Introduction
Filamin A (FlnA) is an actin cross-linking protein
(1), which serves a role in the
organization of the orthogonal actin network, and is considered an
essential component of the cytoskeleton networks that support
various cellular functions (2). In
addition to filamentous actin, FlnA interacts with >60
functionally diverse cellular proteins, including transmembrane
receptors, signaling molecules and DNA damage repair proteins
(1,3–5). These
diverse interactions suggest that FlnA is essential for versatile
cell signaling. Previous research has investigated the involvement
of FlnA in different types of cancer (6,7). FlnA was
originally identified as able to promote cancer progression.
However, a recent study demonstrated that the effect of FlnA on
cancer cells depended on its subcellular localization and
corresponding binding partners (8).
Breast cancer type 1 susceptibility gene (BRCA1) is
a DNA repair gene commonly associated with familial breast and
ovarian cancers (9,10). Studies in BRCA1-knockout models have
indicated that mice lacking BRCA1 have poor genomic integrity and
cannot survive (11). It has been
reported that FlnA may act as a binding partner of BRCA1, and that
it was required for efficient regulation of early stages of DNA
repair in a yeast two-hybrid screening model (12). However, to the best of our knowledge,
the association of these genes in cancer patients and cell lines
remains to be investigated.
In the present study, the expression pattern of FlnA
and BRCA1 proteins were investigated in breast cancer tissues. The
clinicopathological and prognostic value of the subcellular
localizations of these proteins, and the association between
FlnA/BRCA1 expression and clinicopathological characteristics of
breast cancer patients were also investigated. Furthermore, the
association between FlnA and BRCA1 genes was investigated through
RNA interference in the breast cancer cell line, MCF-7. The results
of the present study provide a basis for researching FlnA and BRCA1
function in tumor progression, as well as insight into the
potential prognostic and therapeutic value of FlnA/BRCA1
expression.
Materials and methods
Patients
Tumor tissue specimens were collected from 424 women
(mean age, 56.49 years; range, 26–94 years), who had been diagnosed
with breast cancer. Patients who had received any preoperative
chemotherapy or radiotherapy prior to surgery were excluded. A
total of 88 tissues were collected at the Department of General
Surgery of the Suzhou Municipal Hospital (Jiangsu, China) between
January 2009 and December 2010. The remaining 336 tissues were
collected from the Department of General Surgery of Shanghai Renji
Hospital (Shanghai, China). All 424 cases were pathologically
confirmed as breast cancer by hematoxylin and eosin (H&E)
staining at the time of surgical resection. The clinical parameters
of the patients are presented in Table
I. Survival data were collected through patient follow-up each
year for seven years. The present study was approved by the Ethics
Review Board of Suzhou Vocational Health Technology College
(Jiangsu, China), and written informed consent was obtained from
each participant.
 | Table I.Correlations between FlnA/BRCA1
expression and clinicopathological factors in breast cancer. |
Table I.
Correlations between FlnA/BRCA1
expression and clinicopathological factors in breast cancer.
|
|
| FlnA expression |
| BRCA1 expression |
|---|
|
|
|
|
|
|
|---|
| Factor | Number | Low (%) | High (%) | P-value | Number | Low (%) | High (%) | P-value |
|---|
| Age (years) |
|
|
| 0.719 |
|
|
| 0.307 |
|
<60 | 259 | 177 (40.78) | 82 (18.8) |
| 259 | 145 (33.41) | 114 (26.27) |
|
| ≥60 | 175 | 120 (27.65) | 55 (12.67) |
| 175 | 89 (20.51) | 86 (19.82) |
|
| Tumor size |
|
|
| 0.027 |
|
|
| 0.064 |
| <2.5
cm | 218 | 137 (32.5) | 81 (19.2) |
| 172 | 127 (30.2) | 91 (21.6) |
|
| ≥2.5
cm | 203 | 148 (35.2) | 55 (13.1) |
| 163 | 100 (23.8) | 103 (24.5) |
|
| Lymph node
metastasis |
|
|
| 0.869 |
|
|
| 0.535 |
|
Negative | 72 | 47 (36.4) | 25 (19.4) |
| 72 | 24 (18.6) | 48 (37.2) |
|
|
Positive | 57 | 38 (29.5) | 19 (14.7) |
| 57 | 22 (17.1) | 35 (27.1) |
|
| Differentiation |
|
|
| 0.392 |
|
|
| 0.099 |
| I | 3 | 3 (1.2) | 0 (0) |
| 3 | 3 (1.2) | 0 (0) |
|
| II | 171 | 118 (45.7) | 53 (20.5) |
| 171 | 96 (37.2.8) | 75 (29.1) |
|
|
III | 84 | 62 (24) | 22 (8.5) |
