Introduction
In the current research, cancer statistics show that
invasive breast cancer is projected to have 234,190 new cases and
deaths, for which 231,840 will be women (1). Approximately 40,290 women will die from
invasive breast cancer in 2018 (2).
Although the percentage of mortalities has decreased over the last
few years, breast cancer still ranks as the second leading cause of
cancer death in women; treatments usually entail invasive
surgeries, including breast-conserving surgeries and mastectomies
(1). From a clinical standpoint,
invasive ductal carcinoma (IDC) is the most common form of breast
cancer, affecting 50–75% of breast cancer diagnoses (2). Breast cancer is a heterogeneous disease,
and research has shown that the top mutated genes in breast
carcinomas and carcinomas in situ are
Phosphatidylinositol-4,5-Bisphosphate 3-Kinase Catalytic Subunitα
(PIK3CA), Tumor Protein P53 (TP53), and E-cadherin (CDH1) (3). However, this investigation shows a link
between protein kinase C-ζ (PKC-ζ) protein overexpression and
breast cancer development, particularly in invasive behavior. For
this reason, breast cancer biomarkers are of interest, as they may
help to predict breast cancer incidents and contribute to better
therapeutic regimens.
The complete understanding of histological and
pathological features of the lobular and ductal carcinomas are far
from full elucidation. In an investigation by Ruibal et al,
the authors concluded that, in the absence of the axillary node,
lobular carcinomas had a higher concentration of breast cancer
estrogen-inducible protein (pS2) than ductal carcinomas (4). Lobular carcinomas were also shown to
have a higher frequency of diploidy, which suggests that lobular
carcinomas are less aggressive and grow slower (4).
An overexpression of PKC-ζ protein promotes
carcinogenesis by stimulating cancer cell proliferation through
pathways such as the Nuclear factor-κB (NF-κB), which plays an
essential role in cancer initiation and progression (5). Previous studies suggest that PKC-ζ is a
regulatory factor for the nuclear translocation of NF-κB that in
turn represses E-cadherin (6,7). Although the link between the loss of
E-cadherin and cancer prognosis remains ambiguous, recent findings
showed that E-cadherin possesses a vital tumor suppressive role
(8). Moreover, some researchers have
paralleled PKC-ζ to the phosphorylation of the Inhibitor of κB
kinase (IKK) complex, which in turn phosphorylates the Inhibitor of
κB (IκB) and triggers IκB degradation (7,9). The
degradation of IκB releases NF-κB, allowing its translocation from
the cytosol into the nucleus, where it functions as a transcription
factor (9). The transcription factor
applies explicitly to targets such as apoptosis regulators and
stress response genes. Furthermore, studies show that NF-κB also
plays a role in epithelial to mesenchymal transition, a crucial
carcinogenic event (7).
PKC-ζ has also been linked to metastatic behaviors
of cancer cells. In a study by Islam et al, the Ras-related
C3 botulinum toxin substrate 1 (Rac1)/Pak1/β-Catenin signaling
cascade in colorectal cancer cell lines was evaluated after the
inhibition of PKC-ζ (10). The
knockdown of PKC-ζ decreased the nuclear translocation of β-Catenin
which ultimately leads to reduced colorectal cell proliferation and
metastasis (10). These data were
further supported by another investigation performed by Wu et
al, which determined that inhibition of PKC-ζ in breast cancer
cell lines decreased adhesion and actin polymerization (11). These studies advocate the theory that
PKC-ζ as a critical component of the invasive behaviors of cancer
cells.
Although PKC-ζ has been studied in invasive breast
cancers (most commonly ductal), there have been no examinations of
PKC-ζ expression in other types of breast cancer (such as
carcinomas in situ). An investigation by Lin et al
confirmed that atypical PKC isoforms were elevated in breast cancer
tissues (IDC, specifically) when compared to adjacent healthy
breast tissue (12). Additionally,
Schöndorf et al determined that antineoplastic agents affect
the activation of PKC in IDC breast cancer tumors (13). In this study, our focus was to further
investigate the PKC-ζ expression profile in the four histological
subtypes of breast cancer such as lobular carcinoma in situ
(LCIS), invasive lobular carcinoma (ILC), ductal carcinoma in
situ (DCIS) and IDC. We also evaluated the difference in the
PKC-ζ expression among healthy, invasive and non-invasive tissues.
Moreover, the invasive characteristics of MDA-MB-231 breast cancer
cells were examined upon the inhibition of PKC-ζ. We found that
PKC-ζ is overexpressed in IDC and ILC tissue specimens compared to
other subtypes. In addition, the inhibition of PKC-ζ decreased the
invasion of MDA-MB-231 breast cancer cells.
Materials and methods
Specimen collection and tissue
fractionation
The NCI-supported Cooperative Human Tissue Network
(CHTN; Birmingham, AL, USA) collected and provided the breast
tissue samples (normal, LCIS, ILC, DCIS, and IDC). The specimens
were collected and snap-frozen in liquid nitrogen or placed in dry
ice and stored in a liquid nitrogen vapor phase freezer (−196°C),
where the tissues stayed until shipment. The tissues were shipped
in dry ice. Formalin-fixed paraffin-embedded tissues (FFPE) were
also provided for immunohistochemistry staining. Tissues were
selected based on their histological features (normal, LCIS, ILC,
DCIS, and IDC). The mean age of patient samples collected was 51
years and the collection period of the samples was 2001–2015.
Normal tissues were selected from breast reduction patients with no
previous diagnosis of cancer or the area adjacent to a patient's
malignant tumors. Patients with DCIS and LCIS were selected based
on the lack of invasive tissue adjacent to the extraction site;
some tissues were taken from patients with invasive tissues in the
opposite breast.
The tissue specimens were then resuspended and
sonicated for 3×5 sec cycles on ice in 1 ml of homogenization
buffer (Pierce® Immuno Precipitation Lysis Buffer, 25 mM
Tris-HCl pH 7.4, 150 mM NaCl, 1 mM EDTA, 1% NP-40 and 5% glycerol;
Thermo Fisher Scientific, Inc., Waltham, MA, USA), followed by,
centrifugation at 12,879 × g for 15 min at 4°C to obtain cell
extracts. Subsequently, 250 µl of albumin removal resin
(Pierce™ Albumin Depletion kit; Thermo Fisher
Scientific, Inc.) was added to the lysate and left at 4°C
overnight. The samples were further centrifuged at 12,879 × g for
15 min at 4°C, the supernatant was subsequently collected, and
protein content was measured according to Pierce® 660 nm
Protein Assay reagent protocol (Thermo Fisher Scientific, Inc.) and
NanoDrop 2000 (Thermo Fisher Scientific, Inc., Wilmington, DE,
USA).
Cell culture
The metastatic breast cancer cell line, MDA-MB-231
was obtained from American Type Tissue Culture Collection (ATCC;
Manassas, VA, USA). The MDA-MB-231 cells were sub-cultured and
maintained in T75 flasks containing Dulbecco's modified Eagle's
media (DMEM; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA), supplemented with 10% fetal bovine serum (FBS) and 1%
antibiotics (penicillin 10 U/ml and streptomycin 10 mg/ml). Cells
were incubated at 37°C and 5% CO2. Cells were used at
70–80% confluency for the experiments.
