Introduction
Non-small cell lung cancer (NSCLC) accounts for 85%
of lung cancer cases. It is one of the most common malignant tumors
worldwide and is the leading cause of cancer death (1). Chemotherapy is a widely used treatment
method for NSCLC in clinic, however, drug resistance usually
develops during the course of therapy and limits the efficacy of
chemotherapy. Cancer stem cell (CSC) is a fraction of stem-like
cells in tumor, and has been considered as the root of tumor
growth, relapse and metastasis. Different from other cancer cells,
CSC demonstrates exclusive ability of self-renewal and tumor
formation (2,3). Moreover, CSC has also been identified
with impaired apoptosis, enhanced DNA damage response and
overexpression of multidrug resistance protein and membrane
transporter, which further drive tumor progression and recurrence
after chemotherapy (4). Given these,
CSC represents a promising tool for investigating cancer drug
response and developing novel approaches to overcome
chemo-resistance.
Yes-associated protein (YAP) is a core effector of
the Hippo tumor-suppressor pathway. Recently, YAP has been
implicated in chemoresistance in a variety of cancer cells. It has
been shown that silencing of YAP1 increased cell sensitivity to
anti-tumor drugs in multiple cancers, including ovarian CSC
(5), esophageal cancer (6), pancreatic cancer (7), liver cancer (8), oral squamous cell carcinoma (9) and gastric CSC (10). YAP functions as a transcriptional
coactivator. Overexpression of YAP has been associated with
induction of genes involved in cell proliferation, apoptosis
suppression, epithelial-mesenchymal transition and
anchorage-independent growth. The activated YAP function has been
found to mediate protease-activated receptor 1 (PAR1)-induced tumor
initiation and spheroid colony formation in gastric cancer cells,
and is also responsible for the acquired resistance to cisplatin
(CDDP), 5-FU and paclitaxel in gastric CSCs (10). In addition, via upregulating the
expression of glycogen synthesis kinase 3A (GSK3A) and ATP-binding
cassette subfamily B member 1 (ABCB1). It has also been shown that
YAP could enhance chemotherapy resistance of ovarian CSCs to CDDP,
paclitaxel and (5).
All these evidences supported the role of YAP as a
drug resistance mediator and implied the potential of YAP to be a
novel target for cancer therapy. However, little information is
available regarding the relationship between YAP and drug response
in lung cancer especially in lung CSCs (LCSCs). CDDP is the first
line chemotherapy drug for lung cancer. Since YAP was found to be
responsible for CDDP resistance in gastric and ovarian CSCs, we
hypothesized that YAP may likewise play a role in regulating
cellular response to CDDP in LCSCs. Previously, we showed that A549
lung cancer cells could form tumor spheres with LCSC properties and
enhanced drug resistance (11). In
this study, we explored YAP expression in LCSCs, and inhibited YAP
expression by siRNA to observe the proliferation inhibition and
apoptosis of LCSCs. We presumed that inhibiting YAP is a new
therapeutic strategy for human lung cancer though restraining the
chemotherapy resistance of LCSCs.
Materials and methods
Reagents and antibodies
Cell culture media Dulbecco's modified Eagle's
medium (DMEM), DMEM/F-12 and fetal bovine serum (FBS) were obtained
from Biological Industries (Cromwell, CT, USA). Trypsin and EDTA
were obtained from Solarbio (Beijing, China), B27 was purchased
from Gibco (Thermo Fisher Scientific, Inc., Waltham, MA, USA), EGF
and bFGF were obtained from Proteintech (Rosemont, IL, USA). YAP
rabbit monoclonal antibody was obtained from Cell Signaling
Technology, Inc. (Danvers, MA, USA), β-actin mouse monoclonal
antibody was purchased from Proteintech. The Cell Counting Kit-8
(CCK-8), TransZol Up Plus RNA kit, Transcript One-Step gDNA Removal
and cDNA Synthesis Supermix, Goat Anti-Mouse/Rabbit IgG (H&L)
were purchased from TransGen Biotech (Beijing, China). Annexin
V-FITC apoptosis detection kit was obtained from BD Biosciences
(San Diego, CA, USA). Western blot reagents were obtained from
Bio-Rad Laboratories, Inc. (Hercules, CA, USA), and Pierce BCA
Protein Assay kit was purchased from Applygen Technologies Inc.
(Beijing, China). Small interfering RNA oligos were purchased from
Invitrogen (Thermo Fisher Scientific, Inc.) and
Lipofectamine® 2000 reagent purchased from TransGen
(Beijing, China).
Cell line
The human NSCLC cell line A549, obtained from the
Second Affiliated Hospital of Nanchang University, was cultured in
DMEM supplemented with 10% FBS (v/v), streptomycin and penicillin
and incubated at 37°C under a humid atmosphere with 5%
CO2. A549 cell spheres were generated using the method
as previously described (11).
