Introduction
Gastric cancer is one of the malignant tumors with
highest mortality all over the world, especially in East Asia
(1). Many patients have missed the
best time for diagnosis and treatment when they are certainly
diagnosed, resulting in the metastasis of tumor cells and
developing into the terminal stage of cancer. Nowadays, lack of
efficient biomarkers for early diagnosis, comprehensive treatment
and cancer monitoring has been considered as one of the main
obstacles for better prognosis of gastric cancer (2). As a result, it is of great importance to
further explore the molecular mechanism during the occurrence and
development of gastric cancer, hoping to provide new strategy for
diagnosis, prognosis and treatment (3).
During the recent years, non-coding RNAs have been
generally concerned because of their diverse roles in the
post-transcriptional regulation and they are considered to have
great influence on human diseases (4). Long non-coding RNAs (lncRNA) are one of
the functional non-coding RNAs, which have more than 200
nucleotides, and they are usually lack of open reading frames and
the ability of coding proteins (5).
LncRNAs are located in both cell nucleus and cytoplasm, and
involved in the regulation of several cellular events, such as cell
development, proliferation, apoptosis and so on (6).
More and more lncRNAs have been reported to be
tightly connected with the occurrence and development of malignant
tumors (7). These lncRNAs are proved
to be engaged in the imbalanced gene regulation and aberrant
biological processes (BPs) that contribute to malignant
transformation. Especially, the functions and therapeutic potential
of cancer-related lncRNAs have been greatly focused in the past few
years. To this end, we consider that the overall knowledge about
lncRNAs in gastric cancer needs to be fully elucidated.
In our study, we analyzed the qualified public data
from He et al study (8) and
identified 50 differentially expressed lncRNAs in gastric cancer.
We adopted the systems biology-based approach of weighted gene
co-expression network analysis (WGCNA) to construct a co-expression
network about these lncRNAs and mRNAs, and identified 6 significant
co-expression modules. Next, we carried out Gene Ontology (GO)
enrichment and Kyoto Encyclopedia of Genes and Genomes (KEGG)
pathway analysis to predict the possible functions of the lncRNAs
in modules. Besides, to further confirm the roles of these lncRNAs
in gastric cancer, we performed survival analysis and found two of
the lncRNAs could be considered as prognostic factors, one of which
was novel. In all, our study identified several gastric
cancer-related lncRNAs, confirmed their biological functions, and
most importantly, we found LINC00982 and LL22NC03-N14H11.1 were
associated with patients' survival time and could be considered as
potential prognosis for gastric cancer. Besides, we performed cis,
trans or ceRNA regulation analysis and found the potential target
genes of these two lncRNAs were mainly involved in the
proliferation of gastric cancer cells. For further validation, we
overexpressed or downregulated the lncRNAs in gastric cancer cell
lines and found that LINC00982 inhibited while LL22NC03-N14H11.1
promoted the proliferation of gastric cancer cells. As a result
they can be considered as hopeful prognostic factors in gastric
cancer.
Materials and methods
Collection of gene expression
datasets
Data preprocessing and microarray data were
downloaded from Gene Expression Omnibus (GEO) database (www.ncbi.nlm.nih.gov/geo/) under the accession
number GSE79973 (8). This dataset was
acquired from the study by He et al (8). Totally, 20 samples were included in this
dataset, which consisted of 10 gastric cancer samples and 10 normal
samples. We used array Quality package to quality control and limma
package to apply raw data in R software. The normalization criteria
were quantile normalization. Genes having fold-changes ≥2 and FDR
<0.05 were selected as of significantly differential expression
(9).
Enrichment analysis
To conduct enrichment analyses, the package
clusterProfiler (version 3.5.5) of R (version 3.4.0) was used for
analyzing the KEGG pathways and GO processes, and DOSE (version
3.3.2) of R (version 3.4.0) as was used for Gene Set Enrichment
Analysis (GSEA) (10,11).
WGCNA
WGCNA is a typical systemic biological method for
describing the correlation patterns among genes and identifying
modules of highly correlated genes by using average linkage
hierarchical clustering coupled with the topological overlap
dissimilarity measure based on high-throughput chip data or RNA-Seq
data (12). In current study, WGCNA
package (version 1.60) in R was used to construct lncRNA-mRNA
co-expression network and identify modules based on the expression
levels of the differentially expressed mRNAs and lncRNAs. The gene
modules were signified by different colors and the grey module
showed the genes that cannot be merged (13).
LncRNAs function prediction
Cis-regulation target genes were predicted based on
the nearby genes of lncRNAs and we chose the promoters that located
in the 1M bp regions around the lncRNAs as potential target genes
by Bedtools and hg19 as reference genome (14). Trans-regulation target genes were
predicted by the sequence of lncRNAs based on RBPDP, and relative
score >80% was used to select potential target genes (15). Co-expression target genes were
predicted by the psych package in R (version 3.4.0) to calculate
Pearson correlation coefficient between genes and two lncRNAs. The
absolute value of co-expression coefficient >0.7 and P-value
<0.05 were used to select co-expression genes. The construction
of ceRNA network was as followings: i) We predicted the potential
target microRNAs of the lncRNAs through RegRNA 2.0 (16); ii) Then, we analyzed the differential
expression of the microRNAs that got from 1 in gastric cancers
through The Cancer Genome Atlas (TCGA); iii) Next, we found the
potential target genes of the microRNAs based on miRTarBase
(17); and iv) Finally, the target
genes and lncRNAs were used to construct ceRNA network when the
co-expression coefficient was >0.7 (9).
Cell culture
The SGC-7901 human gastric carcinoma cell line were
cultured in RPMI-1640 medium (Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA), supplemented with 10% fetal bovine serum
(Gibco; Thermo Fisher Scientific, Inc.) and incubated in an
atmosphere of 5% CO2 at 37°C. SGC-7901 cell line was
obtained from the cell bank of the Committee on Type Culture
Collection of the Chinese Academy of Sciences.
Cell transfection
For overexpression of LINC00982, the coding region
was PCR-amplified from cDNA generated from the SGC-7901 cell line
and was subcloned into a PINCO retroviral vector. For knocking down
LL22NC03-N14H11.1, we firstly designed the lncRNA specific small
interfering (si)RNAs (the sequence is 5′-GCACUCACCUACACGUUUAGG-3′),
and cloned it into plko.1 vector. SGC-7901 cells (1×105
per well) were inoculated in six-well plates and cultured for 24 h.
The cells were transfected with LINC00982-overexpression and the
corresponding negative control, or the siRNAs targeting
LL22NC03-N14H11.1 and the corresponding negative control retroviral
vectors using Lipofectamine 3000 transfection reagent (Invitrogen;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) following the
manufacturer's instructions.
Cell proliferation assay
An MTT assay was used to determine the cell
proliferative capacity after overexpression of LINC00982 or
down-regulation of LL22NC03-N14H11.1. In brief, transfected cells
(2×104 cells/well) were seeded into 96-well culture
plates and cultured in RPMI-1640 medium containing 10% FBS. After
culturing the seeding cells for 12, 24, 48 or 72 h, MTT reagent
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was added to each
well, followed by incubation at 37°C for an additional 4 h.
Subsequently, 150 µl dimethyl sulfoxide (Sigma-Aldrich; Merck KGaA)
was added to dissolve the crystals for 10 min at 37°C. The
spectrometric absorbance at 490 nm was measured by an EnSpire
Multimode Plate Reader (PerkinElmer, Inc., Waltham, MA, USA).
Flow cytometry
Anti-mouse-ki-67-PE flow cytometric antibody
(eBioscience; Thermo Fisher Scientific, Inc.) was purchased from
eBioscience. And the intracellular staining of ki-67 was performed
by the eBioscience™ Foxp3/Transcription Factor Staining Buffer Set
(eBioscience; Thermo Fisher Scientific, Inc.), following the
manufacturer's instructions. Briefly, cells were incubated with the
mixture of fixation/permeabilization concentrate and diluent (at
the ratio of 1:3) at 4°C for 2 h. Then after washing with
fixation/permeabilization buffer, cells were incubated with
anti-mouse-ki-67-PE flow cytometric antibody for 0.5 h in 4°C.
