Introduction
Hydrofluoric (HF) acid is a highly dangerous toxic
substance, widely used in industrial engineering. Exposure could
result in unique severe cutaneous burns, systemic toxicity, and
occasionally lethal toxicity. HF acid burns are notorious for
progressive tissue necrosis and severe pain, which can penetrate
the skin and destruct deep tissue layers, as well as the bone. In
tissues, HF acid can react with calcium and magnesium ions,
producing insoluble fluoride salts which can induce myocardium
defects, hypocalcemia and changes in levels of active substances.
High concentrations or prolonged exposure may cause death (1). As low as 2.5% HF acid has been reported
to cause skin burns and even induce death.
Numerous topical and parenteral treatments have been
developed and are currently available. For example, calcium salts
have been proven effective for systemic dermal injuries. Calcium
gluconate (CG), aloe gel, A + D ointment, magnesium ointment and
Zephiran (benzalkonium chloride) are applied topically, and their
efficacy has been extensively investigated. Of these, topical CG
gel has emerged as an effective treatment of HF acid burns, with
recovery signs becoming apparent soon after treatment initiation.
Previous studies found that topical applications and subcutaneous
injections of CG could significantly deter the damage caused by
burns in the first 24 h, as well as enhance tissue recovery as
thetreatment progresses. It has been reported that CG can react
with HF acid to form insoluble, non-toxic calcium fluoride.
However, little is known regarding the molecular mechanism
underlying its therapeutic effect.
Wound healing involves the repairing of tissue
defects through enhanced cellular proliferation, migration and
differentiation. Human dermal fibroblasts (HDFs) are critical cells
for wound healing and tissue repairing after skin injuries
(2,3).
HDFs are responsible for generating connective tissue in the dermis
and repairing skin lesions. Moreover, HDFs produce proteins, such
as laminin and fibronectin, which can aid the formation of
extracellular matrix between dermis and epidermis, effectively
combining epidermal cells and, thus, forming a new top layer of the
skin (4,5). Therefore, HDFs are essential for proper
recovery following skin injury (6–8).
In this study, we investigated the possible roles of
CG in attenuating the toxic effect of HF acid on HDFs. We studied
the CG-driven activation of the wound healing process, by assessing
HDF proliferation, collagen synthesis and migratory properties. The
obtained results may offer new insights into the use of CG for the
treatment of dermal lesions.
Materials and methods
Cell culture
HDFs were purchased from ATCC (Guangzhou, China) and
cultured in DMEM medium (Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) containing 10% fetal bovine serum (FBS; Gibco;
Thermo Fisher Scientific, Inc.) and 1% penicillin/streptomycin
(Invitrogen; Thermo Fisher Scientific, Inc.) at 37°C, in a 5%
CO2 incubator. Cells of logarithm phase were used in our
study. Cell morphology of HDFs was observed by an optical
microscope (Olympus, Tokyo, Japan).
Vimentin immunofluorescence
staining
Cell morphology of HDFs was evaluated by vimentin
immunofluorescence staining assay. HDF cells were inoculated on
sterile coverslips in 6-well plates and cultured for 2 days at 37°C
in a 5% CO2 incubator. Paraformaldehyde functioned on
HDFs and membranes were broken then. HDFs were probed with rabbit
anti-human vimentin primary antibody and then with the appropriate
FITC-conjugated donkey anti-rabbit secondary antibody. After that,
HDFs were immunofluorescence stained by
4′,6-diamidino-2-phenylindole (DAPI). Finally, images were obtained
by DM5000 epifluorescence microscope (Leica, Wetzlar, Germany).
Cell viability assay
The effect of HF acid on cell viability of HDFs was
evaluated by the Cell Counting Kit-8 (CCK8) assay. HDFs were
treated with HF acid of different concentrations [0, 2, 4, 6, 8, 10
and 20% (v/v)], for determined amounts of time (0, 2, 4, 6, 8, 10
and 20 min). Briefly, following each treatment, cells were seeded
in 96-well plates, at an initial density of 5×103
cells/well, and incubated for the indicated amounts of time.
Subsequently, 20 µl CCK-8 reagent were added into each well. The
plates were then incubated for another 1 h. The optical density
(OD) values were read at 450 nm, using a microplate reader (Thermo
Fisher Scientific, Inc.). Data were expressed as percentage of
viable cells as follows: Relative viability (%)=[A450
(treated)-A450 (blank)]/[A450
(control)-A450 (blank)] ×100%.
The effect of CG on cell viability of
HF-acid-injured HDFs was also evaluated by the CCK8 assay. HDFs
were pre-treated with CG at different concentrations (50, 100 and
200 µmol/l) for 6, 12 and 24 h, respectively, after which they were
treated with 8% (v/v) HF acid for 6 min. The results were compared
with cell viability of HDFs solely treated with 8% (v/v) HF acid
for 6 min (HF control). The detection was performed as described
above.
Apoptosis detection
The apoptosis status was measured by Annexin V/PI
double-stain assay, according to the manufacturer's protocol
(Biovision, Mountain View, CA, USA). Briefly, after HF acid
treatment or additional CG pre-treatment, both floating and
trypsinized adherent cells (5×105) were collected and
resuspended in 500 µl binding buffer containing 5 µl Annexin V
fluorescein isothiocyanate and additional 5 µl Propidium Iodide
(PI; Invitrogen; Thermo Fisher Scientific, Inc.). Then cells were
incubated for 5 min in the dark, at the room temperature. Analysis
was immediately performed with a flow cytometer (BD Biosciences,
Franklin Lakes, NJ, USA). Cell Quest software was used to analyze
the apoptosis rate.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
The mRNA expression levels were measured by RT-qPCR.
