Introduction
Each year, >1 million new cases of colorectal
cancer (CRC) are diagnosed worldwide. CRC is the third most common
malignancy and the fourth most common cause of cancer-associated
mortality (1). Despite novel
biological insights and advances in therapy, the prognosis of
patients with CRC remains poor. The molecular mechanisms underlying
malignant tumor occurrence and progression was investigated, with
the aim of developing more effective therapies against CRC.
MicroRNAs (miRNAs) are a group of endogenous small
noncoding RNAs that are 21–25 nucleotides in size. The primary
function of miRNA is to suppress gene expression, either by
inhibiting mRNA translation or inducing mRNA degradation (2). It is estimated that miRNAs regulate the
expression of >60% of human protein-encoding genes (3), and are involved in modulating multiple
cellular pathways, including cell proliferation, differentiation
and apoptosis (4). miRNAs can serve
as tumor suppressors or oncogenes, depending on their target mRNAs,
and their expression can in turn be regulated by tumor suppressors
(5) and oncogenes (6,7). Aberrant
expression of certain miRNAs has been observed in an array of human
cancer types, and miRNAs are thought to serve important roles in
tumorigenesis. Recently, a number of miRNAs have been identified as
oncogenes or tumor suppressor genes in CRC (8–12).
miR-106a is expressed in cancer cells and in stool
samples of patients with CRC with varying degrees (13–15).
However, the molecular role that miR-106a serves in CRC remains
debatable. Thus, in the present study, the biological consequence
of overexpressing miR-106a in two distinct CRC cell lines was
investigated.
Materials and methods
Cell lines and cultures
Human CRC cell lines HCT116 and SW620 were obtained
from the Type Culture Collection of the Chinese Academy of Sciences
(Shanghai, China). Cells were maintained in RPMI-1640 with 10%
fetal bovine serum (both from Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) and 1% antibiotics (100 U/ml penicillin and
100 µg/ml streptomycin sulfate) in a humidified atmosphere
containing 5% CO2 at 37°C.
miRNA mimics and transfections
miR-106a mimics and nontarget control were purchased
from Shanghai GenePharma Co., Ltd. (Shanghai, China). The sequences
of these synthetic miRNAs were as follows: miR-106a mimic forward,
5′-AAAAGUGCUUACAGUGCAGGUAG-3′ and reverse,
5′-ACCUGCACUGUAAGCACUUUUUU-3′; nontarget control forward,
5′-UUCUCCGAACGUGUCACGUTT-3′ and reverse,
5′-ACGUGACACGUUCGGAGAATT-3′. Cells were transfected with the miRNA
(100 nM/l) using Lipofectamine™ 2000 reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) following
manufacturer's protocol.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA from cells was extracted using TRIzol
(Invitrogen; Thermo Fisher Scientific, Inc.) according to
manufacturer's protocol. First-strand cDNA synthesis was performed
using the TaqMan MicroRNA Reverse Transcription kit and miR-106a RT
primers (Applied Biosystems; Thermo Fisher Scientific, Inc.). The
temperature protocol of first-strand cDNA synthesis was: 16°C for
30 min, 42°C for 30 min and 85°C for 5 min. qPCR was performed
using a miR-106a TaqMan MicroRNA Assay and TaqMan Universal Master
Mix II on an ABI PRISM 7500 (Applied Biosystems; Thermo Fisher
Scientific, Inc.). miR-106a primer sequence:
GGAAAAGTGCTTACAGTGCAGGTAG. Thermocycler conditions used as follow:
95°C for 5 min, followed by 40 cycles at 95°C for 30 sec, 55°C for
40 sec and 72°C for 30 sec. The expression of miR-106a was
normalized to that of U6 (U6-forward primer:
5′-GTCGTATCCAGTGCAGGGTCCGAGGT-3′; U6-reverse primer:
5′-GCACTGGATACGACAAAATATGGAAC-3′). Data analysis was performed
using the 2−ΔΔCq method (16).
