Introduction
Colorectal cancer (CRC) is the third most commonly
diagnosed malignancy in males and the second in females, accounting
for ~0.6 million mortalities annually (1,2). The
development and progression of colorectal cancer is associated with
multiple factors including lifestyle, environmental factors, and
genetic and epigenetic changes (3,4). Despite
investigations into the etiology of CRC, the underlying molecular
mechanisms of the disease remain incompletely understood.
Stromal interaction molecule 1 (STIM1) has
previously been identified as a novel human gene, located in the
imprinted gene domain of 11p15.5 which is also reported to be an
important tumor suppressing gene region (5). Ca2+ is an almost ubiquitous
intracellular messenger regulating a variety of physiological and
pathological processes (6).
Store-operated Ca2+ channels provide the major
Ca2+ entry pathway in non-excitable cells, particularly
in cancer cells. It has been demonstrated that STIM1 is an
endoplasmic reticulum (ER) Ca2+ sensor and controls
store-operated Ca2+ entry (SOCE) with ORAI1 (7). Once Ca2+ is released from the
ER, STIMs are able to aggregate and translocate to the ER-plasma
membrane to activate Orai1 (8). STIM1
has also been implicated in the proliferation, migration, invasion
and metastasis in a number of types of cancer, including cervical
cancer, non-small lung cancer, breast cancer and melanoma (9–12).
Furthermore, STIM1 is a potential molecular marker for the
diagnosis and progression of cancer (13,14).
However, there remain conflicting results about the functions of
STIM1 in different cancer studies. Upregulation of
STIM1-ORAI1-mediated SOCE appears to promote metastasis by
stimulating the migration of breast tumor cells (15). Downregulation of STIM1-ORAI1-mediated
SOCE was demonstrated to contribute to the increased resistance to
apoptosis in prostate cancer cells (16). The molecular mechanisms underlying the
regulation of signaling pathways by STIM1 in cancer require further
investigation. Previous studies have demonstrated that STIM1 is
associated with tumor metastasis in CRC (17,18).
However, the importance of the role served by STIM1 in cell growth
and proliferation of CRC remains unclear.
In the present study, small hairpin RNA (shRNA)
silencing of STIM1 in CRC cell lines was performed and it was
revealed that STIM1 knockdown was able to inhibit proliferation and
induce apoptosis of CRC cells. Several pro-apoptotic markers were
also studied to reveal the molecular mechanism underlying STIM1
silencing-promoted cell apoptosis in CRC. The results of the
present study suggest that STIM1 may be essential for the
proliferation and tumorigenesis of CRC.
Materials and methods
Cell lines and culture
Human CRC cell lines SW1116 and HCT116, and 293T
cells were obtained from the Cell Bank of the Chinese Academy of
Sciences (Shanghai, China). HCT116 cells were cultured in RPMI-1640
medium (GE Healthcare, Chicago, IL, USA) with 10% fetal bovine
serum (FBS; Biowest, MO, USA). SW1116 cells and 293T cells were
cultured in Dulbecco's modified Eagle's medium (GE Healthcare)
supplemented with 10% FBS at 37°C in a humidified incubator
containing 5% CO2.
Lentiviral packaging and cell
infection
An shRNA targeting human STIM1 (shSTIM1;
NM_003156.3) was designed using online shRNA tools (Invitrogen;
Thermo Fisher Scientific, Inc., Waltham, MA, USA): shRNA
(5′-CTAGCGCCTGGATGATGTAGATCATAATTCAAGAGATTATGATCTACATCATCCAGGTTTTTT-3′).