| 84 | 56 (21.7) | 28 (10.9) |
|
| ER |
|
|
| 0.694 |
|
|
| 0.003 |
| ER
(−~+) | 186 | 162 (38.3) | 74 (17.5) |
| 187 | 86 (20.3) | 101 (23.9) |
|
| ER
(++~+++) | 236 | 125 (29.6) | 62 (14.7) |
| 236 | 143 (33.8) | 93 (22) |
|
| PR |
|
|
| 0.021 |
|
|
| 0.023 |
| PR
(−~+) | 280 | 201 (47.5) | 79 (18.7) |
| 280 | 163 (38.5) | 117 (27.7) |
|
| PR
(++~+++) | 143 | 86 (20.3) | 57 (13.5) |
| 143 | 66 (15.6) | 77 (18.2) |
|
| HER2 |
|
|
| 0.861 |
|
|
| 0.023 |
| HER2
(−~+) | 153 | 103 (24.3) | 50 (11.8) |
| 153 | 94 (22.2) | 59 (13.9) |
|
| HRE2
(++~+++) | 270 | 184 (43.5) | 86 (20.3) |
| 270 | 135 (31.9) | 135 (31.9) |
|
| Subtype |
|
|
| 0.521 |
|
|
| <0.001 |
| Lumin
A | 83 | 57 (13.5) | 26 (6.2) |
| 83 | 49 (11.6) | 34 (8.1) |
|
| Lumin
B | 116 | 76 (18.1) | 40 (9.5) |
| 116 | 44 (10.5) | 72 (17.1) |
|
| HER2
(+) | 147 | 103 (24.5) | 44 (10.5) |
| 147 | 85 (20.2) | 62 (14.7) |
|
| Basal
like | 51 | 37 (8.8) | 14 (3.3) |
| 51 | 31 (7.4) | 20 (4.8) |
|
| Normal
like | 24 | 13 (3.1) | 11 (2.6) |
| 24 | 19 (4.5) | 5 (1.2) |
|
Construction of tissue microarray
(TMA)
Two specialized histopathologists (authors of this
paper)isolated 2 1.6-mm sections from the centre and periphery of
the tumors, and placed them into a paraffin block using a tissue
puncher/arrayer (patent no. 200920350099.2). Sections of 4-µm were
sliced from the paraffin block, deparaffinized in xylene for 10 min
and conducted three times at room temperature and rehydrated using
a descending ethanol series (100, 95, 80 and 70%) for
immunohistochemical staining. The deparaffinized reagent was xylene
and rehydrated reagent was ethanol (100, 95, 80, 70 and 0%).
Immunohistochemistry
A rabbit anti-human Filamin A antibody (dilution,
1:100; clone, PM61317; cat. no. MAB1678), and a mouse anti-human
BRCA1 antibody (dilution, 1:150; clone, MS110; cat. no. OP92) were
purchased from Abcam (Cambridge, UK). Other antibodies [rabbit
anti-human ER antibody (dilution, 1:100; clone, SP1; cat. no.
GT205611) rabbit anti-human PR antibody (dilution, 1:100; clone,
SP2; cat. no. GT205711) rabbit anti-human HER2 antibody (dilution,
1:100; clone, SP3; cat. no. GT210011)] and the IHC kit (cat. no.
KIT-5010) were purchased from Fuzhou Maixin Biotech, Co., Ltd.
(Fuzhou, China). Immunohistochemical staining of FlnA and BRCA1 was
performed according to the manufacturer's instructions.
Immunohistochemical staining of FlnA and BRCA-1 was
examined by 2 independent pathologists (Suzhou Municipal Hospital,
Jiangsu, China) who were blinded to the clinicopathological
parameters of the breast cancer patients. A semi-quantitative
scoring system was used, based on the distribution of tumor cells
stained positively for FlnA and BRCA-1: 0, 0–10%; 1, 11–30%; 2,
31–60%, and 3, 61–100%. The distribution score was multiplied by
the intensity factor: 1, staining intensity just exceeding
background; 2, weak positive; 3, middle positive; 4, dark brown
staining evident upon macroscopic inspection of the slide. The
resulting score was divided by 4. For FlnA, final scores of 0–1
were considered to indicate low expression, and scores of 2–3 were
considered to indicate high expression. For BRCA1, final scores of
0–2 were considered to indicate low expression, and a score of 3
was considered to indicate high expression.
Cell culture and transfection
The human breast cancer cell line, MCF-7, was
obtained from the American Type Culture Collection (Manassas, VA,
USA) and cultured in RPMI-1640 medium (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal
bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.). To
acquire a better interference efficiency a total of 3 stealth RNA
interference (RNAi) small interfering RNAs (siRNAs) target FlnA and
2 control RNAi were designed by Invitrogen (Thermo Fisher
Scientific, Inc.) based on the FlnA GenBank accession no.
NM_001456.3 (13). According to the
FlnA sequence, the siRNAs sequences designed were:
5′-UGCAUUUGGCGGAAAGUGGGC-3′; 5′-UUUCUUCGGGUUCAGUUUGGG-3′; and
5′-UAUACUUUGACCUUGUUGGGG-3′. The control RNAi sequences were
5′-UUCUCCGAACGUGUCACGUTT-3′; and 5′-ACGUGACACGUUCGGAGAATT-3′. The
RNAi with the greatest FlnA knockdown effect was also selected.
MCF-7 cells were seeded at 1×105 in 24-well plates, and
incubated for 24 h prior to transfection. The individual RNAi were
transfected into MCF-7 cells at 10 µM using
Lipofectamine® 2000 (Thermo Fisher Scientific, Inc.).