Total RNA was extracted from HDFs with different treatment
respectively, using RNeasy kit (Qiagen, Valencia, CA, USA), and
cDNA was reversely transcribed with 1 µg RNA using a kit of
Quantiscript Reverse Transcriptase (Qiagen), according to the
protocol provided by the manufacture. PCR amplification was
performed for 30 sec at 95°C, followed by 40 cycles: Denaturation
at 95°C for 15 sec, annealing/extension at 60°C for 30 sec in ABI
7300 Thermocycler (Applied Biosystems; Thermo Fisher Scientific,
Inc.) using Fast SYBR Green Master Mix (Applied Biosystems; Thermo
Fisher Scientific, Inc.). The oligo nucleotide primer sequences
were displayed in Table I.
 | Table I.Primers used in reverse
transcription-quantitative polymerase chain reaction analysis. |
Table I.
Primers used in reverse
transcription-quantitative polymerase chain reaction analysis.
| Name | Type | Sequence (5′-3′) |
|---|
| Caspase-3 | Forward | TGTGAGGCGGTTGTAGAAG
AGT |
|
| Reverse | CACACCCACCGAAAACCA
GAG |
| Bax | Forward |
TGCTTCAGGGTTTCATCCA |
|
| Reverse |
GGCCTTGAGCACCAGTTT |
| Bcl-2 | Forward | ACGGTGGTGGAGGAGCT
CTT |
|
| Reverse |
CGGTTGACGCTCTCCACAC |
| Wnt2 | Forward | CTGACCTGATGCAGACGC
AAG |
|
| Reverse | AGGAGCCACCTGTAGCTCT
CATGTA |
| Wnt3a | Forward | GATGGTGTCTCGGGAGT
TCG |
|
| Reverse |
CCGTGGCACTTGCACTTGA |
| β-catenin | Forward | ATAAGAGCTCCTTGTGC
GGC |
|
| Reverse | GGCCATGTCCAACTCCA
TCA |
| GAPDH | Forward | TGACTTCAACAGCGACAC
CCA |
|
| Reverse | CACCCTGTTGCTGTAGCC
AAA |
Western blot analysis
The concentrations of proteins were determined by
BCA assay (Beyotime, Nantong, China). Then proteins were subjected
to sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE) and electroblotted onto a polyvinylidene fluoride (PVDF)
membrane (Amersham, Amersham, UK). Following the blockage of 5%
nonfat dry milk for 1 h, the blotting membranes were probed with
the primary antibodies at 4°C overnight respectively. which were
then probed with appropriate HRP-conjugated secondary antibodies.
The PVDF membranes were exposed to X-ray film, and the
immunoreactive bands were detected by reaction with enhanced
chemiluminescense (ECL) detection system reagents (Amersham,
Arlington Heights, IL, USA). For loading control, the membrane was
probed with a monoclonal antibody for glyceraldehyde-3-phosphate
dehydrogenase (GAPDH). Lab Works Image Acquisition and Analysis
Software (UVP, Upland, CA, USA) were used to quantify band
intensities. Antibodies were purchased from Abcam (Cambridge,
UK).
Enzyme-linked immunosorbent assay
(ELISA)
The amounts of proteins were determined by an ELISA
kit (R&D Systems, Minneapolis, MN, USA), following the
manufacturer's instructions. Samples and standard substances were
added into 96-well plates. After incubation for 90 min at 37°C,
biotinylated antibodies were added into the wells and incubated for
another 60 min at 37°C. After being rinsed for 3 times, Avidin
peroxidase complex was added and the plates were incubated for 30
min. Next, after being rinsed for 3 times, tetramethylbenzidine
coloring reagent was added and incubated for 15 min at 37°C.
Finally, the OD values were read at 450 nm by a microplate reader
(Thermo Fisher Scientific, Inc.). Protein amounts were calculated
by the generated standard curve.
Colorimetric assay
The level of L-hydroxyproline (L-HYP) in HDFs was
determined by the colorimetric method. In this assay, L-HYP was
oxidized by chloramine T, and redundant chloramine T was removed by
perchloric acid. After that, dimethyl amino benzaldehyde was added
to form a red complex. OD values were detected at 558 nm, by an
ultraviolet spectrophotometer (Mettler Toledo, Greifensee,
Switzerland). The amount of L-HYP in each sample was calculated
using the obtained standard curve.
Statistical analysis
All results were expressed as mean ± standard
deviations of three independent experiments. Statistical analysis
was performed using a SPSS 13.0 statistical package (SPSS, Inc.,
Chicago, IL, USA) and data were subjected to one-way analysis of
variance (ANOVA), followed by Dunnett's test. P<0.05 was
considered significant; P<0.01 was considered especially
significant.