The non-silencing shRNA (shCon) sequence
(5′-TTCTCCGAACGTGTCACGTCTCGAGACGTGACACGTTCGGAGAATTTTT-3′) was used
as a negative control. All oligonucleotides were synthesized and
cloned into NheI/PacI-linearized pFH-L vector
(Shanghai Hollybio Co., Ltd., Shanghai, China), which contains
green fluorescent protein (GFP) as a reporter marker. The
lentiviruses were subsequently generated by co-transfecting 293T
cells with the shSTIM1 plasmid and pVSVG-I and pCMV-dR8.92 helper
(Shanghai Hollybio Co., Ltd.). Lentiviruses were purified using
ultracentrifugation with centrifugal force of 80,000 × g at 4°C for
60 min. To determine the viral titer, 293T cells were transfected
with lentiviruses and following this, green fluorescent cells under
fluorescence microscopy were observed and the viral titer was
estimated. For cell infection, HCT116 (1×105 cells/well)
and SW1116 cells (4×104 cells/well) were plated in
6-well plates and then infected with lentiviruses (shCon and
shSTIM1) at a multiplicity of infection of 60 and 50, respectively,
for 96 h. GFP-positive signals were observed using fluorescence
microscopy to evaluate infection efficiency.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. RNA (1 µg) was reverse-transcribed into
cDNA using a Takara Reverse Transcription system (Takara
Biotechnology Co., Ltd., Dalian, China). RT-qPCR was conducted on a
BioRad Connect RT-PCR platform using CFX Manager software and the
Precision Melt Analysis system (version 1.0; Bio-Rad Laboratories,
Inc., Hercules, CA, USA) using a volume of 20 µl (10 µl 2X SYBR
Premix Ex Taq, 0.5 µl forward and reverse primers, 5 µl cDNA and
4.5 µl double-distilled water) under the following conditions:
Initial denaturation at 95°C for 1 min and 40 cycles of
denaturation at 95°C for 5 sec and annealing and extension at 60°C
for 20 sec. The relative expression levels of STIM1 mRNA were
calculated using the 2−ΔΔCq method (19), and β-actin was used as the reference
gene. The primers used were as follows: STIM1,
5′-AGCCTCAGCCATAGTCACAG-3′ (forward) and
5′-TTCCACATCCACATCACCATTG-3′ (reverse); B-cell lymphoma 2
(Bcl-2)-associated death promoter (Bad), 5′-GGCACAGCAACGCAGATG-3′
(forward) and 5′-CGGGTGGAGTTTCGGGATG-3′ (reverse); Bcl-2-associated
X protein (Bax), 5′-TGGGCTGGACATTGGACTTC-3′ (forward) and
5′-AGCACTCCCGCCACAAAG-3′ (reverse); and β-actin,
5′-GTGGACATCCGCAAAGAC-3′ (forward) and 5′-AAAGGGTGTAACGCAACTA-3′
(reverse).
Western blot analysis
Total protein was extracted using 2X SDS Lysis
Buffer (100 mM Tris, 4% SDS, 10% glycerol, 200 mM NaCl, 2 mM EDTA,
pH=6.8) from SW1116 and HCT116 cells, and then 30 g protein was
separated by SDS-PAGE with 10% acrylamide and transferred onto
polyvinylidene fluoride membranes. The membranes were probed with
specific antibodies against STIM1 (cat. no. sc-166840; 1:1,000 for
HCT116 cells and 1:500 for SW1116 cells; Santa Cruz Biotechnology,
Inc., Dallas, TX, USA), poly(ADP-ribose) polymerase (PARP; cat. no.
9542; 1:1,000; Cell Signaling Technology Europe, B.V., Leiden, The
Netherlands) or GAPDH (cat. no. 10494-1-AP; 1:50,000 for HCT116
cells and 1:10,000 for SW1116 cells; Proteintech Group Inc.,
Chicago, IL, USA) at 4°C overnight. Membranes were subsequently
incubated with horseradish peroxidase-conjugated goat anti-mouse
(cat. no. sc-2005; 1:5,000; Santa Cruz Biotechnology, Inc.) or goat
anti-rabbit (cat. no. sc-2054, 1:5,000; Santa Cruz Biotechnology,
Inc.) immunoglobulin G antibody and visualized using the Pierce
enhanced chemiluminescence kit (Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol.
MTT assay
Cell proliferation was determined using an MTT assay
(Beyotime Institute of Biotechnology, Haimen, China), according to
the manufacturer's protocol. Briefly, following lentivirus
infection for 120 h, HCT116 cells and SW1116 cells were seeded into
96-well plates at a density of 2×103 or
2.5×103 cells/well, respectively. At the indicated time
points (1, 2, 3, 4 and 5 days), MTT solution (5 mg/ml) was added to
each well prior to incubation at 37°C in a humidified chamber for 4
h. Acidic isopropanol (10% SDS, 5% isopropanol and 0.01 M HCl) was
added to stop the reaction. After 10 min of incubation, the
absorbance was measured using a microplate reader at a wavelength
of 595 nm.