The cells were and cultured for 24 h prior to subsequent
experimentation.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted from MCF-7 cells using an
RNeasy Mini kit (Qiagen GmbH, Hilden, Germany) and cDNA was
synthesized using a Primescript First cDNA synthesis kit (Takara
Bio, Inc., Otsu, Japan), according to the manufacturers' protocols.
RT-qPCR was performed in triplicate using a Bio-Rad iCycler CFX96
(Bio-Rad Laboratories, Inc., Hercules, CA, USA) and a SYBR Green
PCR Master mix (Takara Bio, Inc.), according to the manufacturer's
protocol. The following primers were used: FlnA, forward,
5′-AGCCTCCACGAGACATCATC-3′ and reverse, 5′-CCAGTGTGTACTCCCCCTTG-3′;
BRCA-1, forward, 5′-GGCTATCCTCTCAGAGTGACA-3′ and reverse,
5′-CTGATGTGCTTTGTTCTGGA-3′; GAPDH forward,
5′-GCACCGTCAAGGCTGAGAAC-3′ and reverse, 5′-TGGTGAAGACGCCAGTGGA-3′.
The levels of gene expression were normalized to the expression of
GAPDH. The thermocycling conditions were as follows: 95°C for 5
min, followed by 40 cycles of 95°C for 30 sec, 60°C for 30 sec and
72°C for 30 sec, with a final extension of 72°C for 3 min. The
2−∆∆Cq method was used for the quantification of FlnA
and BRCA-1 gene expression (14).
Statistical analysis
All data are presented as the mean ± standard
deviation. The χ2 test was used to analyze the
association between FlnA/BRCA1 expression and clinicopathological
features. Unpaired Student's t-test was used for all other
comparisons. All tests were performed using software SPSS 17.0
(SPSS Inc., Chicago, IL, USA). P<0.05 was considered to indicate
a statistically significant difference. Kaplan-Meier survival plots
were generated, and comparisons between the survival curves were
made with the log-rank test.
Results
FlnA protein expression levels in
breast cancer tissue
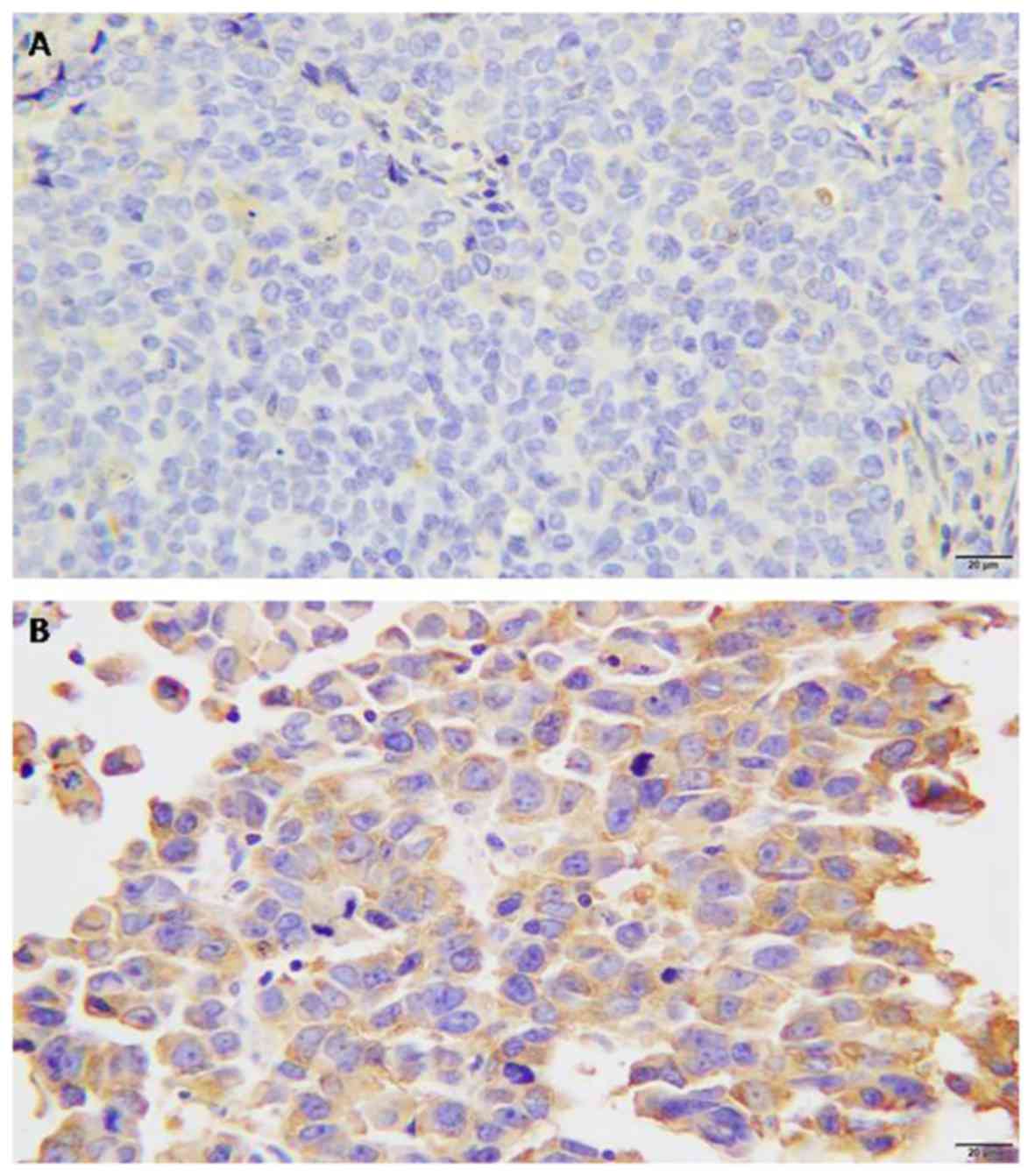
FlnA protein expression was assessed in breast
cancer and normal breast tissue specimens (Fig. 1A and B). FlnA protein was mainly
detected in the myoepithelial and basal cell cytoplasm or
intercellularly in breast cancer tissue. FlnA protein expression
was detected in 52.6% breast cancer tissues, whereas FlnA was
rarely detectable in normal breast tissues or benign tumor ductal
epithelium sections. The rate of positive expression in normal
breast epithelium was 5.9%.
