Introduction
Hepatocellular carcinoma (HCC) is the sixth most
common malignancy and the third leading cause of cancer-associated
mortality worldwide (1,2). In particular, China has a high
prevalence of HCC attributable to significant hepatitis B virus
infection rates (3). The majority of
patients with HCC present with intrahepatic or distant metastasis
at the time of diagnosis, thereby this reduces the possibility of
successful surgical resection or transplantation. Therefore, the
development of novel therapeutics for the treatment of advanced HCC
is crucial.
Sorafenib, the first-line systemic drug approved by
the US Food and Drug Administration for advanced hepatocellular
carcinoma, is a multi-kinase inhibitor that exerts inhibitory
effects on tumor cell proliferation and angiogenesis through the
inactivation of serine-threonine kinase Raf-1, which participates
in the Ras/Raf/MEK/mitogen-activated protein kinase (MAPK)
signaling cascade. Additionally, other receptor tyrosine kinases
are also the target of sorafenib including vascular endothelial
growth factor receptor, platelet-derived growth factor receptor,
and fibroblast growth factor receptor (4,5).
Previously, a series of clinical randomized controlled trials have
shown that sorafenib provides a survival benefit compared with
placebo therapy (6,7).
However, acquired drug resistance in patients with
HCC is prevalent in patients who are exposed to sorafenib and other
chemotherapeutic agents over a long period of time. Thus, further
studies examining the molecular mechanisms by which sorafenib
resistance occurs in patients with HCC are needed in order to
develop novel therapeutic strategies. Previous studies have
attempted to address the mechanisms involved in the development of
sorafenib resistance. Indeed, Chen et al (8) demonstrated that the activation of
phosphoinositide 3-kinase (PI3K)/Akt signaling is associated with
acquired resistance to sorafenib in HCC. Furthermore, previous
studies have also revealed that epithelial-mesenchymal transition
(EMT) is able to render cancer cells not only with drug resistance,
but also with invasive and metastatic properties (9). Additionally, it has been suggested that
the activation of the PI3K/Akt signaling pathway may promote EMT
(8,10). However, whether activation of the
PI3K/Akt cascade results in enhanced invasion of sorafenib
resistant cells associated with EMT remains unclear.
In the present study, the changes in Huh7 cells with
regard to morphology, migration and invasion properties following
sorafenib treatment were examined. Furthermore, alterations in the
PI3K/Akt signaling pathway and activation of EMT following
long-term exposure to sorafenib were analyzed. Finally, the effect
of a PI3K inhibitor, LY294002, on the viability, migration and
invasion of resistant Huh7 cancer cells was investigated.
Materials and methods
Reagents and antibodies
Sorafenib was purchased from Bayer AG (Leverkusen,
Germany). The PI3K inhibitor LY294002 and all antibodies for
immunoblotting were obtained from Cell Signaling Technology, Inc.
(Danvers, MA, USA), unless specified below.
Cell culture
Huh7, a human HCC cell line, was obtained from the
Cell Bank Type Culture Collection of Chinese Academy of Sciences
(Shanghai, China). The cells were cultured in Dulbecco's modified
Eagle's medium (DMEM; Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) containing 10% fetal bovine serum (FBS; Gibco;
Thermo Fisher Scientific, Inc.) in a humidified 5% CO2
incubator at 37°C.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted using Trizol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.), and the concentration
of RNA was quantified by SmartSpec Plus spectrophotometer (Bio-rad
Laboratories, Inc., Hercules, CA, USA). The first-strand
complementary DNA was synthesized using the Prime Script RT reagent
kit with gDNA Eraser (Takara Biotechnology Co., Ltd., Dalian,
China). qPCR reactions were performed using SYBR Premix Ex Taq
(Takara Biotechnology Co., Ltd). Briefly, the amplification
protocol was carried out using an initial denaturation at 95°C for
30 sec, followed by 40 cycles for 5 sec at 95°C, 30 sec at 60°C.
Following the completion of the PCR amplification reaction, the
relative expression levels of the gene of interest were analyzed by
the 2−ΔΔCq method (11),
and the amplification and melting curves were analyzed. All primer
sequences (Sangon Biotech, Shanghai, China) used are listed in
Table I.
 | Table I.Primer sequences for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primer sequences for reverse
transcription-quantitative polymerase chain reaction.
| Gene | Sequence (5′-3′) |
|---|
| ABCB1 |
|
| Forward |
GTGCTGGTTGCTGCTTACAT |
| Reverse |
AGCCTATCTCCTGTCGCATT |
| ABCC1 |
|
| Forward |
GCTGTGAAGACCCAGGAGAG |
| Reverse |
AAGCACCAGGAAACCACTTG |
| ABCG2 |
|
| Forward |
ATCCTTCCATCTTGTTCTTGG |
| Reverse |
CGTCCCTGCTTAGACATCCT |
| Snail |
|
| Forward |
GTGGTTCTTCTGCGCTACTG |
| Reverse |
AGGGCTGCTGGAAGGTAAAC |
| Slug |
|
| Forward |
GAGCATTTGCAGACAGGTCA |
| Reverse |
TCCTCATGTTTGTGCAGGAG |
Cell viability assay
The effect of individual treatment agents on cell
viability was assessed with the Cell Counting Kit-8 (CCK-8) assay
(Dojindo Molecular Technologies, Inc., Kumamoto, Japan) in three
replicates. The cells (5,000/well) were seeded in 96-well plates
and incubated for 24 h at 37°C. The cells were then incubated with
100 µl of fresh medium containing indicated concentrations (0, 1,
2, 4, 6, 8, 10 and 12 µM) of sorafenib for 48 h at 37°C. Finally,
10 µl CCK-8 was added to each well for 1 h at 37°C, and the optical
density was measured at 450 nm using a microplate reader (Bio-Rad
Laboratories, Inc.). The experiments were repeated three times.