Colony formation
Following lentivirus infection for 120 h, SW1116
cells in the exponential growth phase were plated into 6-well
plates at 500 cells/well. Following continuous culture for 10 days,
the RPMI-1640 medium was then discarded. The cells were washed with
PBS (pH 7) twice, fixed with methyl alcohol for 20 min, stained
with crystal violet for 20 min, washed with distilled water and
dried. Colonies were counted by eye.
Cell apoptosis analysis
Apoptosis was evaluated using the Annexin
V/7-aminoactinomycin-D (7-AAD) double staining assay (eBioscience;
Thermo Fisher Scientific, Inc.). Briefly, SW1116 cells were
reseeded into a 6-well plate at 2×105 cells/well 5 days
after lentiviral infection. When cell confluence reached 80%, cells
were collected, washed twice with PBS at 4°C and resuspended in
binding buffer. Finally, cells were stained with Annexin V and
7-AAD solution, and the apoptotic cells were analyzed using flow
cytometry.
Statistical analysis
Data analyses were performed using GraphPad Prism
software (GraphPad Software 5.0, Inc., La Jolla, CA, USA). All
results are expressed as the mean ± standard deviation from three
independent replicate experiments. P<0.05 was considered to
indicate a statistically significant difference, using one-way
analysis of variance followed by Student's t-test.
Results
Lentivirus-mediated shRNA efficiently
suppresses STIM1 expression in CRC cells