Results
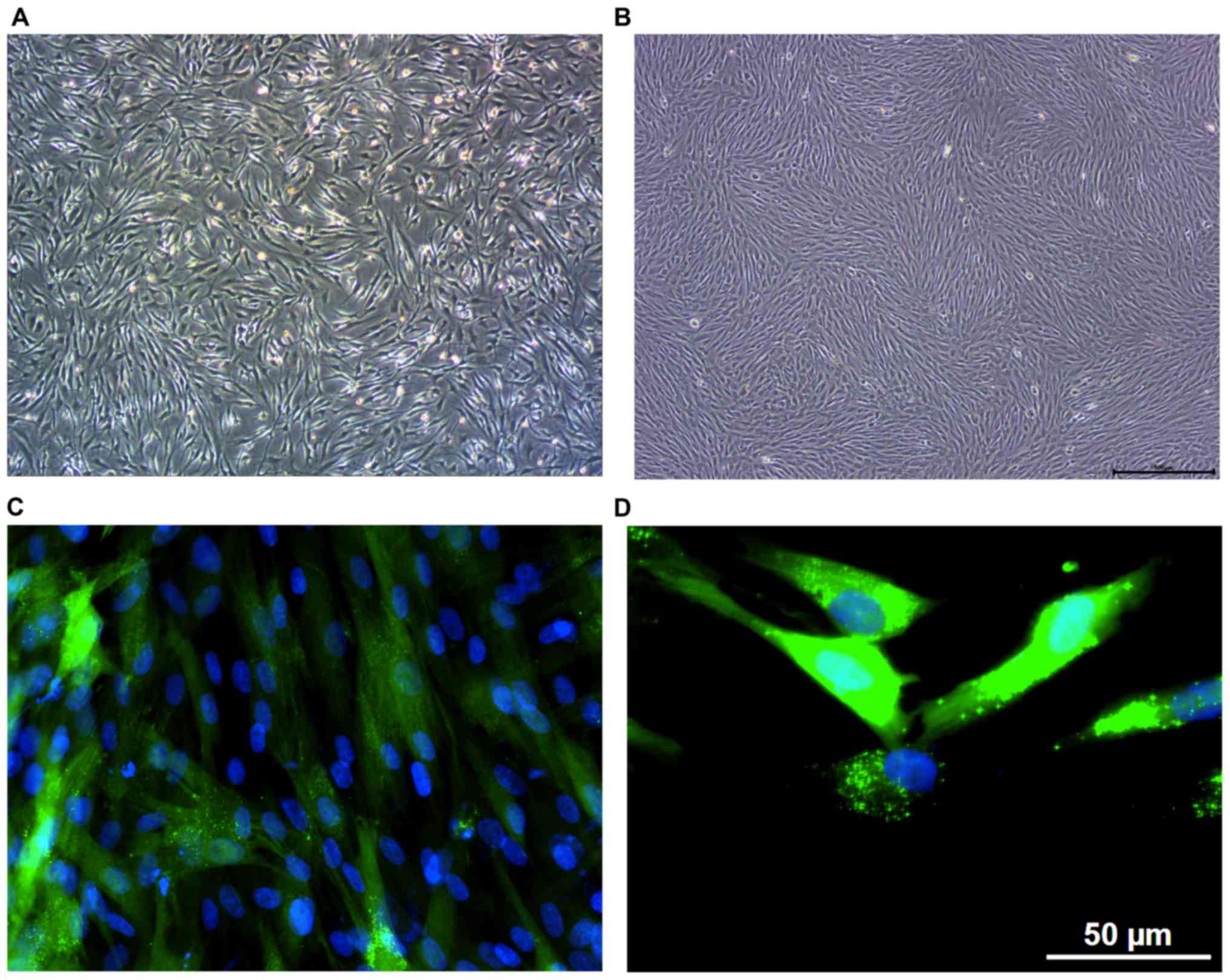
HDF morphology
HDF morphology was detected by optical microscopy
and the vimentin immunofluorescence staining assay. When observed
under an optical microscope, HDF cells appeared oval or
applanate-star-like, with plump cytoplasm, and clear nucleolus,
consistent with the classical morphology of HDFs. When the number
of cells increased, HDFs arranged closely with decreased cell
space, and became spiral- or fence-shaped (Fig. 1A and B). Following the microscopic
assessment, HDF morphology was detected by the vimentin
immunofluorescence staining assay. As a member of the intermediate
filament protein family, vimentin is the main component of the
cytoskeleton, being expressed in cells derived from mesoblastema,
especially HDFs. Consequently, HDFs can be detected by
intracellular vimentin in the cytoplasm. Results showed that the
oval cell nucleus was dyed blue by DAPI, whereas the spindle- or
applanate-star-shaped cytoplasm was dyed green, specifically
indicating the presence of vimentin around the nucleus (Fig. 1C and D).
Analysis of cell viability and
apoptosis of HDFs treated with HF acid
The effect of HF acid on the viability of HDFs,
measured by the CCK8 assay, was shown in Fig. 2A. Cell viability was suppressed in the
HF-treated groups in dose-dependent (2, 4, 6, 8, 10 and 20% (v/v))
and time-dependent manners (2, 4, 6, 8, 10 and 20 min), compared
with the control group. HDF viability varied significantly when
treated with 8, 10 and 20% (v/v) HF acid for 6, 8, 10 or 20 min
(P< 0.05), compared with the control group. However, the
viability of HDFs treated with 20% (v/v) HF acid for 10 or 20 min
was similar to that of cells treated with 8 or 10% (v/v) HF acid
for 6 or 8 min, so detection of apoptosis was performed on HDFs
treated with lower concentrations of HF acid [8 and 10% (v/v)] for
6 and 8 min.
