-healing assays. U87 and U251 cells were seeded
in multiwell plates and cultured until reaching confluence. A 10-µl
pipette tip was used to make a straight scratch. The cells were
washed with PBS, and treated for 24 h with MAP30 (2.7 µM) or PBS
(0.01 M, pH 7.2–7.4) as a control. Images of the wounds were
captured under an inverted light microscope (×400 magnification) at
0 and 24 h, and measured.
A cell invasion assay was performed using 24-well
Transwell chambers with an 8 µm aperture (Costar; Corning
Incorporated, Corning, New York, NY, USA). The inserts were coated
with Matrigel (BD Biosciences, Franklin Lakes, NJ, USA). U87 and
U251 cells were trypsinized and transferred to the upper Matrigel
chamber with FBS-free DMEM at a density of 1×105
cells/well. The lower chamber was filled with DMEM containing 20%
FBS as a chemoattractant. After 24 h of incubation at 37°C, the
non-invasive cells on the upper membrane surface were gently
removed with a cotton tip, and the cells that passed through the
filter were stained with hematoxylin at room temperature for 1 h
and counted under an inverted light microscope (×400
magnification).
Plate colony formation assays
Colony formation ability of the glioblastoma cell
lines was determined using plate colony formation assays. Briefly,
U87 and U251 cells were cultured in DMEM containing 10% FBS in
6-well plates at a concentration of 1,000 cells/well. The cells
were treated with MAP30 0.51 and 0.60 µM at 1/4 of half-maximal
inhibitory concentration (IC50) for U87 and U251 cells
at 48 h. or PBS. After 7 days, the colonies were washed gently with
PBS, fixed with 4% paraformaldehyde at room temperature for 5 min
and stained with Giemsa (Sigma; Merck KGaA, Darmstadt, Germany) at
room temperature for 30 min; images were captured under an inverted
light microscope (×400 magnification) and quantified.
Hoechst 33342/propidium iodide (PI)
staining
U87 cells were treated for 24 h with MAP30 (1.4 and
2.7 µM) or PBS (0.01 M, pH 7.2–7.4), fixed with ice-cold 70%
ethanol (4°C), washed three times with PBS, stained with Hoechst
33342/PI solution (50 µg/ml, Sigma-Aldrich; Merck KGaA) for 10 min
and washed three times with PBS. The nuclear morphological changes
were observed and images were captured using fluorescence
microscopy (×400 magnification; EVOS-FL; Thermo Fisher Scientific,
Inc.).
Cell cycle and apoptosis assays
U87 cells were cultured in 25 ml flasks for 24 h
with MAP30 (1.4 and 2.7 µM) or PBS (0.01 M, pH 7.2–7.4). For cell
cycle analysis, the cells were harvested, washed, fixed with
ice-cold 70% ethanol at −20°C overnight, prior to washing with PBS
and suspended in 300 µl staining buffer (BD Biosciences) containing
10 µl propidium iodide (PI) and 5 µl RNase (BD Biosciences) for 15
min at room temperature in the dark.
An annexin V-fluorescein isothiocyanate (FITC)/PI
Apoptosis Detection kit (BD Biosciences) was used to determine the
effects of MAP30 on apoptosis of U87 cells. The cells were
harvested, washed twice with ice-cold PBS, and resuspended in
binding buffer at a concentration of 1×106 cells/ml. A
500 µl volume of the solution was transferred into a 5-ml culture
tube, and 5 µl annexin V-FITC and 5 µl PI were added to the
solution with slight shaking, prior to incubation for 15 min at
room temperature in the dark.
Samples prepared using the aforementioned methods
were analyzed using flow cytometry (FC500 instrument; Beckman
Coulter, Inc., Brea, CA, USA) within 1 h of staining.
Western blotting
U87 cells were incubated with MAP30 at 1.4 and 2.7
µM for 24 and 48 h, and total cell proteins were extracted with
ice-cold lysis buffer (1 mM EDTA, pH 8.0, 50 mM Tris/HCl, pH 7.4,
150 mM NaCl, 1% NP-40, 0.1% SDS and 0.5% sodium deoxycholate, pH
7.4). After 30 min on ice, cell debris was removed by
centrifugation at 14,000 × g for 20 min at 4°C. A basic protein
quantification kit (BioVision, Inc., CA, USA) was used to determine
total protein concentrations. Equal amounts (20 µg) of whole cell
protein extracts from each sample were size-fractionated by 8%
SDS-PAGE and transferred onto a polyvinyl difluoride membrane
(BioVision, Inc., Milpitas, CA, USA). Membranes were first blocked
in 5% non-fat milk powder (suspended in Tris-buffered saline with
Tween-20; cat. no. AB64204) at room temperature for 2 h, and
incubated with a specific primary antibody (using GAPDH as a
control; cat no. AB181602; dilution 1:2,000; Abcam) overnight at
4°C. Following washing with PBS containing Tween-20, membranes were
incubated with a horseradish peroxidase-conjugated secondary
antibody (cat. no. AB6721; Abcam; dilution, 1:3,000) at room
temperature for 2 h, and protein bands were developed with Amersham
ECL Western Blotting Detection reagent (GE Healthcare Life
Sciences, Little Chalfont, UK) and visualized using a gel imaging
analysis system (Bio-Rad Laboratories, Inc., Hercules, CA,
USA).