Finally, the cells were washed and analyzed by BD
FACSCanto™.
Reverse transcription-quantified
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted from cells with
GenElute™ Total RNA Purification kit (Sigma-Aldrich;
Merck KGaA) according to the manufacturer's instructions. Then the
RNA was reversely transcribed into cDNA with
PrimeScript™ RT reagent kit (Takara Bio, Inc., Otsu,
Japan). RT-PCR reactions were carried out with SYBR®
Premix Ex Taq™ II (Takara Bio, Inc.) using an ABI Prism
7700 Sequence Detector. Relative mRNA expression levels were
calculated by normalizing the relative cycle threshold value to the
control group after normalization to the internal control, β-actin
and then the results of the semi-quantitative RT-PCR were
quantified (18). The primer pairs
used are as follows: Human-LL22NC03-N14H11.1-Foward:
5′-GAGTCTGGGGATCAGCATCG-3′, Human-LL22NC03-N14H11.1-Reverse:
5′-TCCAGGGGGCTGGATAATGA-3′; Human-LINC00982-Forward:
5′-AAGTCGTGCTGAGTGTCTGG-3′, Human-LINC00982-Reverse:
5′-CACAACGTGCCACGAACAAT-3′; Human-β-actin-Forward:
5′-CAGGGCGTGATGGTGGGCA-3′, Human-β-actin- Reverse:
5′-CAAACATCATCTGGGTCATCTTCTC-3′.
Statistical analysis
Statistical analyses were performed using R (version
3.4.0; http://www.r-project.org/). The GSE15459
dataset, which includes 200 gastric cancer samples, together with
lncRNA and mRNA expression and clinical information, were used to
analyze the associations between lncRNA expression signatures and
the corresponding overall survival in patients with gastric cancer
(19). Survival curves were
constructed with the Kaplan-Meier method and the log-rank test was
used to determine survival differences between groups. The survival
data were evaluated by univariate and multivariate Cox regression
analyses to search for independent prognostic factors. P<0.05
was considered to indicate a statistically significant difference
(20). When two independent groups
were compared, if the data was normally distributed, an unpaired
Student's t-test was used.
Results
Differentially expressed mRNAs and
lncRNAs in gastric cancer
Based on the qualified data from He et al
study (8), we identified the lncRNAs
related to gastric cancer through GenCode V24 (21). By the criteria of FDR <0.05 and
fold-change >2, we identified 953 mRNAs and 50 lncRNAs
differentially expressed in gastric cancer compared with
para-carcinoma tissues. As a result, 499 mRNAs and 14 lncRNAs were
upregulated, and 454 mRNAs and 36 lncRNAs were downregulated
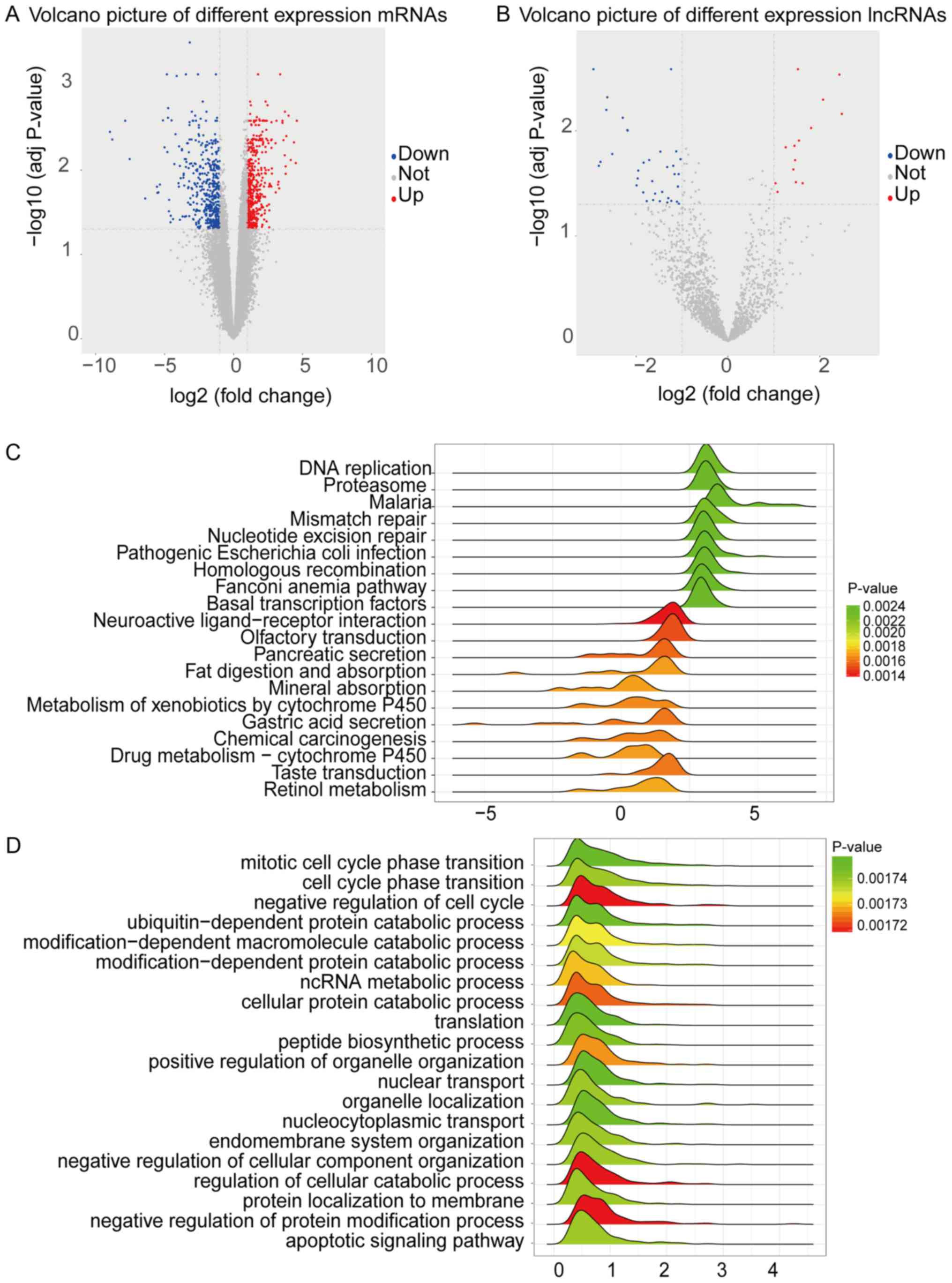
(Fig. 1A and B).
GSEA identified important pathway and
GO in gastric cancer
We assumed that the 50 differently expressed lncRNAs
we identified might be involved in various cellular pathways in
gastric cancer. To figure out the issue, we firstly run GSEA to
identify the dysregulated pathways and BP in gastric
adenocarcinoma. Through KEGG pathways, we found 9 upregulated
enrichment pathways, including DNA replication, Proteasome,
Nucleotide excision repair and 11 downregulated enrichment
pathways, including Neuroactive ligand-receptor interaction,
Pancreatic secretion, Gastric acid secretion and so on (Fig. 1C). Further, we also conducted GO BP
analysis to assign pathways and functionally classified the
dysregulated genes. As shown in Fig.
1D, 20 GO BP term, such as mitotic cell cycle phase transition,
and ncRNA metabolic process, were significantly enriched.