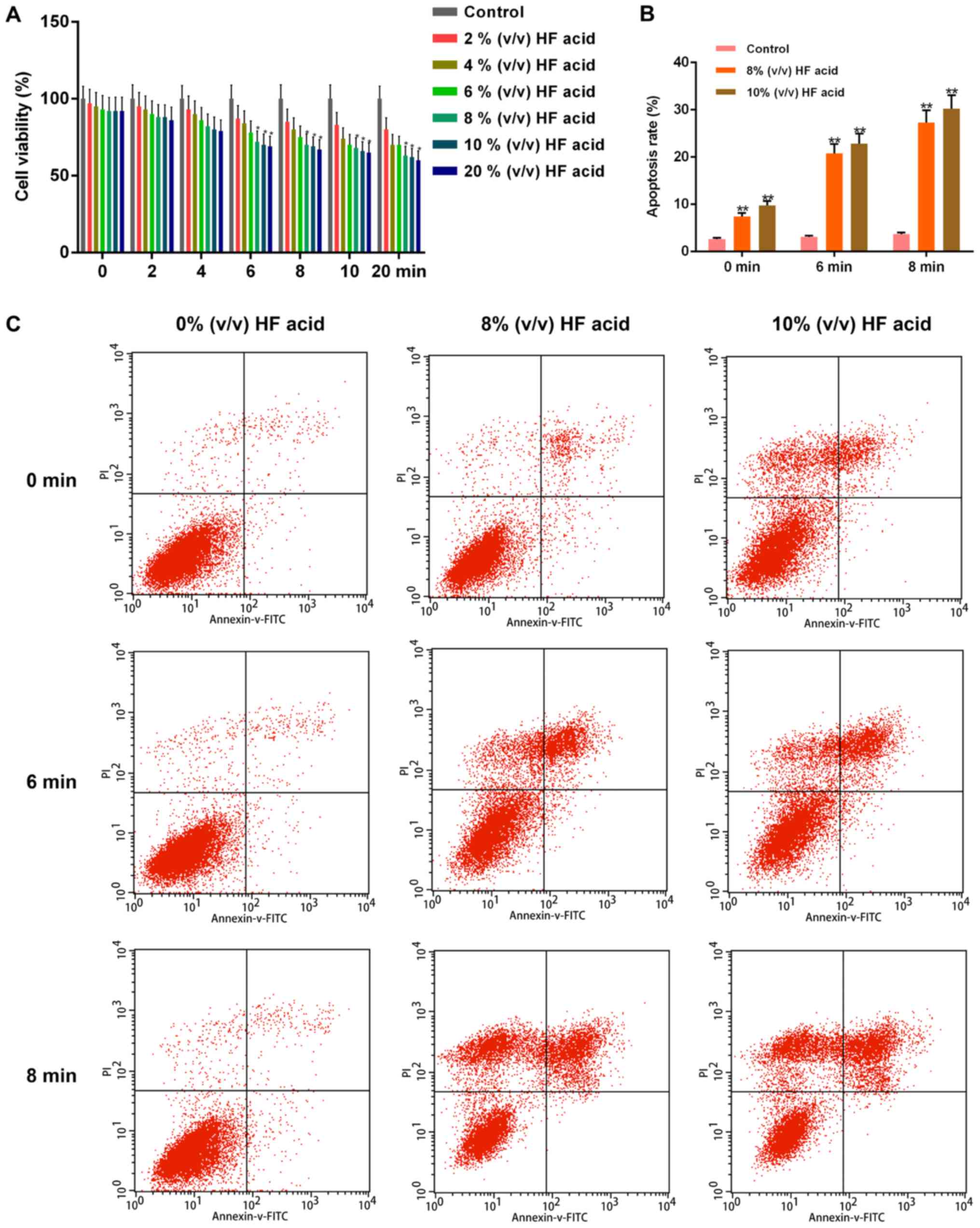 | Figure 2.Analysis of cell viability and
apoptosis of HDFs treated with HF. (A) Cell viability of HDFs were
detected by Cell Counting Kit-8 assay when treated with HF acid of
different concentrations [0, 2, 4, 6, 8, 10 and 20% (v/v)] for
determined times (0, 2, 4, 6, 8, 10 and 20 min), in a dose- and
time-dependent manners. (B) Apoptosis of HDFs was promoted when
treated with HF acid of different concentrations [0, 2, 4, 6, 8, 10
and 20% (v/v)] for specific times (0, 2, 4, 6, 8, 10 and 20 min) in
a dose- and time-dependent manners, as determined by (C) flow
cytometry. Data were presented as the mean ± standard deviation
(n=6/group). *P<0.05 and **P<0.01 vs. control. HDFs, human
dermal fibroblasts; HF, hydrofluoric; FITC, fluorescein
isothiocyanate; PI, propidium iodide. |
We performed Annexin V/PI double-staining to
quantify cellular apoptosis, and used flow cytometry for analysis
(Fig. 2B and C). Apoptosis increased
as the level of phosphatidylserine in the outer leaflet of the
plasma membrane increased, being detected by Annexin V binding to
the surface of the cells. The apoptotic rate of HF-acid-treated
HDFs increased significantly, about 3-fold of control. The
treatment of 8% (v/v) HF acid for 6 min was subjected to subsequent
investigations.
Function of CG on cell viability,
apoptosis status and apoptosis-related factors of HDFs treated with
HF acid
The effect of CG on the viability of HF-acid-injured
HDFs was measured by the CCK8 assay, and the results were shown in
Fig. 3A. Cell viability was
suppressed in 8% (v/v) HF-acid-treated HDFs (treatment duration: 6
min) without CG pre-treatment. When HDFs were pre-treated with CG,
cell viability increased in dose-dependent (50, 100 and 200 µmol/l)
and time-dependent (6, 12 and 24 h) manners, compared with HF
control. The statistical variation was considered significant when
cells were treated with 100 or 200 µmol/l CG for 12 or 24 h. CG
pre-treatment with different concentrations (50, 100, 200 µmol/l)
for 24 h was chosen to assess apoptosis, as well as for the
following detections. We performed Annexin V/PI double-staining to
quantify apoptosis, and used flow cytometry for the analysis
(Fig. 3B and C). The apoptotic rates
of HDFs decreased when pre-treated with CG for 24 h, prior to the 6
min-treatment with 8% (v/v) HF acid, in a dose-dependent (50, 100
and 200 µmol/l) manner.
