Introduction
Sirtuins, which are class III histone deacetylases,
are oxidized nicotinamide adenine dinucleotide
(NAD+)-dependent deacetylases that are implicated in the
control of a wide range of biological processes, including
apoptosis, stress responses, DNA repair, cell cycling, metabolism
and senescence (1,2). In total, seven sirtuins (Sirt1-7), with
considerably different functions and catalytic activities, are
found in mammals (3). Sirt1 has
attracted considerable attention in the last decade as a result of
its role in tumorigenesis in the last decade, due to its strong
catalytic activity and close similarity to the yeast Sir2 protein
(4).
Few studies have investigated the role of Sirtuin7
(Sirt7) in diseases (5). Sirt7 is
located mainly in the nucleus, particularly the nucleolus, where it
binds to rRNA. Sirt7 activity is required to resume rDNA
transcription during mitosis (6).
Additionally, previous studies have highlighted the role of Sirt7
in various notable cellular processes, particularly protein
synthesis, chromatin remodeling, cellular survival and lipid
metabolism (2,7). Of particular interest are reports
concerning the high expression level of Sirt7 in certain types of
cancer, indicating that Sirt7 could be considered to be an oncogene
(5,8,9). However,
the role of Sirt7 in glioma remains unclear, prompting this
investigation into the biological characteristics of Sirt7 in human
glioma.
According to the World Health Organization (WHO)
clinical classification, based on the phenotype of glioma (10), >50% of glioma are glioblastoma
multiform (GBM, grade IV astrocytoma). However, despite
considerable therapeutic efforts, glioblastoma (GBM) remains the
most common form of human brain tumor, and is associated with high
morbidity, recurrence and mortality rates (11). The overall 5-year survival rate of GBM
is <5%, and is even less for elderly patients (12). Thus, to understanding the mechanisms
of GBM tumorigenesis, a wide range of scientific studies are
required; for example, to identify a the biological marker for
targeted therapy so as to decrease the recurrence rate and improve
patient quality of life (13). The
current study investigated the biological roles of Sirt7 in glioma
tumorigenesis.
The regulation of mitogen-activated protein kinase
(MAPK)/extracellular signal-regulated kinase (ERK)-dependent
pathways serves a role in a number of biological functions, which
are particularly relevant in cancer cells, including integrin
activation chemoresistance and migration (14). The Ras/Raf ERK1/2 cascade is essential
for the induction of matrix metalloproteinase-9 (MMP-9) expression
(15).
Signal transducer and activator of transcription 3
(STAT3) serves roles in GBM tumorigenesis and central nervous
system development (16). A previous
study revealed that STAT3 is activated by growth factors or
extensively existed cytokines. Upon tyrosine phosphorylation by
receptor-associated tyrosine kinases, STAT3 translocates to the
nucleus and regulate gene transcription (17).
The present study examined Sirt7 expression in 15
clinical samples of glioma tissues and the corresponding non-tumor
specimens by western blot analysis. Sirt7 was overexpressed in
human glioma tissues, and its high expression level is associated
with the degree of glioma malignancy. These data indicated that
Sirt7 may serve as a sensitive prognostic indicator for glioma
patients. In addition, Sirt7 expression was decreased in glioma
cell lines using small interfering RNA (siRNA) interference. The
results revealed that Sirt7 was involved in cellular proliferation,
colony formation and the invasion ability of glioma cells, and that
downregulation of Sirt7 decreased the expression of phosphorylated
(p)-ERK, cyclin-dependent kinase 2 (CDK2) and STAT3. These results
indicated that Sirt7 may be a potential target in the glioma
therapy.
Materials and methods
Patient samples
Samples of glioma and matched non-tumor tissues were
collected from 15 patients at Affiliated Chenggong Hospital
(Xiamen, China). A total of 40% patients were female and 60% were
male, the median age was 45 years, ranging from 14–74 years.
Patients with malignant peripheral nerve sheath tumors or other
unrelated neurologic tumors were excluded. Informed consent was
obtained from all patients or family members, and the study
protocol was approved by the Ethical Committee of Affiliated
Chenggong Hospital. All tissues were snap-frozen and stored in
liquid nitrogen prior to use. A further 20 paraffin-embedded
samples obtained from Pathology Department of Affiliated Chenggong
Hospital. Only samples that were diagnosed with WHO stage II and IV
disease were chosen.
Analysis of Sirt7 expression using an
online microarray database
Human glioma expression microarray data were
downloaded from The Cancer Genome Atlas (TCGA) (http://cancergenome.nih.gov/), and were used to
analyze difference in the expression of SIRT7 between normal brain
and glioma tissues.
Cell culture and transfection
The U251 cell line was obtained from the China
Center for Typical Culture Collection. Professor Xiuwu Bian (The
Third Military Medical University, Chongqing, China) kindly
provided the glioma CHG5 and SHG44 cell lines. U87 cell lines were
purchased from the American Type Culture Collection (Manassas, VA,
USA). This cell line has been reported to be
misidentified/contaminated. This is not the original glioblastoma
cell line established in 1968 at the University of Uppsala; as
described in PubMed=27582061 (18).
However, this cell line exhibits Sirt7 overexpression and meets our
research requirements, it does not interfere with our experimental
conclusions. Cells were cultured in DMEM (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 10% FBS
(Gemini Bio Products, West Sacramento, CA, USA) at 37°C in a
humidified atmosphere containing 5% CO2.
Owing to transfection efficiency considerations and
the background high expression of Sirt7 in U87 and U251 cell lines,
these two cell lines were chosen in the knockdown experiment. Two
siRNAs targeting at Sirt7 (target sequence:
5′-GTGGACACTGCTTCAGAAA-3′ and 5′-CGAAGCTTTACATCGTGAA-3′) were
synthesized by Guangzhou RiboBio Co., Ltd., (Guangzhou, China).
Cells were seeded into a 6-well plate at 50–60% confluence, and
were then transfected with oligonucleotides at a final
concentration of 20 nM with the DharmaFECT reagent (Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions.
After 6 h, the transfection medium was replaced with 2 ml DMEM. The
cells were collected for the following assays after 48 h.