Construct lncRNA-mRNA co-expression
network and predict the function of key lncRNAs
Co-expression analyses of protein-coding RNAs and
lncRNAs reflect the potential function of lncRNAs (22). To further confirm the functions of
lncRNAs in gastric cancer, we used WGCNA to construct a
co-expression network for both mRNAs and lncRNAs that were
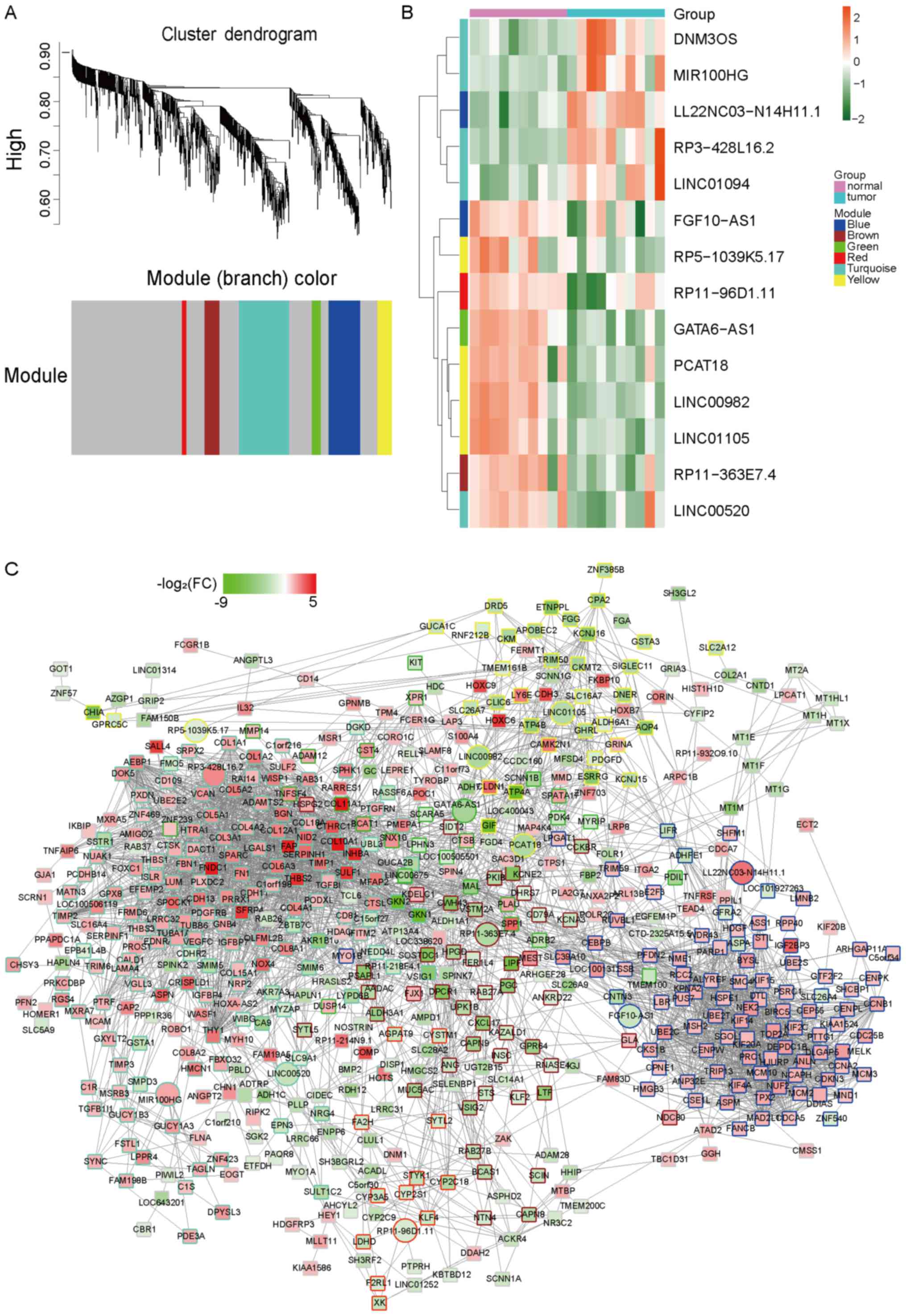
identified as differentially expressed. We identified 6
co-expression modules in which the highly co-expressed mRNAs and
lncRNAs were clustered in the same module (Fig. 2A) and we specifically identified 14
differently expressed lncRNAs in all the modules (Fig. 2B and Table
I). The co-expression network from WGCNA as showed in Fig. 2C and hub-genes in different modules
were listed in Table II. In order to
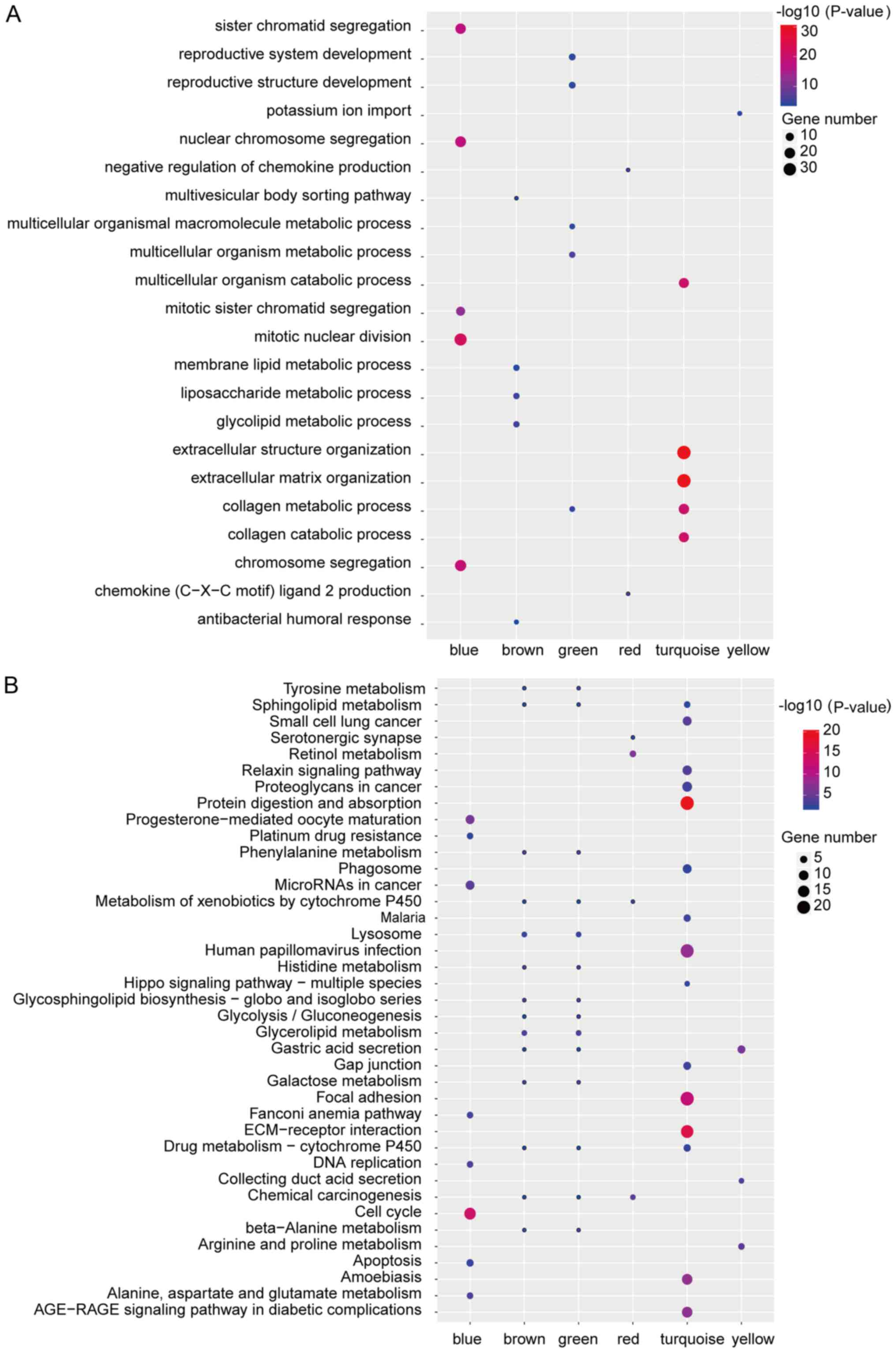
further explore the functions of the module, we performed GO
enrichment and KEGG pathway analysis and found several dysregulated
pathways, such as mitotic nuclear division, extracellular structure
organization, extracellular matrix organization, ECM-receptor
interaction or cell cycle, which were key signal pathways in cancer
generation or development (Fig. 3A and
B).
