Introduction
Prostate cancer (PCa) is the second most commonly
diagnosed form of cancer and the sixth leading cause of
cancer-associated mortality among males globally (1). It has been reported that the 5-year
survival rate is ~100% for patients with localized cancer but is
only 31% for those with distant metastases (2). Therefore, it is of great importance to
identify an effective treatment method for PCa therapy.
Nicotinamide N-methyltransferase (NNMT) was
identified as an S-adenosyl-L-methionine-dependent cytoplasmic
enzyme (3). Previous studies have
indicated its critical role in the biotransformation and
detoxification of multiple drugs and xenobiotic compounds (3,4). Abnormal
upregulation of NNMT has been extensively identified in various
tumor types. For instance, in the progression of PCa,
overexpression of NNMT has been frequently determined (4). Furthermore, NNMT has been frequently
reported to be a non-invasive biomarker of cancer in body fluids,
including serum (5), saliva (6) and urine (7). It was originally defined as the enzyme
responsible for nicotinamide methylation, which is an important
form of vitamin B3 (8). A current
study has demonstrated that the role of NNMT is far from the
clearance of excess vitamin B3 (9).
It has demonstrated that the global epigenetic histone profiles are
regulated by NNMT, and it mainly exerts the acetylation
modifications through regulating the expression of sirtuin 1
(SIRT1) (9). Furthermore, enhanced
NNMT expression in the liver was reported to stabilize SIRT1
protein, thereby maintaining hepatic nutrient metabolism (9); however, whether NNMT may upregulate
SIRT1 in PCa has yet to be elucidated.
Elevated SIRT1 expression has been extensively
identified in multiple types of human malignancy, including
ovarian, liver, stomach, ductal and pancreatic cancer (10,11). In
addition, various molecules are indicated to be regulated by SIRT1,
including p53, forkhead box O1-4, nuclear factor-κB and E2F1
(12–14); however, the regulatory control of
SIRT1 remains poorly understood in PCa. In the present study, it
was demonstrated that increased NNMT expression elevated the
expression level of SIRT1, thereby prompting the malignant
progression of PCa.
Materials and methods
Tissue samples
Patient-matched PCa and adjacent non-cancerous
prostate tissues (15 pairs) were obtained from patients (mean age,
68.7±13.7 years; age range 45–83 years) who had undergone radical
prostatectomy at the Department of Oncology, 202 Hospital of
Chinese People's Liberation Army (Shenyang, China) between January
2015 to Feb 2016. The adjacent non-cancerous prostate tissues from
the patients with PCa were used as controls. PCa tissue specimens
(n=15) were identified as prostatic adenocarcinoma. None of the
cases had received any previous cancer-associated treatment or had
a history of any other type of cancer. Tumors were staged according
to the 2010 revised TNM system (15).
The whole blood samples (5 ml) from the
aforementioned patients with PCa and healthy donors (69.2±14.3)
were collected in tubes containing EDTA between January 2015 and
February 2016 at the Department of Oncology, 202 Hospital of
Chinese People's Liberation Army. Total RNA was extracted using
RNAzol LS (Vigorous Biotechnology Beijing Co., Ltd., Beijing,
China, http://www.vigorousbiol.com/),
according to the manufacturer's protocols. The concentration and
purity of the RNA samples were determined by OD260/OD280.
The clinicopathological characteristics are
presented in Table I. All the samples
were snap-frozen in liquid nitrogen immediately and were stored at
−80°C following surgery until RNA extraction. The study protocols
were approved by the Local Ethics Committees of 202 Hospital of the
Chinese People's Liberation Army and written informed consent was
obtained from all patients and healthy donors prior to tissue or
blood collection.
 | Table I.Clinicopathological characteristics of
patients with PCa and healthy controls. |
Table I.
Clinicopathological characteristics of
patients with PCa and healthy controls.
| Characteristic | Healthy control | PCa | P-value |
|---|
| Age, years
(range) | 69.2 (46–85) | 68.7 (48–86) |
|
| PSA±SD, ng/ml | 2.4±1.6 | 124.3±250.6 | <0.001 |
| Gleason score, no.
(%) |
|
|
|
| ≤7 |
| 7 (46.7) |
|
| ≥8 |
| 8 (53.3) |
|
| Stage, no. (%) |
|
|
|
|
T1-4N0M0 |
|
|
|
|
T1-4N0M0 |
| 9 (60.0) |
|
|
Metastasis (any T, N>0,
M>0) |
| 6 (40.0) |
|
Cell culture
The PCa PC-3 cell line was purchased from the
Chinese Academy of Sciences Cell Bank and cells were cultured in
RPMI-1640/F12 medium (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA), supplemented with 10% fetal bovine serum (FBS;
Invitrogen; Thermo Fisher Scientific, Inc.), streptomycin (100
mg/ml) and penicillin (100 U/ml) at 37°C in a humidified atmosphere
containing 5% CO2.