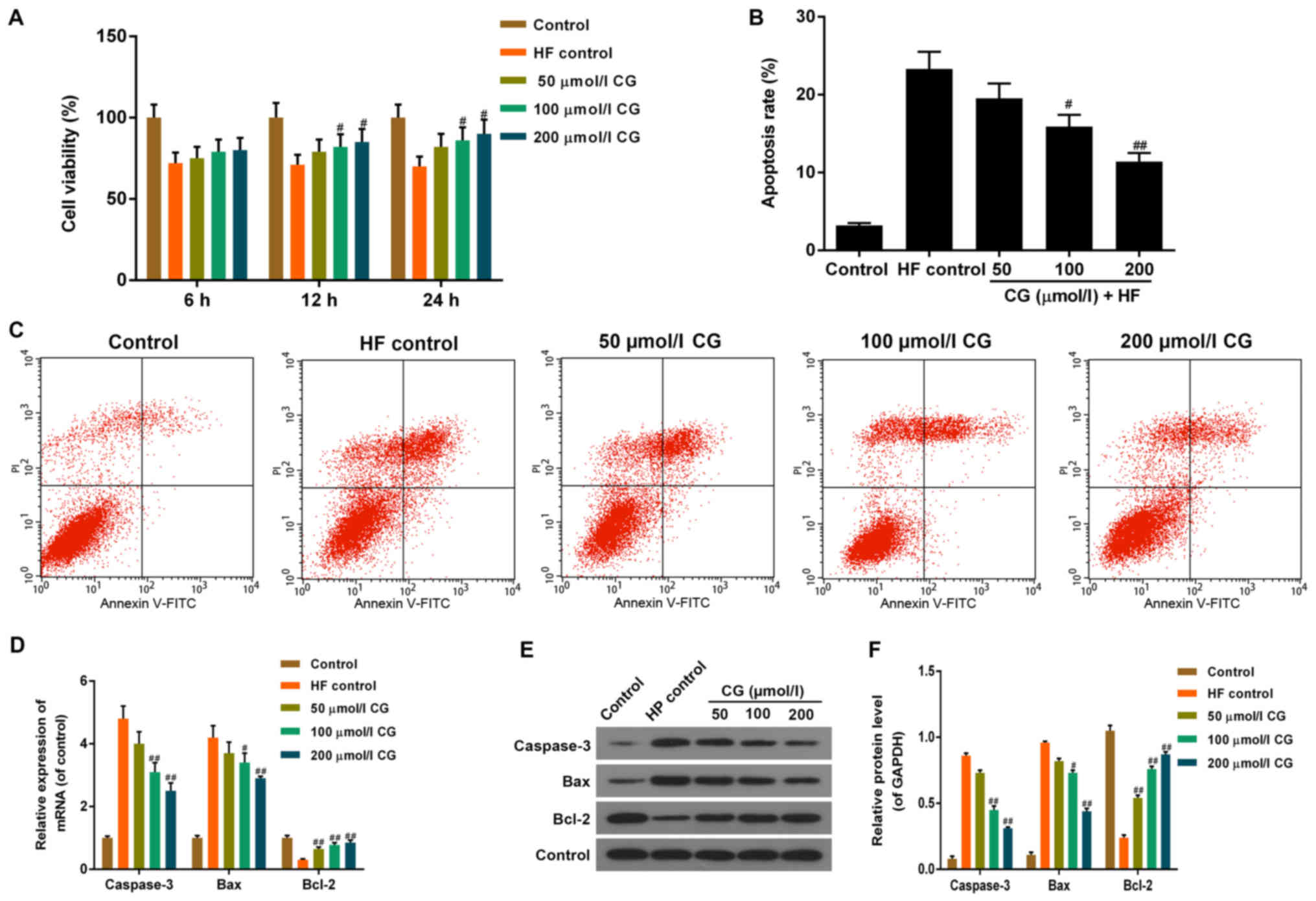 | Figure 3.Function of CG on cell viability and
apoptosis of HDFs treated with HF. (A) Compared with HDFs treated
with HF acid (HF control), the cell viability of HDFs was elevated
when pre-treated with CG of different concentrations (50, 100 and
200 µmol/l) for 6, 12 or 24 h respectively, then treated with 8%
(v/v) HF acid for 6 min, in a dose- and time-dependent manners, as
determined by Cell Counting Kit-8 assay. (B) Compared with HDFs
treated with HF acid, apoptosis of HDFs was reduced when
pre-treated with CG of different concentrations (50, 100 and 200
µmol/l) for 6, 12 or 24 h respectively, then treated with 8% (v/v)
HF acid for 6 min, in a dose- and time-dependent manner, (C) as
determined by flow cytometry. (D) The mRNA expression levels of
Caspase-3 and Bax were decreased, and the levels of Bcl-2 increased
following CG treatment, in dose-dependent manner, when compared
with HF control. (E and F) The protein levels of Caspase-3 and Bax
were decreased, and the levels of Bcl-2 increased following CG
treatment, in dose-dependent manner when compared with HF control.
Data were presented as mean ± standard deviation (n=6/group).
#P<0.05 and ##P<0.01 vs. HF control.
HDFs, human dermal fibroblasts; HF, hydrofluoric; CG, calcium
gluconate; Bcl-2, B-cell lymphoma 2; Bax, Bcl-2-assocaited X
protein; FITC, fluorescein isothiocyanate; PI, propidium
iodide. |
RT-qPCR and western blotting were performed to
detect mRNA and protein expression levels of important
apoptosis-related factors, such as Caspase-3, Bcl-2 associated X
protein (Bax) and B-cell lymphoma (Bcl)-2 (Fig. 3D-F). The mRNA and protein levels of
Caspase-3 and Bax increased, whereas the level of Bcl-2 decreased
significantly in HF-acid-treated cells, compared with control. When
pre-treated with CG, the mRNA and protein levels of Caspase-3 and
Bax decreased, and the level of Bcl-2 was restored in a
dose-dependent manner, compared with HF control.
Function of CG on cell division and
fiber-hyperplasia-related factors of HDFs treated with HF acid
The critical repair function of CG on
HF-acid-induced cell viability suppression and apoptosis promotion
in HDFs, prompted us to further explore the potential underlying
mechanism by measuring the relevant genes and proteins.
