Introduction
China has a high incidence of esophageal cancer; as
the third most common cancer in men, and the fifth in women, the
disease ranks fourth for mortality rate among malignant tumors,
regardless of sex (1). The most
common type of esophageal cancer is squamous cell carcinoma (SCC),
accounting for >90% of cases. Radiation therapy is one of the
principal treatments for advanced esophageal cancer, although
treatment efficacy remains unsatisfactory. Although radiotherapy
has markedly improved, in addition to combination treatment with
chemotherapy and radiotherapy, the 5-year survival rate of patients
with local advanced esophageal SCC treated with concurrent
radiation therapy and chemotherapy is only 23–34% (2). The local recurrence of lesions within
the radiation field caused by radiation resistance is a primary
cause of treatment failure, accounting for ~50% of cases (2). Therefore, enhancing the radiosensitivity
of esophageal cancer cells may improve the success rate of
radiotherapy, and reduce local recurrence of esophageal cancer
following radiotherapy.
A principle cause of radioresistance in esophageal
cancer is the abnormal secretion of radiation-induced vascular
endothelial growth factor (VEGF), which contributes to tumor
angiogenesis and protects tumor vessels from radiation-associated
damage. Studies have revealed that the expression of VEGF is
closely associated with tumor vascular invasion, lymph node
metastasis and poor prognosis (3,4). VEGF
binds to the vascular endothelial growth factor receptor (VEGF-R)
on the endothelial cell membrane to initiate tumor angiogenesis,
and ultimately results in angiogenesis and lymphangiogenesis, and
subsequent radioresistance (4,5).
Apatinib (YN968D1) is a small molecule tyrosine
kinase inhibitor that selectively inhibits the activity of VEGFR-2,
blocking signal transduction following binding to VEGF. This
inhibits the proliferation and migration of endothelial cells,
thereby inhibiting tumor angiogenesis (6). It is indicated that apatinib is a VEGF
inhibitor with the potential to reverse the radioresistance of
esophageal cancer cells. However, whether concurrent radiotherapy
and apatinib is useful for the treatment of esophageal cancer, and
its underlying mechanism of action, remains to be identified.
Therefore, the present study investigated the effects of apatinib
on esophageal squamous cell carcinoma and its impact on the
radiosensitivity of the cancer cells, using the human cell line
KYSE-150; a potential mechanism of action was also explored. The
present study aimed to provide guidance for apatinib use in
enhancing the success rate of radiotherapy and reducing local
recurrence, in addition to improving the prognosis of patients with
esophageal SCC.
Materials and methods
Reagents and equipment
RPMI 1640 culture medium (containing 0.25% EDTA),
trypsin, penicillin and streptomycin was purchased from Gibco
(Thermo Fisher Scientific, Inc., Waltham, MA, USA), and diluted
prior to experimentation. Fetal bovine serum was purchased from
Hangzhou Sijiqing Biological Engineering Materials Co., Ltd.,
(Hangzhou, China). Apatinib solution at a concentration of 10
mmol/l was purchased from MedChemExpress (Monmouth Junction, NJ,
USA). The human VEGF ELISA kit was purchased from NeoBioscience
Technology Co., Ltd., (Shenzen, China; catalog no. EHC-108.96). The
Cell Counting Kit-8 (CCK-8) kit was purchased from Dojindo
Molecular Technologies, Inc., (Kumamoto, Japan). Crystal violet
staining solution, the Annexin V-fluorescein isothiocyanate (FITC)
apoptosis detection kit, and the cell cycle and apoptosis detection
kits were purchased from Beyotime Institute of Biotechnology
(Jiangsu, China). The Primus-H medical linear accelerator (Siemens
AG, Munich, Germany) was used for x-ray irradiation, with an
absorbed dose rate of 1 Gy/min at 100 cm distance between the
source and the target. The flow cytometer was purchased from BD
Biosciences (San Jose, CA, USA).
Cell lines and culture
The human esophageal squamous cell carcinoma cell
line KYSE-150 was purchased from the Cell Bank of Type Culture
Collection of Chinese Academy of Sciences (Shanghai, China; cat.
no; TCHu230) and cultured in RPMI 1640 medium supplemented with 10%
fetal bovine serum and 1% penicillin/streptomycin, in a 37°C
incubator with 5% CO2; cells were passaged every 2–3
days. Cells in the exponential growth phase were used for the
experiment.