Construction of adeno-associated virus
(AAV) vectors
AAV-NNMT or AAV-NC was constructed by Shanghai
GeneChem Co., Ltd. (Shanghai, China). The PCa PC-3 cells were
seeded at the density of 106 cells/well. At 70%
confluency, AAV-NNMT or AAV-NC was transfected without transfection
reagent into PC-3 cells at 30 multiplicity of infection (MOI) for
48 h. Following 48 h transfection, the cells were collected for
subsequent experimentation.
Small interfering RNA (siRNA)
transfection
siRNA was synthesized based on the human NNMT target
sequence (3′-GCTCAAGAGCAGCTACTACAT-5′; Shanghai Genchem Co., Ltd.,
Shanghai, China). In brief, PC-3 cells were seeded onto a 6-well
plate and were transfected with 20 µM siRNA using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocols.
Following transfection for 24 h, cells were collected for further
experiments.
MTT assay
To investigate the effect of NNMT on PC-3 cell
viability, PC-3 cells were seeded in 96-well tissue culture plates
at a density of 5×104 cells per well in DMEM medium.
When the confluence reached 70%, AAV-NNMT was transfected each well
at 37°C for 24, 48 and 72 h. When the confluence reached 70%, siRNA
targeting NNMT was transfected into each well for 24, 48 and 72 h,
respectively. Cell viability was examined with MTT assay kits
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). The blue formazan
products in the cells were dissolved in 150 µl dimethyl sulfoxide
(Sigma-Aldrich; Merck KGaA) and spectrophotometrically measured at
a wavelength of 550 nm. All experiments were performed in
triplicate.
Scratch assay
PC-3 cells were seeded onto a 6-well plate at a
density of 5×105 cells/well until 90% confluence. Next,
a scratch was produced in each well using 10-µl pipette tips.
Following AAV-NNMT or AAV-NC transfection for 48 h at 37°C, the
cells were washed with phosphate-buffered saline (PBS) three times
and the width of the scratch was measured under an inverted
microscope (IX83, Olympus Corporation, Tokyo Japan) (×10
magnification) at different time points following transfection. All
the experiments were repeated three times.
Transwell assay
Migration and invasion assays were performed using
transwell chambers with membrane pore size of 8.0 µm (Corning
Incorporated, Corning, NY, USA). Membranes were uncoated for the
migration assays and coated with 25 µg Matrigel (BD Biosciences,
Franklin Lakes, NJ, USA) for the invasion assays. They were
incubated with PBS (migration) or Matrigel during 1 h at 37°C, 5%
CO2 atmosphere. PC-3 cells at log-phase were prepared
and fasted for 12 h in serum-free RPMI-1640/F12 as aforementioned.
The cells were collected and rinsed with PBS three times. Next,
cells were re-suspended at 6×105 cells/ml. Additionally,
0.1 ml cell suspension was added into 24-well Transwell chambers,
with the lower chamber containing 0.5 ml RPMI-1640/F12 medium,
supplemented with FBS. Following 24 h of incubation, cells were
stained with 0.1% crystal violet for 10 min at room temperature and
were rinsed with PBS, followed by rinsing with 33% acetic acid.
Subsequently, cells were visualized under an inverted microscope
(magnification, ×200) to examine the microporous membrane lower
cell. Subsequently, 10 randomly selected fields were used to
calculate the average.
Western blot analysis
Total protein was extracted from PC-3 cells or PCa
tissues or adjacent non-cancerous prostate tissues using
radioimmunoprecipitation assay buffer (Beijing Solarbio Science
& Technology Co., Ltd., Beijing, China). A BCA protein assay
kit (Pierce; Thermo Fisher Scientific, Inc.) was used to determine
the protein concentration. A total of 20 µg protein in each lane
was separated by 12% SDS-PAGE, prior to being transferred onto a
polyvinylidene fluoride membrane. Following blocking with 5%
skimmed milk powder for 2 h at room temperature, monoclonal
antibodies against NNMT (cat. no., ab58743), SIRT1 (cat. no.,
ab32441) and GAPDH (cat. no., ab9485) (all dilution, 1:1,000;
Abcam, Cambridge, MA, USA) were applied for overnight incubation at
4°C. Non-specific binding was blocked using 8% (w/v) milk in
Tris-buffered saline with 1% Tween-20 (TBST; Beijing SolarBio
Science & Technology Co., Ltd.) for 2 h at room temperature.