Western blot analysis
Cells were harvested and lysed in
radioimmunoprecipitation assay lysis buffer (50 mM Tris (pH 7.4),
150 mM NaCl, 1% NP-40 and 0.5% sodium deoxycholate) The BCA Protein
Assay kit was used to determine the protein concentrations
according to the manufacturer's instruction (Pierce; Thermo Fisher
Scientific, Inc.). Equal amounts of total proteins (25 µg) were
subjected to 10% SDS-PAGE. They were then transferred to
polyvinylidene difluoride membrane, incubated with 5% BSA (Sangon
Biotech, Co, Ltd., Shanghai, China) blocking buffer for 1 h at room
temperature and then incubated overnight with primary antibody at
4°C. The primary antibodies used were as follows: Sirt7 (cat. no.
5360), STAT3 (cat. no. 4904), p-STAT3 (cat. no. 9145), CDK2 (cat.
no. 2546), MMP-9 (cat. no. 3852) (all from Cell Signaling
Technology, Inc., Danvers, MA, USA, and used at a dilution of
1:1,000), and β-actin (cat. no. SC-130300, Santa Cruz
Biotechnology, Inc., Dallas, TX, USA). The membranes were then
incubated with the horseradish peroxidase-conjugated secondary
antibody (Anti-mouse IgG, cat. no. 7076; Anti-rabbit IgG, cat. no.
7076; from Cell Signaling Technology, Inc., Danvers, MA, USA, and
used at a dilution of 1:10,000). Immunoreactive bands (cat. no.
WBKLS0500; Millipore Immobilon Western, ECL; EMD Millipore,
Billerica, MA, USA) were quantified using ImageJ software (v.1.48;
National Institutes of Health, Bethesda, MD, USA).
Immunohistochemistry (IHC)
The archived paraffin- embedded tissues were cut to
a thickness of 4 µm. Immunostaining was performed using the
two-step Elivision Plus kit system (Fuzhou Maixin Biotech Co.,
Ltd., Fuzhou, China). The sections were dewaxed in xylene,
rehydrated with a series of ethanol solutions (100, 95, 80 and
70%), and then boiled in citrate buffer (pH 6.0) for 2 min in an
autoclave. Next, 0.3% H2O2 was used to block
endogenous peroxidase activity at room temperature for 15 min, and
the sections were incubated with normal goat serum (dilution ratio
1:20) for 20 min at room temperature to reduce non-specific
binding. Tissue sections were incubated with the Sirt7 antibody
(cat. no. 5360s; 1:150 dilution; Santa Cruz Biotechnology, Inc.)
for 2 h at room temperature. The secondary antibody was applied
using the Envision Detection kit (Dako; Agilent Technologies, Inc.,
Santa Clara, CA, USA). Slides were stained for 2 min with
diaminobenzidine tetrahydrochloride (DAB) and then counterstained 2
min with hematoxylin at room temperature. The stained tumor cells
were assessed with a Nikon microscope in 10 independent fields at
magnification, ×400.
Cell viability assay
An MTS assay was used to detect cell proliferation.
Cells were plated in 96-well plates at 1×104 cells/well and
cultured for 4 h before each of the treatment durations (24, 36 and
48 h). A total of 20 µl MTS was added to each well and then
incubated for 4 h at 37°C. The absorbance of each sample was
measured at 490 nm using a microplate reader.
Colony formation assay
Cells were seeded at a density of 1,000 per well
into a 6-well culture dish and incubated at 37°C. Culture medium
was replaced with fresh medium every two days. After 3 weeks, cells
were washed with PBS and stained with Gimesa stain for 20 min at
room temperature (EMD Millipore). Only colonies with >50 cells
were counted. A total of 12 random fields were counted under an
upright light microscope at magnification, ×200. All experiments
were performed in triplicate.
Cell invasion assay
An in vitro cell invasion assay was performed
using a 24 well-plate transwell insert (EMD Millipore) coated with
Matrigel (EMD Millipore). A total of 0.5 ml of prepared serum-free
suspension of transfected cells (24 h after transfection), with a
total cell count of 1×105, were added into the upper
chamber (8-µm pore size), 0.5 ml cell culture medium containing 10%
FBS was added to the lower chamber of the insert. After incubation
for 24 h, non-invading cells in the interior of the insert were
removed using a cotton swab. The invasive cells on the lower
surface of the insert were treated with Gimesa stain (1:9) for 20
min at room temperature and average percentage of positive tumor
cells were assayed in 10 independent fields at magnification, ×200
under an upright light microscope for research and clinical use.
All experiments were performed in triplicate.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
Total RNA was extracted from tissues and cells using
Tripure Isolation reagent (Roche Applied Science, Penzberg,
Germany), according to the manufacturer's instructions. The
Transcription First Strand cDNA Synthesis kit (Roche Applied
Science) was used to synthesize cDNA from 1 µg total RNA. The
primers sequences used are as follows: SIRT7, forward,
CAGGGAGTACGTGCGGGTGT, and reverse, TCGGTCGCCGCTTCCCAGTT; GAPDH,
forward, AAGTGAAGCAGGAGGGTGGAA, and reverse, CAGCCTCACCCCATTTGATG.
The thermocycling conditions were as follows: Initial denaturation
at 94°C for 5 min, followed by 30 cycles at 94°C for 45 sec, 59°C
for 45 sec and 72°C for 45 sec, followed by 72°C for 45 sec and
finally one cycle at 72°C for 10 min. Experiments were performed in
triplicate with SYBR Green I Master mix (Roche Applied Science);
GAPDH was used as the internal control. The results were normalized
using the 2−ΔΔCq method (19).
Statistical analysis
All quantified data represented an average of at
least three samples. Graph Pad Prism v.5.0 (GraphPad Software,
Inc., La Jolla, CA, USA) and SPSS v.16 (SPSS, Inc., Chicago, IL,
USA) were used for statistical analysis. The data are expressed as
the mean ± standard deviation. Comparisons between different groups
were performed with Student's t-test and one way analysis of
variance followed by a Student-Newman-Keuls post hoc test.
P<0.05 was considered to indicate a statistically significant
difference. TCGA statistical analysis was performed using the
Mann-Whitney U-test test.