ELISA for VEGF secretion
Experimental groupings were as follows: Blank
control, x-ray (6 Gy), apatinib (10 and 20 µmol/l), and apatinib
combined with x-ray. The cells in each group were collected
following x-ray treatment and cultured in fresh medium for 24 h.
Supernatants were collected and 100 µl (200 pg/ml) was added to
each ELISA plate coated with a VEGF monoclonal antibody. The
detection was performed according to the manufacturer's protocol.
The concentration of VEGF in the supernatant was calculated
according to the absorbance of the specimen, which was determined
using a microplate reader at a wavelength of 450 nm.
Cell proliferation assay
Cells in the exponential growth phase were digested
with trypsin and resuspended. A total of 5,000 cells were seeded
into each well of a 96-well culture plate. Following 24 h, the
culture medium in the apatinib group was replaced with 200 µl
medium containing apatinib at concentrations of 5, 10, 20, 30 and
40 µmol/l. The cells of the blank control group were cultured in
complete medium without apatinib. Each group contained six parallel
wells. The plate was incubated in a 37°C incubator containing 5%
CO2 for 24, 48 and 72 h. Following treatment, 10 µl of
CCK-8 solution was added to each well, and the plate was further
cultured for 4 h. The absorbance (A) at 450 nm was measured and the
proliferation inhibition rate was calculated [cell proliferation
inhibition rate (%) = 1-(A value of experimental group-A value of
blank group)/(A value of control group-A value of blank group)
×100%]. The half-maximal inhibitory concentration (IC50)
was calculated. Each experiment was repeated three times
independently.
Colony formation assay
The experiment included the control, x-ray (2, 4, 6,
and 8 Gy), apatinib (10, 20 and 40 µmol/l) and combination groups
(with the same apatinib concentrations as in the apatinib group and
the same x-ray doses as in the x-ray group). Cells were seeded in
60×60 mm cell culture dishes at 500 cells/dish. Following
incubation for 24 h, the medium in the apatinib and combination
groups was replaced with fresh medium containing 10, 20 and 40
µmol/l apatinib. The cells were incubated for an additional 48 h,
followed by x-ray irradiation. Following irradiation, the medium
was replaced with fresh culture medium without apatinib and
incubated for 8 days. The colonies were washed with PBS, fixed with
4% paraformaldehyde a room temperature for 15 min, and stained with
crystal violet at room temperature for 5 min. The colonies
consisting of ≥50 cells were counted under a microscope and the
colony formation rate (%) was calculated as follows: number of
colonies counted/number of cells seeded ×100%. The surviving
fraction (SF) was determined as follows: SF = number of
colonies/number of cells seeded under the same condition × colony
formation rate. The cell survival curves were fitted by
multi-target model SF = 1-(1-e−D/D0)N, the
radiological parameters, including the extrapolation number (N),
inactivation does (D0), quasi domain dose
(Dq) and survival rate (SF2) were determined,
and the radiosensitization ratio was calculated. All experiments
were repeated three times independently.
Cell cycle analysis
The experimental groups included the blank control,
x-ray (4 Gy), apatinib (20 µmol/l) and apatinib combined with x-ray
groups. The cell culture and apatinib treatment duration were the
same as those in the colony formation assay. Following x-ray
irradiation, the cells were cultured in fresh medium for 24 h,
trypsinized, and centrifuged at 106 × g for 5 min. Cells were
washed with PBS and fixed in 75% ethanol at 4°C overnight. Ethanol
was the removed by centrifugation at 106 × g for 5 min. The cells
were washed with PBS and resuspended in propidium iodide (PI)
solution containing RNase (1.87%), incubated at 37°C in the dark
for 30 min, and analyzed with a flow cytometer. FlowJo 7.6 software
(FlowJo LLC, Ashland, OR, USA) was used to calculate the cell cycle
distribution. Each experiment was repeated three times
independently.
Detection of apoptosis using the
Annexin V-FITC assay
Cells were divided into the same four groups as
described above. The cells were incubated for 24 h after a change
of the medium following x-ray irradiation. The cells were
trypsinized and counted under a light microscope (magnification,
×40). The cell suspensions were stained at a density of
1×106 cells/ml using the Annexin V-FITC Apoptosis
detection kit, as per the manufacturer's protocol; subsequent flow
cytometric analysis was performed. The apoptosis rate was analyzed
using FlowJo software. All experiments were repeated three times
independently.