Following several washes with TBST, the membranes were incubated
with horseradish-peroxidase (HRP)-conjugated goat anti-rabbit or
anti-mouse IgG, or HRP-conjugated mouse anti-goat IgG (all
dilution, 1:5,000; OriGene Technologies, Inc., Beijing, China) for
2 h at room temperature, prior to being washed with TBST. GAPDH was
used as the internal control. Signals were detected using enhanced
chemiluminescence, according to the manufacturer's protocols (EMD
Millipore, Billerica, MA, USA). ImageJ 1.8.0 software (National
Institutes of Health, Bethesda, MD, USA) was used for
densitometry.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from PC-3 cells or PCa
tissues or adjacent non-cancerous prostate tissues using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) and was used as the template to synthesize cDNA using a
reverse transcription kit (Takara Bio, Inc., Otsu, Japan) according
to the instructions. Briefly, 1 µl random primer, 1 µl dNTP mixture
and 1 µl RNA were mixed at 65°C for 5 min. Then, 2 µl 5X
PrimeScript™ Buffer, 1 µl PrimeScript RTase, and 4 µl
DEPC H2O were added at 30°C for 10 min, 42°C for 20 min, and 95°C
for 4 min. Then, the cDNA were collected. RT-qPCR was performed
using a fluorescent qPCR kit (Qiagen GmbH, Hilden, Germany) and
specific primers. Subsequently, PCR amplification was performed. A
total of 1 µg cDNA was used for qPCR using the SYBR®
green Master mix (Roche Diagnostics, Basel, Switzerland) and a
Roche Lightcycler 480 (Roche Diagnostics) at 95°C for 10 min
followed by 50 cycles of: 95°C for 10 sec; 55°C for 10 sec; 72°C
for 5 sec; 99°C for 1 sec; 59°C for 15 sec; 95°C for 1 sec; and
cooling to 40°C. All experiments were performed in triplicate
against a GAPDH housekeeping gene. Relative expression was
normalized against the endogenous control, GAPDH, using the
2−∆∆Cq method (16). The
primers for NNMT, SIRT1, and GAPDH were listed as follows:
NNMT-forward (f), 5′-CTGCCTAGACGGTGTGAAGG-3′; NNMT-reverse (r),
5′-CTTGACCGCCTGTCTCAACT-3′; SIRT1-f, 5′-CCTGCCTGGATCCCCTTAGT-3′;
SIRT1-r, 5′-GGCCTGTTGCTCTCCTCATT-3′; GAPDH-f, GAGAAGGCTGGGGCTCATTT;
GAPDH-r, AGTGATGGCATGGACTGTGG.
Nicotinamide treatment
10 µg/µl Nicotinamide (72340, Sigma-Aldrich; Merck