Cell proliferation assay
Cell proliferation was measured using a Cell
Counting Kit-8 (CCK-8) assay (Beyotime Institute of Biotechnology,
Haimen, China) according to manufacturer's protocol. Briefly, cells
were plated in 96-well plates at a density of 0.5×104
cells/well. CCK-8 (10 µl) was added to each well prior to the
colorimetric measurement. After 1 h incubation at 37°C, the
absorbance at 450 nm was measured using a microplate luminometer
reader.
Cell apoptosis assay
Apoptosis assays were performed using an Annexin V
apoptosis detection kit according to the manufacturer's protocol
(BD Biosciences, San Jose, CA, USA). Briefly, CRC cells were
collected after 72 h transfection, washed with PBS and resuspended
in binding buffer containing 10 mM HEPES (pH 7.4), 2.5 mM
CaCl2, and 140 mM NaCl. Annexin V-PE and 7-AAD were then
added and flow cytometry analysis was performed after 15 min of
incubation at room temperature.
Immunoblotting
Total protein was extracted using whole cell lysis
buffer (Beyotime Institute of Biotechnology). Immunoblotting was
performed as previously described (13). The following antibodies were used: 1,
Anti-E2F transcription factor 1 (E2F1) (1:1,000; Cell Signaling
Technology, Inc., Danvers, MA, USA); 2, anti-caspase-9 antibodies
(1:3,000; Cell Signaling Technology, Inc.); 3, anti-GAPDH
antibodies (1:10,000; Zhongshan Goldenbridge Biotechnology,
Guangzhou, China). The membrane was washed with Tris-buffered
saline with Tween-20 and incubated with a peroxidase-conjugated
secondary antibody (1:1,000; Santa Cruz Biotechnology) for 1 h.
Protein bands were visualized using enhanced chemiluminesence
substrates (EMD Millipore, Billerica, MA, USA).
Statistical analysis
Data are expressed as the mean ± standard deviation
and were analyzed using SPSS 13.0 software (SPSS Inc., Chicago, IL,
USA). One-way analysis of variance was used for comparing multiple
groups, followed with the Student-Newman-Keuls post hoc test.
P<0.05 was considered to indicate a statistically significant
difference. All experiments were performed in biological
triplicates.
Results
miR-106a overexpression inhibits the
proliferation of colorectal cancer cells in vitro
The effect of miR-106a expression on the
proliferation of CRC cells was examined first. HCT116 and SW620
cells were transiently transfected with nontarget control or
miR-106a mimics. The RT-qPCR results demonstrated that cells
transfected with miR-106a mimics exhibited a significantly higher
level of miR-106a compared with cells transfected with a nontarget
control miRNA or with mock transfection (Fig. 1A). In addition, cells overexpressing
miR-106a mimics revealed a significant decrease in the cell
proliferation rate 48 and 72 h after transfection (Fig. 1B).