Briefly, A549 cells were seeded in the 6-well plates at the density
of 1,000–1,500 cells per well and cultured in 2 ml 10% FBS
supplemented DMEM for 8–10 days. Different single-cell derived
clones formed after 8–10 days, including holoclone, meroclones and
paraclones, of which holoclone consisted of tightly packed cells
and demonstrated stem cell properties, such as continuous passage
and unlimited proliferation. The holoclones were collected and
cultured in serum-free media DMEM/F-12 supplemented with bFGF, EGF,
insulin and B27. Primary A549 cell spheres were shaped after 10–14
days and were continuously grown in serum-free media to obtain
secondary A549 cell spheres.
Transfection with siRNA for YAP
A549 cells were cultured in DMEM supplemented with
10% FBS, while A549 cell spheres were cultured in serum-free
DMEM/F-12 supplemented with bFGF, EGF, insulin and B27. After 24 h,
small interfering RNA targeting YAP (siYAP:
5′-GGUCAGAGAUACUUCUUAATT-3′) and negative control (siNC:
5′-UUCUCCGAACGUGUCACGUTT-3′) were introduced into cells using
Lipofectamine® 2000 reagent. CDDP was added after two
days at a concentration of 2.5 µg/ml.
Total RNA isolation and reverse
transcription-PCR
Total RNA of A549 cells and A549 cell spheres under
different treatment was extracted using TransZol Up Plus RNA kit
following the manufacturer's instructions and then the RNA
concentration were measured through ultraviolet spectrophotometer.
cDNA synthesis was in progress with 1 µg of RNA using Transcript
One-Step gDNA Removal and cDNA Synthesis Supermix according to the
manufacturer's instructions. PCR was performed for 35 cycles using
the following temperature profiles 94°C for 30 sec, 55°C for 30
sec, and 72°C for 60 sec. β-actin was served as an internal
control. The sequence of primers includes: YAP_F:
5′-TGACCCTCGTTTTGCCATGA-3′, YAP_R: 5′-GTTGCTGCTGGTTGGAGTTG-3′.
ABCB1_F: 5′-GTCTGGACAAGCACTGAAA-3′, ABCB1_R:
5′-AACAACGGTTCGGAAGTTT-3′. β-actin: F: 5′-CACGGCATCGTCACCAACT-3′,
R: 5′-GTCCTACGGAAAACGGCAGA-3.
Western blot analysis
Total protein of A549 cells and A549 cell spheres
under different treatments was extracted using RIPA lysis buffer
and protein concentration was determined by BCA method. Then
proteins were denatured in 5× SDS-PAGE protein loading buffer,
separated by 10% SDS-PAGE and transferred onto polyvinylidene
difluoride (PVDF) membranes. The membranes were incubated with
antibody recognizing YAP (1:500) and β-actin (1:2,000), and protein
bands were detected with the ECL detection system and image lab
software.
Assessment of proliferation by
CCK-8
10% CCK was added into the culture media of A549
cells and A549 cell spheres under different treatments. After
incubation for another 4 h, cell culture absorbance at 450 nm was
read by microplate reader.
Annexin V-FITC staining
A549 cells and A549 cell spheres under different
treatments were stained with Annexin V-FITC/PI following
manufacturer's instructions. And apoptotic cells were analyzed
determined by BD flow cytometer.
Statistical analysis
Image J software was used for data quantification
and GraphPad Prism 6 software was used for the statistical
analysis. Mean ± SD of three replicates are shown. An unpaired
two-tailed Student's t test was used to compare the means of two
groups. Two-way analysis of variance (ANOVA) was used to analyze
the differences among group means, followed by a
Student-Newman-Keuls (SNK) test to compare the differences between
different groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
Generation of A549 tumor spheres
Previously, we and others showed that adherent lung
cancer A549 cells cultured in serum-free medium could form tumor
spheres enriched for stem-like cells (11,12).
Notably, A549 tumor spheres exhibited enhanced proliferation,
cell-cycle progression as well as drug-resistant properties vs.