Statistical analysis
Results are expressed as the mean ± standard
deviation of multiple independent experiments. Statistical analyses
were performed using two-way analysis of variance with
Student-Newman-Keuls post hoc test or Student's t-test using SPSS
software (version 20.0; IBM Corp., Armonk, NY, USA). The
IC50 value was estimated from CCK-8 dose-response data
via linear regression and log transformation. P<0.05 was
considered to indicate a statistically significant difference.
Results
MAP30 affects the viability of
glioblastoma cells
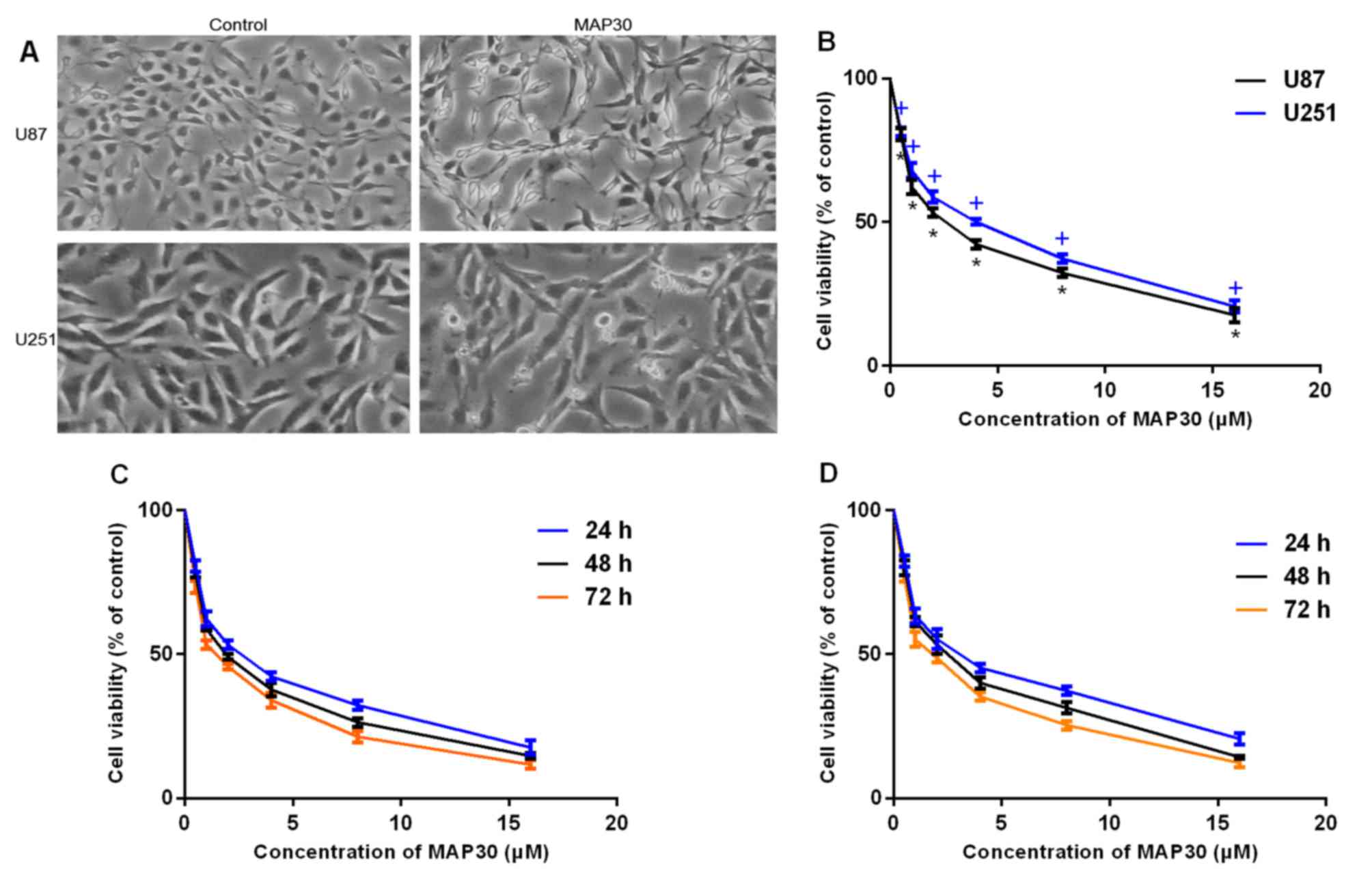
After treat with MAP30 at IC50
concentration for 24 h, the morphological changes in the glioma
cells were observed (Fig. 1A).
Following treatment of U87 and U251 cells with different
concentrations of MAP30 (0.5, 1, 2, 4, 8 and 16 µM) for 24–72 h,
cell viability was measured by CCK-8. It was identified that MAP30
significantly inhibited the viability of U87 and U251 cells
(Fig. 1B). The IC50 values
for MAP30 were identified to be 2.7±0.22 and 3.9±0.18 µM at 24 h,
2.03±0.03 and 2.4±0.03 µM at 48 h, and 1.6±0.15 and 1.9±0.16 µM at
72 h for U87 and U251 cells, respectively (Fig. 1C and D). These results reveal a time-
and dose-dependent effect of the action of MAP30.