Results
Expression of Sirt7 in human glioma
tissues
To examine the physiological relevance of Sirt7 in
human glioma development, the expression level of Sirt7 was
analyzed in clinical patient samples using a Sirt7-specific
antibody. The samples were obtained from 15 patients who were
classified into different grades according to their pathological
diagnoses. The characteristics of the glioma patients are
summarized in Table I. As shown in
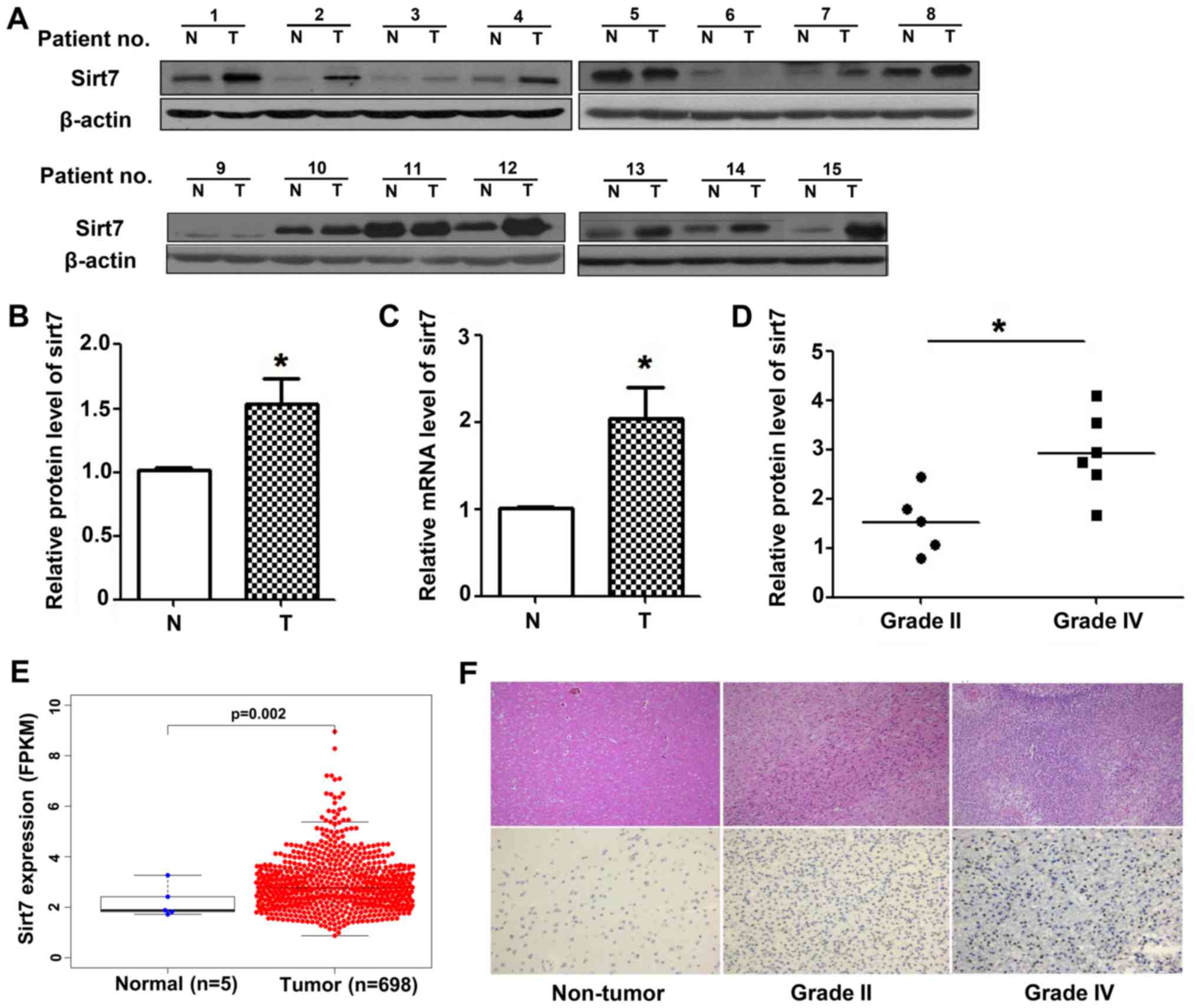
Fig. 1A, Sirt7 was positively
expressed in tumors compared with paired adjacent non-tumor
tissues. To confirm this finding, RT-qPCR and western blot analysis
was performed, which revealed that Sirt7 mRNA was significantly
elevated in glioma compared to the normal tissues, as was that of
Sirt7 protein (Fig. 1B and C).
Besides, to determine whether the levels of Sirt7 expression were
associated with the grade of pathological classification, the
expression level of Sirt7 was assessed in grade II and grade IV
tumors. The results of this analysis revealed that Sirt7 expression
was higher in grade IV than grade II gliomas (Fig. 1D).
 | Table I.Characteristics of Glioma
patients. |
Table I.
Characteristics of Glioma
patients.
| Patient | Age, years | Sex | WHO
classification | Pathological
diagnosis |
|---|
| 1 | 75 | Male | III | Anaplastic
astrocytoma |
| 2 | 51 | Female | IV | Glioblastoma |
| 3 | 43 | Male | II | Astroglioma |
| 4 | 38 | Male | IV | Glioblastoma |
| 5 | 14 | Female | IV | Glioblastoma |
| 6 | 41 | Male | II |
Oligodendrocyte |
| 7 | 45 | Female | II | Astroglioma |
| 8 | 39 | Male | I | Pilocytic
astrocytoma |
| 9 | 55 | Male | I | Pilocytic
astrocytoma |
| 10 | 52 | Female | II |
Oligodendrocyte |
| 11 | 52 | Male | III | Anaplastic
astrocytoma |
| 12 | 44 | Male | IV | Glioblastoma |
| 13 | 58 | Female | IV | Glioblastoma |
| 14 | 53 | Male | II | Astroglioma |
| 15 | 45 | Female | IV | Glioblastoma |
Owing to the limited number of fresh samples, TCGA
database was used to analyze the difference in expression between
the normal and glioma groups. Data from 698 GBM patients and 5
control group patients were extracted from TGCA database.
Expression of Sirt7 was significantly higher in the GBM patients
than in the control individuals (Fig.
1E). Additionally, the pathology database was accessed and
paraffin specimens (20 specimens, 9 for grade II and 11c for grade
IV) were obtained. IHC assays revealed the presence of strong Sirt7
staining in the nuclei of high-grade gliomas (Fig. 1F). These results indicated that Sirt7
may have a role in the development of glioma.