KGaA) was dissolved in ddH2O. To evaluate the effects of
nicotinamide on the expression of SIRT1, nicotinamide was added in
the RPMI-1640/F12 culture at the final concentration of 20 ng/µl in
the presence or absence of AAV-NNMT.
Statistical analysis
SPSS 19.0 software (IBM Corp., Armonk, NY, USA) was
used to process all of the collected data, and those that fit a
normal distribution are presented as the mean ± standard deviation.
Multiple-group comparisons were performed using analysis of
variance, followed by the least significant difference post hoc
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
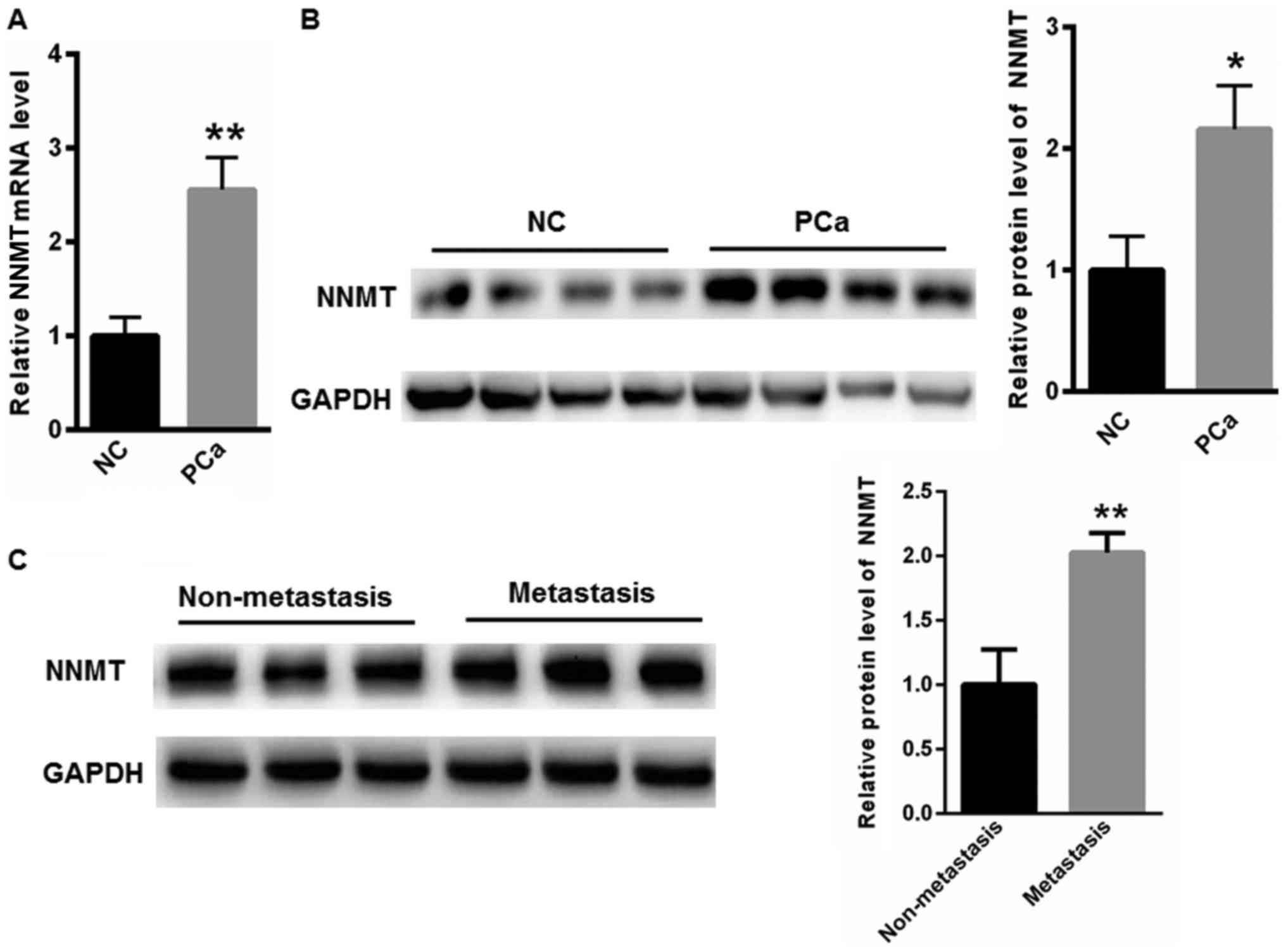
Elevated levels of NNMT in the
peripheral blood and tissues of patients with PCa
In the present study, the NNMT mRNA level in the
peripheral blood of healthy donors was used as a negative control
(NC) when comparing with the NNMT mRNA level of NNMT in the
peripheral blood of patients with PCa. In order to compare the
expression of NNMT in prostate tissues, the PCa tissues and
adjacent non-cancerous prostate tissues were collected from
patients with PCa. To begin with, the expression of NNMT in the
peripheral blood and tissues of patients with PCa was evaluated. As
depicted in Fig. 1A, the mRNA level
of NNMT was notably elevated in the peripheral blood of patients
with PCa, compared with that in the NC samples. The expression of
NNMT in the tissues of patients with PCa was also evaluated and the
data demonstrated that NNMT expression was notably increased,
compared with that in the NC adjacent non-cancerous prostate
tissues (Fig. 1B). Furthermore, the
protein expression level of NNMT was determined to be increased in
the tissues of patients with metastases, compared with that of the
patients without metastases (Fig.
1C).
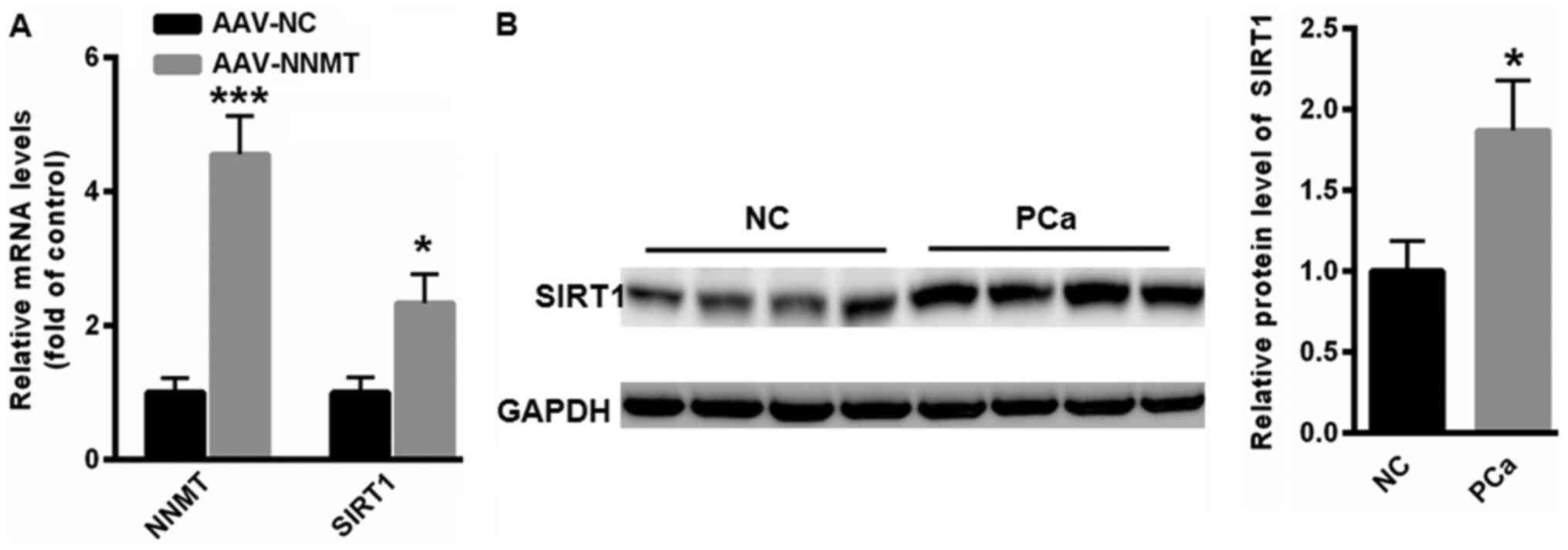
NNMT enhances the expression of SIRT1
in PC-3 cells
Next, the possible association between NNMT and SIRT
expression was evaluated. Overexpression of NNMT significantly
increased the mRNA expression of SIRT1 in PC-3 cells (P<0.05;
Fig. 2A). Furthermore, western blot
analysis indicated that the protein expression of SIRT1 was
elevated in the tissues of patients with PCa, compared with that in
the tissues of NC subjects (Fig.