Western blot analysis
Experimental groupings were as described above. Cell
inoculation, treatment with apatinib and x-ray irradiation were the
same as those in the above experiments. Following x-ray
irradiation, the cells were cultured for 24 h. The protein was
extracted from the cells with radioimmunoprecipitation assay buffer
and quantified using a BCA kit (catalog no. P0011; Beyotime
Institute of Biotechnology). A total of 30 µg protein was loaded
per lane for western blot analysis. 10% SDS-PAGE and 5% stacking
gels were prepared, and the protein samples were electrophoresed.
The proteins were then transferred to a polyvinyl difluoride
membrane (catalog no. IPVH00010; EMD Millipore, Billerica, MA, USA)
and blocked with TBS and Tween-20 (Sangon Biotech Co., Ltd.,
Shanghai, China) with 5% skim milk for 1 h at room temperature. The
membranes were then incubated with the following primary
antibodies: Anti-PARP/cleaved-PARP (catalog no. 9915; 1:1,000; Cell
Signaling Technology, Inc., Danvers, MA, USA);
anti-phospho-serine/threonine-protein kinase-2 (pCHK2; catalog no.
9917; 1:1,000; Cell Signaling Technology, Inc.); and anti-VEGFR-2
(catalog no. 2472; 1:1,000; Cell Signaling Technology, Inc.) at 4°C
overnight. Subsequently, the membranes were incubated with
anti-rabbit IgG, horseradish peroxidase-linked secondary antibody
(catalog no. 9915; 1:1,000; Cell Signaling Technology, Inc) for 1.5
h at room temperature. Visualization was performed with enhanced
chemiluminescence reagent (catalog no. 32106; Thermo Fisher
Scientific, Inc.). Image J 1.8.0 processing software (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) was used for grayscale
analysis. Anti-GAPDH (catalog no. 5174; 1:1,000; Cell Signaling
Technology, Inc.) was used as an internal reference. Each
experiment was repeated three times independently.
Statistical analysis
All data are expressed as the mean ± standard
deviation. SPSS version 20.0 was used for the statistical analyses
(IBM Corp., Armonk, NY, USA). For comparisons between two groups,
the Student's t-test was used. One-way analysis of variance was
used for comparisons between ≥2 groups, followed by the