To study the biological role of STIM1 in CRC, the
expression of STIM1 was specifically knocked down in CRC cell
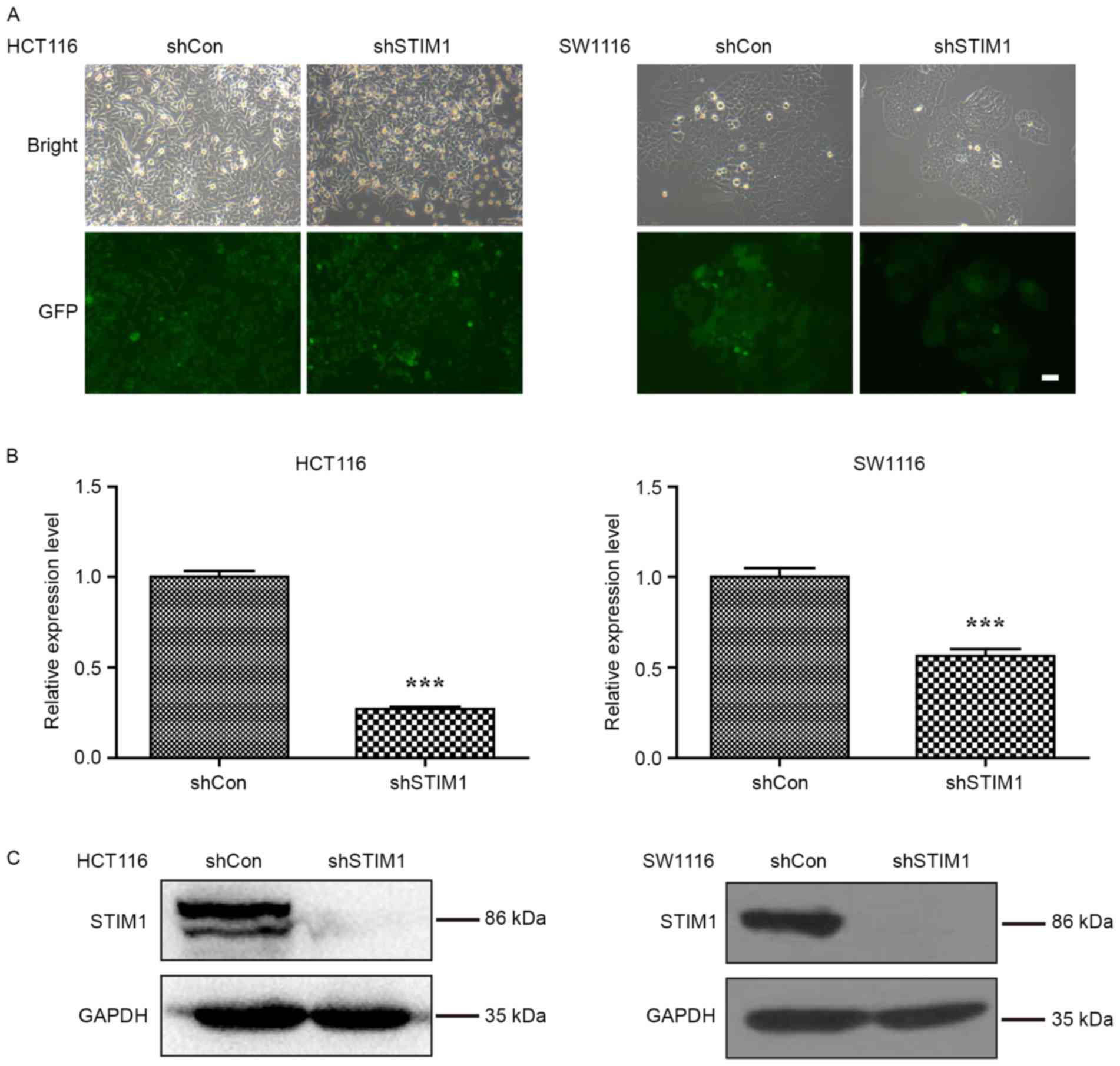
lines, HCT116 and SW1116. As presented in Fig. 1A, the majority of HCT116 and SW1116
cells expressed GFP-positive signals, suggesting satisfactory
infection efficiency. Furthermore, the knockdown efficiency of
STIM1 was investigated in these two cell lines using RT-qPCR and
western blot analysis. As presented in Fig. 1B, the mRNA levels of STIM1 were
significantly decreased in HCT116 and SW1116 cells following STIM1
knockdown (P<0.001 when compared with shCon). Consistent with
qPCR results, the protein levels of STIM1 were decreased in the
shSTIM1 group compared with the shCon group in HCT116 and SW1116
cells, respectively (Fig. 1C). These
results suggest that the lentivirus-mediated shRNA was able to
significantly suppress the endogenous STIM1 expression in HCT116
and SW1116 cells.
Knockdown of STIM1 significantly
inhibits CRC cell proliferation and colony formation ability
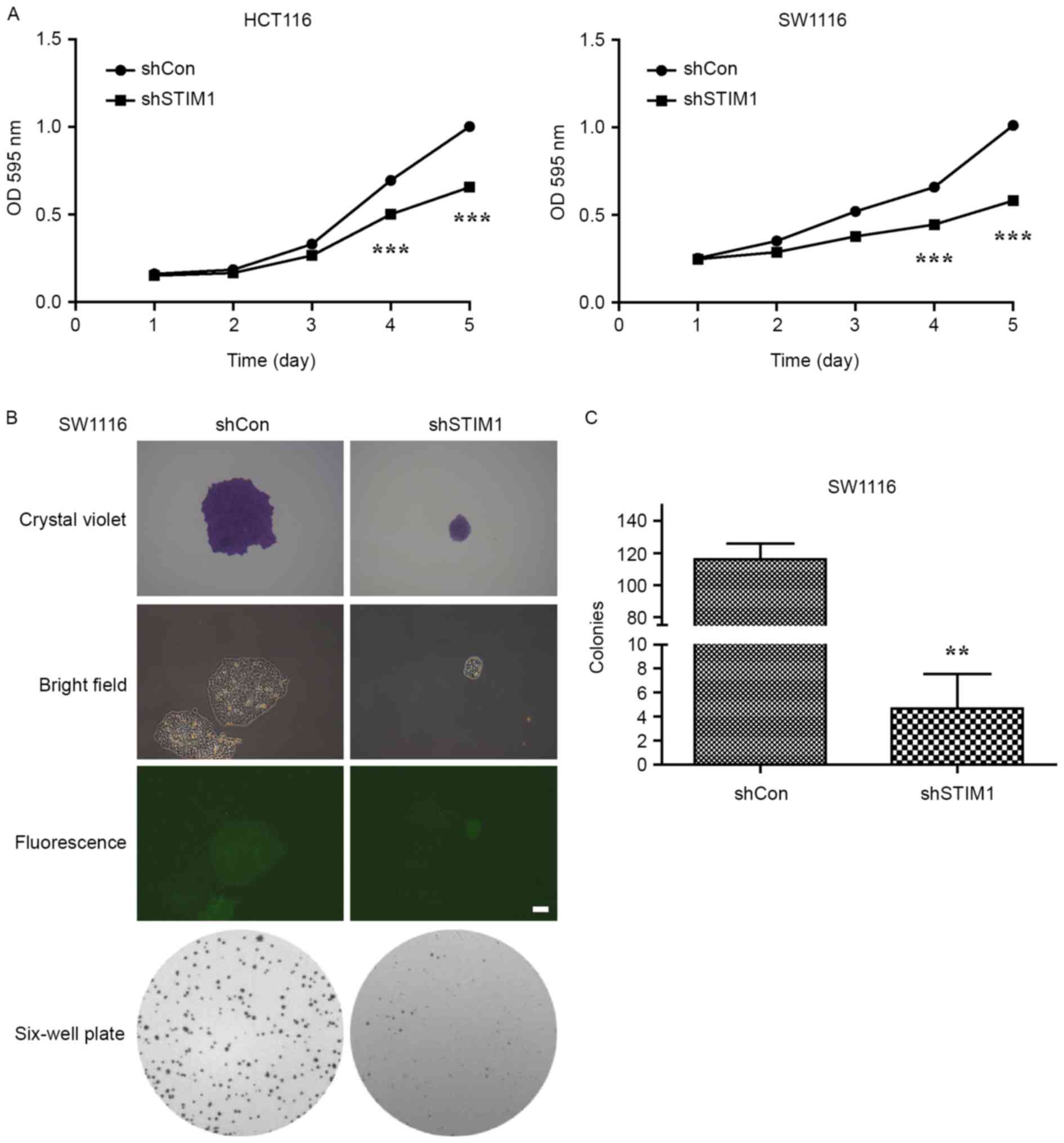
Proliferation of shCon- or shSTIM1-transfected
HCT116 and SW1116 cells was measured using an MTT assay daily for 5
days. The result demonstrated that STIM1-specific shRNA caused a
significant decrease in the proliferation of HCT116 and SW1116
cells (P<0.001 vs. shCon; Fig.
2A). Notably, suppression of cell proliferation following