Effects of MAP30 on cell migration and
invasion
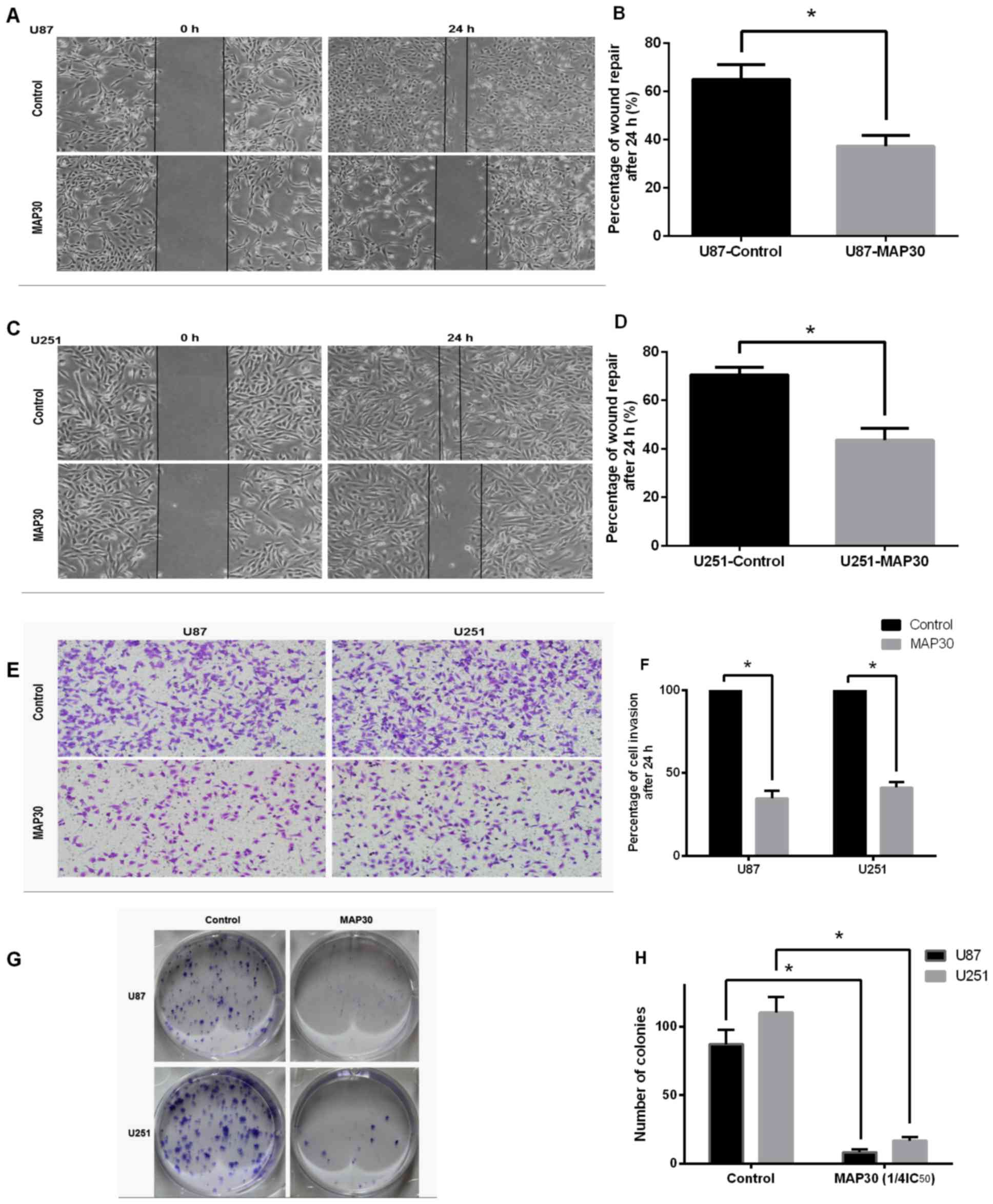
Wound healing assays and Transwell assays were used
to determine the effects of MAP30 on the migration and invasion of
glioblastoma cells. Fig. 2A presents
the wound healing of U87 cells treated with MAP30 in comparison
with the control. The wound healing rate was 37.3% for
MAP30-treated U87 cells compared with 60% for the control cells,
representing a significant decrease (P<0.05; Fig. 2B). Fig.
2C presents the wound healing of U251 cells treated with MAP30
in comparison with the control. The wound healing rate was 43.5%
for MAP30-treated U251 cells compared with 70.5% for the control
cells, representing a significant decrease (P<0.05; Fig. 2D). Fig.
2E presents images of the invading control and MAP30-treated
U87 and U251 cells on the bottom of the membrane. The proportions
of invading MAP30-treated U87 and U251 cells were respectively
decreased to 34.7 and 41.3% compared with their control
counterparts (P<0.05; Fig.
2F).
Plate colony formation assays
Compared with cells treated with PBS, application of
MAP30 at 0.51 and 0.60 µM (1/4 of the IC50 for U87 and
U251 cells, respectively, at 48 h) significantly suppressed colony
formation of U87 and U251 cells. The number of colonies formed was
5 and 12 for MAP30-treated U87 and U251 cells, respectively,
compared with 78 and 113 for control U87 and U251 cells,
respectively, representing a significant decrease (P<0.05;
Fig. 2G).