Association of FlnA protein expression
with clinicopathological and patient prognosis features
The associations between FlnA protein expression
level and various clinicopathological features of the patients with
breast cancer were investigated. The protein expression level of
FlnA was negatively associated with tumor size (P<0.05; Table I). The rate of positive FlnA staining
increased with progesterone receptor expression (P<0.05). No
significant association was identified between FlnA protein
expression level and age, lymph node metastasis, differentiation,
estrogen receptor status or Her2/neu protein expression (P>0.05;
Table I).
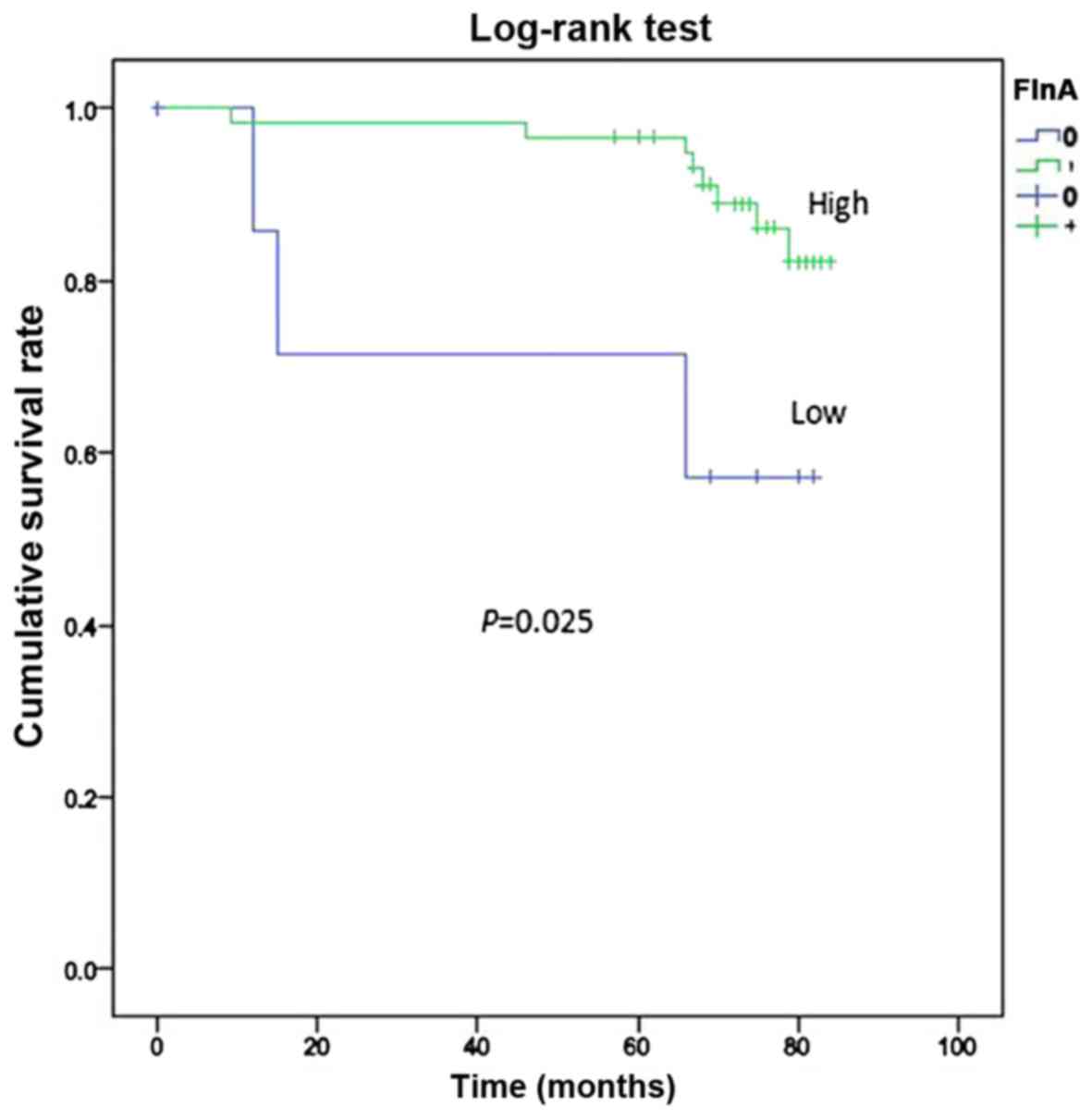
Only 84 cases in Suzhou Municipal Hospital were
investigated for survival. The mean duration of follow-up was 84
months after surgery and 75th percentile of duration was 74 months.