Student-Newman-Keuls test when equal variance was assumed, and
Dunnett's T3 test when equal variance was not assumed.
Liner-regression analysis was used to analyze the cell
proliferation rate. Radiation survival curves were fitted according
to the single hit, multi-target equation (7). P<0.05 was considered to indicate a
statistically significant difference.
Results
Apatinib inhibits VEGF secretion in
KYSE-150 cells in a dose-dependent manner
To evaluate the impact of apatinib on VEGF secretion
in KYSE-150 cells, the cells were treated with varying doses of
apatinib and the level of VEGF secretion determined by ELISA. In
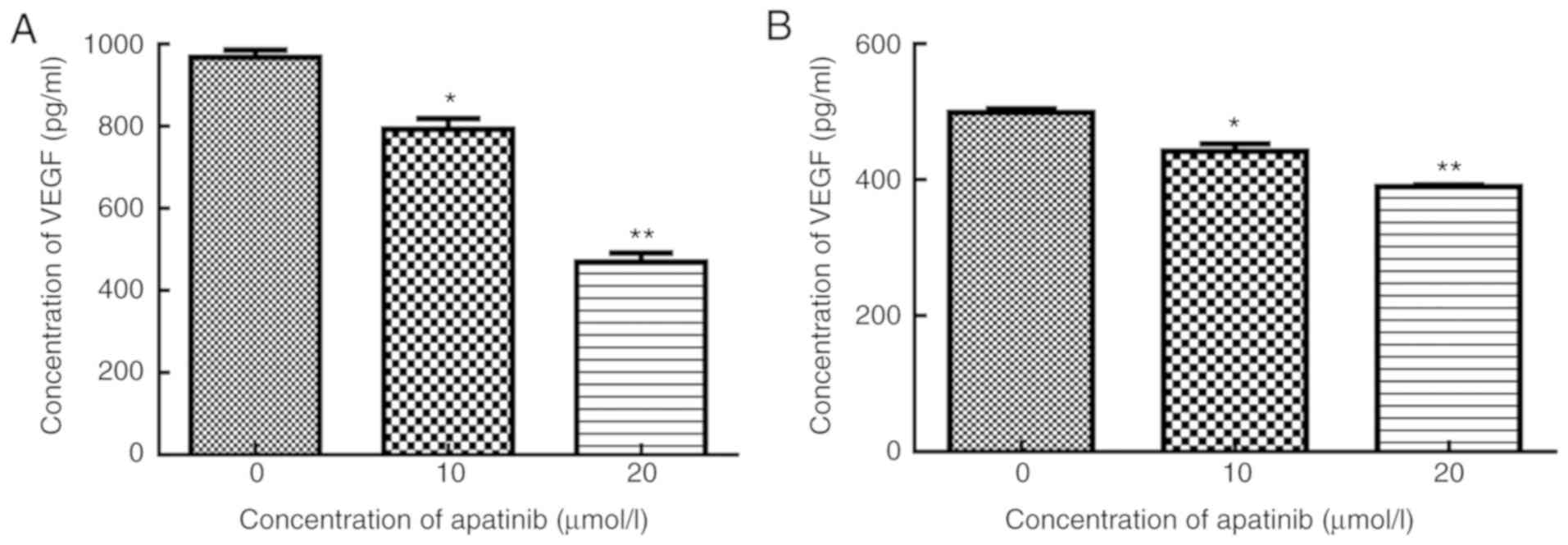
cells treated with 10 and 20 µmol/l apatinib, the levels of VEGF
were 792.6±27.10 and 469.6±22.58 pg/ml, respectively, which were
significantly lower compared with that of the control group,
969.1±7.44 pg/ml (P<0.05). VEGF level was even lower in cells
treated with the combination of apatinib and x-ray. In cells
treated with x-ray and 10 or 20 µmol/l apatinib, the levels of
secreted VEGF were 441.3±11.43 and 390.2±15.54 pg/ml, respectively,
significantly lower compared with that of the x-ray group,
498.8±15.81 pg/ml (t=4.44, 17.12, P<0.05). As illustrated in
Fig. 1, Apatinib inhibited VEGF
secretion in KYSE-150 cells in a dose-dependent manner
(r2=0.96–0.97; P<0.05).
Apatinib inhibits the proliferation
rate of KYSE-150 cells in a dose- and time-dependent manner
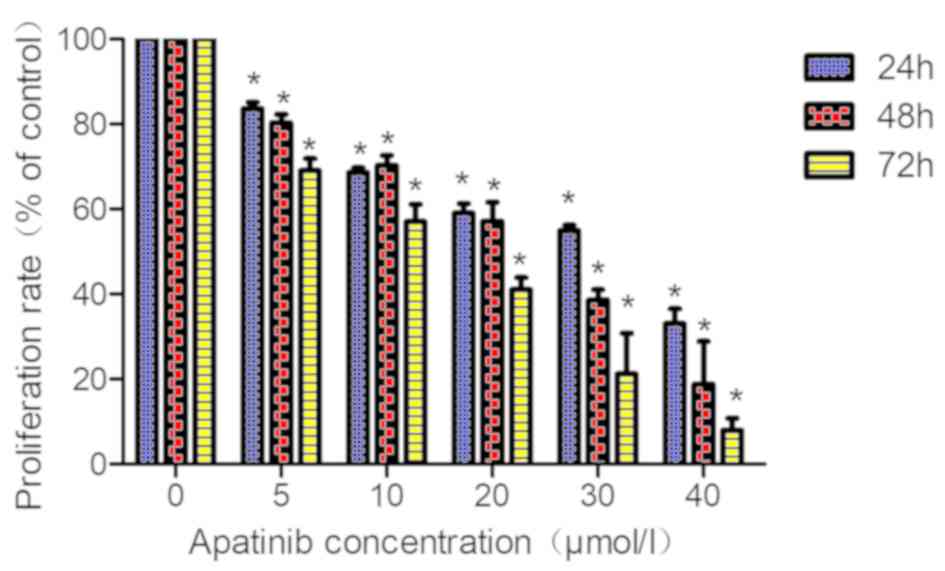
The proliferation rate of KYSE-150 cells was
determined using the CCK-8 assay. With an increase in apatinib
concentration and treatment time, the cell proliferation rate of
KYSE-150 cells decreased significantly (Fig. 2; Table
I). The IC50 values for apatinib in KYSE-150 cells treated for
24, 48 and 72 h were 26.53±0.61, 18.86±0.42, and 11.15±0.26 µmol/l,
respectively, suggesting that the inhibitory effect was dose- and
time-dependent (r2=0.89–0.96; P<0.05). Based on these
findings, KYSE-150 cells were treated with 10, 20 and 40 µmol/l
apatinib for 48 h to further analyze the impact of apatinib on the
radiosensitivity of KYSE-150 cells.