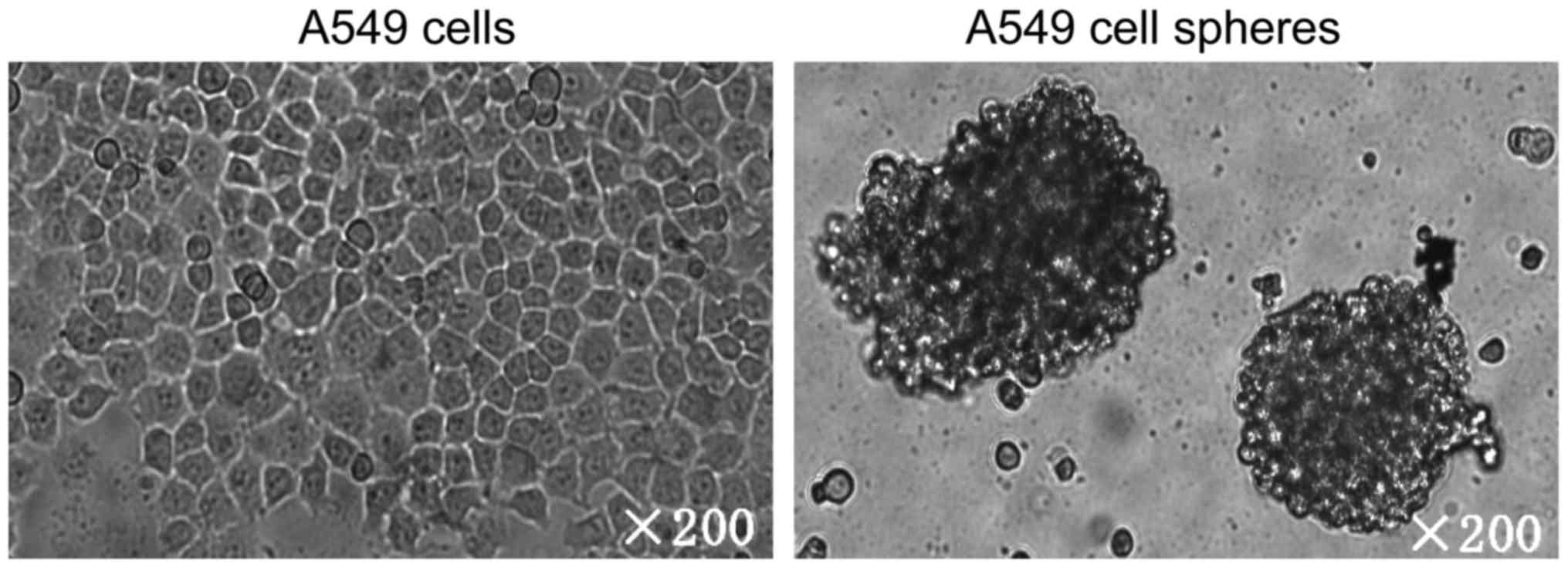
A549 adherent cells. The morphologies of adherent A549 cells and
tumor spheres are shown in Fig. 1.
Specifically, A549 cells formed three morphologically different
colonies: holoclone, meroclone and paraclone by using single-cell
cloning culture. Only holoclones were selected, digested and
further incubated with serum-free media. After three to four weeks,
highly clustered tumor cell spheres were formed (Fig. 1).
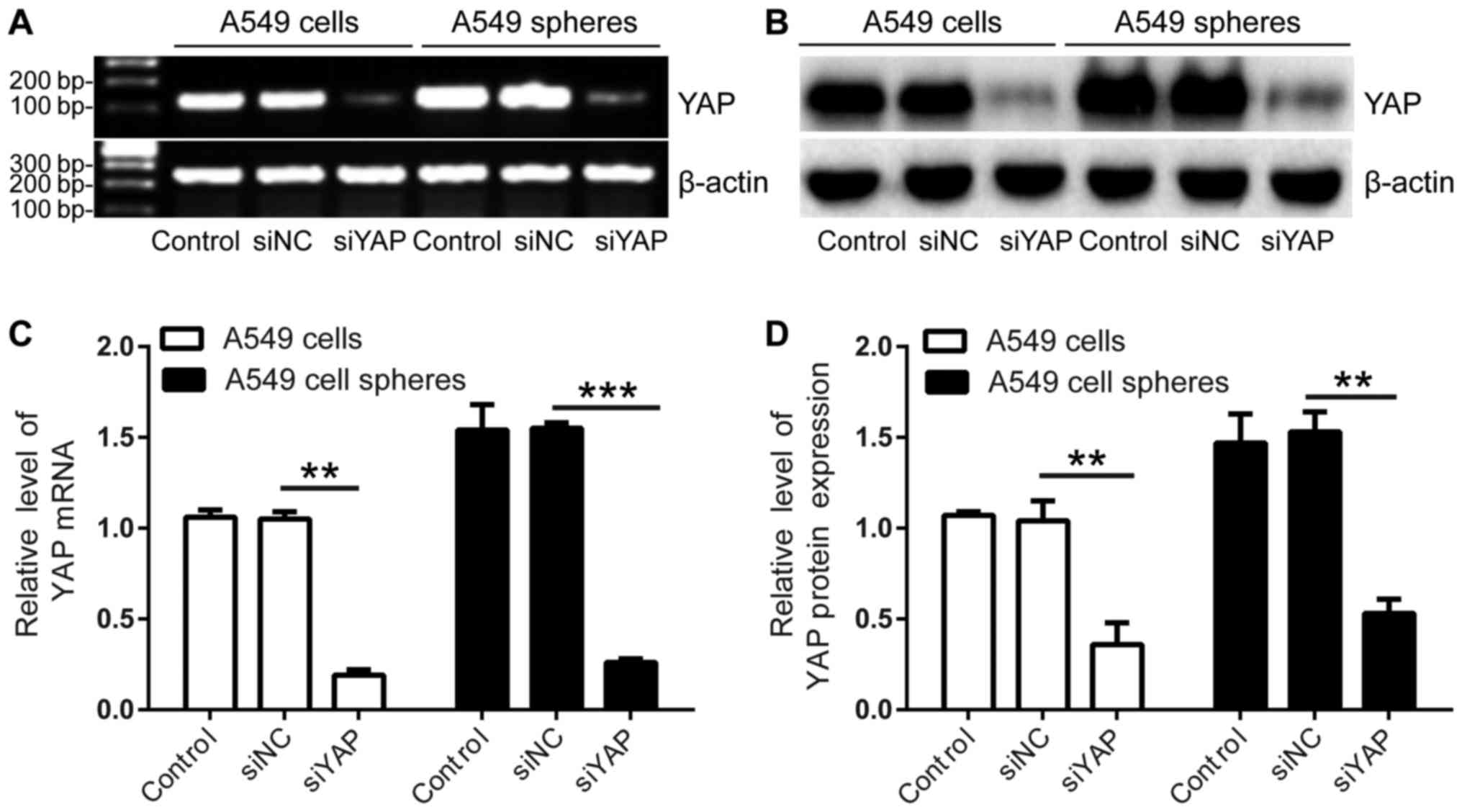
Elevated YAP expression in A549 cell
spheres
Our previously studies revealed some stem cell-like
properties of A549 cell spheres, including elevated expression of
various stem cell markers (e.g., Sca-1, CD133, CD44s, Oct4, Sox3,
Nanog) and the capability of multilineage differentiation (11). Interestingly, here our results showed
that expression of YAP was likewise enhanced in A549 tumor spheres
as compared with adherent A549 culture at both mRNA and protein
levels (Fig. 2A and B), supporting
the association of YAP with the stemness of A549 cell spheres.