During the follow-up period, 14 mortalities occurred (16.67%). The
overall survival rate of patients exhibiting FlnA overexpression
was determined by the by the Kaplan-Meier survival analysis and
log-rank test, and was significantly increased compared with those
who did not exhibit FlnA overexpression (P=0.025; Fig. 2).
Nucleic BRCA1 expression in breast
cancer tissue and its association with clinicopathological
parameters
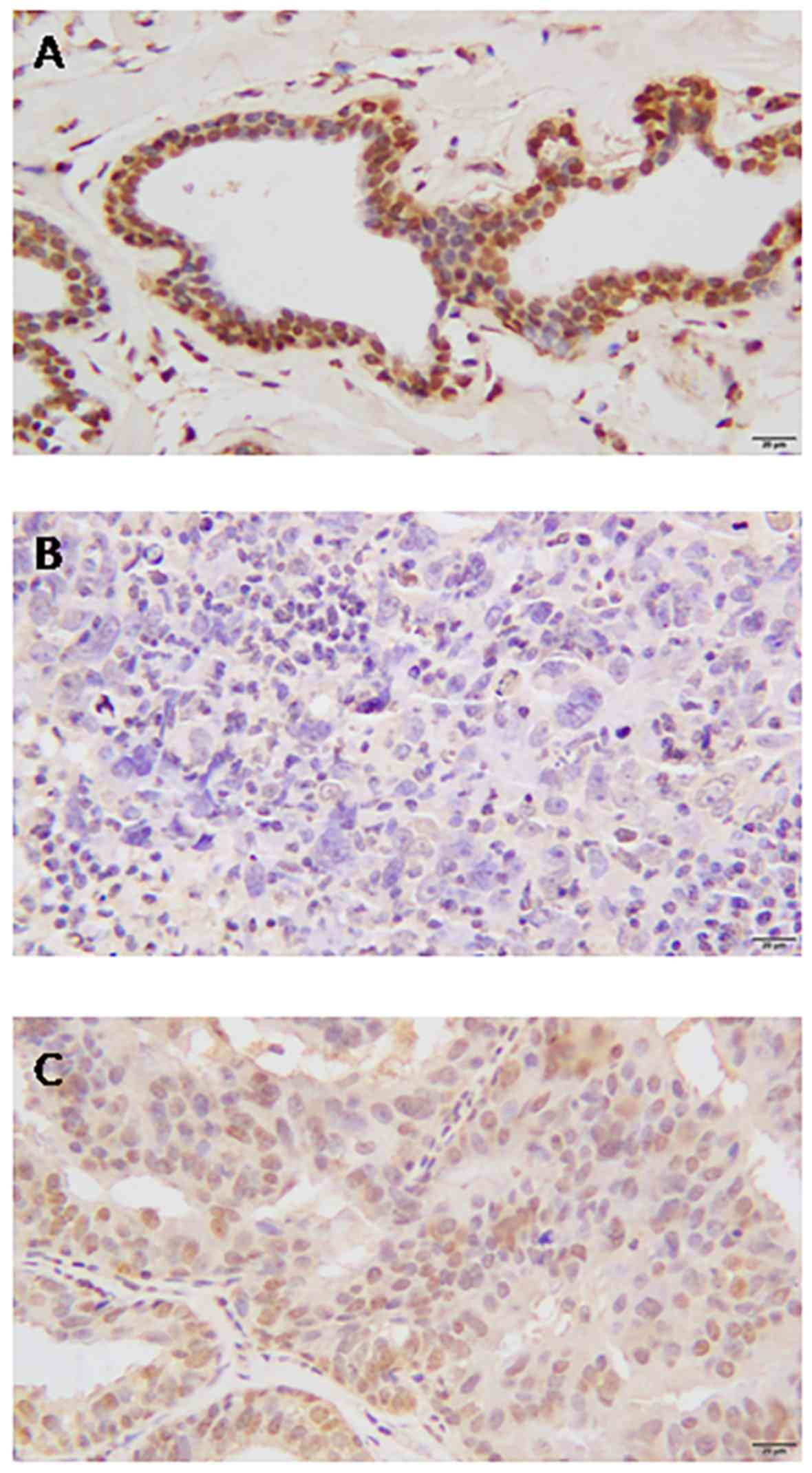
Adjacent normal breast tissue stained strongly
positive for BRCA1 in the cell nuclei, and no cytoplasmic or
membranous staining was observed (Fig.