 | Table I.Cell proliferation inhibition rate in
KYSE-150 cells treated with different concentrations of apatinib
(%; mean ± standard deviation). |
Table I.
Cell proliferation inhibition rate in
KYSE-150 cells treated with different concentrations of apatinib
(%; mean ± standard deviation).
|
| Treatment
duration |
|---|
|
|
|
|---|
| Apatinib
concentration (µmol/l) | 24 h | 48 h | 72 h |
|---|
| 0 | 0 | 0 | 0 |
| 5 |
16.40±1.56a |
19.72±2.04a |
31.00±2.87a,c |
| 10 |
31.37±1.10a |
29.69±2.28a |
43.00±4.12a–c |
| 20 |
40.92±2.20a |
42.84±4.48a |
59.00±2.91a–c |
| 30 |
45.08±1.29a |
61.38±2.45a,b |
78.79±9.60a–c |
| 40 |
67.00±3.58a |
81.23±10.08a |
92.06±2.82a,b |
Apatinib enhances the radiosensitivity
of KYSE-150 cells
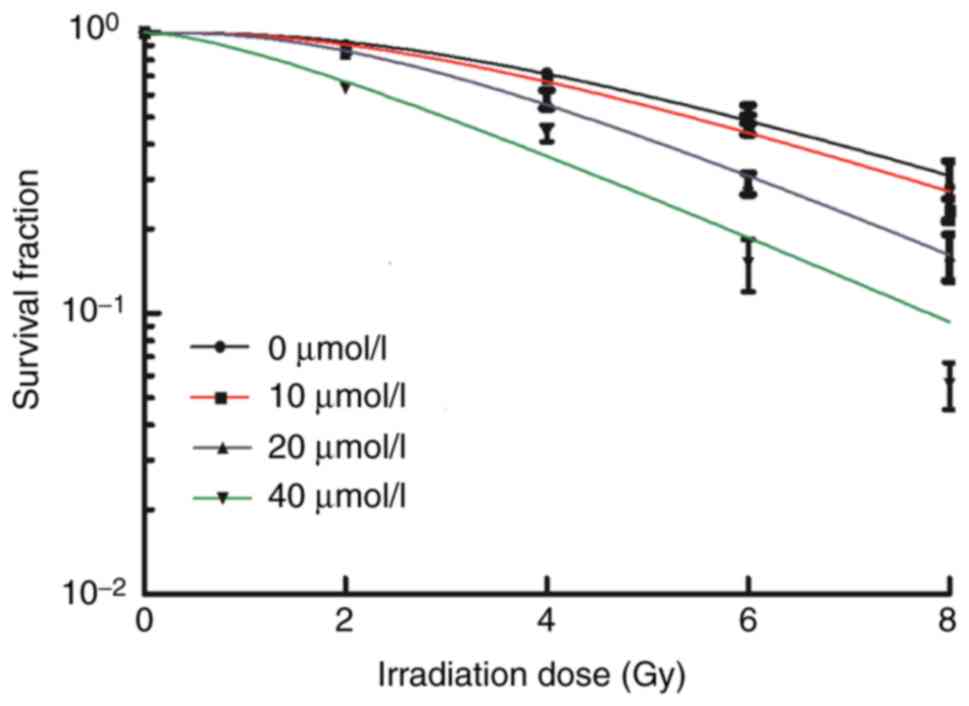
To determine whether apatinib acts as a
radiosensitizer, KYSE-150 cells were treated with apatinib for 48 h
prior to irradiation (2–8 Gy). The radiosensitivity of the cells
was evaluated by the clone formation assay. The survival fraction
of KYSE-150 cells decreased with an increase in radiation dose
(Fig. 3). Compared with the control
group, the higher the concentration of apatinib, the lower the
survival fraction of the cells following x-ray irradiation.