ELISA and Western blot assays showed that, protein
levels of matrix metalloproteinase (MMP)-1 and basic fibroblast
growth factor (bFGF) decreased significantly in HF control,
compared with the control group. When pre-treated with CG, protein
levels of MMP-1 and bFGF were restored in a dose-dependent manner,
compared with HF control (Fig. 4A-C).
ELISA was performed to detect the level of secretory protein
carboxyterminal propeptide of type I collagen (CICP) (Fig. 4D). The level of CICP increased
remarkably in HF control, compared with the control group. When
pre-treated with CG, the protein level of CICP decreased in a
dose-dependent manner, compared with HF control. As detected by the
colorimetric method, the protein level of L-HYP decreased
significantly in HF control, compared with the control group. When
pre-treated with CG, the level of L-HYP was restored in a
dose-dependent manner, compared with HF control (Fig. 4E).
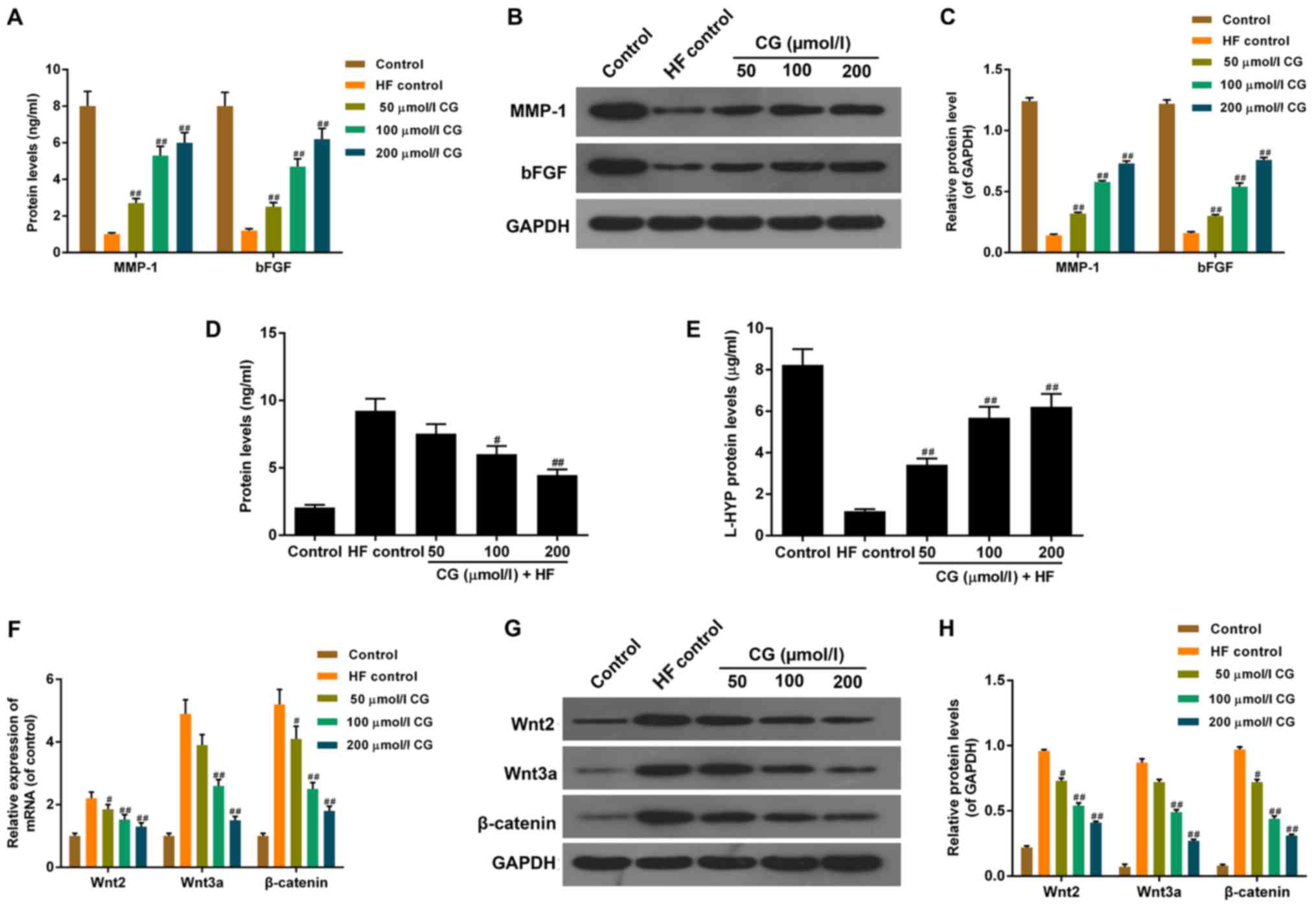 | Figure 4.Function of CG on cell division and
fiber hyperplasia associated factors in HDFs treated with HF. (A)
ELISA assay revealed that when pre-treated with CG, protein levels
of MMP-1 and bFGF increased in a dose-dependent manner when
compared with HF control. (B) Western blot confirmed that, (C) when
pre-treated with CG, the protein levels of MMP-1 and bFGF increased
in a dose-dependent manner when compared with HF control. (D) ELISA
assay showed that protein levels of carboxyterminal propeptide of
type I collagen in CG pre-treated HDFs decreased in a
dose-dependent manner, compared with HF control. (E) Colorimetric
method demonstrated that protein levels of L-HYP increased in a
dose-dependent manner compared with HF control. (F) Reverse
transcription-quantitative polymerase chain reaction showed that
mRNA expression levels of Wnt2, Wnt3a and β-catenin decreased in a
dose-dependent manner compared with HF control. (G) Western blot
showed that (H) protein levels of Wnt2, Wnt3a and β-catenin
decreased in a dose-dependent manner when compared with HF control.