3A). In contrast, the protein expression level of BRCA1 in
breast cancer tissue was demonstrated to be either nucleic or
cytoplasmic (Fig. 3B). Of the 424
breast cancer tissues stained for BRCA1, complete loss of nucleic
expression was detected in 207 (48.82%) cases. Absent nucleic
staining was considered to indicate low expression.
High nucleic BRCA1 protein expression level was
associated with clinicopathological features, including breast
cancer molecular subtype, phenotype, and estrogen and progesterone
receptor expression (P<0.05; Table
I). However, no association was detected between nucleic BRCA1
protein expression and lymph node metastasis, age, tumor size,
differentiation or Her2/neu expression (P>0.05; Table I).
Correlation of FlnA and BRCA1
expression
A positive correlation between FlnA expression and
BRCA1 nucleic expression was identified, indicating that high
expression of BRCA1 was associated with high expression of FlnA
(P<0.001; Table II).
| Table II.Correlation between FlnA and BRCA1
protein expression in patients with breast cancer
(χ2=0.203; P<0.001). |
Table II.
Correlation between FlnA and BRCA1
protein expression in patients with breast cancer
(χ2=0.203; P<0.001).
|
| FlnA
expression |
|---|
|
|
|
|---|
| BRCA1
expression | Negative | Positive | Total |
|---|
| Negative | 176 | 54 | 230 |
| Positive | 111 | 83 | 194 |
| Total | 287 | 137 | 424 |
Silencing FlnA expression
downregulates BRCA1 expression in MCF-7 cells
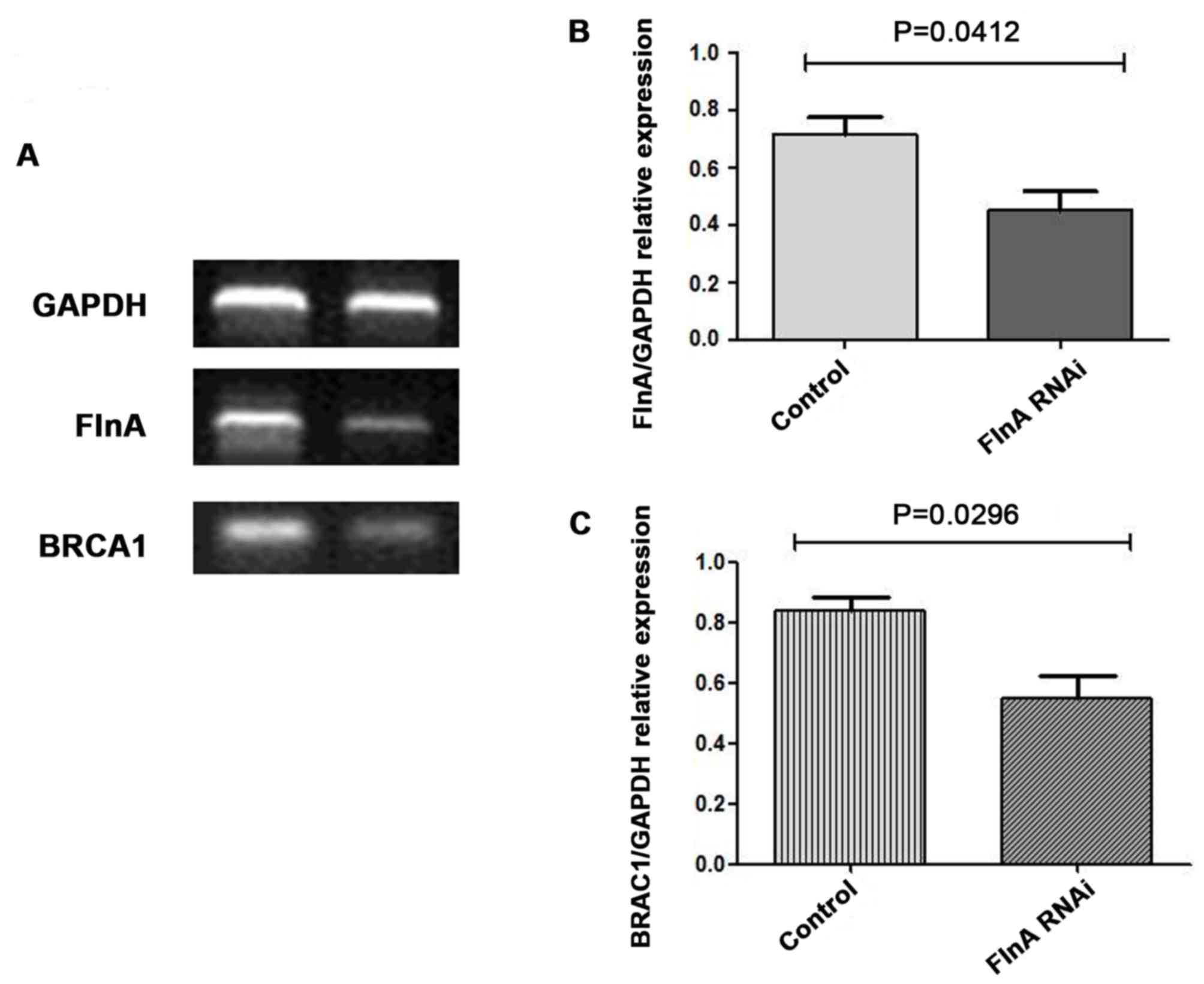
To assess whether FlnA regulates BRCA1 expression,
RNAi technology was used to knock down FlnA expression in MCF-7
cells. RT-qPCR analysis revealed effective and specific knockdown
of FlnA after 48 h of transfection (Fig.