knockdown of STIM1 was more evident in SW1116 cells than in HCT116
cells. As a result, SW1116 cells were selected to perform a colony
formation assay. As presented in Fig. 2B
and C, STIM1-depleted cells grew slowly and formed a few
smaller colonies when compared with the control group during a
10-day period, which indicated that knockdown of STIM1
significantly impaired the colony formation ability of SW1116 cells
(P<0.01). These results indicate that STIM1 silencing may
inhibit the proliferation and colony formation ability of CRC cells
in vitro.
STIM1 silencing promotes the apoptosis
of CRC cells
Genes that are capable of inducing apoptosis are
generally considered potential anticancer targets. To confirm
whether STIM1 is a suitable potential molecular target for CRC, the
Annexin V/7-AAD double staining apoptosis detection assay was used
to investigate cell apoptosis in SW1116 cells following STIM1
knockdown. As presented in Fig. 3A,
the proportion of cells in early apoptotic stage (Annexin
V+/7-AAD−) was increased from 2.70 to 18.00%
and those in the late apoptotic stage (Annexin
V+/7-AAD+) were also increased from 17.36 to
39.72%. Statistical analysis indicated that knockdown of STIM1
significantly promotes cell early- and late-stage apoptosis in
SW1116 cells (P<0.001; Fig. 3B)
which provides evidence that STIM1 silencing may induce apoptosis
in SW1116 cells.