Western blot analysis
Like tissue lysates, MDA-MB-231 cell extracts were
also prepared after the addition of cell lysis buffer, sonication,
and centrifugation (at 12,879 × g for 15 min at 4°C). For western
blot analysis, an equal amount (20–30 µg) of protein from tissue
and cell lysates were loaded in 10% polyacrylamide gels and
separated by using sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE). The proteins were then transblotted
onto a 0.4 µM nitrocellulose membrane. Subsequently, the membranes
were incubated with 5% milk blocking solution, followed by, primary
solution of anti-PKC-ζ (SC-17781; 1:1,000; Santa Cruz
Biotechnology, Dallas, TX, USA; and 9372s; 1:1,000; Cell Signaling
Technology, Danvers, MA, USA), anti-E-cadherin (701134; 1:1,000;
Invitrogen; Thermo Fisher Scientific, Inc.), anti-Ras homolog gene
family member A (RhoA; ab54835, 1:4,000; Abcam, Cambridge, UK) and
anti-Rac1 (4651s; 1:1,000; Cell Signaling Technology) in 5% bovine
serum albumin (BSA). Finally, the membranes were incubated with
secondary antibodies. All the secondary antibodies (anti-rabbit and
anti-mouse) were obtained from Bio-Rad Laboratories, Hercules, CA,
USA (cat. no. 170-6515 and cat. no. 170-6516; 1:2,000). The
immunoreactive bands were then visualized by chemiluminescence
reaction, according to the manufacturer's instructions
(SuperSignal™ West Pico PLUS Chemiluminescent Substrate;
Thermo Fisher Scientific, Inc.). A monoclonal antibody to β-actin
(SC-1616; Santa Cruz Biotechnology) was used as a loading
control.
Densitometry
The densitometry was performed using Image J
(National Institutes of Health, Bethesda, MD, USA) software by the
subtraction of background noise from the density of each band to
derive the corrected intensity. All samples were normalized based
on the intensity of β-actin bands on each blot.
Immunohistochemistry
FFPE tissues received from CHTN were sent to the
Tissue Core, Moffitt Cancer Center (Tampa, FL, USA). Two different
patient tissues were selected for each subtype (normal, LCIS, ILC,
DCIS, and IDC) based on the criteria mentioned previously, with a
total N=10. Briefly, tissues were stained with haemotoxylin
and eosin (H&E), and pathology quality control (PQC) were
performed by a Tissue Core pathologist to confirm breast tissue
subtype and diagnosis. The slides were prepared and stained with
antibodies for PKC-ζ (ab59364; 1:1,000; rabbit polyclonal; Abcam).
A uterine carcinoma specimen was selected as a positive control
based on antibody data sheet and the Human Protein Atlas
recommendations. Slides were stained using a Ventana Discovery XT
automated system (Ventana Medical Systems, Tucson, AZ, USA) as per
the manufacturer's protocol with proprietary reagents. Briefly,
slides were deparaffinized on the automated system with EZ Prep
solution (Ventana Medical Systems). The heat-induced antigen
retrieval method (CC1 standard) was used with the PKC-ζ primary
rabbit antibody (Ventana Medical Systems). This antibody that
reacts to the human isoform of PKC-ζ was used at a 1:1,000
concentration in Dako antibody diluent (Dako; Agilent Technologies,
Inc., Santa Clara, CA, USA) and incubated for 32 min. The Ventana
OmniMap Anti-Rabbit Secondary Antibody (Ventana Medical Systems)
was used for 16 min. The detection system used was the Ventana
ChromoMap kit and slides were then counterstained with
haematoxylin. Slides were subsequently dehydrated and cover-slipped
as per normal laboratory protocol. The Moffitt Cancer Center Tissue
Core pathologist selected the optimal condition, titration, and
incubation time to be used on the control and the breast selected
slides. Subsequently, the pathologist evaluated the slides using
the combinative semi quantitative scores (score, 0–3) (14). Images were taken on a light microscope
Olympus BX51 (Olympus Corp., Tokyo, Japan).
Knockdown of PKC-ζ for invasion
pathway analysis
Human breast cancer cells MDA-MB-231 were grown in
100 mm plates and transfected with 20 nM of scrambled RNA and
siPRKCZ (5′-GCAUGAUGACGAGGAUAUUGACUGG-3′, SR303747A; Origene,
Rockville, MD, USA) for 48 h. Cells were lysed as previously
described and the lysates were run on western blots.
Cell invasion assay by crystal violet
staining of invaded cells
Cells were serum starved for 24 h, followed by
detachment and plating into the upper chamber of a 96-well 8 µm
Transwell permeable support, coated with 0.5X basement membrane
extract (BME; both Corning Inc., Corning, NY, USA) for the
evaluation of invasion. Serum (10%) containing media was loaded
into the receiver plate (lower chamber) as a chemoattractant.
MDA-MB-231 cells at the upper chamber were transfected with 20 nM
siPRKCZ for 24 h. Four experimental treatment groups for the cells
were performed: Control (non-treated), Si-Tran (transfection
reagent), scrambled siRNA (random RNA) and siRNA for PRKCZ (for
knockdown of PKC-ζ protein expression). The cells were treated with
the transfection reagent (Si-Tran) and universal scrambled RNA to
establish the effect of targeted small interfering RNA (siPRKCZ)
only. The invasive cells that passed into the lower chamber were
then fixed with 4% paraformaldehyde, stained with 2% crystal violet
in 2% ethanol, washed with distilled water and photographs were
captured after drying using a light microscope Motic AE31E.
Phalloidin staining of filamentous
actin (F-actin)
Human breast cancer cells MDA-MB-231
(1×104 cells) were grown in 2-well chamber slides,
followed by transfection with 20 nM universal scrambled RNA and
siPRKCZ for 24 h. In addition, cells were also evaluated with the
transfection reagent and without any treatment to establish the
targeted effect of PKC-ζ knockdown. Fixation was performed with 4%
paraformaldehyde. F-actin was subsequently stained with 1X
Phalloidin-iFluor 594 (Abcam) in 1% BSA-phosphate buffered saline
(PBS) solution for an hour at room temperature. Cells were washed,
counterstained with the nuclear stain 4′,6-diamidino-2-phenylindole
(DAPI; Invitrogen; Thermo Fisher Scientific, Inc.) and examined
under Nikon MICROPHOT-FX fluorescence microscope (Ex/Em=590/618).
Photographs were captured using ProgRes®Capture
2.9.0.1.