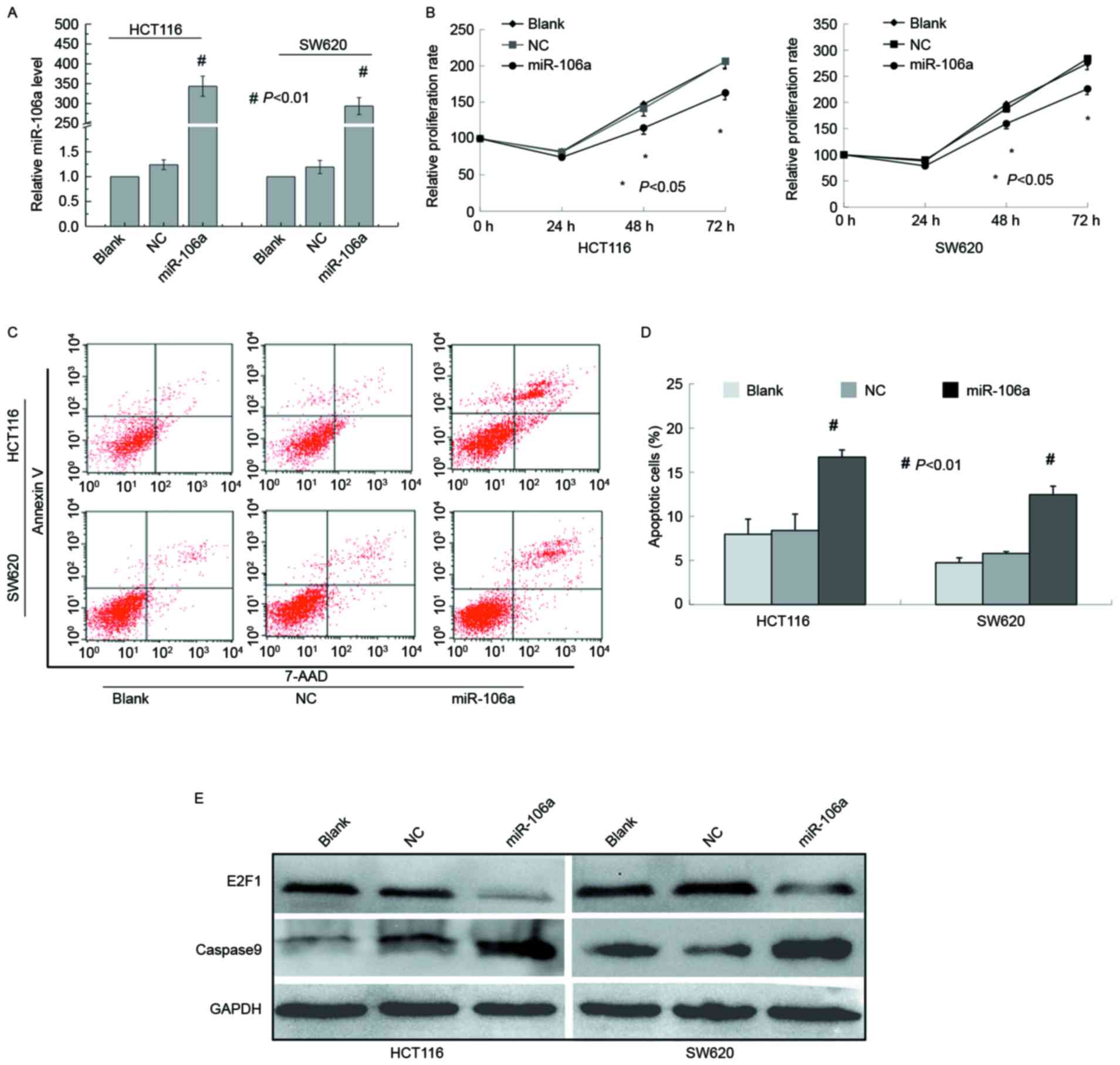 | Figure 1.miR-106a inhibits the proliferation
and induces the apoptosis of CRC cells. (A) Expression of miR-106a
was determined using reverse transcription-quantitative polymerase
chain reaction in HCT116 and SW620 cells 48 h after transfection
with blank, NC, and miR-106a mimics. (B) Relative cell
proliferation was determined using a cell counting kit-8 assay in
HCT116 and SW620 cells transfected with blank, NC, and miR-106a
mimics. (C) Apoptosis was analyzed with flow cytometry in HCT116
and SW620 cells transfected with blank, NC, and miR-106a mimics.