2B).
NNMT promotes PC-3 cell viability,
migration and colony formation
Following this, the effect of NNMT on the malignant
phenotype of PC-3 cells was investigated. An MTT assay demonstrated
that the overexpression of NNMT increased the cell viability by
34.5, 78.6 and 123.2% at 24 (P<0.05), 48 (P<0.05) and 72 h
(P<0.01), respectively (Fig. 3A).
By contrast, the knockdown of NNMT notably suppressed PC-3 cell
viability by 12.3, 22.1 and 34.5% at 24, 48 (P<0.05) and 72 h
(P<0.05), respectively (Fig. 3B).
Furthermore, the upregulation of NNMT elevated the cell migration
and colony formation capacity (Fig. 3C
and D).
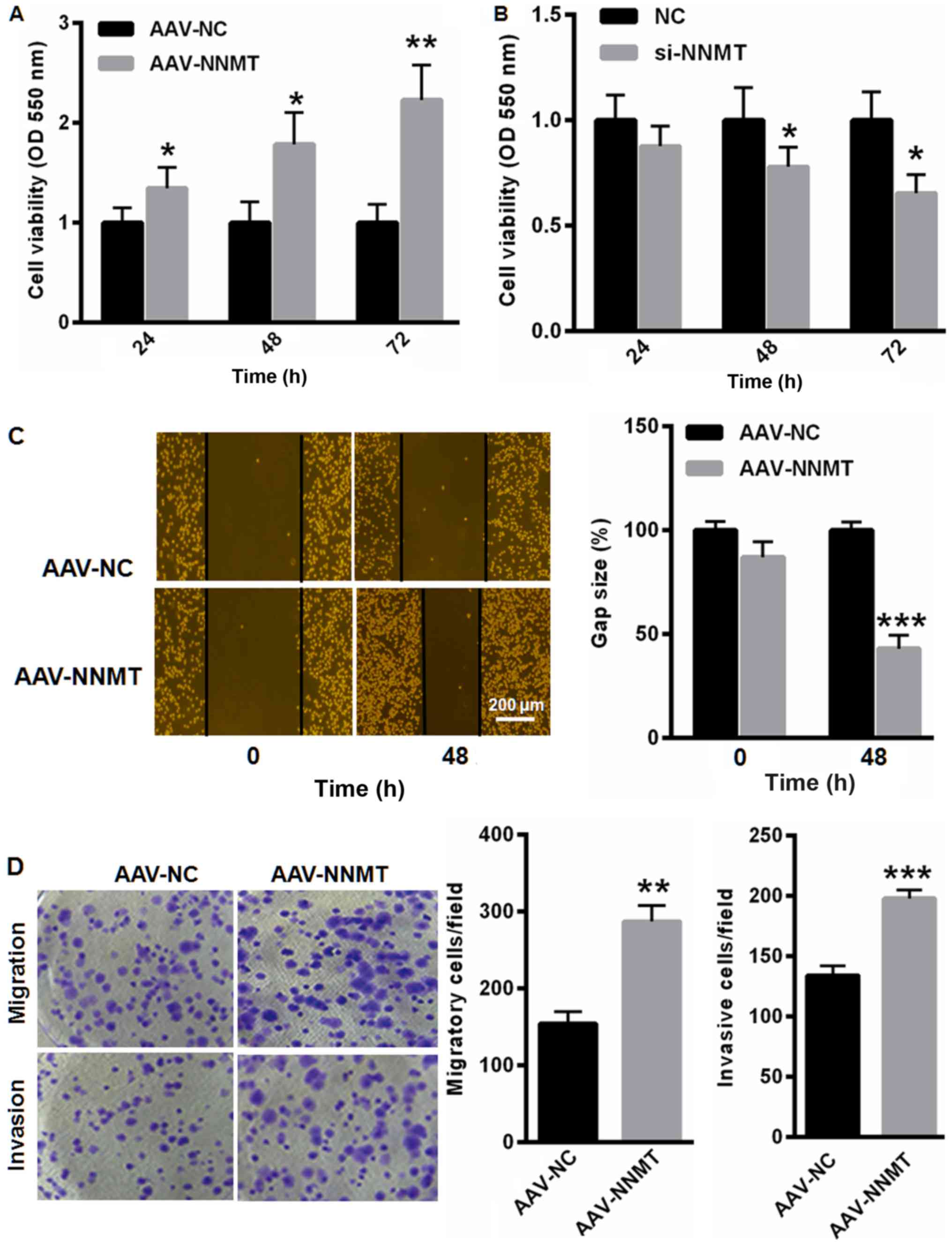 | Figure 3.NNMT promotes PC-3 cell viability,
migration and colony formation. (A) Overexpression of NNMT elevated
the viability of PC-3 cells by 34.5, 78.6 and 123.2% at 24, 48 and
72 h, respectively. (B) Knockdown of NNMT markedly suppressed PC-3
cell viability by 12.3, 22.1 and 34.5% at 24, 48 and 72 h,
respectively. Upregulation of NNMT elevated the cell migration and