 | Table I.lncRNAs in different modules. |
Table I.
lncRNAs in different modules.
| Gene symbol | Chromosome | Ensembl ID | FC | FDR | Module |
|---|
|
LL22NC03-N14H11.1 |
chr22:15,823,197–15,823,890 |
ENST00000608286.1 | 5.37 | 0.0029 | Blue |
| FGF10-AS1 |
chr5:44,388,732–44,413,989 |
ENST00000502457.1 | 0.29 | 0.0466 | Blue |
| RP11-363E7.4 |
chr9:19,453,209–19,455,173 |
ENST00000563205.1 | 0.15 | 0.0197 | Brown |
| GATA6-AS1 |
chr18:22,166,898–22,168,968 |
ENST00000583490.1 | 0.10 | 0.0015 | Green |
| RP11-96D1.11 |
chr16:68,225,969–68,229,145 |
ENST00000571197.1 | 0.42 | 0.0464 | Red |
| RP3-428L16.2 |
chr6:160,990,318–160,992,342 |
ENST00000608721.1 | 4.20 | 0.0051 | Turquoise |
| DNM3OS |
chr1:172,136,531–172,144,794 |
ENST00000417354.2 | 2.39 | 0.0143 | Turquoise |
| LINC01094 |
chr4:78,646,186–78,682,392 |
ENST00000504675.5 | 2.68 | 0.0233 | Turquoise |
| LINC00520 |
chr14:55,782,067–55,796,688 |
ENST00000560336.6 | 0.26 | 0.0239 | Turquoise |
| MIR100HG |
chr11:122,180,338–122,367,973 |
ENST00000527474.5 | 2.79 | 0.0308 | Turquoise |
| LINC00982 |
chr1:3059615-3062531 |
ENST00000606861.1 | 0.13 | 0.0026 | Yellow |
| LINC01105 |
chr2:5,982,571–6,001,275 |
ENST00000450794.1 | 0.16 | 0.0063 | Yellow |
| PCAT18 |
chr18:26,687,621–26,703,638 |
ENST00000579458.1 | 0.14 | 0.0215 | Yellow |
| RP5-1039K5.17 |
chr22:37,950,965–37,951,778 |
ENST00000609976.1 | 0.46 | 0.0472 | Yellow |
| Table II.Hub-genes in different modules. |
Table II.
Hub-genes in different modules.
| Node name | FC | Module | Type | Node number |
|---|
| THY1 | 2.410 | Turquoise | mRNA | 74.000 |
| KPNA2 | 1.313 | Blue | mRNA | 59.000 |
| PSAPL1 | −4.757 | Brown | mRNA | 40.000 |
| ATP4A | −7.860 | Yellow | mRNA | 38.000 |
| BCAT1 | 1.863 | Green | mRNA | 36.000 |
Identify lncRNAs that is associated
with apparent clinical outcome
To further investigate the role of these lncRNAs in
gastric cancer, we compared the expression of the lncRNAs from
different modules, and chose the lncRNAs which changed similarly in
different datasets for further analysis (Table III). Then we used GSE15459 to
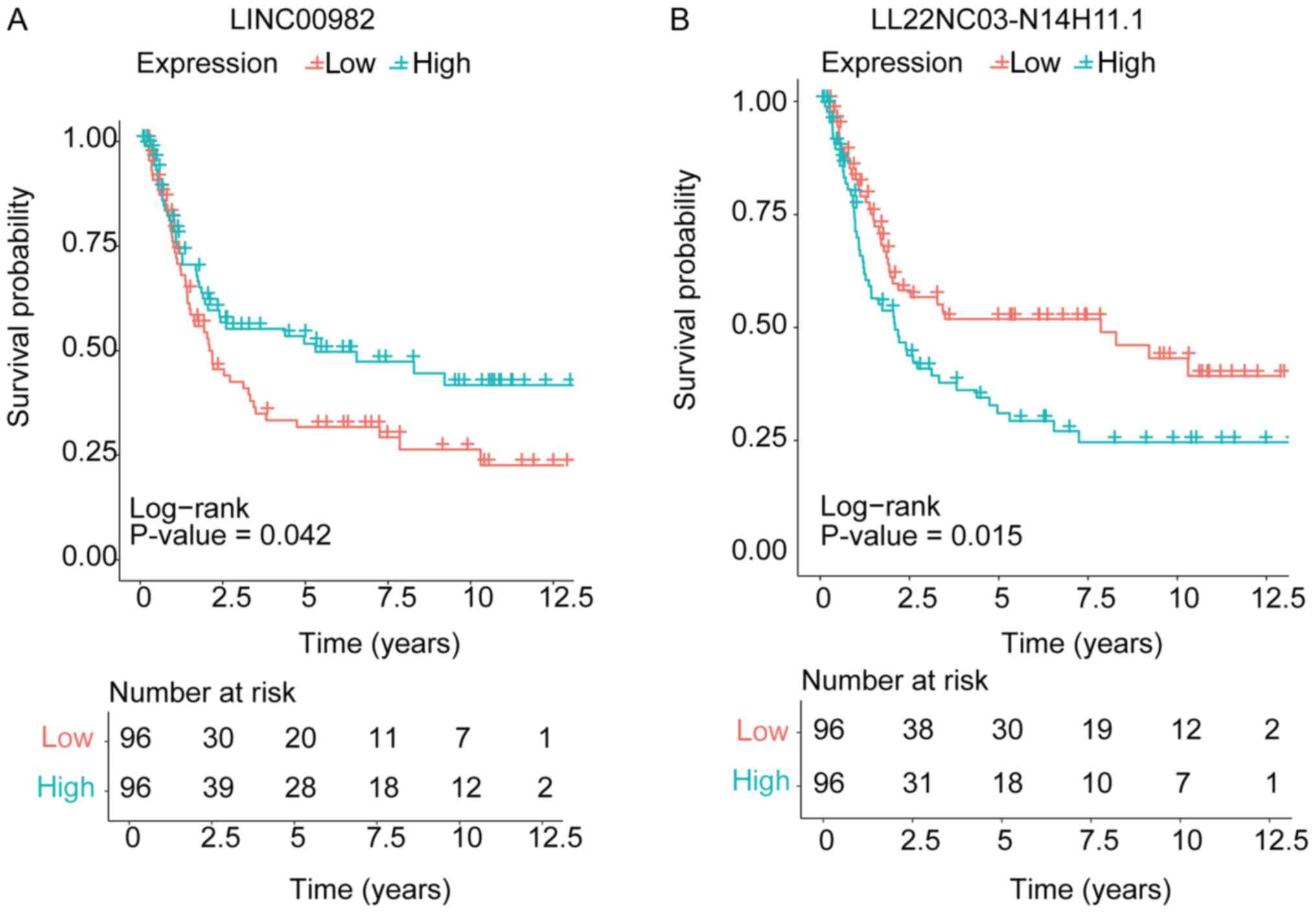
identify lncRNAs associated with clinical outcome (19). Univariate cox survival analysis was
performed to analyze the association of clinico-pathological
variables, including sex, age, clinical stage, and the lncRNAs'
expression with clinical prognosis. Moreover, further multivariate
cox analysis showed that LINC00982 expression (P=0.026) and
LL22NC03-N14H11.1 expression (P=0.015) were independent prognostic
indicators for gastric cancer patients' overall survival (Table IV). Kaplan-Meier survival analysis
was used to show the relationship of the two lncRNAs expression
level and pathology grade on patients' survival time (Fig. 4A and B).
| Table III.The expression of lncRNA in different
datastes. |
Table III.