Statistical analysis
The statistical significance of the western blot
analysis data was evaluated by a Student's t-test (normal
N=32; LCIS N=3; ILC N=13; DCIS N=6; IDC
N=29; overall N=83 at P<0.05; standard error
represented) and the linear regression test (N=20;
R2 value) with GraphPad software (15). A one-way ANOVA was also used to
evaluate the western blot analysis data as well with the post-hoc
Tukey's HSD test (P<0.01), Scheffé multiple comparison
(P<0.05), Bonferroni (P<0.01) and Holm (P<0.01). The
contingency table Chi-squared statistical analysis (normal
N=32; LCIS N=3; ILC N=13; DCIS N=6; IDC
N=29; overall N=83 at P<0.00001) for the
expression of PKC-ζ and breast subtype was performed using the
Chi-Squared Test Calculator from Social Science Statistics
(16). Clinical parameters such as
estrogen receptor expression and Scarff-Bloom-Richardson grade
(presented in the pathology reports) were also investigated with
this statistical software (N=25 and N=22,
respectively). A one-way ANOVA was used to analyze the number of
cells invaded after crystal violet staining (P<0.05).
Results
PKC-ζ is overexpressed in malignant
breast tissues
PKC-ζ protein expression was measured in breast
samples with no tumor complication and malignant breast tissue
samples by western blot analysis. Our investigation showed a
correlation between the overexpression of PKC-ζ and malignant
breast cancer tissue (Fig. 1A and B;
Table III). It was challenging to
obtain DCIS (N=6) and LCIS (N=3) since these tissue
types have lower occurrence rates; hence the results reflect a
comparison of the two main subtypes: IDC, N=29, (50–75%
diagnoses) and ILC, N=13 (10–15% diagnoses). The westerns
represent the majority of samples. Normal tissue only had 3 of 32
samples with expression of PKC-ζ and DCIS only had 1 of 6 samples
with PKC-ζ present (Table II). Among
the IDC subtypes of malignant breast tissue, >74% of tissue
samples overexpressed PKC-ζ protein. Less than 5% of healthy breast
tissue samples exhibited PKC-ζ protein expression. Although a
paired Student's t-test (P<0.05) and one-way ANOVA (P<0.01)
showed no significant correlation between the overexpression of
PKC-ζ protein in malignant tissue (all four subtypes) and healthy
tissue, it did demonstrate a significant correlation between
healthy breast tissue and IDC (Tukey HSD P-value 0.0056404,
P<0.01; Scheffé P-value 0.0121289, P<0.05; Bonferroni and
Holm P-value 0.0063104, P<0.01) (Fig.
1B). According to the significance of the contingency table
(Table III), there is an
established association between the protein expression of PKC-ζ and
sample type (P<0.00001). However, the PKC-ζ expression could not
be statistically linked to the presence of ER and nuclear grade
(Table IV; Fig 3C). These data suggest that it is
unlikely that healthy breast tissue samples overexpress PKC-ζ
protein. Instead, PKC-ζ is overexpressed in malignant breast tissue
samples.
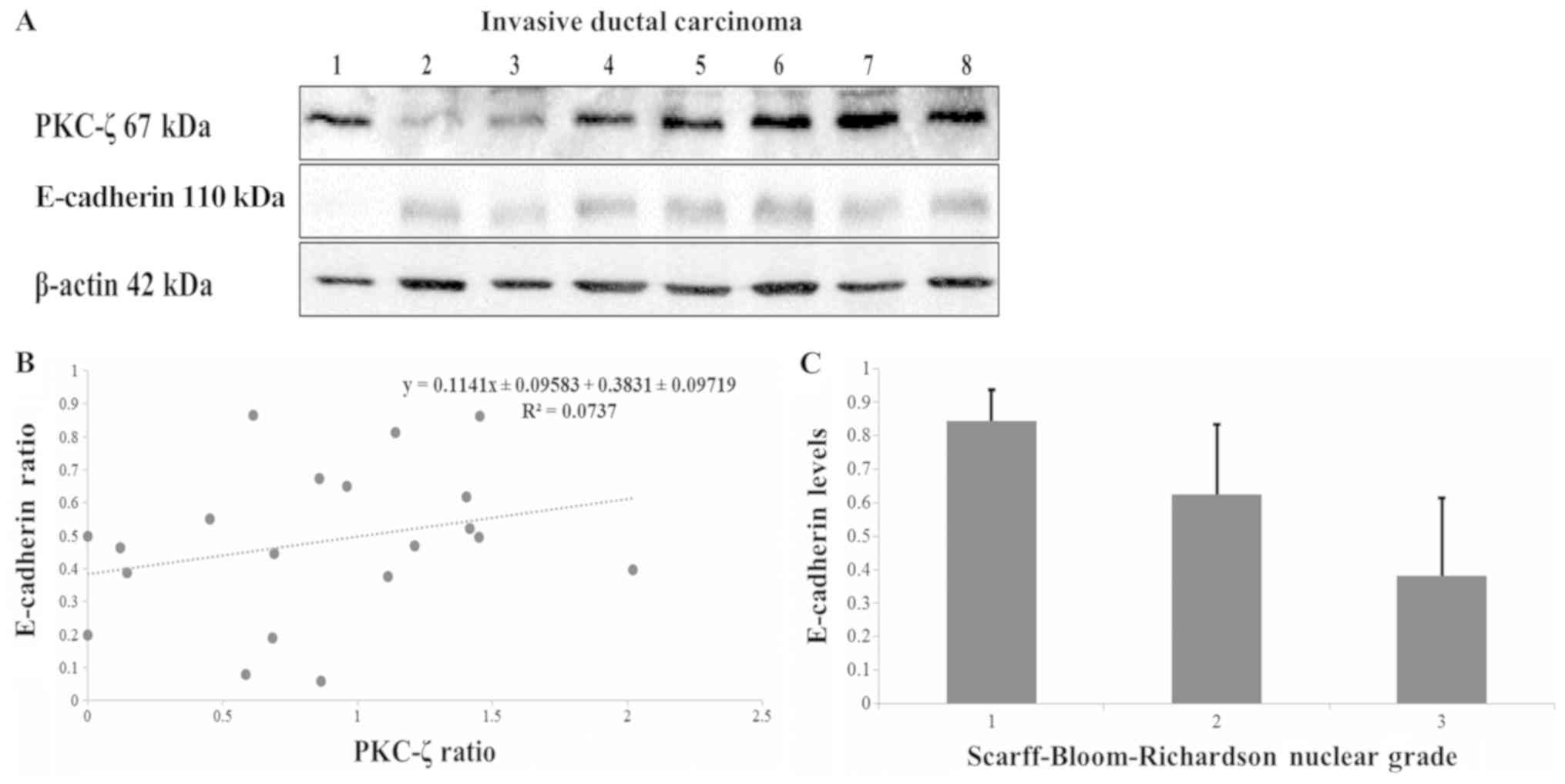
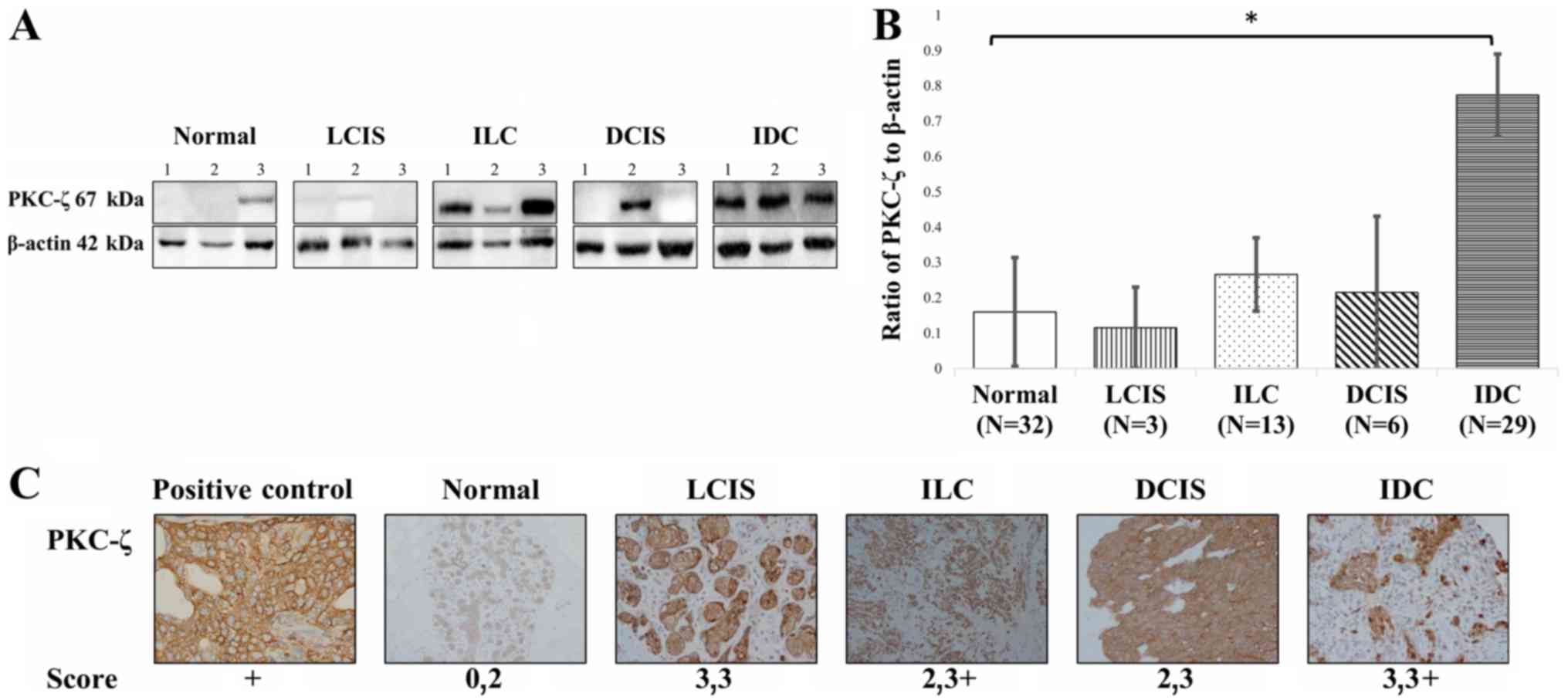 | Figure 1.Western blot and immunohistochemistry
analysis of PKC-ζ expression in normal and malignant tissues.
Malignant tissues were evaluated for the expression of PKC-ζ. (A)
We ran 20–30 µg of tissue lysate and probed with a PKC-ζ antibody.