invasion capacity as analyzed by the (C) scratch assay, and (D)
invasion and migration assays. *P<0.05, **P<0.01,
***P<0.001 vs. control. NNMT, nicotinamide N-methyltransferase;
NC, negative control; AAV, adeno-associated virus; si, small
interfering; OD, optical density. |
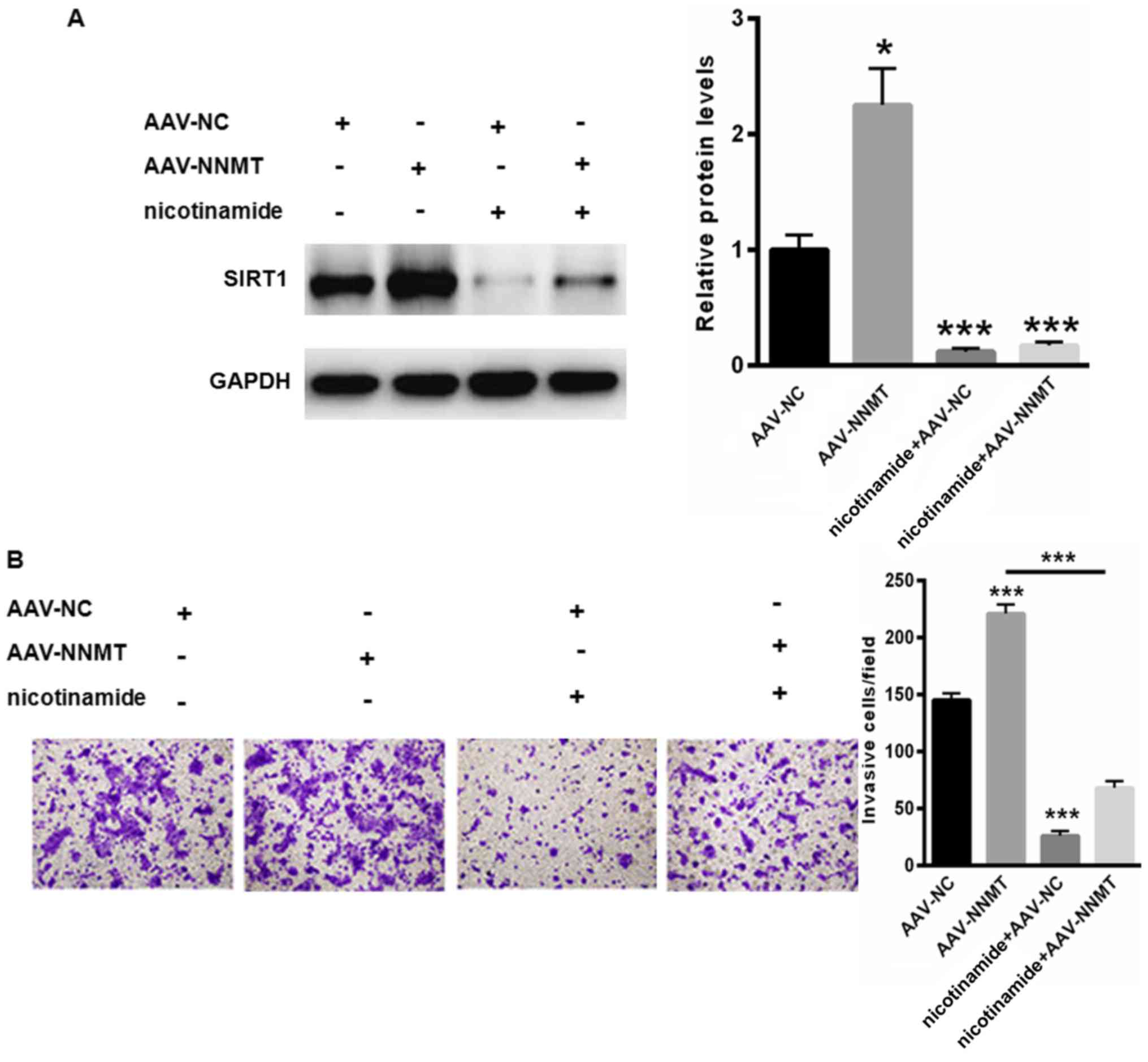
NNMT exerts its oncogenic role
primarily by upregulating SIRT1
Additionally, the underlying mechanism through which
NNMT promotes the malignancy of PCa was investigated. Therefore, an
SIRT1 inhibitor, nicotinamide, was selected. As depicted in
Fig. 4A, treatment with nicotinamide
notably suppressed the expression of SIRT1 even in cells
transfected with adeno-associated virus (AAV)-NNMT (Fig. 4A). At the same time, treatment with
nicotinamide suppressed PC-3 cell migration (Fig. 4B). By contrast, the upregulation of
NNMT notably abolished the nicotinamide-reduced cell migration
capacity (Fig. 4B). These data
indicated that NNMT enhanced PC-3 cell malignancy primarily through
the upregulation of NNMT.