Western blotting
Total protein was extracted using ice-cold cell
lysis buffer (Beyotime Institute of Biotechnology, Haimen, China)
for 30 min. The mixture was centrifuged at 10,000 g for 15 min at
4°C. The upper layer of separation was collected and stored at
−20°C until further use. A total of 30 µg total protein were
separated by 10% SDS-PAGE, followed by wet transfer to
polyvinylidene difluoride membranes. The membranes were blocked
with 5% skimmed milk powder dissolved in phosphate buffered saline
containing 0.1% Tween-20 (PBS-T) at room temperature for 1 h. The
membranes were incubated overnight at 4°C with the following
primary antibodies: PI3K (cat. no. ab-125633; 1:1,000 dilution;
Abcam, Cambridge, UK), phosphorylated (p)-Akt (cat. no. ab-38449;
1:1,000 dilution; Abcam), Akt (cat. no. ab-126580; 1:1,000
dilution; Abcam), p-extracellular signal-regulated kinase (ERK)1/2
(cat. no. 4370; 1:1,000 dilution; Cell Signaling Technology, Inc.),
ERK1/2 (cat. no. 4695; 1:2,000 dilution; Cell Signaling Technology,
Inc.), phosphatase and tensin homolog, E-cadherin (cat. no.
ab-15148; 1:1,000 dilution; Abcam), N-cadherin (cat. no. ab-18203;
1:1,000 dilution; Abcam), Snail (cat. no. 3879; 1:500 dilution;
Cell Signaling Technology, Inc.), Slug (cat. no. 9585; 1:500
dilution; Cell Signaling Technology, Inc., Danvers, MA, USA),
Vimentin (cat. no. ab-92547; 1:1,000 dilution; Abcam) and GAPDH
(cat. no. ab-9485; 1:1,000 dilution; Abcam). After washing three
times with PBST, the membranes were incubated with the secondary
antibodies (horseradish peroxidase-conjugated anti-rabbit IgG; cat.
no. 4414; 1:2,000 dilution; Cell Signaling Technology, Inc.) for 1
h at room temperature. After repetition of the washing step with
PBST, the membranes were visualized using enhanced
chemiluminescence western blotting detection reagents (BeyoECL
Plus; Beyotime Institute of Biotechnology).
Migration and invasion Transwell
assays
Migration and invasion assays were performed using
Transwell chambers containing a membrane filter with 8 µm pores
(Corning Incorporated, Corning, NY, USA) inserted in 24-well
plates. The upper chambers were coated with Matrigel (BD
Biosciences, San Jose, CA, USA), and the top of each well was
seeded with 5×104 cells in a total volume of 100 µl
serum-free DMEM. The lower chamber wells were loaded with 500 µl