Data were presented as mean ± standard deviation (n=6/group).
#P<0.05 and ##P<0.01 vs. HF control.
HDFs, human dermal fibroblasts; HF, hydrofluoric; CG, calcium
gluconate; MMP, matrix metalloproteinase; bFGF, basic fibroblast
growth factor; L-HYP, L-Hydroxyproline. |
RT-PCR and Western-blotting were performed to detect
the mRNA and protein expressions of important factors in the Wnt
pathway, such as Wnt2, Wnt3a and β-catenin (Fig. 4F-H). The mRNA and protein levels of
Wnt2, Wnt3a and β-catenin increased notably in HF control, compared
with the control group. When pre-treated with CG, the mRNA and
protein levels of Wnt2, Wnt3a and β-catenin decreased in a
dose-dependent manner, compared with HF control.
Discussion
HF acid is a highly dangerous, caustic, inorganic
acid, which can cause severe cutaneous burns, systemic toxicity and
occasionally lethal toxicity. CG has been proven to be a more
effective therapy for dermal and systemic injuries. Nevertheless,
the underlying molecular mechanism is yet to be elucidated.
The primary objective of this study was to elucidate
the major molecular mechanism behind the restorative effect of CG
on HF-acid-injured HDFs. In the present study, the classical
morphology of HDFs was observed under an optical microscope,
visible as oval or applanate-star-shaped, with plump cytoplasm and
clear nucleolus; as the cellular density increased HDFs became
spiral- or fence-shaped. Vimentin is a member of the intermediate
filament protein family. As the main component of the cytoskeleton,
it is expressed in cells derived from mesoblastema, particularly
HDFs. Therefore, we further assessed the morphology of HDFs by
detecting the presence of vimentin around the nucleus, using an
immunofluorescent green dye.
HF acid burns are notorious for causing progressive
tissue necrosis and severe pain. Apoptosis is a process of
programmed cell death that occurs in multicellular organisms,
important in maintaining the metabolic balance. As an important
component of granulation tissue, new vessels play key roles in
wound healing, providing the necessary nutrients for the migration
and proliferation of HDFs.
Our study showed that CG could promote cell
viability and decrease apoptotic rates of HDFs, which are destroyed
by HF acid resulting in skin injury, and thus support wound
healing. Furthermore, we studied the expression variations of
critical factors in cell proliferation and apoptosis, such as
Caspase-3, Bcl-2 and Bax, showing consistent changes. Caspase-3 is
an important member of the caspase family (cysteinyl aspartate
specific proteinases), as a conjunct activating factor in apoptosis
signal transduction, which directly participates in cell
regulation, signal transduction and late apoptosis. Bcl-2 activates
the caspase pathway, facilitates cell mitosis and inhibits
apoptosis by dissolving protein structures, resulting in tumor
proliferation (9). In contrast, Bax
can facilitate apoptosis by forming a heterodimer with Bcl-2,
essentially deterring its activity. When pre-treated with CG, mRNA
and protein levels of Caspase-3 and Bax decreased, whereas thelevel
of Bcl-2 increased in dose-dependent manners, compared with
HF-acid-injured HDFs. This suggests that the restorative function
of CG on HF-acid-induced cell viability suppression and apoptosis
promotion of HDFs is related to the molecular mechanism responsible
for apoptosis.
HF acid can penetrate the skin and destruct deep
tissue layers, as well as the bone. Bone metabolic disorder is
considered a common phenomenon in fluorosis and the main
characteristic of fluorosis of the bone. CICP, the carboxyl
terminal cut from type I collagen (Col-I) before fiber formation,
is the characteristic of Col-I and bone development, for Col-I is
the main collagen of the bone scaffold (10). In addition, L-HYP is considered the
characteristic amino acid of collagen. Being the main residue of
collagen proteins degradation, the content of L-HYP is constant,
accounting for more than 10% of collagen amino acids. Therefore,
L-HYP levels could reflect collagen synthesis and HDF status.
MMP-1, secreted by HDFs, is a critical enzyme regulating collagen
degradation, especially Col-I (11).
Self-renewal and damage repair of skin tissues are related to the
metabolic proliferation and directional differentiation of
epidermal stem cells. Cytokines like bFGF, a kind of
heparin-combined polypeptide mitogen, are critical factorspromoting
cell division and proliferation, while regulating cell
differentiation and immunology. Previous studies have even
identified a connection between bFGF and wound healing (11). Our study showed that CG could inhibit
the expression of CICP and promote the expression of L-HYP, MMP-1
and bFGF, as these were increased (CICP) and decreased (L-HYP,
MMP-1 and bFGF), respectively, in HF-acid-treated HDFs. This
strongly suggests that CG could influence cell division or fiber
hyperplasia. Therefore, CG may attenuate the toxic effect of HF
acid on HDFs by regulating the expression of cell division or fiber
hyperplasia-related factors.
Wnt was first discovered in mouse breast cancer
epithelial cells. Though most research on the Wnt pathway is
concentrated on the regulation of stem cell differentiation, organ
development and tumorigenesis (12),
the Wnt pathway also participates in wound healing after skin
injury, progression of fibrotic diseases and activation of fibrotic
effector cells. The related specific function and molecular
mechanism may depend on the type of tissue or organ. Research on
the involvement of the Wnt pathway in wound healing and fibrosis
has been increasing in recent years. Labus et al (13) and Okuse et al (14) found that the Wnt pathway is activated
in the early stage of wound healing, and Konigshoff et al
discovered activated Wnt pathway in pulmonary fibrosis (15,16). There
is crosstalk between the Wnt pathway and the TGF-β/Smad pathway, as
β-catenin is a scaffold protein linking the cytoplasmic tail of
classical cadherins of the endothelium to the actin cytoskeleton in
the Wnt downstream pathway, and its down regulation can give rise
to tumor metastasis and fibrotic diseases (17,18). Wnt2
and Wnt3a both induce abundant cellular changes in several key
signaling molecules, including β-catenin, in the canonical
Wnt/β-catenin pathway. Our study showed that CG could inhibit the
expression of Wnt2, Wnt3a and β-catenin, which were increased in
the HF-acid-treated HDFs. This indicates that CG attenuated the
toxic effect of HF acid on HDFs by regulating the Wnt/β-catenin
pathway. However, as Caspase-3, Bax and Bcl-2, as well as Wnt2,
Wnt3a and β-catenin are classical pathways that can be affected by
many factors, it may be that more specific mechanisms are involved.