The expression of lncRNA in different
datastes.
|
| GSE19826 | GSE13911 | GSE79973 |
|---|
|
|
|
|
|
|---|
| Name | FC | P-value | FC | P-value | FC | P-value |
|---|
| DNM3OS | 1.828 | 0.009 | 1.108 | 0.461 | 2.389 | <0.001 |
| FGF10-AS1 | 0.722 | 0.453 | 0.710 | 0.149 | 0.293 | 0.004 |
| GATA6-AS1 | 0.249 | 0.001 | 0.166 | <0.001 | 0.100 | <0.001 |
| LINC00520 | 0.524 | 0.001 | 0.700 | <0.001 | 0.258 | 0.001 |
| LINC00982 | 0.187 | <0.001 | 0.171 | <0.001 | 0.131 | <0.001 |
| LINC01094 | 1.822 | 0.139 | 1.608 | 0.005 | 2.684 | 0.001 |
| LINC01105 | 0.262 | 0.007 | 0.270 | <0.001 | 0.159 | <0.001 |
|
LL22NC03-N14H11.1 | 2.642 | 0.027 | 3.064 | <0.001 | 5.375 | <0.001 |
| MIR100HG | 2.597 | 0.002 | 1.388 | 0.202 | 2.786 | 0.002 |
| PCAT18 | 0.263 | 0.001 | 0.410 | <0.001 | 0.142 | 0.001 |
| RP11-363E7.4 | 0.234 | 0.003 | 0.146 | <0.001 | 0.146 | 0.001 |
| RP11-96D1.11 | 0.489 | <0.001 | 0.662 | <0.001 | 0.425 | 0.004 |
| RP3-428L16.2 | 4.998 | <0.001 | 3.849 | <0.001 | 4.198 | <0.001 |
| RP5-1039K5.17 | 0.323 | 0.002 | 0.433 | <0.001 | 0.463 | 0.004 |
| Table IV.Univariate and multivariate analyses
of the association between overall survival of 193 patients with
gastric cancer and prognostic factors by Cox proportional hazard
models. |
Table IV.
Univariate and multivariate analyses
of the association between overall survival of 193 patients with
gastric cancer and prognostic factors by Cox proportional hazard
models.
|
| Univariate
analysis | Multivariate
analysis |
|---|
|
|
|
|
|---|
| Variable | HR | 95% CI | P-value | HR | 95% CI | P-value |
|---|
| Age | 1 | 0.98–1.02 | 0.97 | 1 | 0.99–1.02 | 0.706 |
| Gender (male vs.
female) | 0.713 | 0.46–1.10 | 0.127 | 1.13 | 0.71–1.81 | 0.606 |
| Clinical stage
(III+IV vs. I+II) | 6.521 | 3.60–11.82 |
<0.001c | 7.69 | 4.07–14.54 |
<0.001c |
|
LL22NC03-N14H11.1 | 1.29 | 1.11–1.50 |
<0.001c | 1.23 | 1.04–1.46 | 0.015a |
| RP3-428L16.2 | 1.2 | 1.06–1.35 | 0.004b | 0.95 | 0.79–1.14 | 0.594 |
| LINC01105 | 0.7 | 0.54–0.92 | 0.010a | 0.97 | 0.71–1.34 | 0.859 |
| LINC00982 | 0.74 | 0.59–0.93 | 0.011a | 0.7 | 0.51–0.96 | 0.026a |
| LINC01094 | 1.32 | 1.06–1.64 | 0.013a | 0.98 | 0.76–1.27 | 0.889 |
| MIR100HG | 1.15 | 1.03–1.28 | 0.014a | 1.05 | 0.84–1.3 | 0.672 |
| DNM3OS | 1.2 | 1.03–1.41 | 0.020a | 1.2 | 0.88–1.63 | 0.258 |
| GATA6-AS1 | 0.85 | 0.73–0.98 | 0.028a | 1 | 0.81–1.24 | 0.980 |
| RP11-363E7.4 | 0.86 | 0.75–0.99 | 0.032a | 1.06 | 0.87–1.3 | 0.560 |
Potential mechanisms of LINC00982 and
LL22NC03-N14H11.1 in the regulation of gastric cancer cells
Next, we explored the potential mechanisms of
LINC00982 and LL22NC03-N14H11.1 in the regulation of gastric cancer
cells. Firstly, we forecasted the two lncRNAs potential target
genes by cis or trans regulation analysis (Tables V and VI). We found LINC00982 could regulate
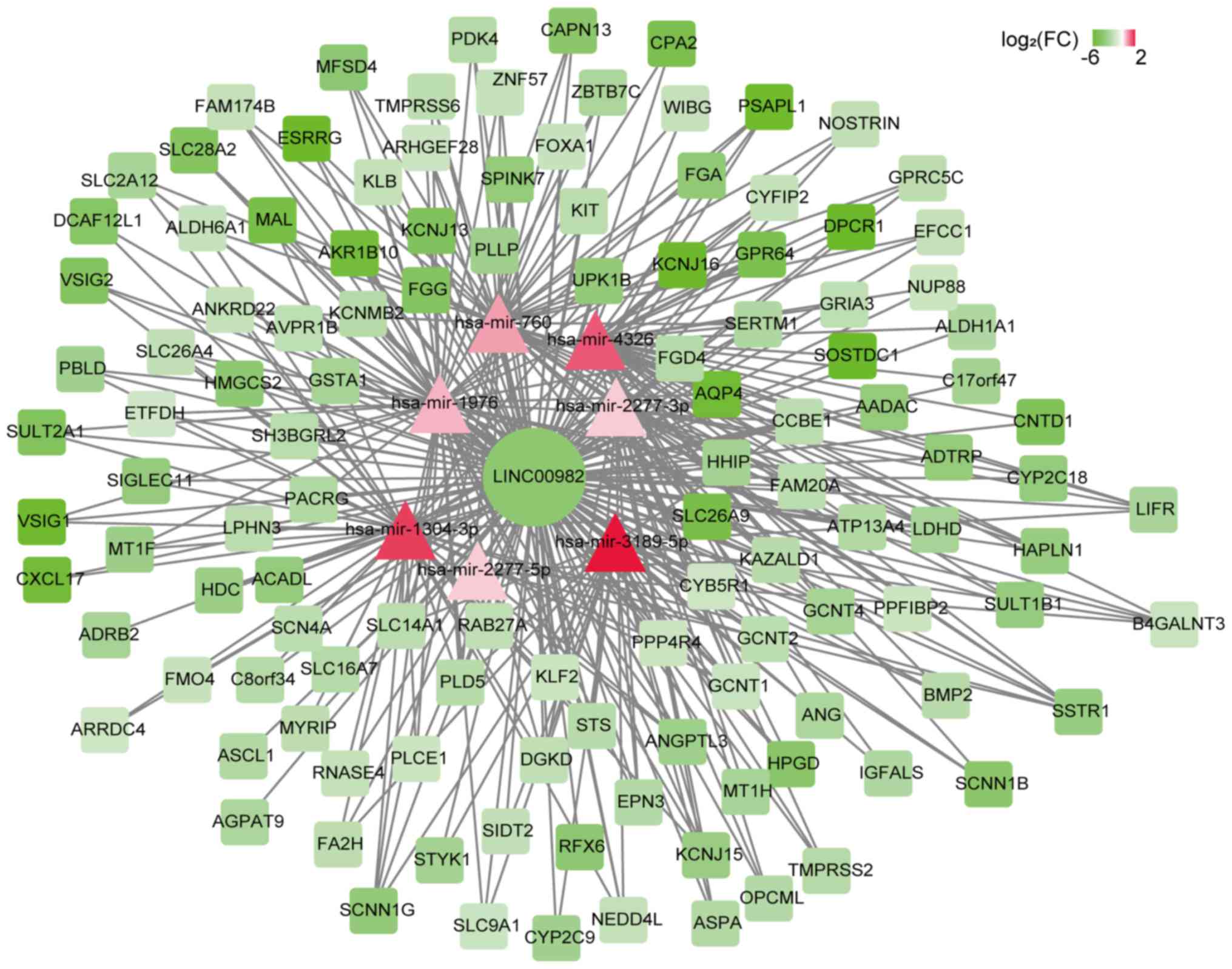
target genes through potential ceRNA network (Fig. 5), but not the same with
LL22NC03-N14H11.1. The microRNAs that could bind to
LL22NC03-N14H11.1 were not differently expressed in gastric
cancers, and these microRNAs were not co-expressed with the target
genes of LL22NC03-N14H11.1. This was not accord with the principles
of ceRNA network and suggested that LL22NC03-N14H11.1 could not
regulate gastric cancer through ceRNA regulation. Then we used
enrichment analysis based on the potential target genes to forecast
potential pathways that the two lncRNAs may be involved in. The
results showed that both LINC00982 and LL22NC03-N14H11.1 may be
involved in drug metabolism, ECM-receptor interaction, cell cycle,
focal adhesion and other pathway in cancer (Table VII). According to previous studies,
cell cycle, ECM-receptor interaction and focal adhesion are
important pathways to regulate cell proliferation which play an
important role in different cancers, so we further inquiried the
ability of the two lncRNAs in regulating tumor cell proliferation
(23,24).