DMEM containing 10% FBS which served as a chemo-attractant. After
60 h of incubation, the invaded cells were fixed with 4%
paraformaldehyde for 15 min at room temperature and then stained
with 0.1% crystal violet for 2 h. The invaded cells were counted
under a phase-contrast microscope in five random fields per insert
at ×100 magnification. A cell migration assay was performed using a
similar protocol with the exception that Matrigel was not added to
the upper chambers.
Statistical analysis
Data are presented as the mean ± standard deviation.
The one-way analysis of variance or Student's t-test was executed
to assess the statistical significance using SPSS version 17.0
software (SPSS, Inc., Chicago, IL, USA) and P<0.05 was
considered to indicate a statistically significant difference.
Results
Characteristics of sorafenib-resistant
cells
Through long-term exposure to increasing
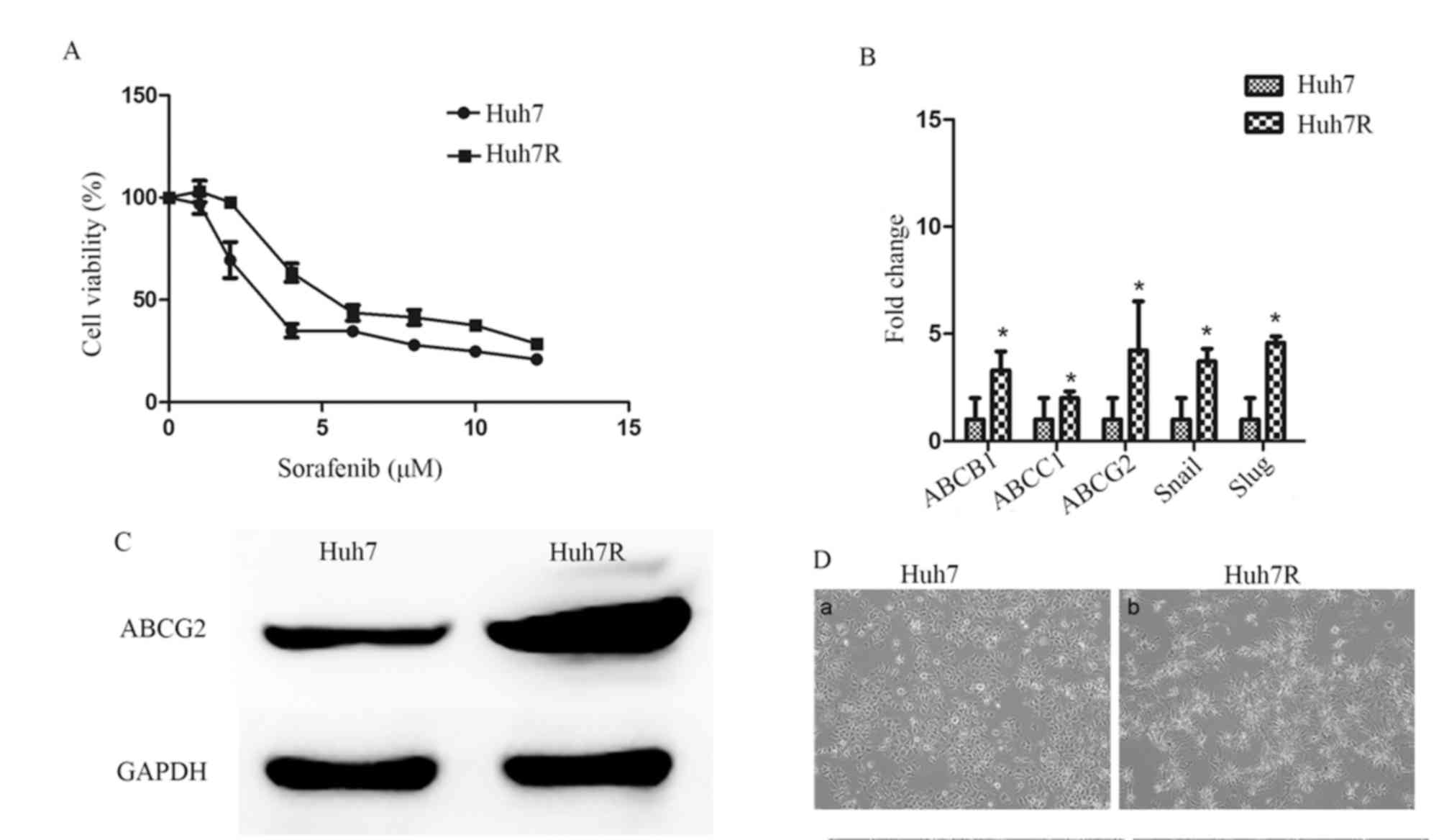
concentrations of sorafenib, Huh7 cells were successfully
transformed a sorafenib-resistant Huh7 cell line (Huh7R) was
established. Huh7R cells were less sensitive to sorafenib treatment
and their viability was markedly higher compared with that of
parental cells when exposed to the same concentration of sorafenib
(Fig. 1A). Furthermore, the
half-maximum inhibitory concentration (IC50) of the
resistant cells increased ~1.7-fold compared with the parental Huh7
cells (Table II). In addition, it
was demonstrated that there was an increased level of ABCB1, ABCC1
and ABCG2 mRNA expression (Fig. 1B)
and increased levels of ABCG2 protein in the Huh7R cell line
(Fig. 1C) compared with Huh7, further
confirming resistance to sorafenib. Notably, Huh7R cells exhibited
microscopically elongated fibroblastic processes and loss of
cell-to-cell contacts (Fig. 1D).
| Table II.IC50 of parental and
resistant cells following sorafenib treatment. |
Table II.
IC50 of parental and
resistant cells following sorafenib treatment.
| Cell line | IC50
(µM) |
|---|
| Huh7 | 3.7 |
| Huh7-R | 6.4 |
Epithelial-mesenchymal transition in
sorafenib-resistant cells
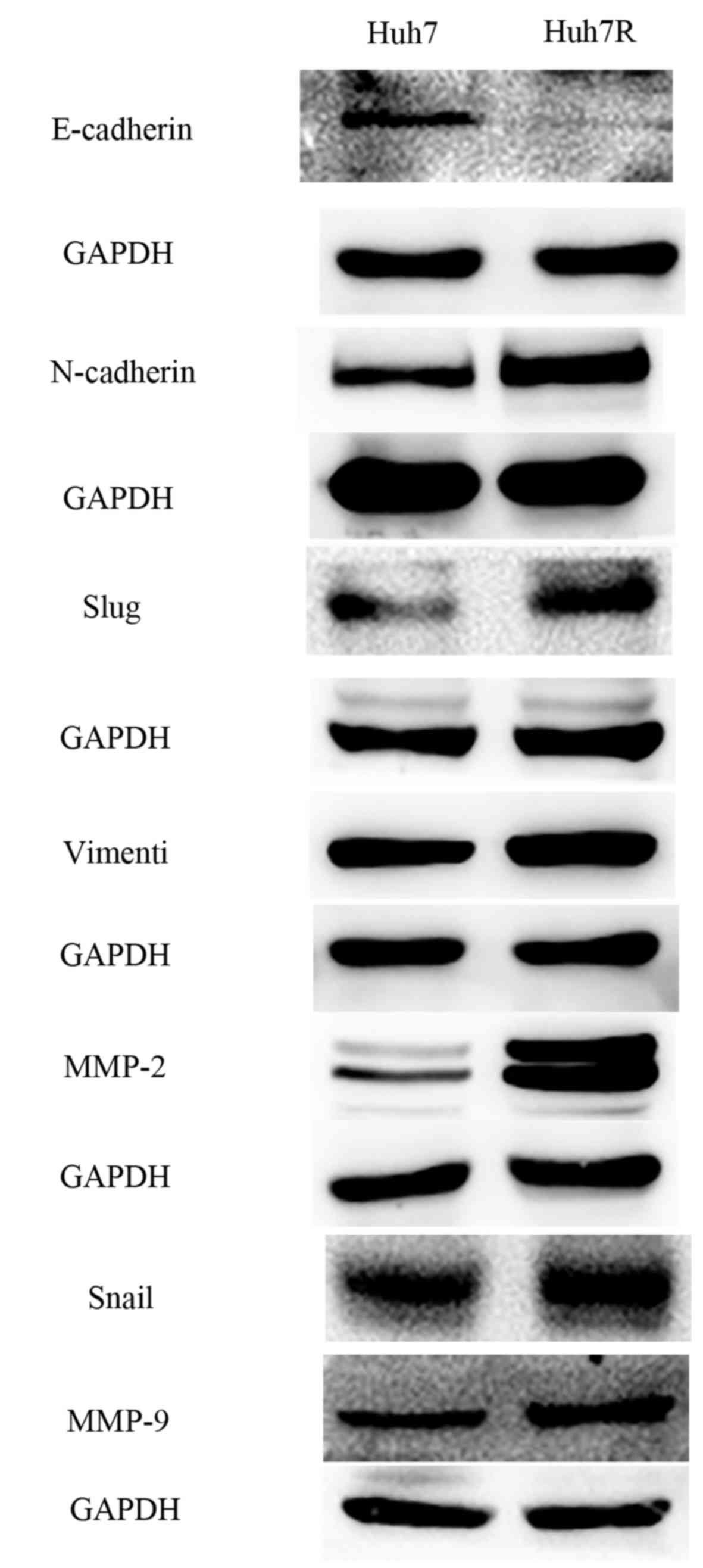
Resistant cells underwent morphological changes,
which were likely to be associated with EMT (Fig. 1D). To evaluate whether these
morphological changes in the resistant cell line Huh7R were
associated with EMT, markers characteristic of EMT were tested.