4A and B). FlnA was decreased by ~50% following transfection of
MCF-7 cells by FlnA-specific stealth RNAi siRNAs, and BRCA-1
expression was also decreased (Fig. 4A
and C).
Discussion
FlnA was first identified as a non-muscle actin
filament cross-linking protein, or gelation factor, in 1975
(15). FlnA serves as a scaffold
protein for >90 binding partners, including channels, receptors,
intracellular signaling molecules, transcription factors,
transmembrane receptors and DNA damage repair proteins (16). FlnA has been associated with cancer
metastasis and progression. Originally it was suggested to be a
cancer-promoting protein (17),
however, it has been more recently established that FlnA serves a
dual role in cancers, depending on its subcellular localization and
the corresponding binding partners (8).
A number of studies have reported abnormal
expression of FlnA in various types of cancer. Overexpression of
FlnA has been observed in multiple malignancies, including
hepatocellular carcinoma, breast cancer, colon cancer, melanoma and
prostate cancer (17–19). In the present study, it was
demonstrated that FlnA was overexpressed in breast cancer tissue
compared with distant normal breast tissue, which is consistent
with previous reports (20–22). Furthermore, it was demonstrated that
FlnA was highly expressed in myoepithelial cells of normal breast
tissues.
BRCA1 is a tumor suppressor gene, which is expressed
in all cells. BRCA1 mutations lead to an increased predisposition
to breast and ovarian cancer (23).
BRCA1 has been implicated in several aspects of the DNA damage
response (DDR) and FlnA has been reported to function in DNA repair
(12,24). A previous study identified FlnA as a
partner of BRCA1, and demonstrated that FlnA is required for the
efficient regulation of the early stages of DNA repair (12). However, to the best of our knowledge,
no further research regarding the association between FlnA and
BRCA1 has been performed.
In the present study, the expression of BRCA1 was
analyzed in 424 breast cancer tissue samples using immunochemistry,
and a positive association between FlnA and BRCA1 was identified
(P<0.001). Knockdown of FlnA expression using stealth RNAi siRNA
in MCF-7 cells resulted in downregulation of BRCA1 expression. It
is speculated that FlnA regulates BRCA1 expression in breast
cancer, which was verified in another cell line (data not shown).
However, further research is required to determine the mechanism by
which FlnA regulates BRCA1 expression.
Overall, the present study indicates that increased
FlnA expression is exhibited in breast cancer originating from
epithelial cells, and that FlnA expression is associated with tumor
progression. A positive association between FlnA and BRCA1
expression was also demonstrated, and it is hypothesized that FlnA
regulates the expression of BRCA1. The precise mechanism, and
whether FlnA may be used as a prognostic indictor of breast cancer,
remains to be determined.
Acknowledgements
The authors would like to thank Dr Yumin Hu (Soochow
University) for providing the breast cancer cell line. The authors
would like to also thank the staff in the Pathology Department of
Suzhou Municipal Hospital for collating the specimens of breast
cancer.
Funding
The present study was supported by grants from
Jiangsu Province University Outstanding Science and Technology
Innovation Team (grant no. 2015023) and Jiangsu Provincial Medical
Youth Talent.
Availability of data and materials
The data used to support the findings of this study
are available from the corresponding author upon request.
Authors' contributions
Research was designed by JS. Patient samples and
data were provided by ML and GB. IHC was performed by XL, LW and
YG. Cell experiments in vitro were performed by ZS and JY.
Statistical analysis of data was performed by YG. The manuscript
was critically reviewed by all authors. The study was supervised by
JS.
Ethics approval and consent to
participate
Prior to commencing this study the written informed
consent was obtained from each patient accepting the surgical
treatment. The present study was approved by the Ethics Review
Board of Suzhou Vocational Health Technology College.
Patient consent for publication
Prior to commencing this study, the approval from
the Ethics Review Board of Suzhou Vocational Health College was
granted.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Feng Y and Walsh CA: The many faces of
filamin: A versatile molecular scaffold for cell motility and
signalling. Nat Cell Biol. 6:1034–1038. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Popowicz GM, Schleicher M, Noegel AA and
Holak TA: Filamins: Promiscuous organizers of the cytoskeleton.
Trends Biochem Sci. 31:411–419. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Meng X, Yuan Y, Maestas A and Shen Z:
Recovery from DNA damage-induced G2 arrest requires actin-binding
protein filamin-A/actin-binding protein 280. J Biol Chem.