Expression levels of Sirt7 in
different immortalized glioma cell lines
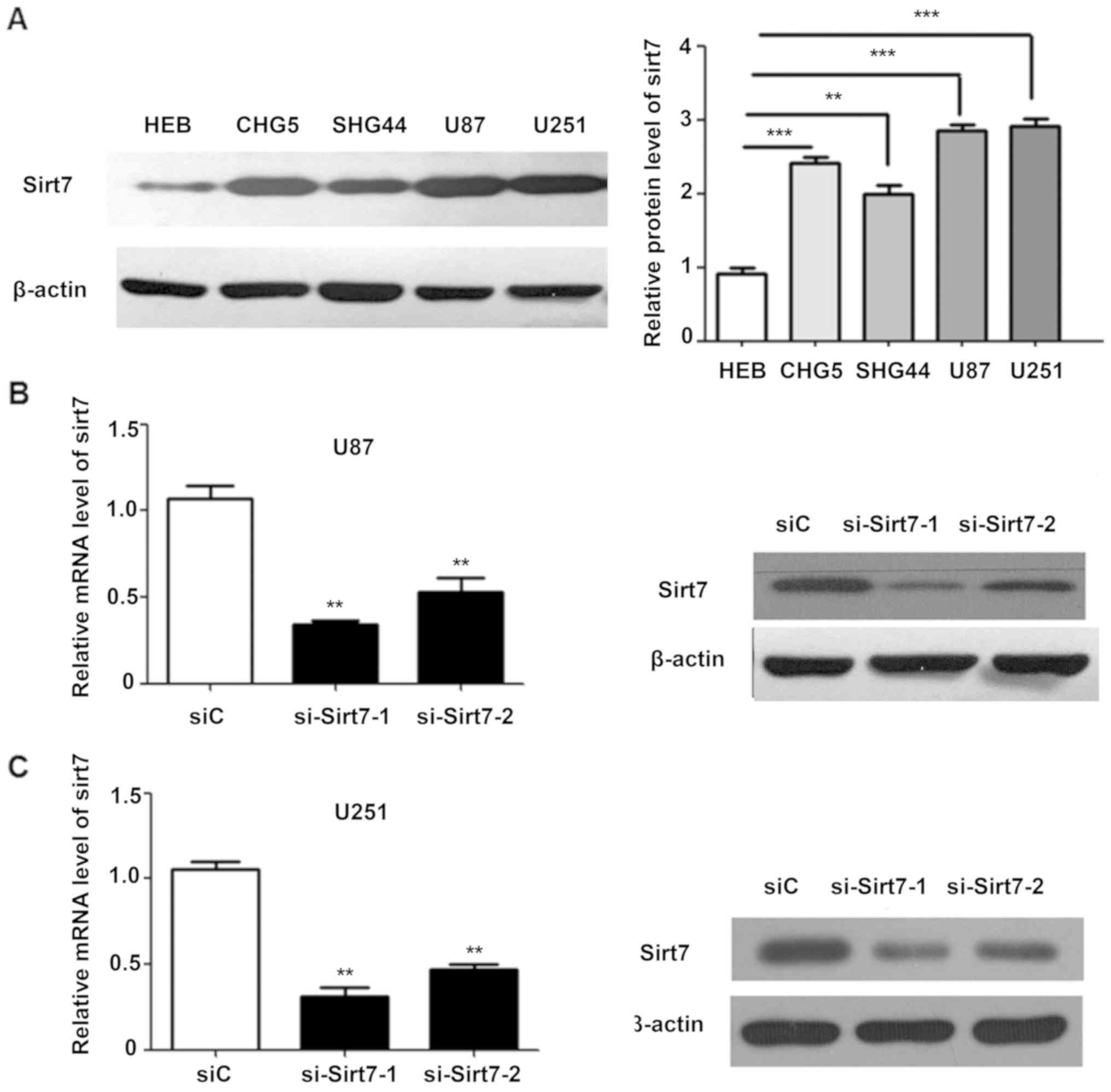
To understand the potential functions of Sirt7 in
glioma cells, the expression level of Sirt7 was analyzed in the
human glioma CHG5, SHG44, U87 and U251 and one normal glial HEB
cell lines (20). The CHG5 and SHG44
cell lines were derived from grade II tissues, whereas U87 and U251
cell lines were obtained from grade IV tissues. Sirt7 expression
was higher in glioma cells than in normal glial cells (Fig. 2A). Compared with the low-grade CHG5
and SHG44 cells, U251 and U87 cells expressed an even higher level
of Sirt7, in accordance with the results from the aforementioned
experiments.
As the glioma cell lines in the present study
exhibited high Sirt7 expression levels, Sirt7 was not
overexpressed. However, two different siRNAs targeting Sirt7 were
transfected into cells, and the efficiency of Sirt7 knock down was
determined by western blot analysis and RT-qPCR. Compared with the
control group, the protein and mRNA expression levels of Sirt7 in
the si-Sirt7 group were significantly decreased (Fig. 2B and C).