Sensitization enhancement ratios (SER) revealed that in cells
treated with 20 and 40 µmol/l apatinib, the SERD0 were
1.36 and 1.36, respectively, and SERDq were 1.35 and
2.96, respectively. The radiobiological parameters (D0,
Dq and SF2 values) of KYSE-150 decreased with
increased concentrations of apatinib (Table II). These results indicated that
apatinib enhanced the sensitivity of KYSE-150 cells to x-ray
irradiation.
| Table II.Radiosensitization effects of apatinib
in KYSEe-150 cells in vitro. |
Table II.
Radiosensitization effects of apatinib
in KYSEe-150 cells in vitro.
| Apatinib
concentration (µmol/l) |
D0 |
Dq |
SF2 |
SERD0 |
SERDq |
|---|
| 0 | 3.79±0.69 | 4.06±0.27 | 0.90±.019 | 1±0 | 1±0 |
| 10 | 3.60±0.41 | 3.66±0.22 | 0.87±0.02 | 1.05±0.20 | 1.11±0.10 |
| 20 |
2.83±0.34a |
3.00±0.13a | 0.84±0.03 |
1.36±0.36a |
1.35±0.11a |
| 40 |
2.81±0.17a |
1.38±0.23a |
0.66±0.02a |
1.36±0.27a |
2.96±0.30a |
Apatinib enhances radiation-induced
apoptosis in KYSE-150 cells
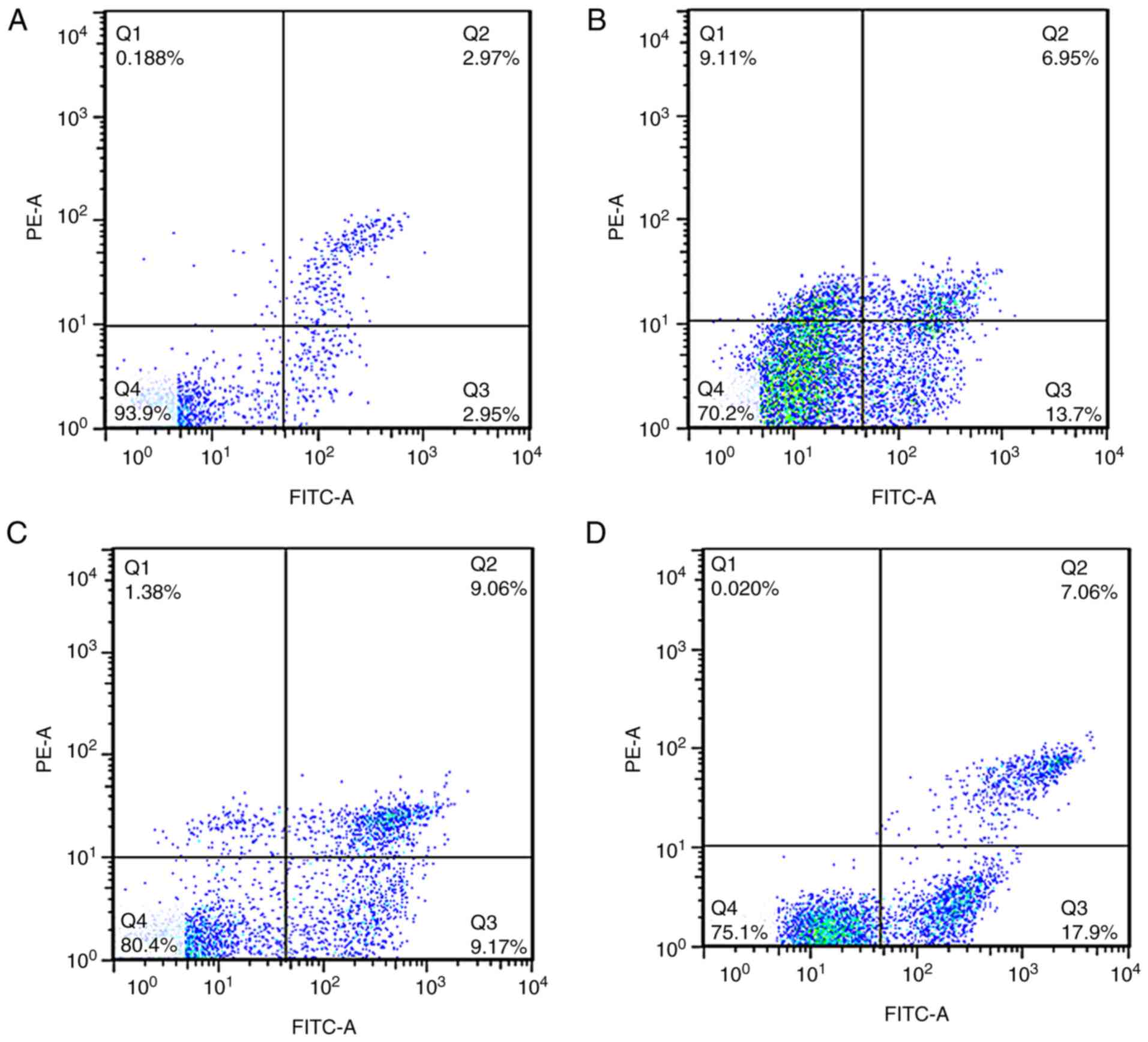
To investigate the mechanism by which apatinib