Notably, YAP expression was effectively knockdown using siRNA, thus
allowing further investigation of YAP function in these cells
(Fig. 2A-D).
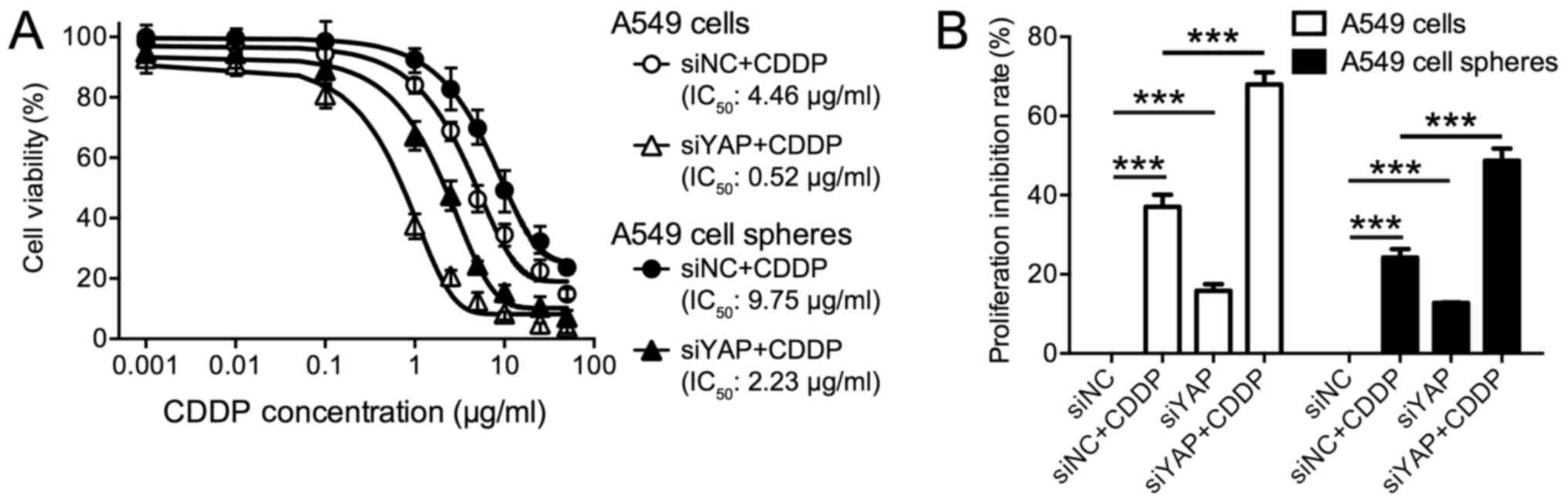
Knockdown of YAP resensitized A549
cells and A549 spheres proliferation to CDDP
YAP overexpression has been implicated in the drug
resistance of various CSCs. Here, we tested if YAP was associated
with lung cancer cell resistance to CDDP. We treated A549 cells and
A549 tumor spheres with increasing dosage of CDDP ranging from
0.001 to 50 µg/ml for 48 h. Cells were transfected with siYAP or
siNC as a control. As a minor toxicity was observed by using the
transfection reagent alone, here we didn't include non-transfected
cells as a control. As shown in Fig.
3A, in siNC treated cells, the half maximal inhibitory
concentration (IC50) of CDDP was determined as 4.46
µg/ml for A549 cells and as 9.75 µg/ml for A549 spheres. The
proliferation of 37.04% (±3.06%) A549 cells and 24.14% (±2.16%)
A549 cell spheres were inhibited by 2.5 µg/ml CDDP (Fig. 3B), indicating the poor efficacy of
CDDP on lung cancer cells. Notably, as compared with adherent
cells, A549 cell spheres demonstrated higher IC50 of
CDDP and lower inhibitory rate, suggesting increased drug
resistance in tumor spheres (Fig. 3A and
B). Of note, upon YAP knockdown, CDDP toxicity was
significantly enhanced in these cells. The IC50 values
of CDDP was reduced to 0.52 µg/ml A549 cells and 2.23 µg/ml for
A549 spheres after YAP knockdown (Fig.