Downregulation of Sirt7 expression
suppresses cell proliferation in vitro
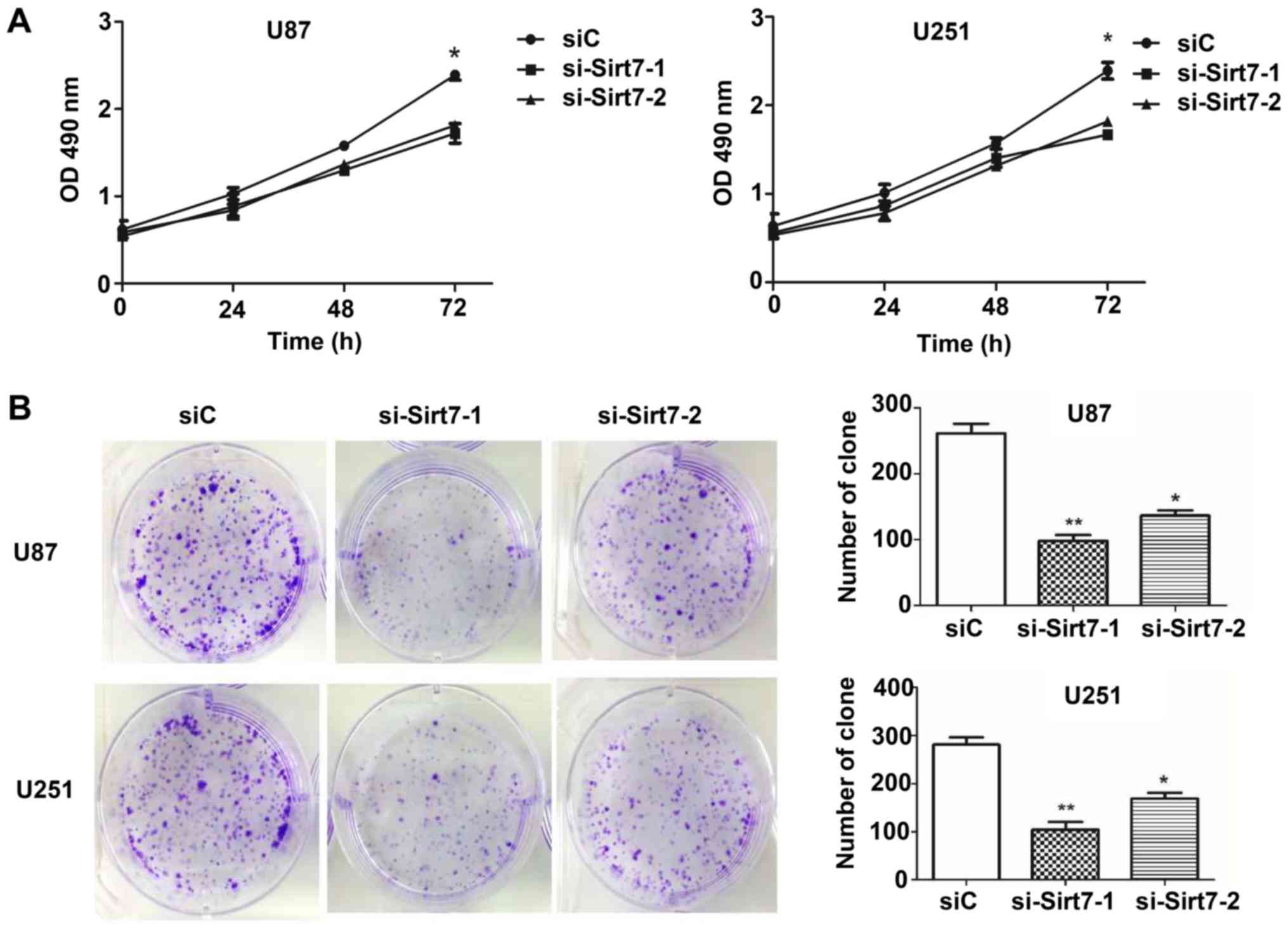
The aforementioned observations prompted the
investigation of the biological function of Sirt7 in the
tumorigenesis of glioma cells, which was achieved by performing MTS
assays to evaluate the effect of Sirt7 on cellular proliferation.
At 24 h after siRNA transfection up to 72 h, the proliferation of
cells was assessed (Fig. 3A). The
results demonstrated that knockdown of Sirt7 reduced the
proliferation of cells in the si-Sirt7 groups when compared to the
group transfected with the nonsense control siRNA. Whether Sirt7
affected the tumorigenesis of glioma cells was next assessed by
detecting the colony formation ability of glioma cells following
transfection. The results showed that Sirt7 knockdown significantly
reduced the number of colonies formed (Fig. 3B).
Sirt7 knockdown weakens cell
invasion
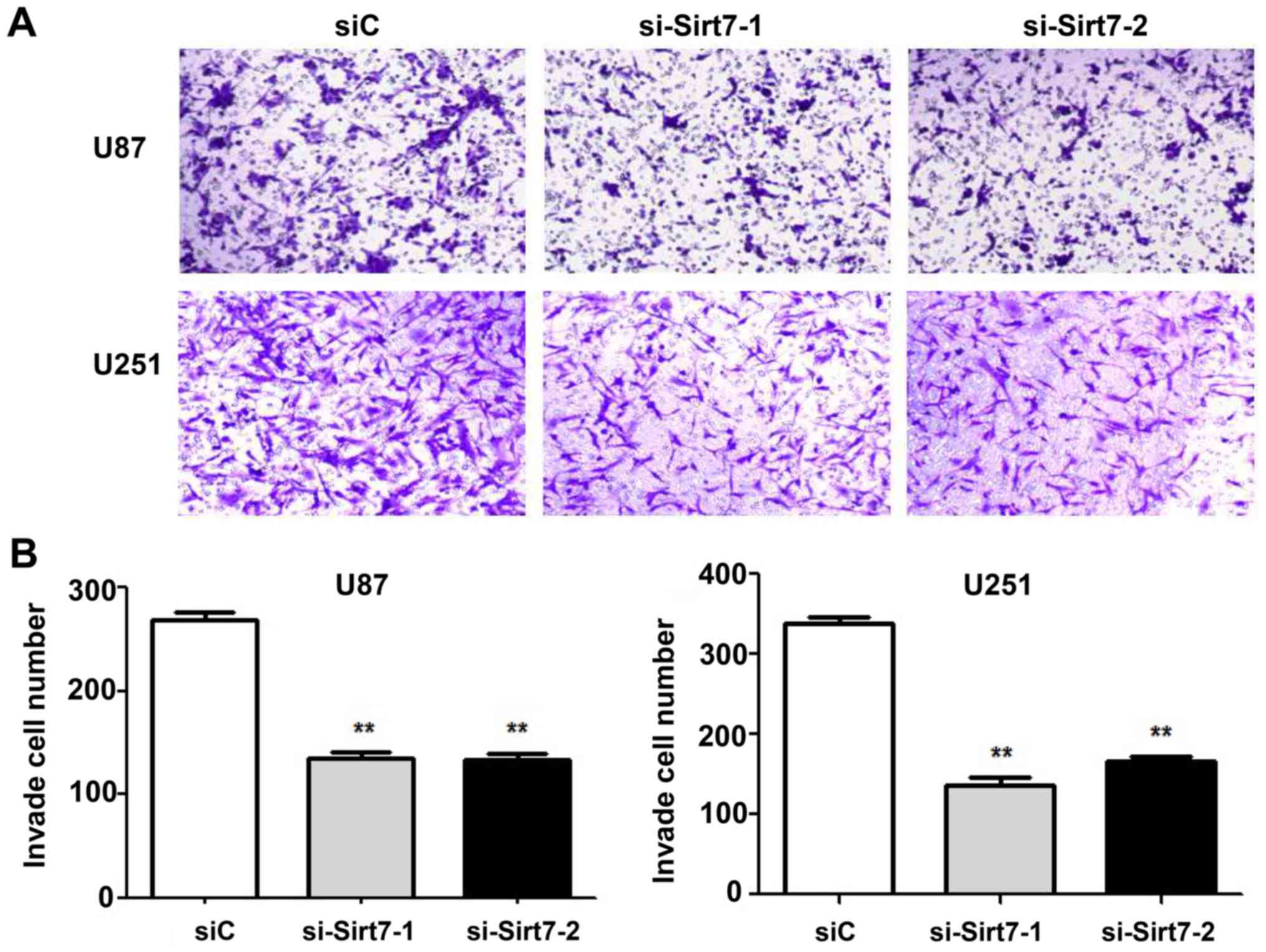
Invasive ability is one of the features that best
reflects the malignancy of glioma cells. Thus, the effects of Sirt7
on cell invasion was assessed by Matrigel invasion assays. Sirt7
was knocked down in the U87 and U251 cell lines by transfection
with si-Sirt7. The si-Sirt7 group exhibited reduced invasion into
the lower chamber compared to the control group (Fig. 4A and B; P<0.01). Depletion of Sirt7
significantly reduced cell invasion, indicating that Sirt7 may
participate in the progression of glioma.