(B) The ratios of PKC-ζ to β-actin in healthy tissue and the four
subtypes of malignant breast tissue were derived using the
densitometry. The white bar (healthy) represents the control and
the error bars represent the standard error. The standard
deviation, a Student's t-test (P<0.05) and a one-way ANOVA was
used to evaluate the western blot data as well with the post-hoc
Tukey's HSD test (P<0.01), Scheffé multiple comparison
(P<0.05), Bonferroni (P<0.01) and Holm (P<0.01).
Statistical tests showed no meaningful relationship between the
healthy and LCIS, or the ILC and DCIS. *, statistically relevant
relationship between normal and IDC breast tissue. (C) FFPE samples
were probed with PKC-ζ and stained for immunohistochemistry. The
scoring was based on the abundance or lack thereof PKC-ζ
expression: Score: 0, no expression; 1, weak expression; 2,
moderate expression; 3, abundant expression; 3 plus (+), strong
expression. Image original magnification, ×20. LCIS, lobular
carcinoma in situ; ILC, invasive lobular carcinoma; IDC,
invasive ductal carcinoma; DCIS, ductal carcinoma in situ;
PKC-ζ, protein kinase C-ζ; FFPE, formalin-fixed paraffin-embedded
tissues. |
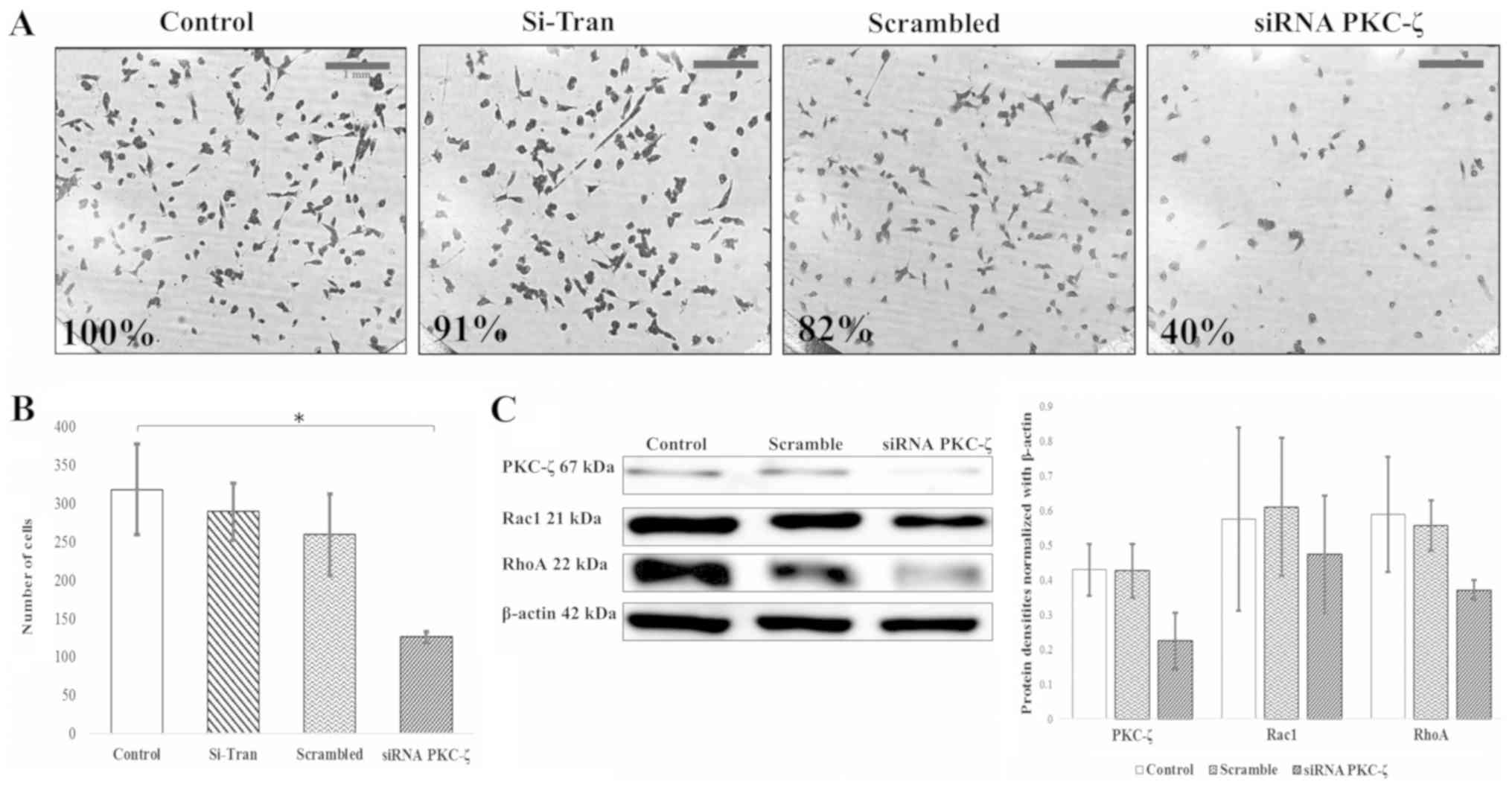 | Figure 3.PKC-ζ knockdown retards invasion of
MDA-MB-231 breast cancer cells. MDA-MB-231 breast cancer cells were
placed in the upper chamber of Transwell plate coated with 0.5X BME
and serum containing media was placed in the lower chamber as a
chemoattractant. Following transfection with siPRKCZ and scrambled
RNA, cells that invaded through the BME and migrated into the lower
chamber were stained with crystal violet and (A) observed under a
microscope at magnification, ×10; scale bar, 1 mm. Untreated cells
and cells treated with the transfection reagent (Si-Tran) were also
used to establish the effect on invasion. (B) The cells were
counted for each treatment image (N=3), averaged and
analyzed with the standard deviation, a Student's t-test
(*P<0.05) and a one-way ANOVA was also used to evaluate the data
with the post-hoc Tukey's HSD test (P<0.01), Scheffé multiple
comparison (P<0.01), Bonferroni (P<0.01) and Holm
(P<0.01). All statistical tests were used to determine the
statistical significance of the invasion data. (C) The cells were
further investigated by western blot to analyze the Rac1/RhoA
pathway. This data was quantified and averaged for the graph with
standard error of the mean represented. PKC-ζ, protein kinase C-ζ;
BME, basement membrane extract; Rac1, Ras-related C3 botulinum
toxin substrate 1; RhoA, Ras homolog gene family member A. |
 | Table III.Chi-square statistical analysis
results of normal and malignant breast subtypes (normal, LCIS, ILC,
DCIS and IDC; N=83) |
Table III.
Chi-square statistical analysis
results of normal and malignant breast subtypes (normal, LCIS, ILC,
DCIS and IDC; N=83)
| Variable | Normal | LCIS | ILC | DCIS | IDC | Row totals | Chi-square | P-value |
|---|
| PKC-ζ present | 3 | 1 | 5 | 1 | 23 | 33 |
a32.6715 | <0.00001 |
| PKC-ζ absent | 29 | 2 | 8 | 5 | 6 | 50 |
|
|
| Column totals | 32 | 3 | 13 | 6 | 29 | 83 (grand
total) |
|
|
| Table II.The status of PKC-ζ in malignant and
healthy breast tissue. |
Table II.
The status of PKC-ζ in malignant and
healthy breast tissue.
| Tissue type | Not present | Weakly present | Positively