Discussion
PCa is the most common neoplasia among males
globally (17). Therefore, it is of
great importance to investigate the molecular mechanisms underlying
PCa development and progression. In the present study, the primary
focus was on NNMT, a phase II metabolizing enzyme that primarily
catalyzes the methylation of nicotinamide and other pyridines
(9). The data indicated that the
level of NNMT was elevated in the peripheral blood and tissues of
patients with PCa, indicating a potential oncogenic role of NNMT in
the progression of PCa.
Previous studies have been focused on the functional
roles of NNMT in metabolic diseases, including diabetes and obesity
(9,18). Other studies reported its involvement
in the development and progression of carcinoma in vivo
(19,20). Multiple subsequent data have indicated
that the upregulation of NNMT in various cancer types is associated
with a poor prognosis (19,21). Furthermore, NNMT was also demonstrated
to enhance the proliferative, migratory, invasive and
differentiation capacities of different types of tumor (20,22). For
instance, the overexpression of NNMT promoted renal carcinoma cell
proliferation and invasion mainly by activating the
phosphoinositide 3-kinase/Akt pathway in nasopharyngeal carcinoma
(21). In the present study, NNMT
expression was demonstrated to be increased in PCa, but the
biological function and underlying mechanisms of this remain
unclear.
The in vitro experiments of the present study
revealed that the overexpression of NNMT significantly enhanced
PC-3 cell viability, cell migration and invasive capacity. These
oncogenic results prompted additional investigation into the
underlying mechanism of NNMT in the development of PCa. Recently,
NNMT was demonstrated to stabilize the mRNA and protein expression
levels of SIRT1, thereby maintaining lipid homeostasis in the liver
(9); however, whether NNMT functions
through SIRT1 in PCa has yet to be investigated. Previous studies
have indicated that SIRT1 serves an important role in multiple
physiological processes, including aging, metabolism, neurogenesis
and cell survival, based on its capacity to deacetylate histone and
non-histone substrates (23,24). For instance, SIRT1 was indicated to
increase the expression of matrix metalloproteinase-2, thereby
enhancing PCa cell invasion (25). In
order to validate the role of SIRT1 in PCa malignancies in the
present study, a specific inhibitor, nicotinamide, was selected. It
was determined that the PC-3 cell invasion capacity was notably
decreased by nicotinamide treatment, even in PC-3 cells transfected
with AAV-NNMT. These data indicated that NNMT enhanced PC-3 cell
migration and invasion primarily through regulating SIRT1
expression.
In summary, the present study indicated that NNMT is
an important regulator of SIRT1 expression in PC-3 cells and may be
a potential therapeutic target for PCa.
Acknowledgements
Not applicable.
Funding
This work is supported by the Cardiovascular System
Disease Transformation Medical Research Center and Collaborative
Research Network Construction, 2013225089.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZY performed the experiments and analyzed the data.
YL performed the western blotting. XL conceived the idea,
supervised the study and wrote the manuscript.
Ethics approval and consent to
participate
The study protocols were approved by the Local
Ethics Committees of 202 Hospital of the Chinese People's
Liberation Army and written informed consent was obtained from all
patients and healthy donors prior to tissue or blood
collection.
Consent for publication
The patients and healthy control provided written
informed consent for the publication of any associated data and
accompanying images.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wang X, Yang B and Ma B: The
UCA1/miR-204/Sirt1 axis modulates docetaxel sensitivity of prostate
cancer cells. Cancer Chemother Pharmacol. 78:1025–1031. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Byles V, Zhu L, Lovaas JD, Chmilewski LK,
Wang J, Faller DV and Dai Y: SIRT1 induces EMT by cooperating with
EMT transcription factors and enhances prostate cancer cell
migration and metastasis. Oncogene. 31:4619–4629. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xu Y, Liu P, Zheng DH, Wu N, Zhu L, Xing C
and Zhu J: Expression profile and prognostic value of NNMT in
patients with pancreatic cancer. Oncotarget. 7:19975–19981.
2016.PubMed/NCBI
|
|
4
|
Zhou W, Gui M, Zhu M, Long Z, Huang L,
Zhou J, He L and Zhong K: Nicotinamide N-methyltransferase is
overexpressed in prostate cancer and correlates with prolonged
progression-free and overall survival times. Oncol Lett.
8:1175–1180. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chlopicki S, Kurdziel M, Sternak M,
Szafarz M, Szymura-Oleksiak J, Kamiński K and Żołądź JA: Single
bout of endurance exercise increases NNMT activity in the liver and
MNA concentration in plasma; the role of IL-6. Pharmacol Rep.