3A). Treatment with 2.5 µg/ml CDDP resulted in a significant
inhibition of cancer cell proliferation in both A549 cells and A549
cell spheres by (Fig. 3B, all
P<0.001). Moreover, a strong synergistic effect was observed
when CDDP was combined with YAP knockdown (Fig. 3B, all P<0.001). All these evidences
implied the potential of YAP silencing as an adjuvant therapy to
chemotherapy.
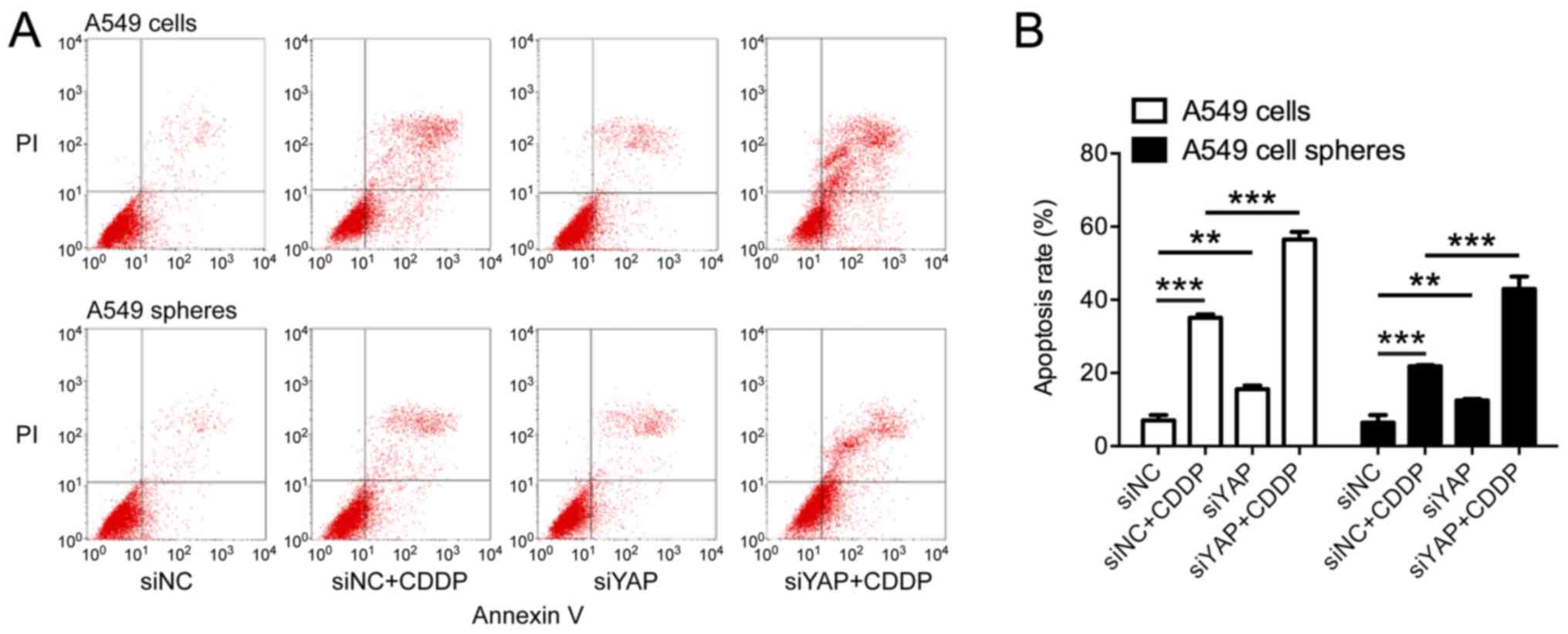
Knockdown of YAP induced
apoptosis
Since YAP is responsible for apoptosis suppression,
we then investigated the apoptosis of lung cancer cells on YAP
depletion. As shown in Fig. 4, in
both A549 cells and A549 spheres, more apoptotic cells were
observed on YAP knockdown, suggesting the activation of apoptotic
cascades. Moreover, we also found that CDDP induced apoptosis in
35.08% (±0.96%) A549 cells and 21.84% (±0.30%) A549 spheres,
whereas much more cells were undergoing apoptosis on the
co-treatment with siYAP, 56.50% (±2.07%) in A549 cells and 35.08%
(±3.38%) in A549 spheres (Fig. 4B,
all P<0.01), suggesting that YAP knockdown may enhance CDDP
efficacy by promoting apoptosis.