(D) Graphical representation of the apoptosis of CRC cells using
flow cytometry analysis as presented in C. (E) E2F1 and caspase-9
were detected using western blot assays in HCT116, and SW620 cells
transfected with blank, NC, and miR-106a mimics. Data are expressed
as the mean ± standard deviation following three independent
experiments. *P<0.05; #P<0.01. NC, nontarget
control; miR, microRNA; CRC, colorectal cancer; E2F1, E2F
transcription factor 1. |
miR-106a overexpression enhances
apoptosis of colorectal cancer cells in vitro
Whether miR-106a overexpression induces apoptosis of
CRC cells was examined next. Cells transfected with miR-106a or
controls were stained with Annexin V and 7-AAD to measure cell
apoptosis 72 h after transfection. miR-106a mimics caused
significant increases in the percentage of apoptotic cells (HCT116:
miR-106a, 16.69±0.82 vs. nontarget control, 8.38±1.86%; SW620:
miR-106a, 12.44±0.97 1 vs. nontarget control, 5.77±0.21%), while no
significant difference was identified between the blank and
nontarget control group (Fig. 1C and
D).
miR-106a overexpression causes a
marked change in the expression of E2F1 and cleaved caspase-9
Immunoblot analysis revealed that the protein level
of E2F1 was decreased following miR-106a overexpression.
Furthermore, caspase-9 cleavage was examined upon miR-106a
overexpression and it was demonstrated that active caspase-9 was
significantly enhanced compared with control cells (Fig. 1E).
Discussion
miRNAs have been demonstrated to serve essential
roles in carcinogenesis (17–21). Aberrant expression of certain miRNAs
has been implicated in a variety of tumor properties, including
growth, carcinogenesis, angiogenesis and apoptosis. The role of
miR-106a serves in the malignant progression of tumors is complex
and controversial. Numerous studies have revealed that miR-106a may
serve as a tumor suppressor or an oncogene in different cancer
types, which may be dependent on the cellular context (22,23). For
example, miR-106a exerts a tumor suppressive effect via suppressing
proliferation and inducing apoptosis in human glioma cells by
targeting E2F1, regardless of p53 status (22). In breasts cancer, miR-106a enhances
cell proliferation and migration by negatively regulating
zinc-finger and BTB domain containing 4 (23). However, the role of miR-106a in CRC
carcinogenesis remains unclear.
The present work identified miR-106a as a
tumor-suppressive miRNA in CRC cells in vitro.
Overexpression of miR-106a induced apoptosis and decreased
proliferation in CRC cells. The oncogene E2F1, target gene of
miR-106a was decreased following miR-106a overexpression while
apoptosis was elicited. Simultaneously, caspase-9 expression, an
important caspase in the apoptotic pathway, was induced. Once
active, caspase-9 triggers a cascade of caspase activation events
leading to apoptosis.
Collectively, the results of the current study
demonstrated that miR-106a may serve as a tumor suppressor in CRC,
potentially through regulating E2F1 and caspase-9 expression in
cultured cancer cells. However, in vivo studies are
warranted to validate these findings.
Acknowledgements
The authors would like to thank Dr Dennis Liang Fei
of National Institutes of Health (Bethesda, USA) for providing
editorial assistance.
References
|
1
|
Tenesa A and Dunlop MG: New insights into
the aetiology of colorectal cancer from genome-wide association
studies. Nat Rev Genet. 10:353–358. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gu S, Jin L, Zhang F, Sarnow P and Kay MA:
Biological basis for restriction of microRNA targets to the 3′
untranslated region in mammalian mRNAs. Nat Struct Mol Biol.
16:144–150. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of microRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Esquela-Kerscher A and Slack FJ:
Oncomirs-microRNAs with a role in cancer. Nat Rev Cancer.
6:259–269. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xi Y, Shalgi R, Fodstad O, Pilpel Y and Ju
J: Differentially regulated micro-RNAs and actively translated
messenger RNA transcripts by tumor suppressor p53 in colon cancer.
Clin Cancer Res. 12:2014–2024. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
O'Donnell KA, Wentzel EA, Zeller KI, Dang
CV and Mendell JT: c-Myc-regulated microRNAs modulate E2F1
expression. Nature. 435:839–843. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Woods K, Thomson JM and Hammond SM: Direct
regulation of an oncogenic micro-RNA cluster by E2F transcription
factors. J Biol Chem. 282:2130–2134. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
He L, Thomson JM, Hemann MT,
Hernando-Monge E, Mu D, Goodson S, Powers S, Cordon-Cardo C, Lowe
SW, Hannon GJ and Hammond SM: A microRNA polycistron as a potential
human oncogene. Nature. 435:828–833. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Johnson SM, Grosshans H, Shingara J, Byrom
M, Jarvis R, Cheng A, Labourier E, Reinert KL, Brown D and Slack
FJ: RAS is regulated by the let-7 microRNA family. Cell.