| Table V.Potential cis-regulation target
genes. |
Table V.
Potential cis-regulation target
genes.
| lncRNA | Target genes |
|---|
|
LL22NC03-N14H11.1 | TPTEP1, CECR7,
OR11H1, POTEH, POTEH-AS1, LINC01297, DUXAP8, BMS1P18, BMS1P17,
BMS1P22, CCT8L2, ANKRD62P1-PARP4P3, LOC101929350, XKR3, HSFY1P1,
GAB4, IL17RA, CECR6, LOC100996342, CECR5 |
| LINC00982 | PRKCZ, PRDM16,
CFAP74, MORN1, PLCH2, MMEL1, TTC34, MEGF6, TP73, GABRD,
LOC105378591, FAAP20, SKI, LOC100129534, RER1, PEX10, PANK4, HES5,
LOC115110, TNFRSF14, LOC100996583, FAM213B, ACTRT2, LINC00982,
MIR4251, ARHGEF16, MIR551A, TPRG1L, WRAP73, TP73-AS1, CCDC27,
SMIM1, LRRC47, CEP104, DFFB, C1orf174, LINC01134, LINC01346 |
| Table VI.Potential trans-regulation target
genes. |
Table VI.
Potential trans-regulation target
genes.
| lncRNA | Target genes |
|---|
| LINC00982 | NONO, PABPC1,
EIF4B, FUS, SNRPA, Pum2, ACO1, SFRS9, MBNL1, KHSRP, YTHDC1, Vts1,
QKI, RBMX, SFRS13A, SFRS1, ELAVL1, KHDRBS3 |
|
LL22NC03-N14H11.1 | HNRNPA1, NONO,
sap-49, PABPC1, a2bp1, EIF4B, FUS, SFRS9, MBNL1, Vts1, RBMX, SFRS1,
KHDRBS3, ELAVL1, SFRS13A |
| Table VII.Correlation pathway of LINC00982 and
LL22NC03-N14H11.1. |
Table VII.
Correlation pathway of LINC00982 and
LL22NC03-N14H11.1.
| A, Positive
correlation KEGG pathway of LINC00982 |
|---|
|
|---|
| Term | Count | P-value | Genes |
|---|
| hsa00982: Drug
metabolism | 10 | 1.35E-07 | FMO4, GSTA1, FMO5,
GSTA3, CYP2C18, CYP2C9, ADH1C, ADH7, ADH1A, UGT2B15, ALDH3A1 |
| hsa00980:
Metabolism of xenobiotics by cytochrome P450 | 9 | 1.39E-06 | GSTA1, GSTA3,
CYP2C18, CYP2C9, ADH1C, ADH7, ADH1A, UGT2B15, AKR1C1, ALDH3A1 |
| hsa00830: Retinol
metabolism | 7 | 9.11E-05 | ALDH1A1, RDH12,
CYP2C18, CYP2C9, ADH1C, ADH7, ADH1A, UGT2B15 |
| hsa00591: Linoleic
acid metabolism | 4 | 0.006654721 | CYP2C18, CYP2C9,
AKR1B10, PLA2G1B |
| hsa00350: Tyrosine
metabolism | 4 | 0.022960524 | GOT1, ADH1C, ADH7,
ADH1A, ALDH3A1 |
|
| B, Negative
correlation KEGG pathway of LINC00982 |
|
| Term | Count | P-value | Genes |
|
| hsa04512:
ECM-receptor interaction | 11 | 3.99E-08 | COL4A2, COL4A1,
COMP, COL6A3, COL3A1, COL1A2, HSPG2, ITGA2, COL5A2, THBS2,
SPP1 |
| hsa04510: Focal
adhesion | 10 | 5.72E-04 | COL4A2, COL4A1,
COMP, COL6A3, COL3A1, COL1A2, ITGA2, COL5A2, THBS2, SPP1 |
| hsa04110: Cell
cycle | 6 | 0.016438397 | CCNB1, MAD2L1,
PTTG1, CCNA2, CDC25A, CDC25B |
| hsa04914:
Progesterone-mediated oocyte maturation | 5 | 0.019154758 | CCNB1, MAD2L1,
CCNA2, CDC25A, CDC25B |
| hsa05130:
Pathogenic escherichia coli infection | 4 | 0.030393085 | ARPC1B, CLDN1,
TUBB6, CD14 |
|
| C, Positive
correlation KEGG pathway of LL22NC03-N14H11.1 |
|
| Term | Count | P-value | Genes |
|
| hsa04512:
ECM-receptor interaction | 9 | 6.49E-06 | COL4A2, COL4A1,
COL6A3, COL3A1, COL1A2, ITGA2, COL5A2, THBS2, SPP1 |
| hsa04110: Cell
cycle | 9 | 1.18E-04 | CCNB1, E2F3,
MAD2L1, MCM2, PTTG1, MCM3, CCNA2, CDC25A, CDC25B |
| hsa04510: Focal
adhesion | 9 | 0.002793081 | COL4A2, COL4A1,
COL6A3, COL3A1, COL1A2, ITGA2, COL5A2, THBS2, SPP1 |
| hsa05222: Small
cell lung cancer | 5 | 0.018703322 | CKS1B, E2F3,
COL4A2, COL4A1, ITGA2 |
| hsa04914:
Progesterone-mediated oocyte maturation | 5 | 0.020218016 | CCNB1, MAD2L1,
CCNA2, CDC25A, CDC25B |
|
| D, Negative
correlation KEGG pathway of LL22NC03-N14H11.1 |
|
| Term | Count | P-value | Genes |
|
| hsa00982: Drug
metabolism | 7 | 4.21E-05 | CYP3A43, FMO4,
FMO5, CYP2C18, CYP2C9, ADH7, ADH1A |
| hsa00830: Retinol
metabolism | 6 | 2.38E-04 | CYP3A43, ALDH1A1,
CYP2C18, CYP2C9, ADH7, ADH1A |
| hsa04960:
Aldosterone-regulated sodium reabsorption | 5 | 8.60E-04 | NR3C2, NEDD4L,
SCNN1G, SCNN1B, SCNN1A |
| hsa00591: Linoleic
acid metabolism | 4 | 0.003074157 | CYP3A43, CYP2C18,
CYP2C9, PLA2G1B |
| hsa00980:
Metabolism of xenobiotics by cytochrome P450 | 5 | 0.00356804 | CYP3A43, CYP2C18,
CYP2C9, ADH7, ADH1A |
LINC00982 and LL22NC03-N14H11.1 have