present |
|---|
| Normal tissue | 29 | 1 | 2 |
| LCIS | 2 | 1 | 0 |
| ILC | 8 | 4 | 1 |
| DCIS | 5 | 0 | 1 |
| IDC | 6 | 5 | 18 |
| Table IV.Chi-square statistical analysis
results of estrogen receptor expression (N=25). |
Table IV.
Chi-square statistical analysis
results of estrogen receptor expression (N=25).
| Variable | ER+ | ER- | Row totals | Chi-square | P-value |
|---|
| PKC-ζ present | 10 | 6 | 16 | 0.0434 |
0.834969 |
| PKC-ζ absent | 6 | 3 | 9 |
|
|
| Column totals | 16 | 9 | 25 (grand
total) |
|
|
PKC-ζ protein levels are higher in
invading breast cancer subtypes
To compare the expression of PKC-ζ in non-invading
tissues and invading tissues, western blot and immunohistochemistry
were performed. Western blot analysis data showed that PKC-ζ
protein expression was higher in IDC when compared to the ILC, LCIS
and the DCIS (Fig. 1A and B).
According to immunohistochemistry findings, breast specimens were
scored for the expression of PKC-ζ by the pathologist (Fig. 1C). Normal tissue had no to moderate
expression (score, 0,2), LCIS had abundant expression (scores,
3,3), ILC had moderate to strong expression (scores, 2,3+), DCIS
had moderate to abundant expression (scores, 2.3), and IDC had
abundant to strong expression (scores, 3,3+). The most robust
expression was found to be in ILC and IDC represented by the 3+
score. However, ILC was found to have moderate to strong
expression, whereas IDC was found to have abundant to strong
expression. These findings suggest that PKC-ζ is found higher in
invading tissues when compared to non-invading tissues and more so
in IDC when compared to ILC. These data suggest that the stage
between in situ and invading malignancy can be correlated to
an increased PKC-ζ protein expression.
PKC-ζ and E-cadherin levels in tissue
specimens
Since the decreased expression of E-cadherin is
indicative of a more aggressive phenotype, the relationship between
PKC-ζ and E-cadherin protein levels was studied. Western blot
analysis was used in the context of probing for PKC-ζ and
E-cadherin in IDC breast tissues (Fig.
2A). We randomly selected 8 samples out of the 29 IDC tissue
specimen to illustrate the data. We could not establish a
significant relationship between PKC-ζ expression and E-cadherin
expression in IDC samples. Our linear regression test (Fig. 2B) showed no significance (P<0.05)
between the expressions of PKC-ζ and E-cadherin protein levels in
20 randomly selected IDC breast tissue samples. The N value
was too low to take into consideration for LCIS, ILC and DCIS.
Normal breast tissues were not taken into consideration since the
PKC-ζ expression was only found in 3 samples out of 32. However,
our investigation established the relationship between E-cadherin
protein expression and nuclear grade diagnosis (Fig. 2C). The Scarff-Bloom-Richardson scale
data was derived from the pathology reports (summarized in Tables I and V). The results show that E-cadherin protein
expression had an inverse relationship to nuclear grade diagnosis.
Even though the PRKCZ gene is not one of the top mutated genes in
breast cancer (carcinomas and carcinomas in situ), CDH1 is
on the top of the list (Table VI)
(3).
| Table I.The selection of breast specimens
with pathological characteristics. |
Table I.