Huh7R cells exhibited upregulated mRNA expression of Snail and Slug
compared with Huh cells (Fig. 1B). In
addition, immunoblotting indicated a marked reduction in the
expression of the epithelial marker E-cadherin and a marked
upregulation of mesenchymal markers, including Snail, Slug,
vimentin and N-cadherin in Huh7R resistant cells compared with
control parental cells (Fig. 2).
Moreover, it was demonstrated that Huh7R resistant cells expressed
expression of matrix metalloproteinase (MMP)-2 and MMP-9 at the
protein level (Fig. 2). Together, the
present study indicated that EMT was a characteristic of
sorafenib-resistant cells.
Enhanced migration and invasion
capacities of sorafenib-resistant cells
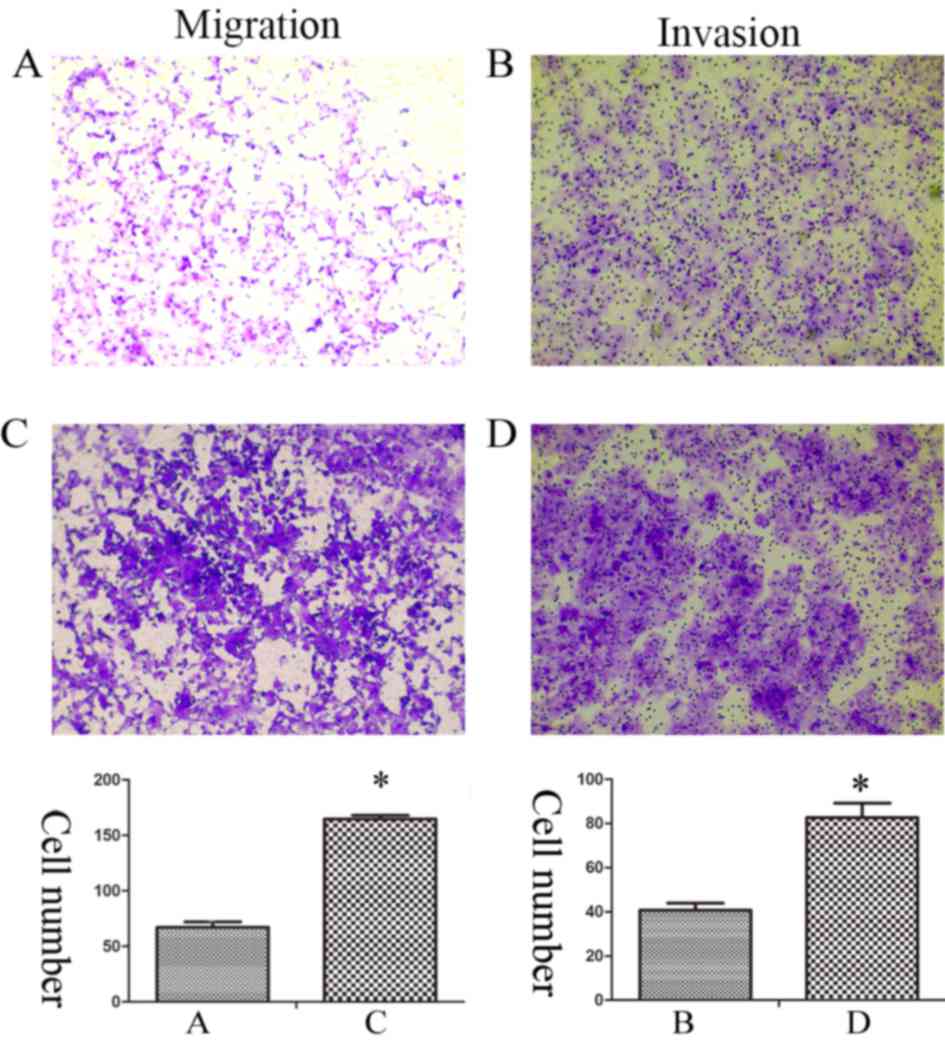
EMT is an important process, which results in
enhanced invasive and metastatic properties in cancer cells
(12). Therefore, Transwell assays
were performed in order to evaluate the migration and invasion
capacities of Huh7 and Huh7R cells. The data revealed that the
migration and invasion of Huh7R cells was significantly increased
compared with Huh7 cells (Fig.
3).