opposite roles in proliferation of gastric cancer cells
To further confirm the functions of LINC00982 and
LL22NC03-N14H11.1 in the regulation of gastric cancer cells'
proliferation, we decided to construct overexpressed or
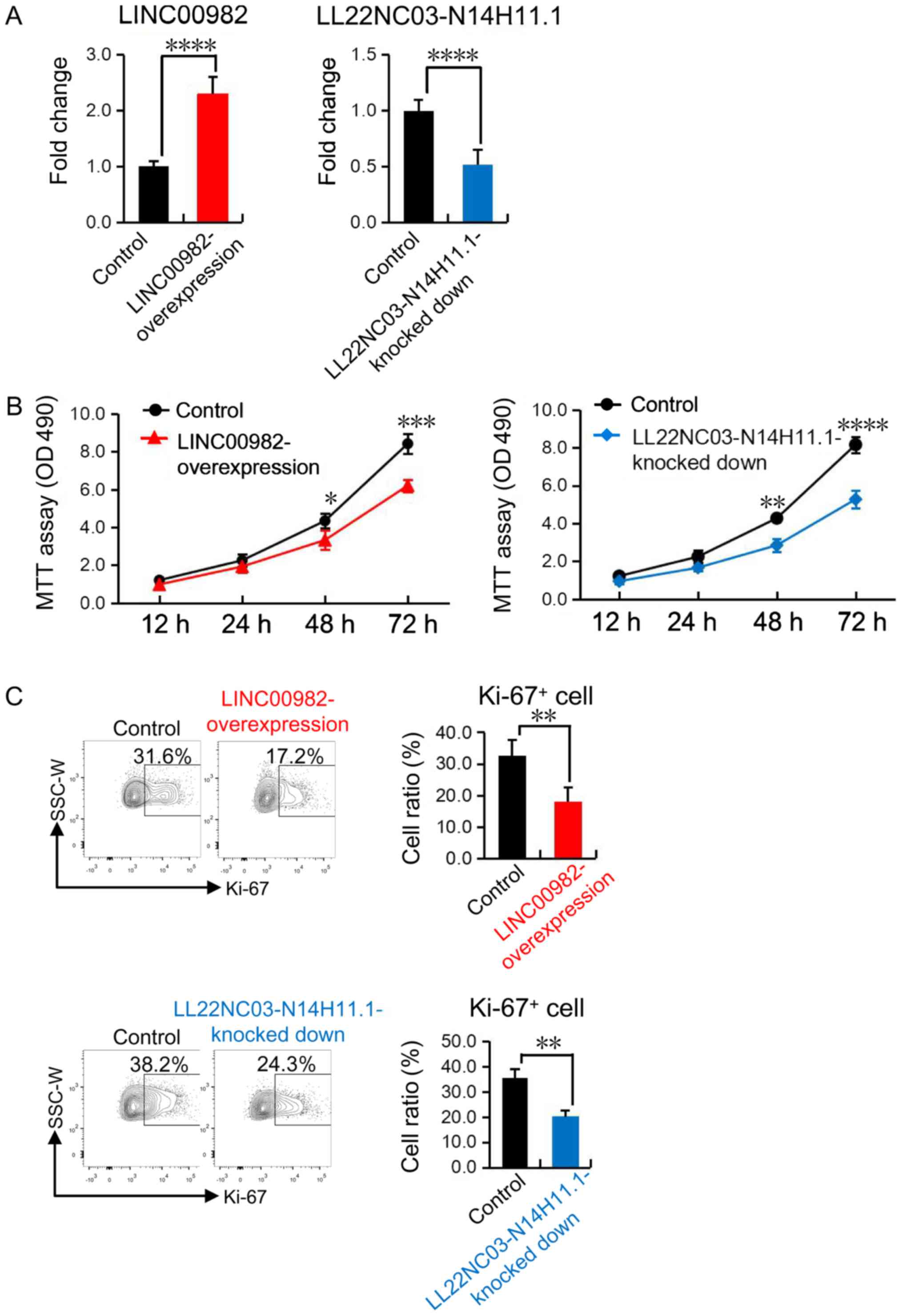
down-regualted cell lines for the two lncRNAs. As our previous
bioinformatics analysis revealed that LINC00982 was reduced while
LL22NC03-N14H11.1 was increased in gastric cancer cells (Table III), we overexpressed LINC00982 and
knocked down LL22NC03-N14H11.1 in gastric cell lines SGC-7901
respectively (Fig. 6A). Then we
performed MTT assay and flow cytometric analysis and found that the
proliferation as well as the expression of ki-67, which acts as a
key marker for cell mitosis, were impaired in LINC00982
overexpressed and LL22NC03-N14H11.1 down-regulated SGC-7901 cell
lines respectively (Fig. 6B and C).
These experimental results provided solid evidence to support our
conclusion that LINC00982 inhibited while LL22NC03-N14H11.1
promoted the proliferation of gastric cancer cells.
Discussion
LncRNAs are endogenous, lncRNAs that participate in
the regulation of diverse cellular process and a great many
evidence demonstrates that the abnormal expression of lncRNAs might
serve as potential diagnostic and prognostic factors for cancers
(25,26).
In this study, we made a deep analysis of the data
from He et al study (8), and
identified 50 differently expressed lncRNAs in gastric cancer
cells. Then we carried out the GSEA and found that several cellular
pathways were changed in gastric cancer, which suggested that the
differently expressed lncRNAs might be involved in these
pathways.
In order to further verify the functions of these
differently expressed lncRNAs, we used WGCNA to construct a
co-expression network including these lncRNAs and the associated
mRNA, and figured out 6 co-expression modules, which involved 14
lncRNAs. The co-expression network revealed that these 14 lncRNAs
were functional, for they were related with various
protein-encoding mRNAs. To further elucidate the functions of these
lncRNAs, we performed KEGG pathway analyses and GO enrichment
analyses, and found the modules were tightly associated with
mitotic nuclear division, extracellular structure organization,
extracellular matrix organization, ECM-receptor interaction or cell
cycle, which were the key pathways involved in cancer generation
and development.
LncRNAs can be considered as independent indicators
for prognosis and we also identified two lncRNAs, LINC00982 and
LL22NC03-N14H11.1, associated with apparent clinical outcome in
gastric cancer. LINC00982 is located on chromosome 1p36.32 and has
two transcript variants. In previous study, Fei et al found
that LINC00982 functioned as an inhibitor of cancer cell's
proliferation and arrested cell cycle partly via regulating P15 and
P16 protein expressions (27). As a
result, they identified LINC00982 as a prognostic biomarker in
gastric cancer, which was consistent with our current results. This
also proved that our bioinformatics analysis process was highly
reliable.
On the other hand, we also identified
LL22NC03-N14H11.1 as a potential prognostic marker for gastric
cancer. LL22NC03-N14H11.1 is located on chromosome 22 and encodes a
transcript of 694 bp, however it has never been reported in any
study. Here in our study, we found it was highly expressed in
gastric cancer and formed the blue module in the co-expression
network we constructed, which was involved in the cell cycle
related cellular process, including sister chromatid segregation,
nuclear chromosome segregation, mitotic nuclear division,
chromosome segregation and so on. As a result, we assumed that
LL22NC03-N14H11.1 might promote the proliferation of cancer cells,
leading to the generation or development of gastric cancer.