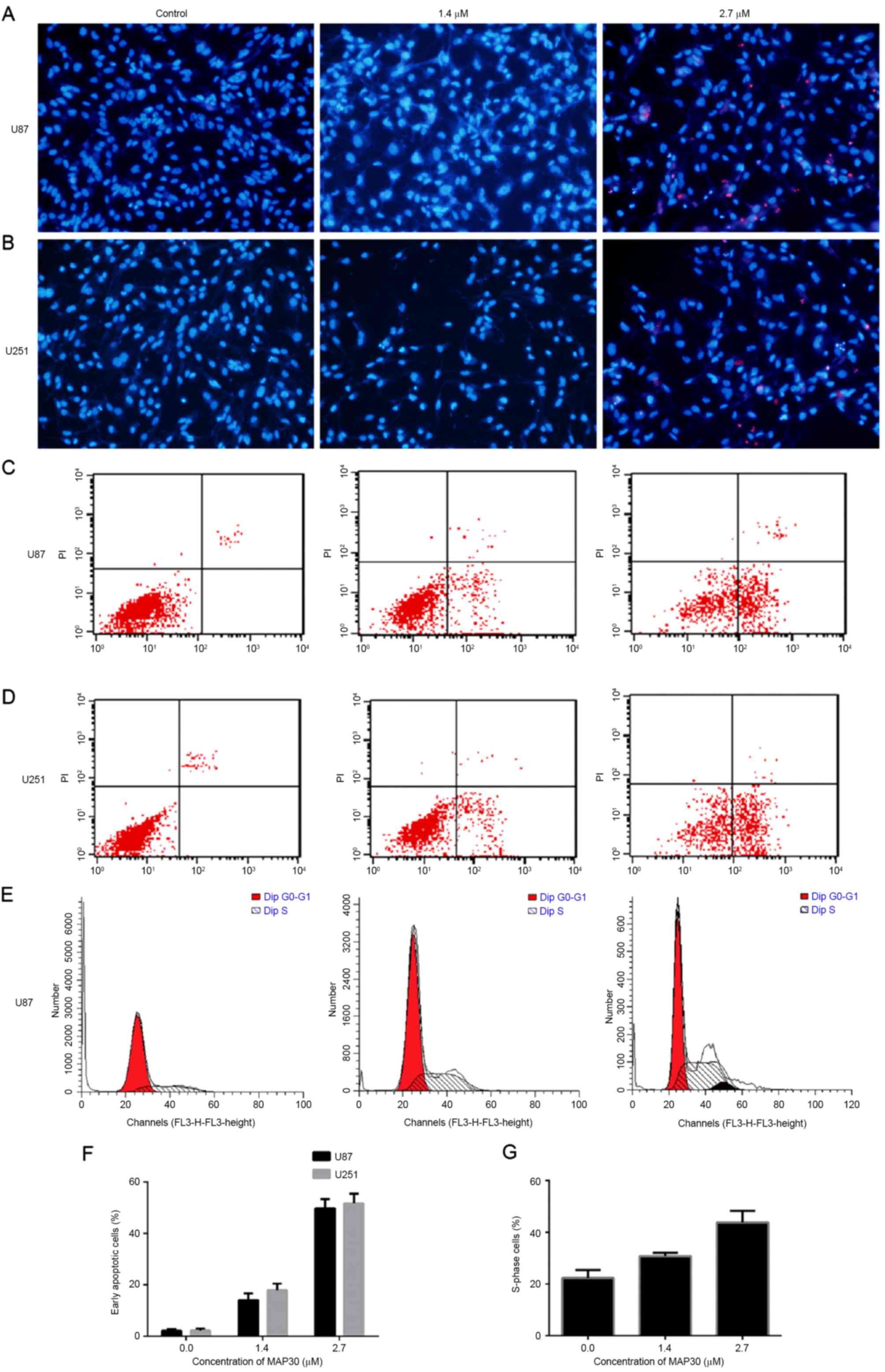
Cell apoptosis and cycle assays
Following Hoechst 33342/PI double staining, U87 and
U251 cells treated with MAP30 exhibited typical apoptotic
morphological changes including chromatic agglutination,
karyopyknosis and nuclear fragmentation (Fig. 3A and B).
After 24 h of treatment with MAP30 at a dose of 1.4
µM, early apoptosis was increased to 13.87 and 16.77% in the U87
and U251 cell lines, respectively. The early apoptosis ratio
increased to 49.61 and 51.17% in these two cell lines when the
concentration of MAP30 was increased to 2.7 µM (Fig. 3C and D). The baseline early apoptotic
proportions were 0.08 and 0.25%, respectively, for U87 and U251
cells treated with PBS (P<0.05; Fig.
3F).
Cell cycle assays indicated that MAP30 induced
S-phase arrest in U87 cells. The proportions of S-phase U87 and
U251 cells treated with MAP30 (1.4 and 2.7 µM) for 24 h
significantly increased from 21.73% (PBS-treated) to 30.79 and
41.33%, respectively (P<0.05; Fig. 3E
and G).