The selection of breast specimens
with pathological characteristics.
| LN | Grade/stage | ER | PR | Histology | Race | PKC-ζ
expression | Number of patient
samples |
|---|
| NA | NA | NA | NA | Normal | Black | No | 9 |
| NA | NA | NA | NA | Normal | NA | No | 4 |
| NA | NA | NA | NA | Normal | White | No | 15 |
| NA | NA | NA | NA | Normal | White | Yes | 4 |
| NA | NA | NA | NA | LCIS | Black | No | 1 |
| + | II/III | − | − | LCIS | White | No | 1 |
| + | II/III | + | + | LCIS | White | Yes | 1 |
| NA | NA | NA | NA | ILC | Black | No | 1 |
| − | II/III | NA | NA | ILC | NA | No | 1 |
| + | III/III | + | + | ILC | NA | No | 1 |
| NA | NA | NA | NA | ILC | NA | No | 1 |
| − | I/III | + | + | ILC | White | No | 1 |
| − | NA | NA | NA | ILC | White | No | 1 |
| + | II/III | + | + | ILC | White | No | 1 |
| NA | NA | NA | NA | ILC | White | No | 1 |
| + | II/III | + | + | ILC | Black | Yes | 1 |
| + | III/III | + | + | ILC | NA | Yes | 1 |
| NA | NA | NA | NA | ILC | NA | Yes | 1 |
| − | NA | + | + | ILC | White | Yes | 1 |
| NA | I/III | + | − | ILC | White | Yes | 1 |
| − | III/III | − | + | IDC | Black | No | 1 |
| + | NA | − | + | IDC | Black | No | 1 |
| NA | NA | NA | NA | IDC | NA | No | 1 |
| − | I/III | + | + | IDC | White | No | 1 |
| NA | III/III | NA | NA | IDC | White | No | 1 |
| NA | NA | + | − | IDC | White | No | 1 |
| NA | NA | + | + | IDC | Black | Yes | 1 |
| − | II/III | NA | NA | IDC | NA | Yes | 1 |
| + | III/III | NA | NA | IDC | NA | Yes | 1 |
| + | III/III | NA | NA | IDC | NA | Yes | 1 |
| NA | NA | + | − | IDC | NA | Yes | 1 |
| NA | NA | NA | NA | IDC | NA | Yes | 2 |
| − | III/III | − | − | IDC | White | Yes | 2 |
| + | II/I | + | + | IDC | White | Yes | 1 |
| + | NA | − | − | IDC | White | Yes | 2 |
| + | NA | − | NA | IDC | White | Yes | 1 |
| + | NA | + | + | IDC | White | Yes | 1 |
| + | NA | NA | NA | IDC | White | Yes | 2 |
| NA | I/III | NA | NA | IDC | White | Yes | 1 |
| NA | NA | − | − | IDC | White | Yes | 1 |
| NA | NA | + | − | IDC | White | Yes | 2 |
| NA | NA | NA | NA | IDC | White | Yes | 3 |
| NA | NA | NA | NA | DCIS | Black | No | 1 |
| NA | NA | NA | NA | DCIS | NA | No | 4 |
| − | NA | NA | NA | DCIS | Black | Yes | 1 |
| Table V.Chi-square statistical analysis
results of Scarff-Bloom-Richardson grading (N=22). |
Table V.
Chi-square statistical analysis
results of Scarff-Bloom-Richardson grading (N=22).
| Variable | NG1 | NG2 | NG3 | Row totals | Chi-square | P-value |
|---|
| PKC-ζ present | 4 | 4 | 5 | 13 | 0.2 | 0.90485 |
| PKC-ζ absent | 3 | 2 | 4 | 9 |
|
|
| Column totals | 7 | 6 | 9 | 22 (grand
total) |
|
|
| Table VI.Top mutated genes in all breast
cancer types. |
Table VI.
Top mutated genes in all breast
cancer types.
| Gene | Protein
product | Protein
function | Chromosome
location | Percentage of
mutated samples tested (%) | Highest percentage
mutation (%) | Mutation type |
|---|
| PIK3CA |
Phosphatidylinosital-4,5-bisphosphate
3-kinase catalytic subunit α | Phosphorylates
certain signaling molecules | 3q26.3 | 27 | 98.97 | Substitution
missense |
| TP53 | p53 | Tumor suppressor,
regulates cell cycle | 17p13.1 | 24 | 50.30, 25.20 | Substitution
missense, other |
| MED12 | Mediator complex
subunit 12 | Initiation of
transcription | Xq13 | 20 | 71.04 | Substitution
missense |
| CDH1 | E-cadherin
(Cadherin 1) | Cell adhesion | 16q22.1 | 11 | 28.07, 18.42,
17.54 | Deletion
frameshift, substitution nonsense, substitution missense |
| GATA3 | GATA binding
protein 3 | Transcription
factor | 10p15 | 10 | 48.58, 16.60 | Insertion
frameshift, deletion frameshift |
PRKCZ gene silencing decreases the
invasion of breast cells
To investigate the effects of PKC-ζ inhibition on
the invasive behavior of breast cancer cells, a Transwell invasion
assay and immunostaining of F-actin were performed. When compared
to the control, PKC-ζ knockdown decreased the invasion of breast
cancer cells by 60% and was significant (Student's t-test
P<0.05, one-way ANOVA Tukey HSD P-value 0.0029646, P<0.01;
Scheffé P-value 0.0049622, P<0.01; Bonferroni P-value 0.0040067,
P<0.01 and Holm P-value 0.0040067, P<0.01) (Fig. 3A and B). In addition, the levels of
two important components of metastatic pathways, Rac1 and RhoA,
were also decreased dramatically (Fig.