Furthermore, we performed realtime PCR assay to
examine the influence of YAP knockdown on ABCB1, which is a
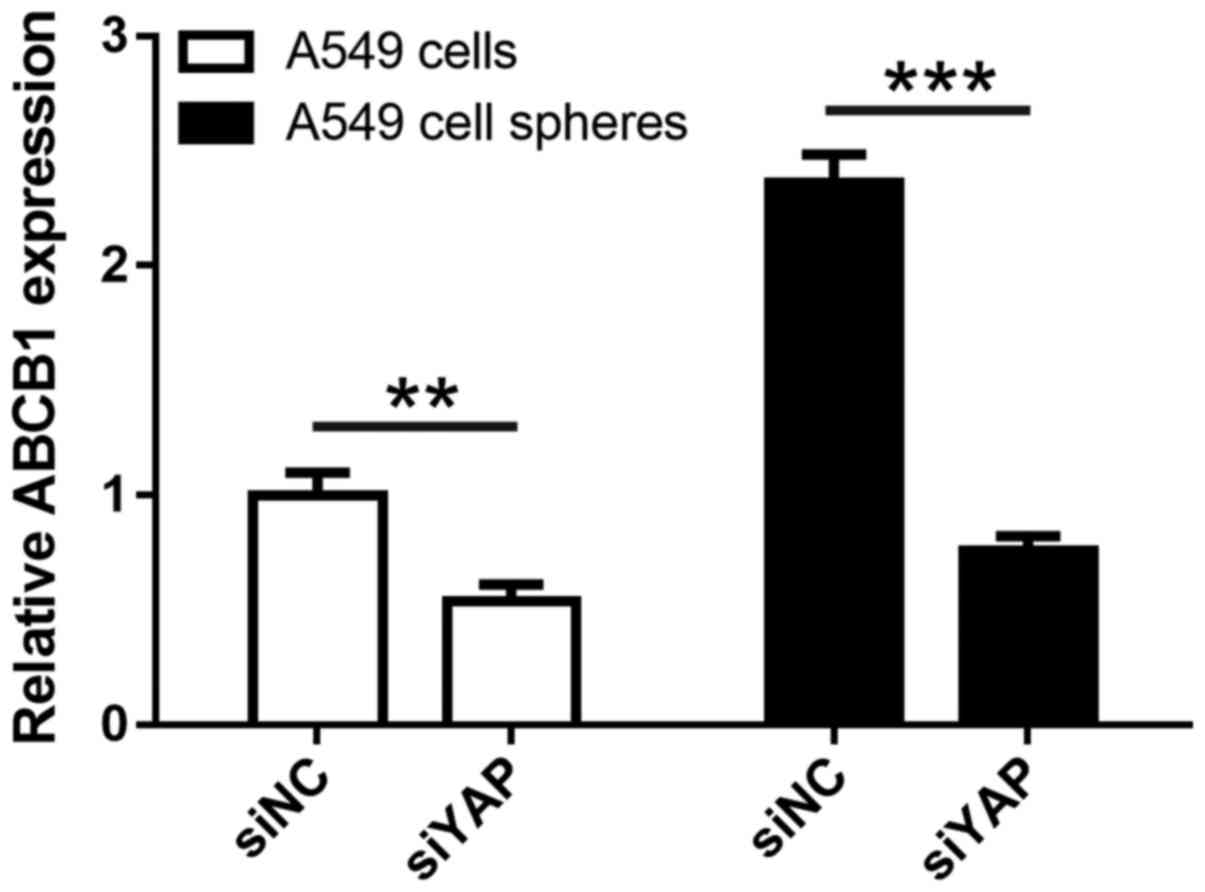
glycoprotein involved in multidrug resistance. As shown in Fig. 5, ABCB1 expression was significantly
enhanced (P<0.01) in tumor spheres vs. adherent A549 cells,
consistent with upregulation of YAP. Moreover, YAP silencing
resulted in significant reduction of ABCB1 expression in both
adherent A549 and tumor spheres (all P<0.01), indicative of a
direct role of YAP in transcriptional regulation of ABCB1
expression in lung cancer cells.
Discussion
According to our previous study, A549 cell spheres
were characterized with stem cell properties, including elevated
expression of stem cell makers and the potential of self-renewal
and multilineage differentiation (11). In the current study, we also found
that A549 cell spheres demonstrated increased resistance to CDDP
treatment, with enhanced cell proliferation and impaired apoptosis
as compared with adherent A549 cells. This was consistent with our
knowledge that CSCs are responsible for treatment failure and tumor
recurrence in cancer patients and also provided A549 spheres as
promising model to investigate drug response and develop novel
strategies to overcome chemoresistance in NSCLC. In the present
study, we found that YAP might be associated with the stemness and
chemoresistance of A549 tumor spheres, while knockdown of YAP
significantly enhanced sensitivity of A549 spheres to CDDP.
YAP is a major effector of Hippo tumor suppressor
pathway, which is implicated in organ size control and tissue
regeneration through regulating cell proliferation and apoptosis
(13). YAP carries out its function
by translocating into nuclear and inducing the transcription of
genes involved in proliferation and anti-apoptosis (14,15),
whereas the activation of Hippo pathway limits YAP function by
inducing YAP phosphorylation and impairing its nuclear
translocation (16,17). The major functions of the Hippo
pathway have been involved in regulation of cell proliferation,
differentiation, and migration in developing organs. High Hippo
signaling activity has been observed in many cancer types, and
functional dysregulation of Hippo signaling enhances the oncogenic
properties of YAP and promotes tumorigenesis. Given these, the
disruption of the balance between Hippo activity and YAP levels may
disturb tissue homeostasis, and lead to a variety of disorders
including cancers. Consistent with this, impaired Hippo pathway and
elevated YAP expression has been frequently observed in solid tumor
tissues, and YAP has been recognized as an oncogene, which is
essential for cancer initiation, progression, or metastasis
(18–25). Moreover, studies have also shown that
overexpression of YAP is closely related to some carcinogenic
properties of CSCs, such as unlimited self-renewal, the loss of
cell contact inhibition (14),
epithelial-mesenchymal transition (21,26) and
anchorage-independent growth (27),
which contribute to the development of drug resistance and result
in cancer relapse (10,28,29). In
the present study, we found that the expression of YAP was higher
in A549 cell spheres than A549 cells, which support a role of YAP
in maintaining certain characteristics of LCSCs.