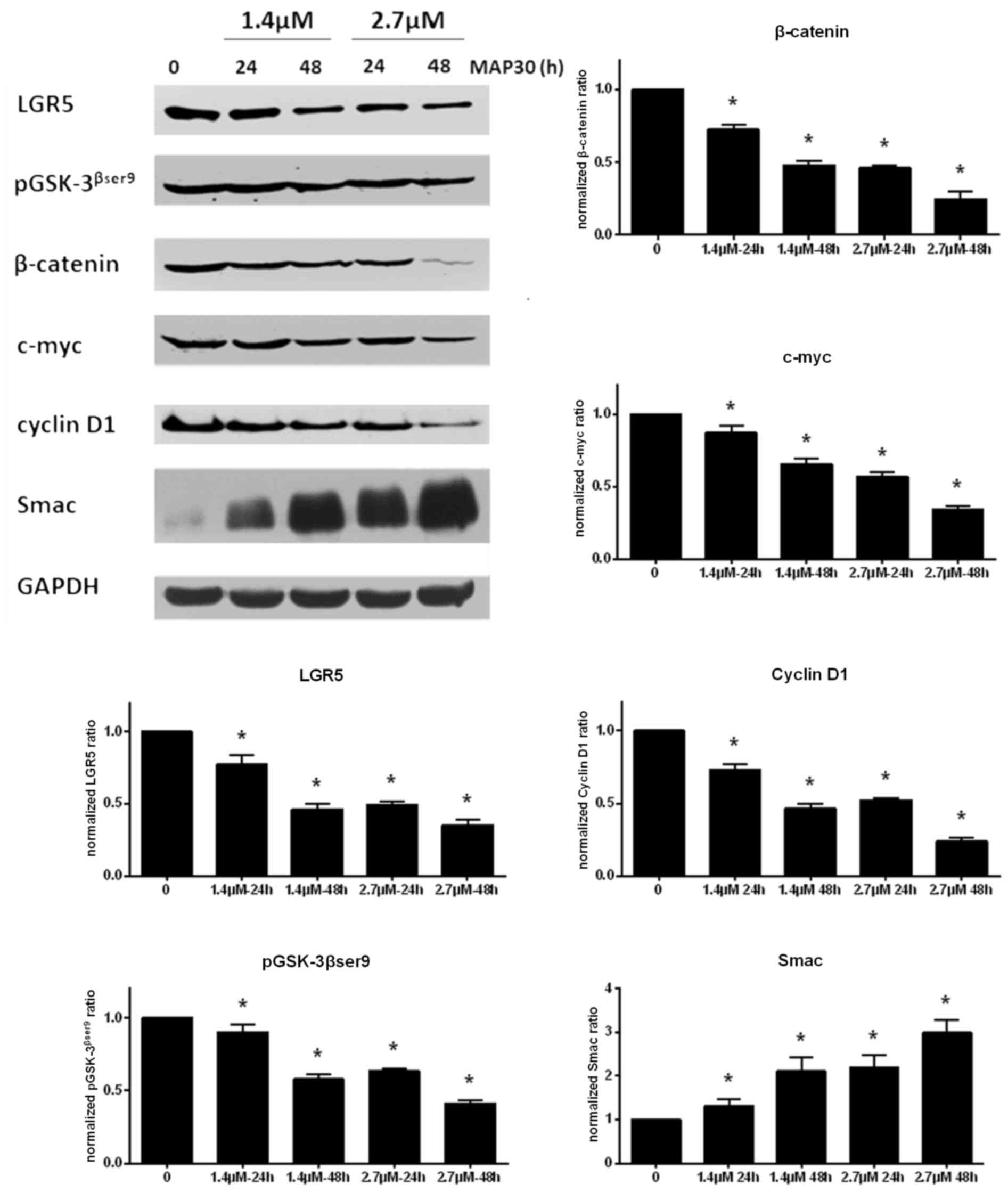
Western blot analysis
The western blotting results demonstrated that the
expression of LGR5, pGSK-3βSer9, β-catenin, c-Myc and
cyclin D1 decreased, and the expression of Smac markedly increased,
as the dose (1.4–2.7 µM) and treated time (24–48 h) of MAP30
increased (Fig. 4; P<0.05).
Discussion
Previously, a promising anti-angiogenic therapy
(bevacizumab) failed to improve overall survival rates in two
independent Phase III trials (AVAglio and RTOG 0825), suggesting
that gliomas are refractory to treatment (19,20).
Therefore, development of novel and effective therapeutic agents is
highly desirable. Although previous studies have identified that
MAP30 possesses antitumor effects in various tumor cells, including
glioblastoma cells, the underlying molecular mechanism of MAP30 on
glioblastoma cells has not yet been elucidated (4,6,12–14,16,21,22).
The results of the present study demonstrated that MAP30 displayed
significant inhibition of proliferation, invasion and migration on
U87 and U251 cell lines in a dose- and time-dependent manner. The
IC50 values for U87 cells were lower compared with those
for U251 cells, indicating that U87 cells might be more sensitive
to MAP30. Therefore, the U87 cell line was selected for additional
experiments. The U87 cell line is the most representative glioma
cell line. Although it was identified that the DNA profile of U87MG
cells was different from that of the original cells in 2016, it is
likely to be a bona fide human glioblastoma cell line of unknown
origin (23). These cells are thought
to exhibit the characteristics of human glioblastoma (24–27),
including proliferation, invasion and migration. The U87 cell line
remains one of the most representative glioma cell lines. In
addition, the U251 cell line was investigated to support the
results from the U87 cells.
The flow cytometric and fluorescence staining
results demonstrated that MAP30 induced apoptosis and arrested cell
cycle in S-phase. The apoptosis-promoting and cell cycle-arresting
abilities of MAP30 were identified in previous studies (14,28–30). MAP30
was considered to cleave adenine-ribose glycosidic bonds by
targeting the conserved host protein synthesis machinery, and
prevent the elongation of DNA/RNA, resulting in DNA replication
arrest in S-phase (3,16,17).