Development of sorafenib resistance is
associated with activation of the PI3K/Akt signaling pathway
To better understand the molecular mechanisms
associated with the development of sorafenib resistance, key
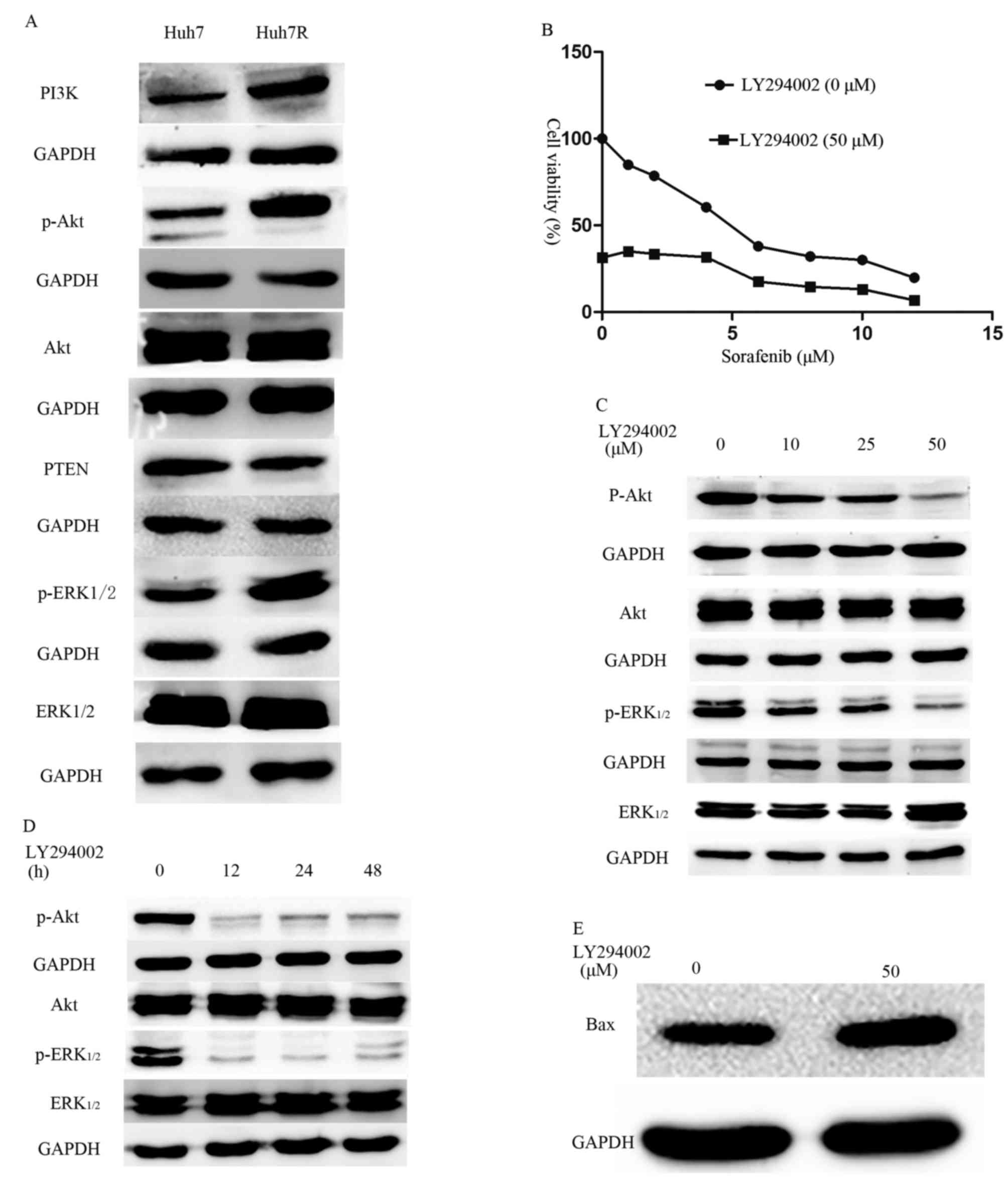
molecules within the PI3K/Akt and ERK pathways were investigated.
Huh7R resistant cells expressed increased levels of PI3K, p-Akt and
p-ERK1/2 compared with their parental Huh7 cells (Fig. 4A). The results from the present study
indicate that long-term exposure of HCC cells to sorafenib may
activate the PI3K/Akt and ERK signaling pathways. Based on findings
by Chen et al (8), the
possibility of inhibiting PI3K with LY294002 in order to reverse
sorafenib resistance was investigated. Indeed, it was demonstrated
that a combination of different concentrations of sorafenib and a
non-toxic dose (50 µM) of LY294002 resulted in decreased viability
of resistant cells (Fig. 4B) compared
with untreated cells. Furthermore, it was observed that the
combined treatment caused attenuation of the p-Akt and increased
expression of pro-apoptotic proteins, including Bax (Fig. 4C-E) compared with the expression in
untreated cells. Furthermore, the activation of Akt and ERK1/2 was
suppressed in Huh7R resistant cells that were treated with
LY294002, in a dose- and time-dependent manner (Fig. 4C-E).
Enhanced migration and invasion is
associated with activation of the PI3K/Akt signaling pathway
To better understand the signaling mechanisms
involved in the invasion of Huh7R cells following long-term
exposure to sorafenib, the activation of PI3K/Akt and ERK signaling
pathways was studied. Treatment of sorafenib resistant cells with
the PI3K inhibitor, LY294002, markedly attenuated the activation of
Akt (Ser473) and ERK1/2 in a concentration and time-dependent
manner. Furthermore, treatment with LY294002 was able to suppress
the expression of mesenchymal markers including N-cadherin,
vimentin, Snail and Slug, and induce the expression of E-cadherin,
which is an epithelial marker (Fig. 5A,
B). Furthermore, the treatment of sorafenib-resistant cells
suppressed the expression of MMP-2 in a concentration- and
time-dependent manner (Fig. 5C, D).
In addition, a dose of 50 µM LY294002 significantly prevented the
increased migratory and invasive abilities observed in
sorafenib-resistant Huh7R cells (Fig.