It has been reported that YAP was responsible for
the overexpression of anti-apoptotic Bcl-xL in hepatocellular
carcinoma cells, which prevented the release of mitochondrial
contents and inhibited caspase activation (30). In addition, YAP was also found to
initiate chemoresistance in ovarian cancer cell via up-regulating
the expression of drug resistance genes ABCB1, ABCC1 and GSK3A
(5). In our study, we also provided
evidences that elevated YAP expression might be associated with
CDDP resistance in LCSCs, while YAP silencing increased CDDP
toxicity to LCSCs by subjecting more cancer cells to apoptosis. The
regulatory mechanism of YAP in lung cancer is still under
investigation. Nevertheless, although YAP knockdown demonstrated
the ability of restoring drug sensitivity, reduced CDDP efficacy
was still observed in A549 spheres as compared with A549 cells,
indicating higher basal line levels of YAP or existence of other
drug resistance mechanisms.
In conclusion, our study identified increased drug
resistance in LCSCs, which might be associated with the
overexpression of YAP. We also provided evidences that YAP
silencing could resensitize LCSCs to chemotherapy and could become
a promising adjuvant therapy for NSCLC patients.
Acknowledgments
Not applicable.
Funding
This work was supported by the National Natural
Science Foundation of China (grant no. 81660493), the Scientific
Research Foundation of the Education Department of Jiangxi Province
(grant no. GJJ14003) and the Natural Science Foundation of Jiangxi
Province (grant no. 20143ACB20011).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
JS, LXX and XQY developed and designed the study and
wrote the manuscript. JS, LXX, XYZ, PH and MFL performed the
experiments. FX and JH performed the statistical analyses. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
MacDonagh L, Gray SG, Breen E, Cuffe S,
Finn SP, O'Byrne KJ and Barr MP: Lung cancer stem cells: The root
of resistance. Cancer Lett. 372:147–156. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Leon G, MacDonagh L, Finn SP, Cuffe S and
Barr MP: Cancer stem cells in drug resistant lung cancer: Targeting
cell surface markers and signaling pathways. Pharmacol Ther.
158:71–90. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Signore M, Ricci-Vitiani L and De Maria R:
Targeting apoptosis pathways in cancer stem cells. Cancer Lett.
332:374–382. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xia Y, Zhang YL, Yu C, Chang T and Fan HY:
YAP/TEAD co-activator regulated pluripotency and chemoresistance in
ovarian cancer initiated cells. PLoS One. 9:e1095752014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Song S, Honjo S, Jin J, Chang SS, Scott
AW, Chen Q, Kalhor N, Correa AM, Hofstetter WL, Albarracin CT, et
al: The hippo coactivator YAP1 mediates EGFR overexpression and
confers chemoresistance in esophageal cancer. Clin Cancer Res.
21:2580–2590. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yuan Y, Li D, Li H, Wang L, Tian G and
Dong Y: YAP overexpression promotes the epithelial-mesenchymal
transition and chemoresistance in pancreatic cancer cells. Mol Med
Rep. 13:237–242. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dai XY, Zhuang LH, Wang DD, Zhou TY, Chang
LL, Gai RH, Zhu DF, Yang B, Zhu H and He QJ: Nuclear translocation
and activation of YAP by hypoxia contributes to the chemoresistance
of SN38 in hepatocellular carcinoma cells. Oncotarget. 7:6933–6947.
2016.PubMed/NCBI
|
|
9
|
Yoshikawa K, Noguchi K, Nakano Y, Yamamura
M, Takaoka K, Hashimoto-Tamaoki T and Kishimoto H: The Hippo
pathway transcriptional co-activator, YAP, confers resistance to
cisplatin in human oral squamous cell carcinoma. Int J Oncol.
46:2364–2370. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fujimoto D, Ueda Y, Hirono Y, Goi T and
Yamaguchi A: PAR1 participates in the ability of multidrug
resistance and tumorigenesis by controlling Hippo-YAP pathway.