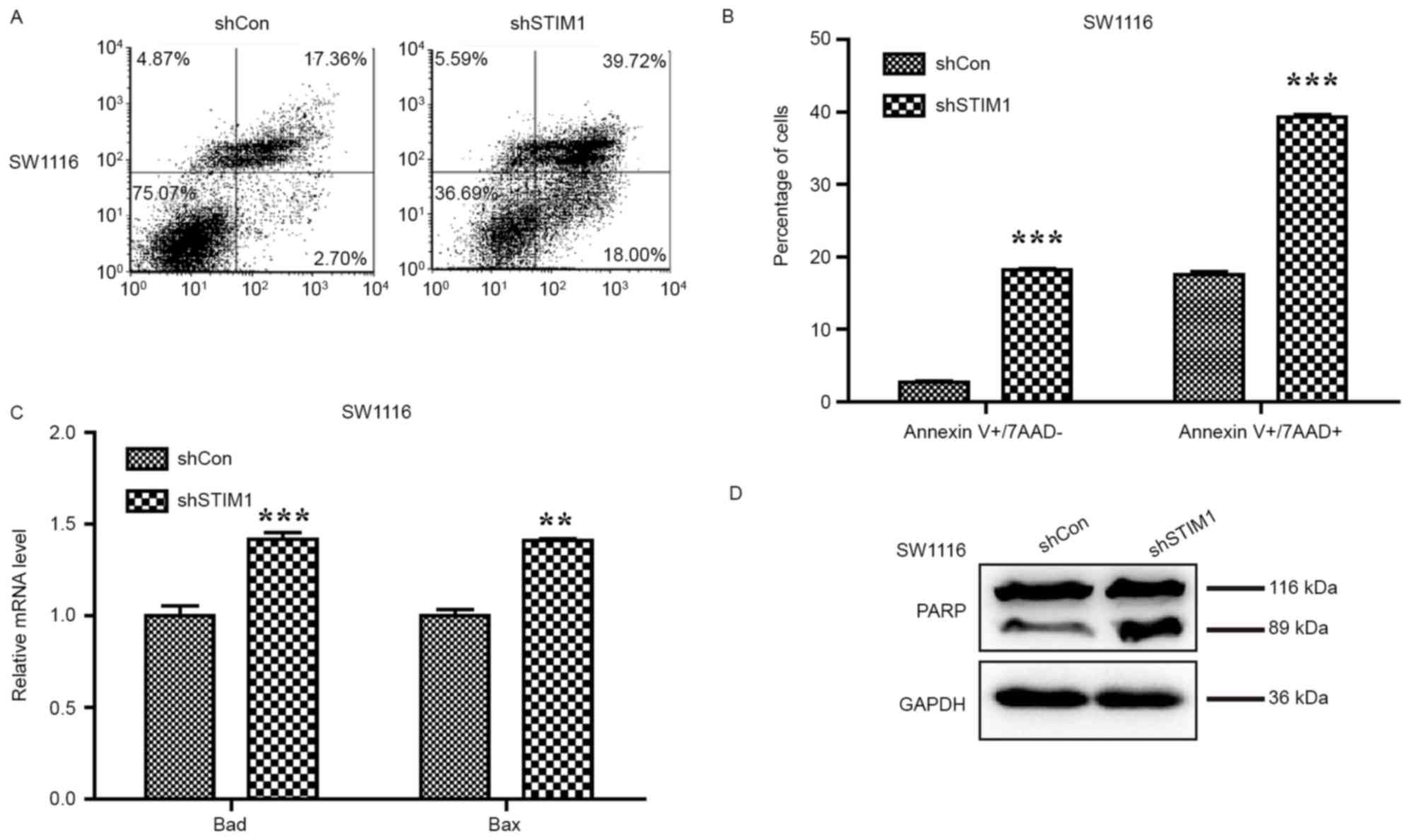 | Figure 3.Knockdown of STIM1 induces cell
apoptosis via activation of the mitochondrial signalling pathway.
(A) Representative images of cell apoptosis of SW1116 cells
following shSTIM1 infection analyzed using flow cytometry and
Annexin V/7-AAD double staining. (B) Quantification of the
proportions of SW1116 cells corresponding to early (Annexin
V+/7-AAD-) and late (Annexin V+/7-AAD+) apoptotic cells.
Upregulation of (C) Bad and Bax, and (D) cleaved PARP measured by
western blot analysis. The experiment was performed in triplicate
and repeated in triplicate. **P<0.01, ***P<0.001 vs. shCon.
STIM1, stromal interaction molecule 1; shSTIM1, shRNA targeting
human STIM1; shCon, non-silencing shRNA; CRC, colorectal cancer;
PARP, poly(ADP-ribose) polymerase; BAD, B-cell lymphoma-2
associated death promoter; Bax, B-cell lymphoma-2 associated X
protein; 7-AAD, 7-aminoactinomycin D. |
To further uncover the molecular mechanism
underlying cell apoptosis promoted by STIM1 silencing, the levels
of Bad and Bax expression, and PARP cleavage were examined to
detect apoptosis following silencing of STIM1 in SW1116 cells. The
qPCR analysis indicated that expression levels of Bad and Bax were
increased in SW1116 cells following STIM1 silencing (P<0.01 and
P<0.001; Fig. 3C). Furthermore, an
increase in PARP cleavage was observed in SW1116 cells with
silenced STIM1, indicating a higher proportion of apoptotic cells
as a consequence of STIM1 silencing (Fig.