To further explore the mechanisms of LINC00982 and
LL22NC03-N14H11.1 in the regulation of gastric cancer cells, we
performed in-depth analysis on the cis, trans or ceRNA regulation
of the lncRNAs in gastric cancer cells. It turned out that the
potential target genes of the two lncRNAs were mainly involved in
the proliferation of gastric cancer cells, which was consistent
with our previous analysis. Besides, in order to provide solid
evidence for their roles in the proliferation, we further reversed
the expression of LINC00982 and LL22NC03-N14H11.1 in gastric cell
linces SGC-7901 with genetic interference. The results completely
supported our conclusions that LINC00982 inhibited while
LL22NC03-N14H11.1 promoted the proliferation of gastric cancer
cells. We believe our findings possess important value, because we
may control the proliferation of gastric cancer cells artificially
by interfering the expression of LINC00982 and
LL22NC03-N14H11.1.
In all, our study contributed a comprehensive
knowledge about lncRNAs in gastric cancer and hopefully,
LL22NC03-N14H11.1 and LINC00982 might be potential prognostic
indicators and clinical targets for gastric cancer.
Acknowledgements
The authors would like to thank Dr Yao Yang (Army
Medical University, Chongqing, China) for technology
consulting.
Funding
No funding was received.
Authors' contributions
DQ and XZ conceived and designed the experiments.
DQ, QW, MW and XZ performed the experiments. DQ, QW, MW and XZ
analyzed the data. DQ and XZ supervised the study and wrote the
paper.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
lncRNA
|
Long non-coding RNAs
|
|
WGNCA
|
weighted gene co-expression network
analysis
|
|
GO
|
Gene Ontology
|
|
KEGG
|
Kyoto Encyclopedia of Genes and
Genomes
|
|
GEO
|
Gene Expression Omnibus
|
|
BP
|
biological process
|
|
FC
|
fold-change
|
|
FDR
|
false discovery rate
|
|
TCGA
|
The Cancer Genome Atlas
|
References
|
1
|
Shin HR, Carlos MC and Varghese C: Cancer
control in the Asia Pacific region: Current status and concerns.
Jpn J Clin Oncol. 42:867–881. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ilson DH: Advances in the treatment of
gastric cancer. Curr Opin Gastroenterol. 33:473–476. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sasaki K, Onodera S, Otsuka K, Satomura H,
Kurayama E, Kubo T, Takahashi M, Ito J, Nakajima M, Yamaguchi S, et
al: Validity of neoadjuvant chemotherapy with docetaxel, cisplatin,
and S-1 for resectable locally advanced gastric cancer. Med Oncol.
34:1392017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Beermann J, Piccoli MT, Viereck J and Thum
T: Non-coding RNAs in development and disease: Background,
mechanisms, and therapeutic approaches. Physiol Rev. 96:1297–1325.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kondo Y, Shinjo K and Katsushima K: Long
non-coding RNAs as an epigenetic regulator in human cancers. Cancer
Sci. 108:1927–1933. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhu J, Fu H, Wu Y and Zheng X: Function of
lncRNAs and approaches to lncRNA-protein interactions. Sci China
Life Sci. 56:876–885. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gibb EA, Brown CJ and Lam WL: The
functional role of long non-coding RNA in human carcinomas. Mol
Cancer. 10:382011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
He J, Jin Y, Chen Y, Yao HB, Xia YJ, Ma
YY, Wang W and Shao QS: Downregulation of ALDOB is associated with
poor prognosis of patients with gastric cancer. Onco Targets Ther.
9:6099–6109. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wen Q, Yang Y, Chen XH, Pan XD, Han Q,
Wang D, Deng Y, Li X, Yan J and Zhou J: Competing endogenous RNA
screening based on long noncoding RNA-messenger RNA co-expression
profile in Hepatitis B virus-associated hepatocarcinogenesis. J
Tradit Chin Med. 37:510–521. 2017. View Article : Google Scholar
|
|
10
|
Yu G, Wang LG, Han Y and He QY:
clusterProfiler: An R package for comparing biological themes among
gene clusters. OMICS. 16:284–287. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yu G, Wang LG, Yan GR and He QY: DOSE: An
R/Bioconductor package for disease ontology semantic and enrichment
analysis. Bioinformatics. 31:608–609. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Langfelder P and Horvath S: WGCNA: An R
package for weighted correlation network analysis. BMC
Bioinformatics. 9:5592008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang C, Peng L, Zhang Y, Liu Z, Li W,
Chen S and Li G: The identification of key genes and pathways in
hepatocellular carcinoma by bioinformatics analysis of
high-throughput data. Med Oncol. 34:1012017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mitchell RD III, Wallace AD, Hodgson E and
Roe RM: Differential expression profile of lncRNAs from primary
human hepatocytes following DEET and fipronil exposure. Int J Mol
Sci. 18:pii: E2104. 2017.
|
|
15
|
Cook KB, Kazan H, Zuberi K, Morris Q and
Hughes TR: RBPDB: A database of RNA-binding specificities. Nucleic
Acids Res. 39:(Database Issue). D301–D308. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chang TH, Huang HY, Hsu JB, Weng SL, Horng
JT and Huang HD: An enhanced computational platform for
investigating the roles of regulatory RNA and for identifying
functional RNA motifs. BMC Bioinformatics. 14 Suppl 2:S42013.
|
|
17
|
Chou CH, Shrestha S, Yang CD, Chang NW,
Lin YL, Liao KW, Huang WC, Sun TH, Tu SJ, Lee WH, et al: miRTarBase
update 2018: A resource for experimentally validated
microRNA-target interactions. Nucleic Acids Res. 46(D1): D296–D302.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang Q, Deng Y, Lai W, Guan X, Sun X, Han
Q, Wang F, Pan X, Ji Y, Luo H, et al: Maternal inflammation
activated ROS-p38 MAPK predisposes offspring to heart damages
caused by isoproterenol via augmenting ROS generation. Sci Rep.
6:301462016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ooi CH, Ivanova T, Wu J, Lee M, Tan IB,
Tao J, Ward L, Koo JH, Gopalakrishnan V, Zhu Y, et al: Oncogenic
pathway combinations predict clinical prognosis in gastric cancer.
PLoS Genet. 5:e10006762009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang YS, Ma LN, Sun JX, Liu N and Wang H:
Long non-coding RNA CPS1-IT1 is a positive prognostic factor and
inhibits epithelial ovarian cancer tumorigenesis. Eur Rev Med
Pharmacol Sci. 21:3169–3175. 2017.PubMed/NCBI
|
|
21
|
Jalali S, Gandhi S and Scaria V:
Navigating the dynamic landscape of long noncoding RNA and
protein-coding gene annotations in GENCODE. Hum Genomics.
10:352016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liao Q, Liu C, Yuan X, Kang S, Miao R,
Xiao H, Zhao G, Luo H, Bu D, Zhao H, et al: Large-scale prediction
of long non-coding RNA functions in a coding-non-coding gene
co-expression network. Nucleic Acids Res. 39:3864–3878. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Schwartz MA and Assoian RK: Integrins and
cell proliferation: Regulation of cyclin-dependent kinases via
cytoplasmic signaling pathways. J Cell Sci. 114:2553–2560.
2001.PubMed/NCBI
|
|
24
|
Lu P, Weaver VM and Werb Z: The
extracellular matrix: A dynamic niche in cancer progression. J Cell
Biol. 196:395–406. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Schmitt AM and Chang HY: Long noncoding
RNAs in cancer pathways. Cancer Cell. 29:452–463. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhu X, Tian X, Yu C, Shen C, Yan T, Hong
J, Wang Z, Fang JY and Chen H: A long non-coding RNA signature to
improve prognosis prediction of gastric cancer. Mol Cancer.
15:602016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fei ZH, Yu XJ, Zhou M, Su HF, Zheng Z and
Xie CY: Upregulated expression of long non-coding RNA LINC00982
regulates cell proliferation and its clinical relevance in patients
with gastric cancer. Tumour Biol. 37:1983–1993. 2016. View Article : Google Scholar : PubMed/NCBI
|