Similarly, there are specific inhibitors for both the Wnt pathway
and cell apoptosis. We will perform more blocking experiments to
assess the specificity of these effects and pathways, in a future
study.
Collectively, these data suggest that CG attenuated
the toxic effect of HF acid on HDFs by promoting cell viability,
while inhibiting apoptosis and related factors. Moreover, CG may
regulate cell division or fiber hyperplasia-related factors, and
also the Wnt/β-catenin pathway. These findings provide novel
evidence in favor of utilizing CG in the treatment of
HF-acid-induced wounds.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
JP designed the research and treated cells. RL, LP
and HJ performed the other experiments and drafted the manuscript.
All authors have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hojer J, Personne M, Hulten P and Ludwigs
U: Topical treatments for hydrofluoric acid burns: A blind
controlled experimental study. J Toxicol Clin Toxicol. 40:861–866.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shamis Y, Hewitt KJ, Carlson MW,
Margvelashvilli M, Dong S, Kuo CK, Daheron L, Egles C and Garlick
JA: Fibroblasts derived from human embryonic stem cells direct
development and repair of 3D human skin equivalents. Stem Cell Res
Ther. 2:102011. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Andriani F, Margulis A, Lin N, Griffey S
and Garlick JA: Analysis of microenvironmental factors contributing
to basement membrane assembly and normalized epidermal phenotype. J
Invest Dermatol. 120:923–931. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Smola H, Thiekötter G and Fusenig NE:
Mutual induction of growth factor gene expression by
epidermal-dermal cell interaction. J Cell Biol. 122:417–429. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chmielowiec J, Borowiak M, Morkel M,
Stradal T, Munz B, Werner S, Wehland J, Birchmeier C and Birchmeier
W: c-Met is essential for wound healing in the skin. J Cell Biol.
177:151–162. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Matsumoto K and Nakamura T: Hepatocyte
growth factor (HGF) as a tissue organizer for organogenesis and
regeneration. Biochem Biophys Res Commun. 239:639–644. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sorrell JM and Caplan AI: Fibroblast
heterogeneity: More than skin deep. J Cell Sci. 117:667–675. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sorrell JM, Baber MA and Caplan AI: Clonal
characterization of fibroblasts in the superficial layer of the
adult human dermis. Cell Tissue Res. 327:499–510. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rashed L, Gharib DM, Hussein RE, Tork O
and Abusree A: Combined effect of bone marrow derived mesenchymal
stem cells and nitric oxide inducer on injured gastric mucosa in a
rat model. Tissue Cell. 48:644–652. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Scalinci SZ, Scorolli L, Meduri A, Grenga
PL, Corradetti G and Metrangolo C: Effect of basic fibroblast
growth factor and cytochrome c peroxidase combination in transgenic
mice corneal epithelial healing process after excimer laser
photoablation. Clin Ophthalmol. 5:215–221. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gallego-Munoz P, Ibares-Frias L,
Valsero-Blanco MC, Cantalapiedra-Rodriguez R, Merayo-Lloves J and
Martinez-Garcia MC: Effects of TGFβ1, PDGF-BB, and bFGF, on human
corneal fibroblasts proliferation and differentiation during
stromal repair. Cytokine. 96:94–101. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sokol SY: Maintaining embryonic stem cell
pluripotency with Wnt signaling. Development. 138:4341–4350. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Labus MB, Stirk CM, Thompson WD and Melvin
WT: Expression of Wnt genes in early wound healing. Wound Repair
Regen. 6:58–64. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Okuse T, Chiba T, Katsuumi I and Imai K:
Differential expression and localization of WNTs in an animal model
of skin wound healing. Wound Repair Regen. 13:491–497. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Konigshoff M, Balsara N, Pfaff EM, Kramer
M, Chrobak I, Seeger W and Eickelberg O: Functional Wnt signaling
is increased in idiopathic pulmonary fibrosis. PLoS One.
3:e21422008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Konigshoff M, Kramer M, Balsara N, Wilhelm
J, Amarie OV, Jahn A, Rose F, Fink L, Seeger W, Schaefer L, et al:
WNT1-inducible signaling protein-1 mediates pulmonary fibrosis in
mice and is upregulated in humans with idiopathic pulmonary
fibrosis. J Clin Invest. 119:772–787. 2009.PubMed/NCBI
|
|
17
|
Cheon S, Poon R, Yu C, Khoury M, Shenker
R, Fish J and Alman BA: Prolonged beta-catenin stabilization and
tcf-dependent transcriptional activation in hyperplastic cutaneous
wounds. Lab Invest. 85:416–425. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dômont J, Salas S, Lacroix L, Brouste V,
Saulnier P, Terrier P, Ranchère D, Neuville A, Leroux A, Guillou L,
et al: High frequency of beta-catenin heterozygous mutations in
extra-abdominal fibromatosis: A potential molecular tool for
disease management. Br J Cancer. 102:1032–1036. 2010. View Article : Google Scholar : PubMed/NCBI
|