5E).
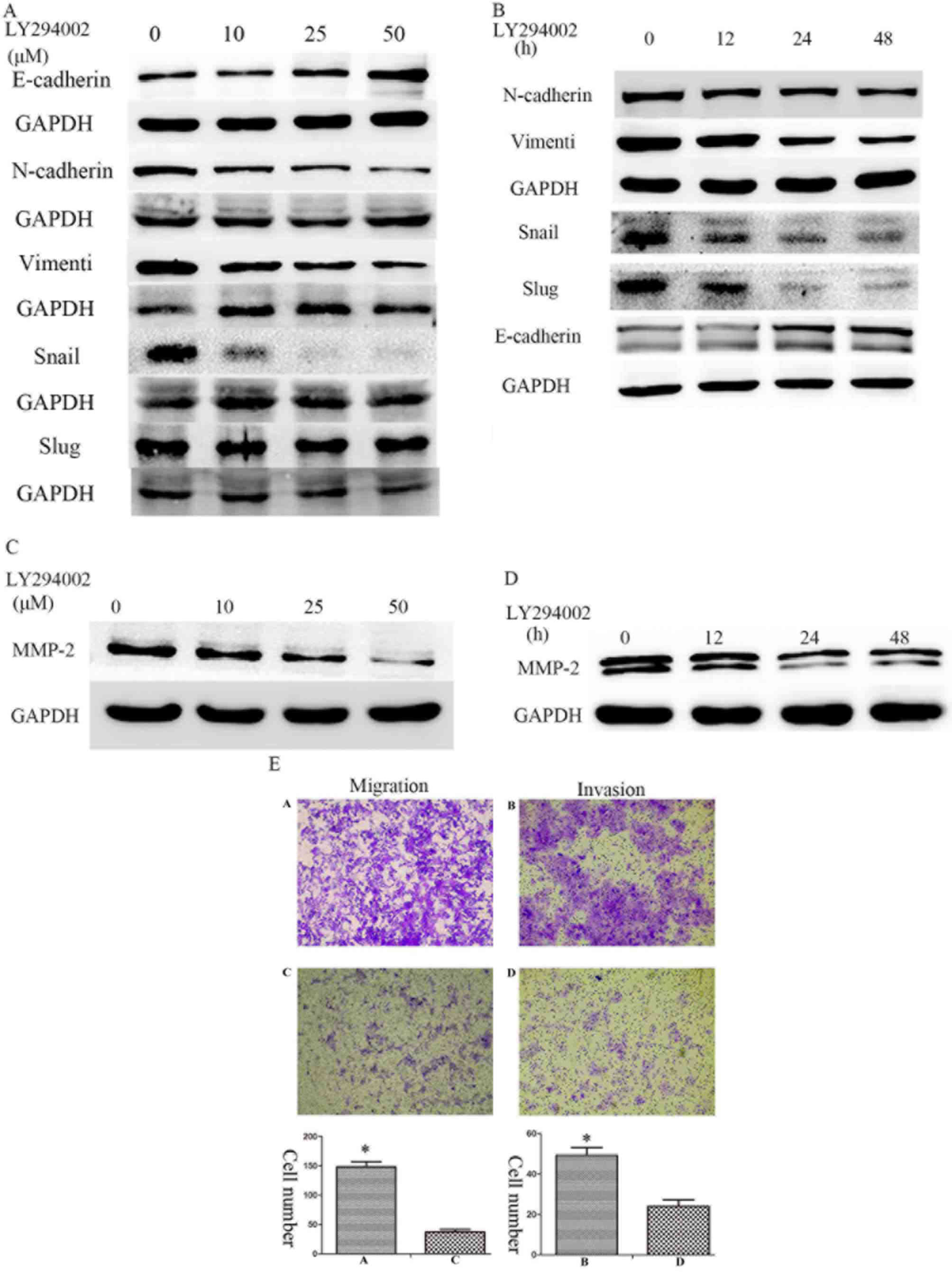 | Figure 5.Enhanced migration and invasion is
associated with the activation of the PI3K/Akt signaling pathway.
Expression of E-cadherin, N-cadherin, Vimentin, Snail and Slug was
examined in Huh7-R cells treated with (A) various doses of LY294002
(0, 10, 25 and 50 µM) and (B) various durations (0, 12, 24 and 48
h) of 50 µM LY294002. Expression of MMP-2 was examined in Huh7-R
cells treated with (C) various doses of LY294002 (0, 10, 25 and 50
µM) and (D) various durations (0, 12, 24 and 48 h) of 50 µM
LY294002. (E) Huh7R cells were treated with 0 and 50 µM LY294002.
The cells in panels (A) and (B) were treated with 0 µM LY294002,
and the cells in panels (C) and (D) were treated with 50 µM
LY294002. Migration and invasion were examined using Transwell
assays. The bar charts demonstrate the average number of migratory
or invasive cells in the experimental groups. Statistical analysis
was performed with Student's t-test. *P<0.05. |
Discussion
Sorafenib is the standard first-line systemic
therapy with clinically significant survival advantages for
patients with advanced HCC. Unfortunately, clinical evidence
confirms that the majority of all patients will eventually become
resistant to sorafenib treatment and experience disease progression
whilst remaining on therapy, consequently limiting the survival
benefit of sorafenib treatment. Chow et al (13) demonstrated that long-term exposure to
sorafenib increased migratory and invasive abilities of HCC cells
in vitro and in vivo, suggesting that an increase in
metastatic potential is a characteristic of resistant cells. Thus,
it is essential to further elucidate the biological mechanisms
involved in the development of chemoresistance and metastasis. In
the present study, it was demonstrated that long-term exposure to
sorafenib led to drug resistance and was able to increase the
migratory and invasive properties of resistant cells in
vitro. Importantly, it was also revealed that the activation of
the PI3K/Akt signaling pathway was a characteristic of
sorafenib-resistant cells.