The apoptotic mechanism of MAP30 in glioma cells has
not been clarified. LGR5, a downstream target gene of the Wnt
signaling pathway, serves a key function in tumorigenesis, and the
Wnt signaling pathway is vital in proliferation, differentiation
and morphogenesis of gliomas (31–36). Our
previous studies revealed that LGR5 expression increased with the
malignant degree of human gliomas, and knockdown of LGR5 was
identified to lead to significant inhibition of the proliferation
of glioma cells in vitro and in vivo (37). Additionally, Hsu et al
(38) demonstrated that depletion of
LGR5 induced apoptosis through the loss of mitochondrial membrane
potential, and inhibited the activity of Wnt/β-catenin signaling by
suppressing the expression of c-Myc and cyclin D in CRC cells
(38). The western blotting results
of the present study indicated that the expression of LGR5,
pGSK-3βSer9, β-catenin, c-Myc and cyclin D1 were
markedly decreased in U87 cells following MAP30 treatment in a
dose- and time-dependent manner. Thus, we hypothesize that
suppression of expression of LGR5 and the Wnt/β-catenin signaling
pathway may be part of the mechanism by which MAP30 inhibits
proliferation and induces apoptosis in glioma cells.
On the other hand, our results indicate that the
expression of Smac is significantly enhanced in U87 cells in a
dose- and time-dependent manner. Evasion of programmed cell death
is a hallmark of human cancer (39).
Smac is considered to be an important anticancer factor in the
mitochondrial apoptosis pathway, which binds directly to and
neutralizes inhibitor of apoptosis (IAP) proteins, thus leading to
activation of the intrinsic pathway of apoptosis (39–42). In
addition, Smac mimetic-mediated depletion of IAP proteins may lead
to ubiquitination, proteasomal degradation of substrates and
activation of nuclear factor κB (NF-κB), a transcription factor
with important functions in cell death and survival signaling, and
all the functions of Smac promote apoptosis (39,41,43).
In addition, previous studies fused MAP30 with
human-derived cell penetrating peptide or poly (ethylene glycol) to
increase the uptake efficiency, cytotoxic activity and in
vivo half-life and decrease immunogenicity of MAP30 (12,44). Even
more importantly, RIPs may exhibit little or no detectable adverse
effects on normal cells as they may recognize features on membranes
dominant to tumor cells (4,17,45–47). All
these characteristics would broaden the number of applications of
MAP30.
On the basis of the results of the present study, it
is concluded that the mechanisms of MAP30 against malignant glioma
cell lines may be associated with potent suppression of LGR5 and
the Wnt/β-catenin signaling pathway, and enhancement of Smac
expression in a dose- and time-dependent manner, indicating that
MAP30 may be a novel and effective therapeutic natural plant agent
for the treatment of gliomas. However, further study using in
vivo models and elucidation of the underlying molecular
mechanism of action of Smac are required to confirm these
results.
Acknowledgements
The present study was supported by the Peking
University People's Hospital Research and Development Funds (grant
nos. 2104000004, RDB2011-14 and RDC2015-05) and by the National
Natural Science Foundation of China (grant no. 81001009). The
authors would like to thank Professor Meng (College of Life
Sciences, Sichuan University, Chengdu, China) for providing the
highly purified MAP30. The authors also thank Professor Xiangjun
He, Dr Junjie Miao, Dr Dongliang Wang (Peking University People's
Hospital, Beijing, China) and Dr Jiangfeng Liang (Peking University
International Hospital, Beijing, China) for providing the
laboratory and technical advice.
References
|
1
|
Louis DN, Ohgaki H, Wiestler OD, Cavenee
WK, Burger P, Jouvet A, Scheithauer BW and Kleihues P: WHO
classification of tumours of the central nervous system. Acta
Neuropathol. 114:97–109. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tchoghandjian A, Soubéran A, Tabouret E,
Colin C, Denicolaï E, Jiguet-Jiglaire C, El-Battari A, Villard C,
Baeza-Kallee N and Figarella-Branger D: Inhibitor of apoptosis
protein expression in glioblastomas and their in vitro and in vivo
targeting by SMAC mimetic GDC-0152. Cell Death Dis. 7:e23252016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lee-Huang S and Huang PL, Nara PL, Chen
HC, Kung HF, Huang P, Huang HI and Huang PL: MAP 30: A new
inhibitor of HIV-1 infection and replication. FEBS Lett. 272:12–18.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fang EF, Zhang CZ, Ng TB, Wong JH, Pan WL,
Ye XJ, Chan YS and Fong WP: Momordica Charantia lectin, a type II
ribosome inactivating protein, exhibits antitumor activity toward
human nasopharyngeal carcinoma cells in vitro and in vivo. Cancer
Prev Res (Phila). 5:109–121. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fang EF and Ng TB: Can bitter gourd
(Momordica Charantia) be a novel therapy for human cancers? Cancer
Prev Res. 1–2. 2012.
|
|
6
|
Fang EF and Ng TB: Bitter gourd (Momordica
charantia) is a cornucopia of health: A review of its credited
antidiabetic, anti-HIV and antitumor properties. Curr Mol Med.