Sirt7 knockdown reduces the
phosphorylation level of ERK and STAT3, and inhibits the expression
of MMP9 and CDK2
To investigate the molecular mechanism by which
downregulation of Sirt7 inhibited glioma cell proliferation and
invasion, whether knockdown of Sirt7 influenced ERK signaling
activation was examined. The level of p-ERK decreased when Sirt7
expression was suppressed (Fig. 5A and
B). Compared with the control group, the si-Sirt7 group
exhibited a decrease in the levels of p-STAT3 and MMP9. To
determine the function of Sirt7 in cell cycle progression, Sirt7
downregulation was performed, which decreased the expression of
CDK2 (Fig. 5A and B), indicating that
Sirt7 may participate in cell cycle regulation, which could further
influence the glioma tumorigenesis.
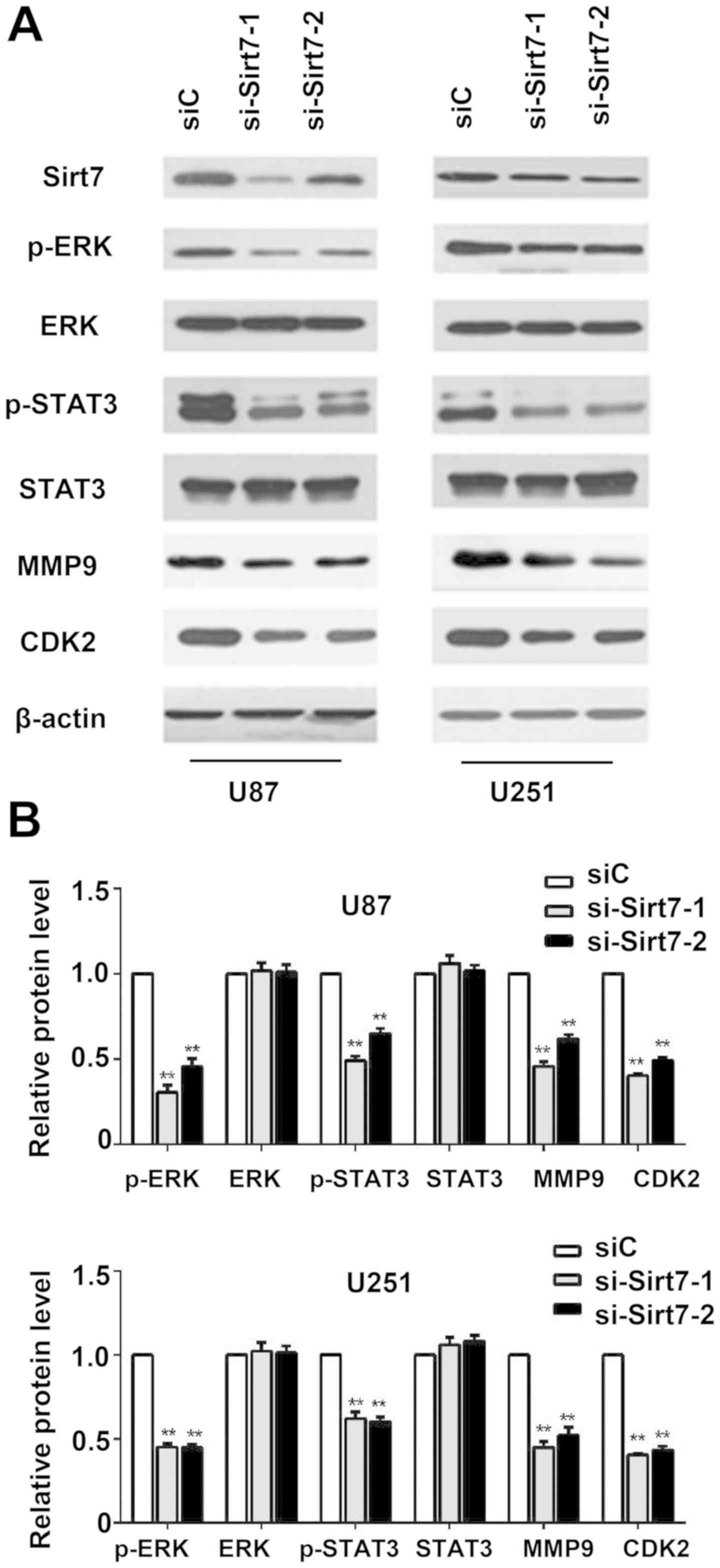 | Figure 5.Sirt7 knockdown decreased the
phosphorylation of ERK, STAT3, and inhibited expression of MMP9 and
CDK2 in U87 and U251 glioma cells. (A) Western blot was used to
detect the key signal molecules, with β-actin used as an internal
control. (B) Quantification of the results in (A) Data are
expressed as the mean ± standard deviation of triplicate
experiments. **P<0.01 indicates significantly different from the
siC group. ERK, extracellular signal-regulated kinase; STAT3,
signal transducer and activator of transcription 3; MMP9, matrix
metallopeptidase 9; CDK2, cyclin-dependent kinase 2; siRNA, small
interfering RNA; si-Sirt7, siRNA targeting Sirt7; siC, control
siRNA. |
Discussion
The mammalian sirtuin family has attracted
considerable attention as regulators of numerous cellular
processes, including metabolism, aging, cell cycle progression and
cell survival under stress conditions (4,21). Sirt7,
the newest member of the sirtuin family, is a nuclear protein that
interacts with RNA polymerase I. Depletion of Sirt7 halts cellular
proliferation and triggers apoptosis (22). The dysregulated expression of Sirt7
has been detected in several malignancies, including colorectal
cancer, gastric cancer, breast cancer and hepatocellular carcinoma
(8,23). Ford et al (22) revealed that Sirt7 is associated with
transcriptionally active rRNA genes, by interacting with RNA
polymerase I (Pol I) and histones. Knockdown of Sirt7 resulted in
decreased association of Pol I with rDNA and a reduction of Pol I
transcription. Sirt7 is an NAD1-dependent histone 3 acetyl Lys18
deacetylase that stabilizes the transformed state of cancer cells
(24).