EMT is a crucial biological process by which
malignant tumor cells undergo multiple morphological and
biochemical changes including increased cellular motility. This
conversion enables tumor cells to acquire drug resistance, and
invasive and metastatic properties (12,14), and
has indeed attracted considerable attention among researchers. The
process of EMT is associated with shorter disease-free survival and
poorer prognosis (9). In the present
study, Huh7R cells underwent morphological changes associated with
EMT, including enhanced migration and invasion. Moreover,
downregulation in E-cadherin and upregulation of Snail, Slug,
Vimentin and N-cadherin further confirmed that EMT was a feature of
Huh7R cells following long-term exposure to sorafenib. Consistent
with the data in the present study, Chow et al (13) previously demonstrated that
sorafenib-induced EMT promoted migration and invasion in HCC cells.
However, the molecular mechanisms involved in the process of EMT
have, to date, not been clearly elucidated.
Recently, increasing evidence has indicated that the
activation of the PI3K/Akt signaling pathway may play an important
role in promoting invasion and metastasis in resistant cells
(15). It is, therefore, of
significant interest to examine whether sorafenib-induced EMT is
attributable to the activation of the PI3K/Akt cascade. It has been
demonstrated that the PI3K/Akt and ERK signaling pathways are
involved in the process of EMT (16).
Thus, analysis of the status of the PI3K/Akt and ERK1/2 pathways
was examined in the present study. Notably, it was demonstrated
that inhibition of the PI3K/Akt signaling pathway with LY294002, a
PI3K inhibitor, had an effect on ERK and thus suppressed the
initiation of EMT. Together, these data suggest not only a
crosstalk between the PI3K/Akt and ERK1/2 signaling pathways, but
also that EMT may directly or indirectly regulate transcription
factors associated with these pathways, including Snail and
Slug.
The MMP-2 and MMP-9 have been associated with
migration and invasion of resistant cells. This phenomenon may be
regulated by the activation of the PI3K/Akt and MAPK signaling
pathways (17). The data of the
present study indicated that the PI3K/Akt cascade may directly or
indirectly regulate the properties of migration and invasion in
sorafenib-resistant cells via MMP-2 and MMP-9. Taken together, the
results suggest that the PI3K/Akt signaling pathway contributes a
pivotal role in directly or indirectly regulating the migratory and
invasive capacities of sorafenib-resistant cells via EMT and
MMPs.
The PI3K/Akt signaling cascade is known to be an
important signal transduction pathway involved in
hepatocarcinogenesis. Multiple studies have reported that the
activation of the PI3K/Akt signaling pathway induces
chemoresistance in HCC cells (18–20).
Meanwhile, the MAPK signaling pathway is a major target of
sorafenib therapy. However, several studies have also clearly
indicated that activation of the MAPK cascade is associated with
drug resistance (21–23). Concomitant with the present study,
overexpression of p-Akt and p-ERK1/2 results in sorafenib
resistance (24). The PI3K inhibitor
LY294002 is able to inhibit the activation of ERK, which indicates
that ERK activation may be induced by the PI3K/Akt signaling
pathway. In the present study, it was also demonstrated that
attenuation of the PI3K/Akt signaling pathway using LY294002 was
able to induce the expression of apoptotic proteins, including Bax,
and cell viability was also decreased. These data suggested that
the PI3K/Akt signaling pathway may have an important role in
sorafenib resistance.
In conclusion, the results from the present study
highlight the activation of the PI3K/Akt signaling pathway as a key
event in the acquisition of resistance and the enhancement of
invasive properties in sorafenib-resistant cells. Furthermore, the
present study indicates that combined treatment with sorafenib and
LY294002 may reduce resistance to sorafenib and prevent metastasis
in patients with HCC, providing an attractive novel therapeutic
regime in patients with advanced HCC.
Acknowledgements
Not applicable.
Funding
This study was supported by grants from Jiaxing
Science and Technology Projects (grant no. 2013AY21042-5), Jiaxing
Science and Technology innovation team projects (grant no.
2013-03), and major projects of Zhejiang Province on the
transformation of the appropriate technical achievements of primary
health care (grant no. 2013T301-12 and 2013T301-15).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Author contributions
HZ performed the preliminary studies on which this
work is based, conceived the study, collected and analysed data and
drafted the manuscript. QW performed the RT-PCR and western-blot
analysis and was also involved in the study design and editing of
the manuscript. JL and HC were involved in the experiment planning,
the drafting of the manuscript and for revising it critically for
content and general supervision of the project. All the authors
have read and approved the final manuscript.