11:417–436. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang X, Sun W, Cao J, Qu H, Bi X and Zhao
Y: Structures of new triterpenoids and cytotoxicity activities of
the isolated major compounds from the fruit of Momordica charantia
L. J Agric Food Chem. 60:3927–3933. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Miao J, Jiang Y, Wang D, Zhou J, Fan C,
Jiao F, Liu B, Zhang J, Wang Y and Zhang Q: Trichosanthin
suppresses the proliferation of glioma cells by inhibiting LGR5
expression and the Wnt/β-catenin signaling pathway. Oncol Rep.
34:2845–2852. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Grover JK and Yadav SP: Pharmacological
actions and potential uses of Momordica charantia: A review. J
Ethnopharmacol. 93:123–132. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dandawate PR, Subramaniam D, Padhye SB and
Anant S: Bitter melon: A panacea for inflammation and cancer. Chin
J Nat Med. 14:81–100. 2016.PubMed/NCBI
|
|
11
|
Raina K, Kumar D and Agarwal R: Promise of
bitter melon (Momordica charantia) bioactives in cancer prevention
and therapy. Semin Cancer Biol 40–41. 1–129. 2016.
|
|
12
|
Lv Q, Yang XZ, Fu LY, Lu YT, Lu YH, Zhao J
and Wang FJ: Recombinant expression and purification of a
MAP30-cell penetrating peptide fusion protein with higher
anti-tumor bioactivity. Protein Expr Purif. 111:9–17. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fan X, He L and Meng Y, Li G, Li L and
Meng Y: A-MMC and MAP30, two ribosome-inactivating proteins
extracted from Momordica charantia, induce cell cycle arrest and
apoptosis in A549 human lung carcinoma cells. Mol Med Rep.
11:3553–3558. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fang EF, Zhang CZ, Wong JH, Shen JY, Li CH
and Ng TB: The MAP30 protein from bitter gourd (Momordica
charantia) seeds promotes apoptosis in liver cancer cells in vitro
and in vivo. Cancer Lett. 324:66–74. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fan JM, Luo J, Xu J, Zhu S, Zhang Q, Gao
DF, Xu YB and Zhang GP: Effects of recombinant MAP30 on cell
proliferation and apoptosis of human colorectal carcinoma LoVo
cells. Mol Biotechnol. 39:79–86. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fang EF, Ng TB, Shaw PC and Wong RN:
Recent progress in medicinal investigations on trichosanthin and
other ribosome inactivating proteins from the plant genus
Trichosanthes. Curr Med Chem. 18:4410–4417. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang YX, Neamati N, Jacob J, Palmer I,
Stahl SJ, Kaufman JD, Huang PL, Huang PL, Winslow HE, Pommier Y, et
al: Solution structure of anti-HIV-1 and anti-tumor protein MAP30:
Structural insights into its multiple functions. Cell. 99:433–442.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Meng Y, Lin S, Liu S, Fan X, Li G and Meng
Y: A novel method for simultaneous production of two
ribosome-inactivating proteins, α-MMC and MAP30, from Momordica
charantia L. PLoS One. 9:e1019982014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chinot OL, Wick W, Mason W, Henriksson R,
Saran F, Nishikawa R, Carpentier AF, Hoang-Xuan K, Kavan P, Cernea
D, et al: Bevacizumab plus radiotherapy-temozolomide for newly
diagnosed glioblastoma. N Engl J Med. 370:709–722. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wick W, Chinot OL, Bendszus M, Mason W,
Henriksson R, Saran F, Nishikawa R, Revil C, Kerloeguen Y and
Cloughesy T: Evaluation of pseudoprogression rates and tumor
progression patterns in a phase III trial of bevacizumab plus
radiotherapy/temozolomide for newly diagnosed glioblastoma. Neuro
Oncol. 18:1434–1441. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hlin H, Zhi-Guo Z, Cong-Hui H, Yan Z, Qing
L, Bo J, Hou-Guang H, Jun-Jie Z and Pei-Ying Z: Expression of
Momordica charantia MAP30 and its anti-tumor effect on bladder
cancer cells. Minerva Urol Nefrol. 68:275–281. 2014.PubMed/NCBI
|
|
22
|
Meng Y, Liu S, Li J, Meng Y and Zhao X:
Preparation of an antitumor and antivirus agent: chemical
modification of α-MMC and MAP30 from Momordica Charantia L. with
covalent conjugation of polyethyelene glycol. Int J Nanomedicine.
7:3133–3142. 2012.PubMed/NCBI
|
|
23
|
Allen M, Bjerke M, Edlund H, Nelander S
and Westermark B: Origin of the U87MG glioma cell line: Good news
and bad news. Sci Transl Med. 8:354re32016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bedini A, Baiula M, Vincelli G, Formaggio
F, Lombardi S, Caprini M and Spampinato S: Nociceptin/orphanin FQ
antagonizes lipopolysaccharide-stimulated proliferation, migration
and inflammatory signaling in human glioblastoma U87 cells. Biochem
Pharmacol. 140:89–104. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Heckler M, Osterberg N, Guenzle J,
Thiede-Stan NK, Reichardt W, Weidensteiner C, Saavedra JE and
Weyerbrock A: The nitric oxide donor JS-K sensitizes U87 glioma
cells to repetitive irradiation. Tumour Biol.
39:10104283177039222017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Alphandéry E, Idbaih A, Adam C, Delattre
JY, Schmitt C, Guyot F and Chebbi I: Development of non-pyrogenic
magnetosome minerals coated with poly-l-lysine leading to full
disappearance of intracranial U87-Luc glioblastoma in 100% of
treated mice using magnetic hyperthermia. Biomaterials.