64:369–376. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sartini D, Pozzi V, Renzi E, Morganti S,
Rocchetti R, Rubini C, Santarelli A, Lo Muzio L and Emanuelli M:
Analysis of tissue and salivary nicotinamide N-methyltransferase in
oral squamous cell carcinoma: Basis for the development of a
noninvasive diagnostic test for early-stage disease. Biol Chem.
393:505–511. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sartini D, Muzzonigro G, Milanese G, Pozzi
V, Vici A, Morganti S, Rossi V, Mazzucchelli R, Montironi R and
Emanuelli M: Upregulation of tissue and urinary nicotinamide
N-methyltransferase in bladder cancer: Potential for the
development of a urine-based diagnostic test. Cell Biochem Biophys.
65:473–483. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pissios P: Nicotinamide
N-Methyltransferase: More than a vitamin B3 clearance enzyme.
Trends Endocrinol Metab. 28:340–353. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hong S, Moreno-Navarrete JM, Wei X,
Kikukawa Y, Tzameli I, Prasad D, Lee Y, Asara JM, Fernandez-Real
JM, Maratos-Flier E and Pissios P: Nicotinamide N-methyltransferase
regulates hepatic nutrient metabolism through Sirt1 protein
stabilization. Nat Med. 21:887–894. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cui Y, Li J, Zheng F, Ouyang Y, Chen X,
Zhang L, Chen Y, Wang L, Mu S and Zhang H: Effect of SIRT1 gene on
Epithelial-mesenchymal transition of human prostate cancer PC-3
cells. Med Sci Monit. 22:380–386. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Duan K, Ge YC, Zhang XP, Wu SY, Feng JS,
Chen SL, Zhang LI, Yuan ZH and Fu CH: miR-34a inhibits cell
proliferation in prostate cancer by downregulation of SIRT1
expression. Oncol Lett. 10:3223–3227. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hoffmann MJ, Engers R, Florl AR, Otte AP,
Muller M and Schulz WA: Expression changes in EZH2, but not in
BMI-1, SIRT1, DNMT1 or DNMT3B are associated with DNA methylation
changes in prostate cancer. Cancer Biol Ther. 6:1403–1412. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jung-Hynes B and Ahmad N: Role of p53 in
the anti-proliferative effects of Sirt1 inhibition in prostate
cancer cells. Cell Cycle. 8:1478–1483. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nakane K, Fujita Y, Terazawa R, Atsumi Y,
Kato T, Nozawa Y, Deguchi T and Ito M: Inhibition of cortactin and
SIRT1 expression attenuates migration and invasion of prostate
cancer DU145 cells. Int J Urol. 19:71–79. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cheng L, Montironi R, Bostwick DG,
Lopez-Beltran A and Berney DM: Staging of prostate cancer.
Histopathology. 60:87–117. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang BD, Ceniccola K, Hwang S, Andrawis R,
Horvath A, Freedman JA, Olender J, Knapp S, Ching T, Garmire L, et
al: Alternative splicing promotes tumour aggressiveness and drug
resistance in African American prostate cancer. Nat Commun.
8:159212017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Trammell SA and Brenner C: NNMT: A bad
actor in fat makes good in liver. Cell Metab. 22:200–201. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Palanichamy K, Kanji S, Gordon N,
Thirumoorthy K, Jacob JR, Litzenberg KT, Patel D and Chakravarti A:
NNMT silencing activates tumor suppressor PP2A, inactivates
oncogenic STKs, and inhibits tumor forming ability. Clin Cancer
Res. 23:2325–2334. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xie X, Liu H, Wang Y, Zhou Y, Yu H, Li G,
Ruan Z, Li F, Wang X and Zhang J: Nicotinamide N-methyltransferase
enhances resistance to 5-fluorouracil in colorectal cancer cells
through inhibition of the ASK1-p38 MAPK pathway. Oncotarget.
7:45837–45848. 2016.PubMed/NCBI
|
|
21
|
Win KT, Lee SW, Huang HY, Lin LC, Lin CY,
Hsing CH, Chen LT and Li CF: Nicotinamide N-methyltransferase
overexpression is associated with Akt phosphorylation and indicates
worse prognosis in patients with nasopharyngeal carcinoma. Tumour
Biol. 34:3923–3931. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yu T, Wang YT, Chen P, Li YH, Chen YX,
Zeng H, Yu AM, Huang M and Bi HC: Effects of nicotinamide
N-methyltransferase on PANC-1 cells proliferation, metastatic
potential and survival under metabolic stress. Cell Physiol
Biochem. 35:710–721. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu T, Liu PY and Marshall GM: The
critical role of the class III histone deacetylase SIRT1 in cancer.
Cancer Res. 69:1702–1705. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yamamoto H, Schoonjans K and Auwerx J:
Sirtuin functions in health and disease. Mol Endocrinol.
21:1745–1755. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lovaas JD, Zhu L, Chiao CY, Byles V,
Faller DV and Dai Y: SIRT1 enhances matrix metalloproteinase-2
expression and tumor cell invasion in prostate cancer cells.
Prostate. 73:522–530. 2013. View Article : Google Scholar : PubMed/NCBI
|