3D).
Discussion
Despite the ongoing advancement of our understanding
of cancer progression and metastasis in the last decade, a number
of oncogenes remain poorly understood. Previous studies indicate
that Ca2+ channels are critical regulators in cancer
(20,21). As a Ca2+ sensor, STIM1 is
required for a number of important physiological and pathological
processes. Previous studies indicate that STIM1 may be a potential
therapeutic target in numerous types of cancer (10,11,17). In
breast cancer, overexpression of STIM1 is able to promote
metastasis and invasion of cancer cells (15). In lung cancer, STIM1 is upregulated in
the carcinoma relative to the para-carcinoma tissues (22). Additionally, STIM1 overexpression
promotes CRC cell migration by inducing cyclo-oxygenase 2
expression and is associated with CRC progression (17); however, the role of STIM1 in CRC cell
proliferation has not been fully elucidated.
Results of the present study suggest that knockdown
of STIM1 inhibits the proliferation and colony formation ability of
CRC cells, which is consistent with results from a previous study
(23). Furthermore, Annexin V/7-AAD
double staining confirmed that knockdown of STIM1 increased the
proportion of early and late apoptotic cells. Cleavage of PARP is
one of the most commonly used diagnostic tools for the detection of
apoptosis in numerous cell types (24). In the present study, the expression of
PARP cleavage was increased in STIM1-silenced cells. Furthermore,
knockdown of STIM1 increased the expression of the pro-apoptotic
mitochondrial genes Bax and Bad in SW1116 cells. Therefore, it may
be inferred that the cell proliferation inhibition by STIM1
silencing in CRC was possibly due to the induction of
mitochondria-associated apoptosis.
In conclusion, the results of the present study
demonstrate that STIM1 knockdown by lentivirus-mediated shRNA may
inhibit the proliferation of CRC cells, which may be a result of
induced apoptosis. These results also for the first time highlight
the crucial role of STIM1 in CRC progression. Further study is
required to illuminate the potential value of STIM1-targeted
therapy in vivo.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Natural
Science Foundation of Ningbo (grant nos. 2013A610224 and
2016A610135), the Municipal Key Disciplines of Ningbo (grant no.
2013-88), the Medical Foundation of Ningbo (grant no. 2011B10), the
Social Development and Scientific and Technological Projects
Foundation of Ningbo (grant no. 2014C50068) and the Huamei
Foundation of Ningbo No. 2 Hospital (grant nos. 2015HMKJ08 and
2016HMKJ26).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
DY performed the vector construction experiments and
drafted the manuscript. XD and KL participated in the research
design, reviewed the literature and examined the data. YX and JZ
participated in the qPCR and western blot experiments. MD and HY
participated in the cellular function experiments. ZK participated
in the data analysis and figure format.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Desantis C and Jemal A:
Colorectal cancer statistics, 2014. CA Cancer J Clin. 64:104–117.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fearon ER: Molecular genetics of
colorectal cancer. Ann N Y Acad Sci. 768:101–110. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Song M, Garrett WS and Chan AT: Nutrients,
foods, and colorectal cancer prevention. Gastroenterology.
148:1244–1260. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Parker NJ, Begley CG, Smith PJ and Fox RM:
Molecular cloning of a novel human gene (D11S4896E) at chromosomal
region 11p15.5. Genomics. 37:253–256. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Prevarskaya N, Skryma R and Shuba Y:
Calcium in tumour metastasis: New roles for known actors. Nature
Rev Cancer. 11:609–618. 2011. View
Article : Google Scholar
|
|
7
|
Roos J, DiGregorio PJ, Yeromin AV, Ohlsen
K, Lioudyno M, Zhang S, Safrina O, Kozak JA, Wagner SL, Cahalan MD,
et al: STIM1, an essential and conserved component of
store-operated Ca2+ channel function. J Cell Biol. 169:435–445.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hogan PG, Lewis RS and Rao A: Molecular
basis of calcium signaling in lymphocytes: STIM and ORAI. Annu Rev
Immunol. 28:491–533. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hu J, Qin K, Zhang Y, Gong J, Li N, Lv D,
Xiang R and Tan X: Downregulation of transcription factor Oct4
induces an epithelial-to-mesenchymal transition via enhancement of
Ca2+ influx in breast cancer cells. Biochem Biophys Res Commun.