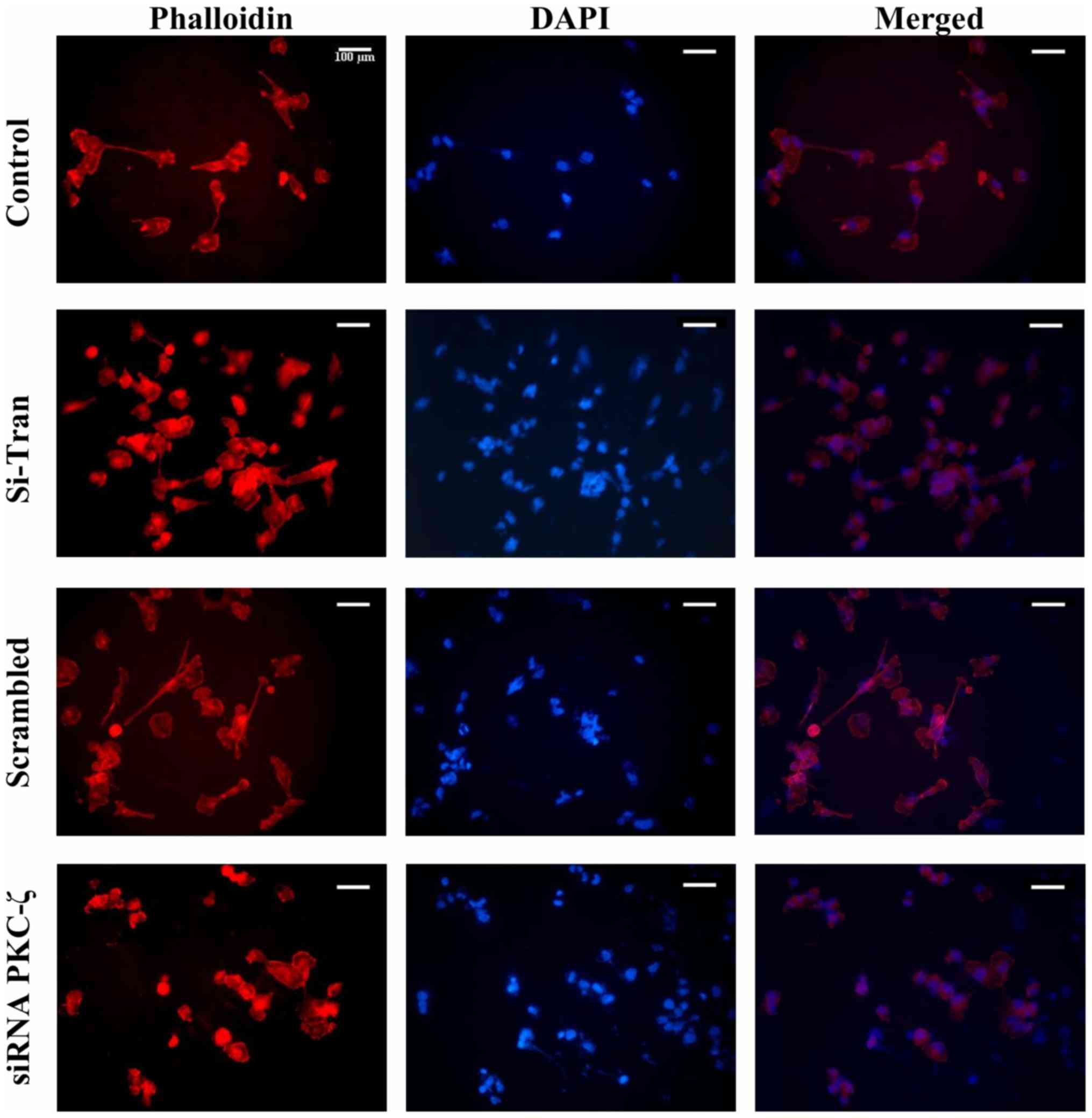
3C). Moreover, MDA-MB-231 breast cancer cells were fixed and
stained with phalloidin probe to visualize the impacts of PRKCZ
gene silencing on F-actin organization. The silencing of PRKCZ
caused the reorganization of F-actin around the cell cytoskeleton
(Fig. 4). Taken together, these data
advance the theory that PKC-ζ modulates the invasive behavior of
breast cancer cells by the regulation of invasion through the
Rac1/RhoA pathway and cytoskeleton filaments.
Discussion
Previously, Yin et al showed that the
expression of PKC-ζ was higher in invading breast tissues compared
to samples uncomplicated with tumors and the highest PKC-ζ protein
expression existed in stage III human breast ductal carcinomas
(17). Likewise, Paul et al
concluded that the depletion of PKC-ζ reduced the invasive
behaviors of MDA-MB-231 cells by upregulating epithelial markers
such as Zonula occludens-1 (ZO-1) and E-cadherin (6). They also found that PKC-ζ activation
(phosphorylated PKC-ζ levels) was found higher in invasive tissues
(i.e., IDC and metastatic tissues) when compared to non-invasive
tissues (DCIS). PKC-ζ expression did not significantly change with
the presence or absence of receptors (ER and HER2) (6). Our western blots and Chi-squared
analysis (Fig. 1A and B; Table IV) support these findings. They
performed a PKC-ζ knockdown mouse study and found that the
depletion of PKC-ζ leads to an approximately 50% reduction in
primary tumor growth compared to the control within five weeks
(6). Similarly, our western blot
analysis and immunohistochemistry data of IDC and ILC (Fig. 1A-C) suggest that PKC-ζ might also be a
novel component in pathways that affect cancer cell invasion and
metastasis.
PKC-ζ assists a transcription factor (NFκB-p65) that
downregulates targets such as E-cadherin and ZO-1 (6). In addition, decreased E-cadherin levels
cause the cells to no longer adhere to the extracellular matrix
causing the cells to migrate, invade, or metastasize (6). Moreover, Chua et al showed that
NF-κB induction elevated expression of Zinc finger E-box binding
homeobox 1 and 2 (ZEB-1 and ZEB-2) which ultimately repressed the
E-cadherin levels (7). In our
investigation, we could not establish a statistical relationship
between PKC-ζ and E-cadherin protein expression in IDC (Fig. 2B) as per the linear regression
analysis. However, previous studies illustrated an increase in
E-cadherin levels in PKC-ζ knockdown MDA-MB-231 cells (6).
In addition, our findings showed an increased
expression of PKC-ζ in IDC tissues when compared to other subtypes
(Fig. 1A-C) which may be because of
the difference in pathological features of ductal and lobular
tumors. Ductal carcinomas are lined with a two-layered stratified
cuboidal epithelium resting on the basement membrane. This cuboidal
epithelium contains tight junctions where E-cadherins are located
and play a central role in cell-to-cell adhesion (18). In contrast, lobular carcinomas vary in
terms of molecular and genetic aberrations. Lobular carcinomas are
epithelial-like, growing individually in sheets or in a single file
(4). In our investigation, E-cadherin
levels were also compared to the nuclear grade listed on the
pathology report (Fig. 2C; Table V), which supports the previous studies
that described a decline in E-cadherin as a part of the epithelial
to mesenchymal transition leads to metastasis (7,19).
During metastasis, directional cell movement
involves five distinct steps: leading edge membrane protrusion,
adhesion of the protrusion to the substrates, cell body
translocation, de-adhesion of the tail from the substrate and
trailing edge retraction (20). These
processes are mainly controlled by reorganization of the actin in
the cell cytoskeleton which in turn are regulated by Guanosine
Triphosphatases (GTPases) (21).
Among the GTPases, RhoA, Rac1, and CDC42 are most commonly studied
because of their crucial role in actin assembly and formation of
metastatic structures of cells, such as filopodia, lamellipodia and
stress fibers (22). Rac1 and CDC42
produce localized actin polymerization at the leading edge which
pushes the membrane forward in slender like structure known as
filopodia and sheet-like structure known as lamellipodia that
ultimately generate locomotive force in migrating cells (23). In contrast, RhoA promotes the assembly
of contractile actomyosin filaments and acts on the rear end of the
migrating cells, inducing tail detachment (24). Thus, Rac1 and CDC42 stimulate leading
edge protrusion, and RhoA stimulates trailing edge retraction in
metastatic cells. We found that the knockdown of PKC-ζ by siPRKCZ
reduced the invasion of MDA-MB-231 breast cancer cells by 60%
(P<0.05) when compared to control (Fig. 3A and B). In addition, there was a
decreased expression of both Rac1 and RhoA in siPRKCZ transfected
MDA-MB-231 breast cancer cells compared to control (Fig. 3C). Furthermore, our immunostaining
analysis of F-actin illustrated that actin filaments were nicely
organized around the cells with the inhibition of PKC-ζ (Fig. 4). Hence, PKC-ζ may play an essential
role in the invasion and migration of breast cancer cells by the
regulation of RhoA and Rac1 pathways.
To conclude, our findings suggest that PKC-ζ is
found to be most abundant in invading tissue subtypes and may be a
functional component of invasion pathways such as Rac1 and RhoA.