However, to the best of our knowledge, no intensive
study of the biological functions of Sirt7 in human glioma has been
conducted. The present study observed that significantly higher
Sirt7 expression levels were observed in glioma tissues than in
adjacent non-tumor tissues. Furthermore, the results of the present
study demonstrated that Sirt7 upregulation was significantly
associated with higher WHO grades, indicating that Sirt7 may
participate in the progression of glioma. More notably, in
functional experiments, Sirt7 knockdown markedly inhibited cell
colony formation and invasion in vitro. Additionally, Sirt7
inactivation suppressed MMP9 and CDK2 expression, indicating that
Sirt7 may lead to mitotic stimulation and uncontrolled cell growth
during glioma progression. This finding was consistent with the
results of a study of human hepatocellular carcinoma tumorigenesis,
which indicated that Sirt7-knockdown was associated with
G1 phase cell cycle arrest (8). However, further research into the
mechanism of suppression is required.
ERKs are required for cell growth, and are reported
to exert a diversity of biological functions in differentiation,
apoptosis, inflammation, oncogenesis and angiogenesis in numerous
tumor types (25–27). ERK signaling is activated in the
majority of gliomas (28). In glioma
cell lines and stem cell-enriched primary cultures of glioblastoma,
constitutive activation of ERK kinases has been observed (29). ERK phosphorylation is a major upstream
activator of MMP9 (30). STAT3, which
was originally identified to be a mediator of inflammatory response
to interleukin-6 (IL-6) (31), is
required for brain tumor formation, as well as for maintenance of
multipotency in GBM stem cells (32,33). We
hypothesized that Sirt7 is important to this activation in two
pathways; knockdown of Sirt7 expression decreased the
phosphorylation level of ERK and STAT3, which indicated that STAT3
and ERK may be downstream effectors of Sirt7. Nevertheless, the
identity of the specific signaling pathway in which Sirt7 is
involved remains unclear. Therefore, additional research is
required to confirm the findings of the present study.
The results of the present study demonstrated that
the targeted knockdown of Sirt7 inhibited glioma cell proliferation
and invasion in vitro. Sirt7 selectively modulated the cell
cycle and invasion-associated protein synthesis, partly through the
ERK/MMP-9 signaling axis. In addition, interference of Sirt7
expression disrupted STAT3 signaling via inhibition of STAT3
phosphorylation, which is required for proliferation and the
maintenance of multipotency in glioblastoma stem cells (34). Unfortunately, phosphatase inhibitors
were not added to the lysis buffer during preparation of the
patient samples; as a result, the phosphorylation status of STAT3
and ERK1/2 in tumor and adjacent normal tissues could not be
detected. Finally, a reduction in CDK2 levels was observed in U87
and U251 cell lines when Sirt7 expression was suppressed,
indicating that the suppression of Sirt7 may cause cell cycle
arrest and inhibit cell growth. Thus, the findings of the present
study demonstrated that Sirt7 could represent a potential
therapeutic target in glioma. These data identified the potential
clinical value of quantifying Sirt7 expression, which may aid
assessment of the clinical prognosis. Future studies should
investigate the detailed association between STAT3, ERK1/2, and
Sirt7. A number of research topics remain to be studied, which
include whether activating p-ERK and p-STAT3 directly or indirectly
can antagonize the induction of Sirt7-inhibited cell proliferation
and invasion.
Acknowledgements
The authors would like to thank Dr. Kathryn Williams
(University of Florida, Gainesville, FL, USA) for writing
assistance, and Professor Xiuwu Bian (Third Military Medical
University, Chongqing, China) for the gift of the glioma cell
lines.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant no. 81402064),
Science and Technology Project of Xiamen (grant nos. 3502Z20164043,
3502Z20154039 and 3502Z20124019).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
PM, KL, QL, WS and TJ conceived the study,
participated in its design and drafted the manuscript. DL
participated in the cell experiments. WY and ZL performed sample
collection and IHC staining. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The research was approved by the Ethics Committee of
the Chinese People's Liberation Army 174 Hospital (Affiliated
Chenggong Hospital, Xiamen University). Informed consent was
provided by all patients or their family members.
Patient consent for publication
Informed permission for publication was acquired
from all patients.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
p-ERK
|
phosphorylated extracellular signal-
regulated kinase
|
|
CDK2
|
cyclin-dependent kinase 2
|
|
STAT3
|
signal transducer and activator of
transcription 3
|
|
siRNA
|
small interfering RNA
|
References
|
1
|
Houtkooper RH, Pirinen E and Auwerx J:
Sirtuins as regulators of metabolism and healthspan. Nat Rev Mol
Cell Biol. 13:225–238. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kiran S, Anwar T, Kiran M and Ramakrishna
G: Sirtuin 7 in cell proliferation, stress and disease. Rise of the
Seventh Sirtuin! Cell Signal. 27:673–682. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
North BJ and Verdin E: Sirtuins:
Sir2-related NAD-dependent protein deacetylases. Genome Biol.
5:2242004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Haigis MC and Sinclair DA: Mammalian
sirtuins: Biological insights and disease relevance. Ann Rev
Pathol. 5:253–295. 2010. View Article : Google Scholar
|
|
5
|
Zhang S, Chen P, Huang Z, Hu X, Chen M, Hu
S, Hu Y and Cai T: Sirt7 promotes gastric cancer growth and
inhibits apoptosis by epigenetically inhibiting miR-34a. Sci Rep.
5:2015.
|
|
6
|
Grob A, Roussel P, Wright JE, McStay B,
Hernandez-Verdun D and Sirri V: Involvement of SIRT7 in resumption
of rDNA transcription at the exit from mitosis. J Cell Sci.