279:6098–6105. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Duval D, Lardeux A, Le Tourneau T, Norris
RA, Markwald RR, Sauzeau V, Probst V, Le Marec H, Levine R, Schott
JJ and Merot J: Valvular dystrophy associated filamin A mutations
reveal a new role of its first repeats in small-GTPase regulation.
Biochim Biophys Acta. 1843:234–244. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nakamura F, Pudas R, Heikkinen O, Permi P,
Kilpeläinen I, Munday AD, Hartwig JH, Stossel TP and Ylänne J: The
structure of the GPIb-filamin A complex. Blood. 107:1925–3192.
2016. View Article : Google Scholar
|
|
6
|
Shao QQ, Zhang TP, Zhao WJ, Liu ZW, You L,
Zhou L, Guo JC and Zhao YP: Filamin A: Insights into its exact role
in cancers. Pathol Oncol Res. 22:245–252. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yue J, Lu H, Liu J, Berwick M and Shen Z:
Filamin-A as a marker and target for DNA damage based cancer
therapy. DNA Repair (Amst). 11:192–200. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Savoy RM and Ghosh PM: The dual role of
filamin A in cancer: Can't live with (too much of) it, can't live
without it. Endocr Relat Cancer. 20:R341–R356. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ford D, Easton DF, Bishop DT, Narod SA and
Goldgar DE: Risks of cancer in BRCA1-mutation carriers. Breast
cancer linkage consortium. Lancet. 343:692–695. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Easton DF, Bishop DT, Ford D and Crockford
GP: Genetic linkage analysis in familial breast and ovarian cancer:
Results from 214 families. The breast cancer linkage consortium. Am
J Hum Genet. 52:678–701. 1993.PubMed/NCBI
|
|
11
|
Gowen LC, Johnson BL, Latour AM, Sulik KK
and Koller BH: Brca1 deficiency results in early embryonic
lethality characterized by neuroepithelial abnormalities. Nat
Genet. 12:191–194. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Velkova A, Carvalho MA, Johnson JO,
Tavtigian SV and Monteiro AN: Identification of Filamin A as a
BRCA1-interacting protein required for efficient DNA repair. Cell
Cycle. 9:1421–1433. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Benson DA, Karsch-Mizrachi I, Lipman DJ,
Ostell J and Wheeler DL: GenBank. Nucleic Acids Res. 33:D34–D38.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hartwig JH and Stossel TP: Isolation and
properties of actin, myosin, and a new actinbinding protein in
rabbit alveolar macrophages. J Biol Chem. 250:5696–5705.
1975.PubMed/NCBI
|
|
16
|
Nakamura F, Stossel TP and Hartwig JH: The
filamins: Organizers of cell structure and function. Cell Adh Migr.
5:160–169. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Flanagan LA, Chou J, Falet H, Neujahr R,
Hartwig JH and Stossel TP: Filamin A the Arp2/3 complex, and the
morphology and function of cortical actin filaments in human
melanoma cells. J Cell Biol. 155:511–517. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ai J, Huang H, Lv X, Tang Z, Chen M, Chen
T, Duan W, Sun H, Li Q, Tan R, et al: FLNA and PGK1 are two
potential markers for progression in hepatocellular carcinoma. Cell
Physiol Biochem. 27:207–216. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lin JF, Xu J, Tian HY, Gao X, Chen QX, Gu
Q, Xu GJ, Song JD and Zhao FK: Identification of candidate prostate
cancer biomarkers in prostate needle biopsy specimens using
proteomic analysis. Int J Cancer. 121:2596–2605. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jiang X, Yue J, Lu H, Campbell N, Yang Q,
Lan S, Haffty BG, Yuan C and Shen Z: Inhibition of filamin-A
reduces cancer metastatic potential. Int J Biol Sci. 9:67–77. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Alper O, Stetler-Stevenson WG, Harris LN,
Leitner WW, Ozdemirli M, Hartmann D, Raffeld M, Abu-Asab M, Byers
S, Zhuang Z, et al: Novel anti-filamin-A antibody detects a
secreted variant of filamin-A in plasma from patients with breast
carcinoma and high-grade astrocytoma. Cancer Sci. 100:1748–1756.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tian HM, Liu XH, Han W, Zhao LL, Yuan B
and Yuan CJ: Differential expression of filamin A and its clinical
significance in breast cancer. Oncol Lett. 6:681–686. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Miki Y, Swensen J, Shattuck-Eidens D,
Futreal PA, Harshman K, Tavtigian S, Liu Q, Cochran C, Bennett LM
and Ding W: A strong candidate for the breast and ovarian cancer
susceptibility gene BRCA1. Science. 266:66–71. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kim KM, Moon YJ, Park SH, Park HJ, Wang
SI, Park HS, Lee H, Kwon KS, Moon WS, Lee DG, et al: Individual and
combined expression of DNA damage response molecules PARP1,
gammaH2AX, BRCA1, and BRCA2 predict shorter survival of soft tissue
sarcoma patients. PLoS One. 11:e01631932016. View Article : Google Scholar : PubMed/NCBI
|