The use of PKC-ζ-specific inhibitors could be used to correlate the
decrease in expression or functionality of PKC-ζ with the decrease
in invasive behavior in breast cancer.
Acknowledgements
The authors would like to thank the Tissue Core,
Moffitt Cancer Center (Tampa, FL, USA) for their assistance. Tissue
samples were provided by the NCI Cooperative Human Tissue
Network.
Funding
This study was funded by the University of South
Florida (Tampa, FL, USA) Foundations 42-0142. The authors would
like to acknowledge the financial contributions from the William
and Ella Owens Medical Research Foundation, Alaska Run for Women,
Save the Ta-Tas Foundation, Mary Ewell Dickens Foundation, Yolanda
and Salvatore Gigante Charitable Foundation Trust, Daniel Tanner
Foundation, Frederick H. Leonhardt Foundation and the Charles and
Ann Johnson Foundation.
Availability of data and materials
The datasets we used and/or analyzed during the
current study are available from the corresponding authors upon
reasonable request. The datasets we generated and/or analyzed
during the current study are available from the COSMIC repository
at https://cancer.sanger.ac.uk/cosmic.
Authors' contributions
TS contributions include tissue selection and
fractionation, western blot analysis, densitometry analysis,
statistical analyses, pathway analysis and writing. SMAI
contributions include the invasion assay, phalloidin staining, and
writing. CA contributions include preliminary western blot
analysis. AA contributions to the paper include statistical
analysis of E-cadherin expression and nuclear grade. MAD
contributions to the paper include concept, design, writing,
editing, resources, supervision and funding acquisition. The
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The right to informed consent was waived by the
Ethics committee of the University of South Florida.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Author information
TS is currently a graduate student at the University
of South Florida, Department of Chemistry. She is working on the
second year of her PhD and continues to work on ovarian cancer and
breast cancer.
Glossary
Abbreviations
Abbreviations:
|
PKC-ζ
|
protein kinase C ζ
|
|
PRKCZ
|
gene name for protein kinase C ζ
|
|
CDH1
|
gene name for E-cadherin
|
|
LCIS
|
lobular carcinoma in situ
|
|
ILC
|
invasive lobular carcinoma
|
|
DCIS
|
ductal carcinoma in situ
|
|
IDC
|
invasive ductal carcinoma
|
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kuhn E: Quick Guide for Most Commonly Used
Breast Cancer Statements. Susan G. Komen. 10–11. 2015.
|
|
3
|
COSMIC: Catalogue of somatic mutations in
cancer. 2016.
|
|
4
|
Ruibal A, Núñez MI, del Rio M, Arias J,
Martínez MI, Rabadán J and Tejerina A: Clinical-biological
differences between invasive ductal carcinomas and breast lobular
carcinomas. Preliminary results. Rev Esp Med Nucl. 18:84–87.
1999.PubMed/NCBI
|
|
5
|
Hoesel B and Schmid JA: The complexity of
NF-κB signaling in inflammation and cancer. Mol Cancer. 12:862013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Paul A, Danley M, Saha B, Tawfik O and
Paul S: PKCζ promotes breast cancer invasion by regulating
expression of E-Cadherin and Zonula Occludens-1 (ZO-1) via
NFκB-p65. Sci Rep. 5:125202015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chua HL, Bhat-Nakshatri P, Clare SE,
Morimiya A, Badve S and Nakshatri H: NF-kappaB represses E-cadherin
expression and enhances epithelial to mesenchymal transition of
mammary epithelial cells: Potential involvement of ZEB-1 and ZEB-2.
Oncogene. 26:711–724. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Singhai R, Patil VW, Jaiswal SR, Patil SD,
Tayade MB and Patil AV: E-Cadherin as a diagnostic biomarker in
breast cancer. N Am J Med Sci. 3:227–233. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hirai T and Chida K: Protein kinase Czeta
(PKCzeta): Activation mechanisms and cellular functions. J Biochem.
133:1–7. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Islam SMA, Patel R and Acevedo-Duncan M:
Protein Kinase C-ζ stimulates colorectal cancer cell carcinogenesis
via PKC-ζ/Rac1/Pak1/β-Catenin signaling cascade. Biochim Biophys
Acta Mol Cell Res. 1865:650–664. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu J, Liu S, Fan Z, Zhang L, Tian Y and
Yang R: A novel and selective inhibitor of PKC ζ potently inhibits
human breast cancer metastasis in vitro and in mice. Tumour Biol.
83:8391–8401. 2016. View Article : Google Scholar
|
|
12
|
Lin YM, Su CC, Su WW, Hwang JM, Hsu HH,
Tsai CH, Wang YC, Tsai FJ, Huang CY, Liu JY and Chen LM: Expression
of protein kinase C isoforms in cancerous breast tissue and
adjacent normal breast tissue. Chin J Physiol. 55:55–61. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Schöndorf T, Kurbacher CM, Becker M, Warm
M, Kolhagen H and Göhring UJ: Heterogeneity of proteinkinase C
activity and PKC-zeta expression in clinical breast carcinomas.
Clin Exp Med. 1:1–8. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim SW, Roh J and Park CS:
Immunohistochemistry for pathologists: Protocols, pitfalls, and
Tips. J Pathol Transl Med. 50:411–418. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
GraphPad Software: GraphPad QuickCalcs.
Linear regression calculator. 2018.
|
|
16
|
Social science statistics: Chi-square test
calculator-up to 5×5 contingency table. 2018.
|
|
17
|
Yin WJ, Lu JS, Di GH, Lin YP, Zhou LH, Liu
GY, Wu J, Shen KW, Han QX, Shen ZZ and Shao ZM: Clinicopathological
features of the triple-negative tumors in Chinese breast cancer
patients. Breast Cancer Res Treat. 115:325–333. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hugo HJ, Gunasinghe NPAD, Hollier BG,
Tanaka T, Blick T, Toh A, Hill P, Gilles C, Waltham M and Thompson
EW: Epithelial requirement for in vitro proliferation and xenograft
growth and metastasis of MDA-MB-468 human breast cancer cells:
Oncogenic rather than tumor-suppressive role of E-cadherin. Breast
Cancer Res. 19:862017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Onder TT, Gupta PB, Mani SA, Yang J,
Lander ES and Weinberg RA: Loss of E-cadherin promotes metastasis
via multiple downstream transcriptional pathways. Cancer Res.
68:3645–3654. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lauffenburger DA: Cell motility. Making
connections count. Nature. 383:390–391. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Raftopoulou M and Hall A: Cell migration:
Rho GTPases lead the way. Dev Biol. 265:23–32. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lawson CD and Ridley AJ: Rho GTPase
signaling complexes in cell migration and invasion. J Cell Biol.
217:447–457. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Small JV, Stradal T, Vignal E and Rottner
K: The lamellipodium: Where motility begins. Trends Cell Biol.
12:112–120. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nobes CD and Hall A: Rho GTPases control
polarity, protrusion, and adhesion during cell movement. J Cell
Biol. 144:1235–1244. 1999. View Article : Google Scholar : PubMed/NCBI
|