120:635–647. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lu J, Getz G, Miska EA, Alvarez-Saavedra
E, Lamb J, Peck D, Sweet-Cordero A, Ebert BL, Mak RH, Ferrando AA,
et al: MicroRNA expression profiles classify human cancers. Nature.
435:834–838. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Roldo C, Missiaglia E, Hagan JP, Falconi
M, Capelli P, Bersani S, Calin GA, Volinia S, Liu CG, Scarpa A and
Croce CM: MicroRNA expression abnormalities in pancreatic endocrine
and acinar tumors are associated with distinctive pathologic
features and clinical behavior. J Clin Oncol. 24:4677–4684. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ventura A and Jacks T: MicroRNAs and
cancer: Short RNAs go a long way. Cell. 136:586–591. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Schetter AJ, Leung SY, Sohn JJ, Zanetti
KA, Bowman ED, Yanaihara N, Yuen ST, Chan TL, Kwong DL, Au GK, et
al: MicroRNA expression profiles associated with prognosis and
therapeutic outcome in colon adenocarcinoma. JAMA. 299:425–436.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Díaz R, Silva J, García JM, Lorenzo Y,
García V, Peña C, Rodríguez R, Muñoz C, García F, Bonilla F and
Domínguez G: Deregulated expression of miR-106a predicts survival
in human colon cancer patients. Genes Chromosomes Cancer.
47:794–802. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Link A, Balaguer F, Shen Y, Nagasaka T,
Lozano JJ, Boland CR and Goel A: Fecal MicroRNAs as novel
biomarkers for colon cancer screening. Cancer Epidemiol Biomarkers
Prev. 19:1766–1774. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Calin GA, Liu CG, Sevignani C, Ferracin M,
Felli N, Dumitru CD, Shimizu M, Cimmino A, Zupo S, Dono M, et al:
MicroRNA profiling reveals distinct signatures in B cell chronic
lymphocytic leukemias. Proc Natl Acad Sci USA. 101:11755–11760.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Calin GA, Sevignani C, Dumitru CD, Hyslop
T, Noch E, Yendamuri S, Shimizu M, Rattan S, Bullrich F, Negrini M
and Croce CM: Human microRNA genes are frequently located at
fragile sites and genomic regions involved in cancers. Proc Natl
Acad Sci USA. 101:2999–3004. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jovanovic M and Hengartner MO: miRNAs and
apoptosis: RNAs to die for. Oncogene. 25:6176–6187. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kent OA and Mendell JT: A small piece in
the cancer puzzle: microRNAs as tumor suppressors and oncogenes.
Oncogene. 25:6188–6196. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Volinia S, Calin GA, Liu CG, Ambs S,
Cimmino A, Petrocca F, Visone R, Iorio M, Roldo C, Ferracin M, et
al: A microRNA expression signature of human solid tumors defines
cancer gene targets. Proc Natl Acad Sci USA. 103:2257–2261. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang G, Zhang R, Chen X, Mu Y, Ai J, Shi
C, Liu Y, Shi C, Sun L, Rainov NG, et al: MiR-106a inhibits glioma
cell growth by targeting E2F1 independent of p53 status. J Mol Med
(Berl). 89:1037–1050. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kim K, Chadalapaka G, Lee SO, Yamada D,
Sastre-Garau X, Defossez PA, Park YY, Lee JS and Safe S:
Identification of oncogenic microRNA-17-92/ZBTB4/specificity
protein axis in breast cancer. Oncogene. 31:1034–1044. 2012.
View Article : Google Scholar : PubMed/NCBI
|