141:210–222. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Alphandéry E, Idbaih A, Adam C, Delattre
JY, Schmitt C, Guyot F and Chebbi I: Chains of magnetosomes with
controlled endotoxin release and partial tumor occupation induce
full destruction of intracranial U87-Luc glioma in mice under the
application of an alternating magnetic field. J Control Release.
262:259–272. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ru P, Steele R, Nerurkar PV, Phillips N
and Ray RB: Bitter melon extract impairs prostate cancer cell-cycle
progression and delays prostatic intraepithelial neoplasia in TRAMP
model. Cancer Prev Res (Phila). 4:2122–2130. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Brennan VC, Wang CM and Yang WH: Bitter
melon (Momordica charantia) extract suppresses adrenocortical
cancer cell proliferation through modulation of the apoptotic
pathway, steroidogenesis and insulin-like growth factor type 1
receptor/RAC-alpha serine/threonine-protein kinase signaling. J Med
Food. 15:325–334. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ray RB, Raychoudhuri A, Steele R and
Nerurkar P: Bitter melon (Momordica charantia) extract inhibits
breast cancer cell proliferation by modulating cell cycle
regulatory genes and promotes apoptosis. Cancer Res. 70:1925–1931.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rot S, Taubert H, Bache M, Greither T,
Würl P, Eckert AW, Schubert J, Vordermark D and Kappler M: A novel
splice variant of the stem cell marker LGR5/GPR49 is correlated
with the risk of tumor-related death in soft-tissue sarcoma
patients. BMC Cancer. 11:4292011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yi GZ, Liu YW, Xiang W, Wang H, Chen ZY,
Xie SD and Qi ST: Akt and β-catenin contribute to TMZ resistance
and EMT of MGMT negative malignant glioma cell line. J Neurol Sci.
367:101–106. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Morris SL and Huang S: Crosstalk of the
Wnt/β-catenin pathway with other pathways in cancer cells. Genes
Dis. 3:41–47. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Barker N and Clevers H: Leucine-rich
repeat-containing G-protein-coupled receptors as markers of adult
stem cells. Gastroenterology. 138:1681–1696. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Barker N, van Es JH, Kuipers J, Kujala P,
van den Born M, Cozijnsen M, Haegebarth A, Korving J, Begthel H,
Peters PJ and Clevers H: Identification of stem cells in small
intestine and colon by marker gene Lgr5. Nature. 449:1003–1007.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hsu SY, Kudo M, Chen T, Nakabayashi K,
Bhalla A, van der Spek PJ, van Duin M and Hsueh AJ: The three
subfamilies of leucine-rich repeat-containing G protein-coupled
receptors (LGR): Identification of LGR6 and LGR7 and the signaling
mechanism for LGR7. Mol Endocrinol. 14:1257–1271. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang D, Zhou J, Fan C, Jiao F, Liu B, Sun
P, Miao J and Zhang Q: Knockdown of LGR5 suppresses the
proliferation of glioma cells in vitro and in vivo. Oncol Rep.
31:41–49. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hsu HC, Liu YS, Tseng KC, Tan BC, Chen SJ
and Chen HC: LGR5 regulates survival through mitochondria-mediated
apoptosis and by targeting the Wnt/β-catenin signaling pathway in
colorectal cancer cells. Cell Signal. 26:2333–2342. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Fulda S: Promises and challenges of smac
mimetics as cancer therapeutics. Clin Cancer Res. 21:5030–5036.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Marschall V and Fulda S: Smac
mimetic-induced upregulation of interferon-β sensitizes
glioblastoma to temozolomide-induced cell death. Cell Death Dis.
6:e18882015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Mizukawa K, Kawamura A, Sasayama T, Tanaka
K, Kamei M, Sasaki M and Kohmura E: Synthetic Smac peptide enhances
the effect of etoposide-induced apoptosis in human glioblastoma
cell lines. J Neurooncol. 77:247–255. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Amendola D, De Salvo M, Marchese R, Verga
Falzacappa C, Stigliano A, Carico E, Brunetti E, Moscarini M and
Bucci B: Myc down-regulation affects cyclin D1/cdk4 activity and
induces apoptosis via Smac/Diablo pathway in an astrocytoma cell
line. Cell Prolif. 42:94–109. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Scheidereit C: IkappaB kinase complexes:
Gateways to NF-kappaB activation and transcription. Oncogene.
25:6685–6705. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sun Y, Sun F, Li J, Wu M, Fan X and Meng Y
and Meng Y: Mono-PEGylation of Alpha-MMC and MAP30 from Momordica
charantia L: Production, identification and anti-tumor activity.
Molecules. 21:E14572016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Huang PL, Sun Y, Chen HC, Kung HF and
Lee-Huang S: Proteolytic fragments of anti-HIV and anti-tumor
proteins MAP30 and GAP31 are biologically active. Biochem Biophys
Res Commun. 262:615–623. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Schreiber CA, Wan L, Sun Y, Lu L, Krey LC
and Lee-Huang S: The antiviral agents, MAP30 and GAP31, are not
toxic to human spermatozoa and may be useful in preventing the
sexual transmission of human immunodeficiency virus type 1. Fertil
Steril. 72:686–690. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Putnam CD and Tainer JA: The food of sweet
and bitter fancy. Nat Struct Biol. 7:17–18. 2000. View Article : Google Scholar : PubMed/NCBI
|