Oncotarget. 6:34788–34799. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ye XQ, Li Q, Wang GH, Sun FF, Huang GJ,
Bian XW, Yu SC and Qian GS: Mitochondrial and energy
metabolism-related properties as novel indicators of lung cancer
stem cells. Int J Cancer. 129:820–831. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sun FF, Hu YH, Xiong LP, Tu XY, Zhao JH,
Chen SS, Song J and Ye XQ: Enhanced expression of stem cell markers
and drug resistance in sphere-forming non-small cell lung cancer
cells. Int J Clin Exp Pathol. 8:6287–6300. 2015.PubMed/NCBI
|
|
13
|
Johnson R and Halder G: The two faces of
Hippo: Targeting the Hippo pathway for regenerative medicine and
cancer treatment. Nat Rev Drug Discov. 13:63–79. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhao B, Wei X, Li W, Udan RS, Yang Q, Kim
J, Xie J, Ikenoue T, Yu J, Li L, et al: Inactivation of YAP
oncoprotein by the Hippo pathway is involved in cell contact
inhibition and tissue growth control. Genes Dev. 21:2747–2761.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhao B, Li L, Tumaneng K, Wang CY and Guan
KL: A coordinated phosphorylation by Lats and CK1 regulates YAP
stability through SCF(beta-TRCP). Genes Dev. 24:72–85. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chan EH, Nousiainen M, Chalamalasetty RB,
Schäfer A, Nigg EA and Silljé HH: The Ste20-like kinase Mst2
activates the human large tumor suppressor kinase Lats1. Oncogene.
24:2076–2086. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Praskova M, Xia F and Avruch J:
MOBKL1A/MOBKL1B phosphorylation by MST1 and MST2 inhibits cell
proliferation. Curr Biol. 18:311–321. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang Y, Dong Q, Zhang Q, Li Z, Wang E and
Qiu X: Overexpression of yes-associated protein contributes to
progression and poor prognosis of non-small-cell lung cancer.
Cancer Sci. 101:1279–1285. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Diep CH, Zucker KM, Hostetter G, Watanabe
A, Hu C, Munoz RM, Von Hoff DD and Han H: Down-regulation of Yes
associated protein 1 expression reduces cell proliferation and
clonogenicity of pancreatic cancer cells. PLoS One. 7:e327832012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gao YB, Chen ZL, Li JG, Hu XD, Shi XJ, Sun
ZM, Zhang F, Zhao ZR, Li ZT, Liu ZY, et al: Genetic landscape of
esophageal squamous cell carcinoma. Nat Genet. 46:1097–1102. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen D, Sun Y, Wei Y, Zhang P, Rezaeian
AH, Teruya-Feldstein J, Gupta S, Liang H, Lin HK, Hung MC and Ma L:
LIFR is a breast cancer metastasis suppressor upstream of the
Hippo-YAP pathway and a prognostic marker. Nat Med. 18:1511–1517.
2012. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu G, Yu FX, Kim YC, Meng Z, Naipauer J,
Looney DJ, Liu X, Gutkind JS, Mesri EA and Guan KL: Kaposi
sarcoma-associated herpesvirus promotes tumorigenesis by modulating
the Hippo pathway. Oncogene. 34:3536–3546. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cao JJ, Zhao XM, Wang DL, Chen KH, Sheng
X, Li WB, Li MC, Liu WJ and He J: YAP is overexpressed in clear
cell renal cell carcinoma and its knockdown reduces cell
proliferation and induces cell cycle arrest and apoptosis. Oncol
Rep. 32:1594–1600. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang W, Nandakumar N, Shi Y, Manzano M,
Smith A, Graham G, Gupta S, Vietsch EE, Laughlin SZ, Wadhwa M, et
al: Downstream of mutant KRAS, the transcription regulator YAP is
essential for neoplastic progression to pancreatic ductal
adenocarcinoma. Sci Signal. 7:ra422014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lau AN, Curtis SJ, Fillmore CM, Rowbotham
SP, Mohseni M, Wagner DE, Beede AM, Montoro DT, Sinkevicius KW,
Walton ZE, et al: Tumor-propagating cells and Yap/Taz activity
contribute to lung tumor progression and metastasis. EMBO J.
33:468–481. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lamar JM, Stern P, Liu H, Schindler JW,
Jiang ZG and Hynes RO: The Hippo pathway target, YAP, promotes
metastasis through its TEAD-interaction domain. Proc Natl Acad Sci
USA. 109:E2441–E2450. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tanaka I, Osada H, Fujii M, Fukatsu A,
Hida T, Horio Y, Kondo Y, Sato A, Hasegawa Y, Tsujimura T and
Sekido Y: LIM-domain protein AJUBA suppresses malignant
mesothelioma cell proliferation via Hippo signaling cascade.
Oncogene. 34:73–83. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Song S, Ajani JA, Honjo S, Maru DM, Chen
Q, Scott AW, Heallen TR, Xiao L, Hofstetter WL, Weston B, et al:
Hippo coactivator YAP1 upregulates SOX9 and endows esophageal
cancer cells with stem-like properties. Cancer Res. 74:4170–4182.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bora-Singhal N, Nguyen J, Schaal C,
Perumal D, Singh S, Coppola D and Chellappan S: YAP1 regulates OCT4
activity and SOX2 expression to facilitate self-renewal and
vascular mimicry of stem-like cells. Stem Cells. 33:1705–1718.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Huo X, Zhang Q, Liu AM, Tang C, Gong Y,
Bian J, Luk JM, Xu Z and Chen J: Overexpression of Yes-associated
protein confers doxorubicin resistance in hepatocellullar
carcinoma. Oncol Rep. 29:840–846. 2013. View Article : Google Scholar : PubMed/NCBI
|