122:489–498. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yoshizawa T, Karim MF, Sato Y, Senokuchi
T, Miyata K, Fukuda T, Go C, Tasaki M, Uchimura K, Kadomatsu T, et
al: SIRT7 controls hepatic lipid metabolism by regulating the
ubiquitin-proteasome pathway. Cell Metab. 19:712–721. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kim JK, Noh JH, Jung KH, Eun JW, Bae HJ,
Kim MG, Chang YG, Shen Q, Park WS, Lee JY, et al: Sirtuin7
oncogenic potential in human hepatocellular carcinoma and its
regulation by the tumor suppressors MiR-125a-5p and MiR-125b.
Hepatology. 57:1055–1067. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Geng Q, Peng H, Chen F, Luo R and Li R:
High expression of Sirt7 served as a predictor of adverse outcome
in breast cancer. Int J Clin Exp Pathol. 8:1938–1945.
2015.PubMed/NCBI
|
|
10
|
Louis DN, Ohgaki H, Wiestler OD, Cavenee
WK, Burger PC, Jouvet A, Scheithauer BW and Kleihues P: The 2007
WHO classification of tumours of the central nervous system. Acta
Neuropathol. 114:97–109. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Stupp R, Mason WP, van den Bent MJ, Weller
M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn
U, et al: Radiotherapy plus concomitant and adjuvant temozolomide
for glioblastoma. New Eng J Med. 352:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen J, McKay RM and Parada LF: Malignant
glioma: Lessons from genomics, mouse models, and stem cells. Cell.
149:36–47. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Aldape K, Zadeh G, Mansouri S,
Reifenberger G and von Deimling A: Glioblastoma: Pathology,
molecular mechanisms and markers. Acta Neuropathol. 129:829–848.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Eckert A, Böck BC, Tagscherer KE, Haas TL,
Grund K, Sykora J, Herold-Mende C, Ehemann V, Hollstein M,
Chneiweiss H, et al: The PEA-15/PED protein protects glioblastoma
cells from glucose deprivation-induced apoptosis via the ERK/MAP
kinase pathway. Oncogene. 27:1155–1166. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zeigler ME, Chi Y, Schmidt T and Varani J:
Role of ERK and JNK pathways in regulating cell motility and matrix
metalloproteinase 9 production in growth factor-stimulated human
epidermal keratinocytes. J Cell Physiol. 180:271–284. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Levy DE and Darnell JE Jr: Stats:
Transcriptional control and biological impact. Nat Rev Mol Cell
Biol. 3:651–662. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Iwamaru A, Szymanski S, Iwado E, Aoki H,
Yokoyama T, Fokt I, Hess K, Conrad C, Madden T, Sawaya R, et al: A
novel inhibitor of the STAT3 pathway induces apoptosis in malignant
glioma cells both in vitro and in vivo. Oncogene. 26:2435–2444.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Allen M, Bjerke M, Edlund H, Nelander S
and Westermark B: Origin of the U87MG glioma cell line: Good news
and bad news. Sci Transl Med. 8:354re3532016. View Article : Google Scholar
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-delta delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dai Z, Wu J, Chen F, Cheng Q, Zhang M,
Wang Y, Guo Y and Song T: CXCL5 promotes the proliferation and
migration of glioma cells in autocrine- and paracrine-dependent
manners. Oncol Rep. 36:3303–3310. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu T, Liu PY and Marshall GM: The
critical role of the class III histone deacetylase SIRT1 in cancer.
Cancer Res. 69:1702–1705. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ford E, Voit R, Liszt G, Magin C, Grummt I
and Guarente L: Mammalian Sir2 homolog SIRT7 is an activator of RNA
polymerase I transcription. Genes Dev. 20:1075–1080. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ashraf N, Zino S, Macintyre A, Kingsmore
D, Payne AP, George WD and Shiels PG: Altered sirtuin expression is
associated with node-positive breast cancer. Br J Cancer.
95:1056–1061. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Longo VD and Kennedy BK: Sirtuins in aging
and age-related disease. Cell. 126:257–268. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mansour SJ, Matten WT, Hermann AS, Candia
JM, Rong S, Fukasawa K, Vande Woude GF and Ahn NG: Transformation
of mammalian cells by constitutively active MAP kinase kinase.
Science. 265:966–970. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chang L and Karin M: Mammalian MAP kinase
signalling cascades. Nature. 410:37–40. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen D, Zuo D, Luan C, Liu M, Na M, Ran L,
Sun Y, Persson A, Englund E, Salford LG, et al: Glioma cell
proliferation controlled by ERK activity-dependent surface
expression of PDGFRA. PLoS One. 9:e872812014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang Z, Guo Q, Wang R, Xu G, Li P, Sun Y,
She X, Liu Q, Chen Q, Yu Z, et al: The D Domain of LRRC4 anchors
ERK1/2 in the cytoplasm and competitively inhibits MEK/ERK
activation in glioma cells. J Hematol Oncol. 9:1302016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Glassmann A, Reichmann K, Scheffler B,
Glas M, Veit N and Probstmeier R: Pharmacological targeting of the
constitutively activated MEK/MAPK-dependent signaling pathway in
glioma cells inhibits cell proliferation and migration. Int J
Oncol. 39:1567–1575. 2011.PubMed/NCBI
|
|
30
|
Lin F, Chengyao X, Qingchang L, Qianze D,
Enhua W and Yan W: CRKL promotes lung cancer cell invasion through
ERK-MMP9 pathway. Mol Carcinog. 54:E35–E44. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Justicia C, Gabriel C and Planas AM:
Activation of the JAK/STAT pathway following transient focal
cerebral ischemia: Signaling through Jak1 and Stat3 in astrocytes.
Glia. 30:253–270. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dasgupta A, Raychaudhuri B, Haqqi T,
Prayson R, Van Meir EG, Vogelbaum M and Haque SJ: Stat3 activation
is required for the growth of U87 cell-derived tumours in mice. Eur
J Cancer. 45:677–684. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yu H and Jove R: The STATs of cancer-new
molecular targets come of age. Nat Rev Cancer. 4:97–105. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sherry MM, Reeves A, Wu JK and Cochran BH:
STAT3 is required for proliferation and maintenance of multipotency
in glioblastoma stem cells. Stem cells. 27:2383–2392. 2009.
View Article : Google Scholar : PubMed/NCBI
|