Ethics approval and consent to
participate
Animal experiments were performed in accordance with
the animal guidelines of the First Hospital of Jiaxing Ethics
Review Committee for Animal Experimentation.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
HCC
|
hepatocellular carcinoma
|
|
EMT
|
epithelial-to-mesenchymal cell
transition
|
|
CCK-8
|
cell counting kit-8
|
|
MMP
|
matrix metalloproteinases
|
|
MAPK
|
mitogen-activated protein kinase
|
|
ERK
|
extracellular signal-regulated
kinase
|
|
IC50
|
half-maximum inhibitory
concentration
|
References
|
1
|
Forner A, Llovet JM and Bruix J:
Hepatocellular carcinoma. Lancet. 379:1245–1255. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu L, Cao Y, Chen C, Zhang X, McNabola A,
Wilkie D, Wilhelm S, Lynch M and Carter C: Sorafenib blocks the
RAF/MEK/ERK pathway, inhibits tumor angiogenesis, and induces tumor
cell apoptosis in hepatocellular carcinoma model PLC/PRF/5. Cancer
Res. 66:11851–11858. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen S, Wang Y, Ruan W, Wang X and Pan C:
Reversing multidrug resistance in hepatocellular carcinoma cells by
inhibiting extracellular signal-regulated kinase/mitogen-activated
protein kinase signaling pathway activity. Oncol Lett. 8:2333–2339.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cheng AL, Kang YK, Chen Z, Tsao CJ, Qin S,
Kim JS, Luo R, Feng J, Ye S, Yang TS, et al: Efficacy and safety of
sorafenib in patients in the Asia-Pacific region with advanced
hepatocellular carcinoma: A phase III randomised, double-blind,
placebo-controlled trial. Lancet Oncol. 10:25–34. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rimassa L and Santoro A: Sorafenib therapy
in advanced hepatocellular carcinoma: The SHARP trial. Expert Rev
Anticancer Ther. 9:739–745. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen KF, Chen HL, Tai WT, Feng WC, Hsu CH,
Chen PJ and Cheng AL: Activation of phosphatidylinositol
3-kinase/Akt signaling pathway mediates acquired resistance to
sorafenib in hepatocellular carcinoma cells. J Pharmacol Exp Ther.
337:155–161. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ombrato L and Malanchi I: The EMT
universe: Space between cancer cell dissemination and metastasis
initiation. Crit Rev Oncog. 19:349–361. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang H, Xu L, Zhu X, Wang P, Chi H and
Meng Z: Activation of phosphatidylinositol 3-kinase/Akt signaling
mediates sorafenib-induced invasion and metastasis in
hepatocellular carcinoma. Oncol Rep. 32:1465–1472. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
van Malenstein H, Dekervel J, Verslype C,
Van Cutsem E, Windmolders P, Nevens F and van Pelt J: Long-term
exposure to sorafenib of liver cancer cells induces resistance with
epithelial-to-mesenchymal transition, increased invasion and risk
of rebound growth. Cancer Lett. 329:74–83. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chow AK, Ng L, Lam CS, Wong SK, Wan TM,
Cheng NS, Yau TC, Poon RT and Pang RW: The Enhanced metastatic
potential of hepatocellular carcinoma (HCC) cells with sorafenib
resistance. PLoS One. 8:e786752013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mir N, Jayachandran A, Dhungel B, Shrestha
R and Steel JC: Epithelial-to-mesenchymal transition: A mediator of
sorafenib resistance in advanced hepatocellular carcinoma. Curr
Cancer Drug Targets. 17:698–706. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xu W, Yang Z and Lu N: A new role for the
PI3K/Akt signaling pathway in the epithelial-mesenchymal
transition. Cell Adh Migr. 9:317–324. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wen W, Ding J, Sun W, Fu J, Chen Y, Wu K,
Ning B, Han T, Huang L, Chen C, et al: Cyclin G1-mediated
epithelial-mesenchymal transition via phosphoinositide 3-kinase/Akt
signaling facilitates liver cancer progression. Hepatology.
55:1787–1798. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu YJ, Neoh CA, Tsao CY, Su JH and Li HH:
Sinulariolide suppresses human hepatocellular carcinoma cell
migration and invasion by inhibiting matrix metalloproteinase-2/-9
through MAPKs and PI3K/Akt signaling pathways. Int J Mol Sci.
16:16469–16482. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kunter I, Erdal E, Nart D, Yilmaz F,
Karademir S, Sagol O and Atabey N: Active form of AKT controls cell
proliferation and response to apoptosis in hepatocellular
carcinoma. Oncol Rep. 31:573–580. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dong J, Zhai B, Sun W, Hu F, Cheng H and
Xu J: Activation of phosphatidylinositol 3-kinase/AKT/snail
signaling pathway contributes to epithelial-mesenchymal
transition-induced multi-drug resistance to sorafenib in
hepatocellular carcinoma cells. PLoS One. 12:e01850882017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang PF, Li KS, Shen YH, Gao PT, Dong ZR,
Cai JB, Zhang C, Huang XY, Tian MX, Hu ZQ, et al: Galectin-1
induces hepatocellular carcinoma EMT and sorafenib resistance by
activating FAK/PI3K/AKT signaling. Cell Death Dis. 7:e22012016.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xu Y, Huang J, Ma L, Shan J, Shen J, Yang
Z, Liu L, Luo Y, Yao C and Qian C: MicroRNA-122 confers sorafenib
resistance to hepatocellular carcinoma cells by targeting IGF-1R to
regulate RAS/RAF/ERK signaling pathways. Cancer Lett. 371:171–181.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen W, Wu J, Shi H, Wang Z, Zhang G, Gao
Y, Jiang C and Ding Y: Hepatic stellate cell coculture enables
sorafenib resistance in Huh7 cells through HGF/c-Met/Akt and
Jak2/Stat3 pathways. Biomed Res Int. 2014:7649812014.PubMed/NCBI
|
|
23
|
Li QL, Gu FM, Wang Z, Jiang JH, Yao LQ,
Tan CJ, Huang XY, Ke AW, Dai Z, Fan J and Zhou J: Activation of
PI3K/AKT and MAPK pathway through a PDGFRβ-dependent feedback loop
is involved in rapamycin resistance in hepatocellular carcinoma.
PLoS One. 7:e333792012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xu J, Zheng L, Chen J, Sun Y, Lin H, Jin
RA, Tang M, Liang X and Cai X: Increasing AR by HIF-2α inhibitor
(PT-2385) overcomes the side-effects of sorafenib by suppressing
hepatocellular carcinoma invasion via alteration of pSTAT3, pAKT
and pERK signals. Cell Death Dis. 8:e30952017. View Article : Google Scholar : PubMed/NCBI
|