411:786–791. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hou MF, Kuo HC, Li JH, Wang YS, Chang CC,
Chen KC, Chen WC, Chiu CC, Yang S and Chang WC: Orai1/CRACM1
overexpression suppresses cell proliferation via attenuation of the
store-operated calcium influx-mediated signalling pathway in A549
lung cancer cells. Biochim Biophys Acta. 1810:1278–1284. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen YF, Chiu WT, Chen YT, Lin PY, Huang
HJ, Chou CY, Chang HC, Tang MJ and Shen MR: Calcium store sensor
stromal-interaction molecule 1-dependent signaling plays an
important role in cervical cancer growth, migration, and
angiogenesis. Proc Natl Acad Sci USA. 108:15225–15230. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fedida-Metula S, Feldman B, Koshelev V,
Levin-Gromiko U, Voronov E and Fishman D: Lipid rafts couple
store-operated Ca2+ entry to constitutive activation of PKB/Akt in
a Ca2+/calmodulin-, Src- and PP2A-mediated pathway and promote
melanoma tumor growth. Carcinogenesis. 33:740–750. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
van de Vijver MJ, He YD, van't Veer LJ,
Dai H, Hart AA, Voskuil DW, Schreiber GJ, Peterse JL, Roberts C,
Marton MJ, et al: A gene-expression signature as a predictor of
survival in breast cancer. N Engl J Med. 347:1999–2009. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Faehling M, Kroll J, Föhr KJ, Fellbrich G,
Mayr U, Trischler G and Waltenberger J: Essential role of calcium
in vascular endothelial growth factor A-induced signaling:
Mechanism of the antiangiogenic effect of carboxyamidotriazole.
FASEB J. 16:1805–1807. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang S, Zhang JJ and Huang XY: Orai1 and
STIM1 are critical for breast tumor cell migration and metastasis.
Cancer cell. 15:124–134. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Flourakis M, Lehen'kyi V, Beck B, Raphaël
M, Vandenberghe M, Abeele FV, Roudbaraki M, Lepage G, Mauroy B,
Romanin C, et al: Orai1 contributes to the establishment of an
apoptosis-resistant phenotype in prostate cancer cells. Cell Death
Dis. 1:e752010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang JY, Sun J, Huang MY, Wang YS, Hou MF,
Sun Y, He H, Krishna N, Chiu SJ, Lin S, et al: STIM1 overexpression
promotes colorectal cancer progression, cell motility and COX-2
expression. Oncogene. 34:4358–4367. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang Z, Liu X, Feng B, Liu N, Wu Q, Han
Y, Nie Y, Wu K, Shi Y and Fan D: STIM1, a direct target of
microRNA-185, promotes tumor metastasis and is associated with poor
prognosis in colorectal cancer. Oncogene. 35:60432016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Monteith GR, Davis FM and Roberts-Thomson
SJ: Calcium channels and pumps in cancer: Changes and consequences.
J Biol Chem. 287:31666–31673. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen YF, Chen YT, Chiu WT and Shen MR:
Remodeling of calcium signaling in tumor progression. J Biomed Sci.
20:232013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li W, Zhang M, Xu L, Lin D, Cai S and Zou
F: The apoptosis of non-small cell lung cancer induced by cisplatin
through modulation of STIM1. Exp Toxicol Pathol. 65:1073–1081.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li G, Zhang Z, Wang R, Ma W, Yang Y, Wei J
and Wei Y: Suppression of STIM1 inhibits human glioblastoma cell
proliferation and induces G0/G1 phase arrest. J Exp Clin Cancer
Res. 32:202013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bressenot A, Marchal S, Bezdetnaya L,
Garrier J, Guillemin F and Plénat F: Assessment of apoptosis by
immunohistochemistry to active caspase-3, active caspase-7, or
cleaved PARP in monolayer cells and spheroid and subcutaneous
xenografts of human carcinoma. J Histochem Cytochem. 57:289–300.
2009. View Article : Google Scholar : PubMed/NCBI
|