increased the radiosensitivity of KYSE-150 cells, cells were
pretreated with 20 µmol/l apatinib for 48 h prior to x-ray
irradiation at 4 Gy. Apoptosis was determined using the Annexin
V/PI assay. The apoptosis rates of the apatinib, x-ray, and the
combination of apatinib and x-ray groups were 15.65±1.54, 8.30±1.18
and 19.70±1.66% respectively, which were significantly higher
compared with that of the control group (3.49±0.74%) (Fig. 4). To further investigate the mechanism
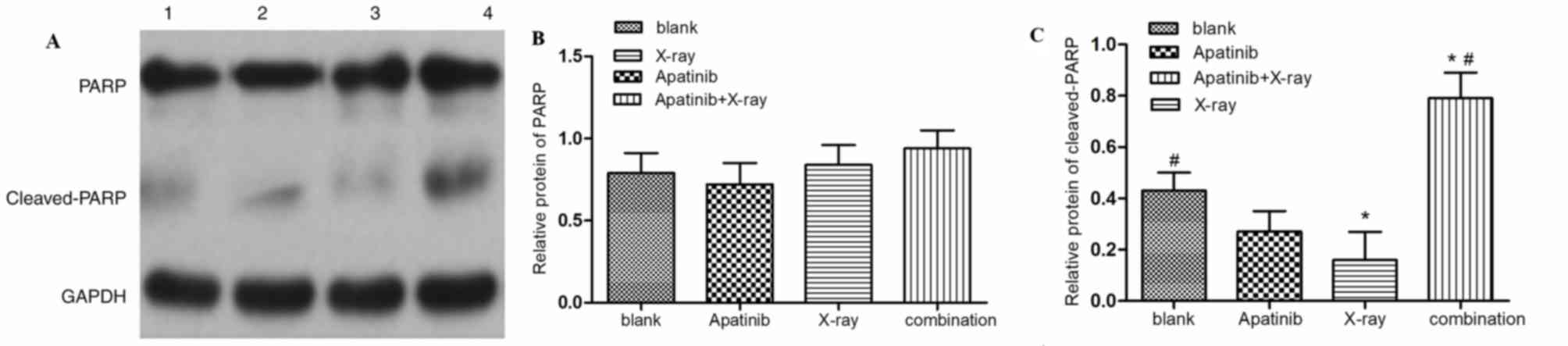
through which apatinib induces apoptosis, the expression of poly
(ADP-ribose) polymerase PARP and cleaved-PARP in KYSE-150 cells was
examined. It was indicted that the level of cleaved-PARP was
markedly increased in the combination group compared with those of
the control and x-ray groups, though the expression of PARP was not
markedly altered, demonstrating that apatinib enhances
PARP-mediated apoptosis in KYSE-150 cells (Fig. 5).
Apatinib accelerates radiation-induced
cell cycle redistribution and causes G2/M-phase arrest in KYSE-150
cells
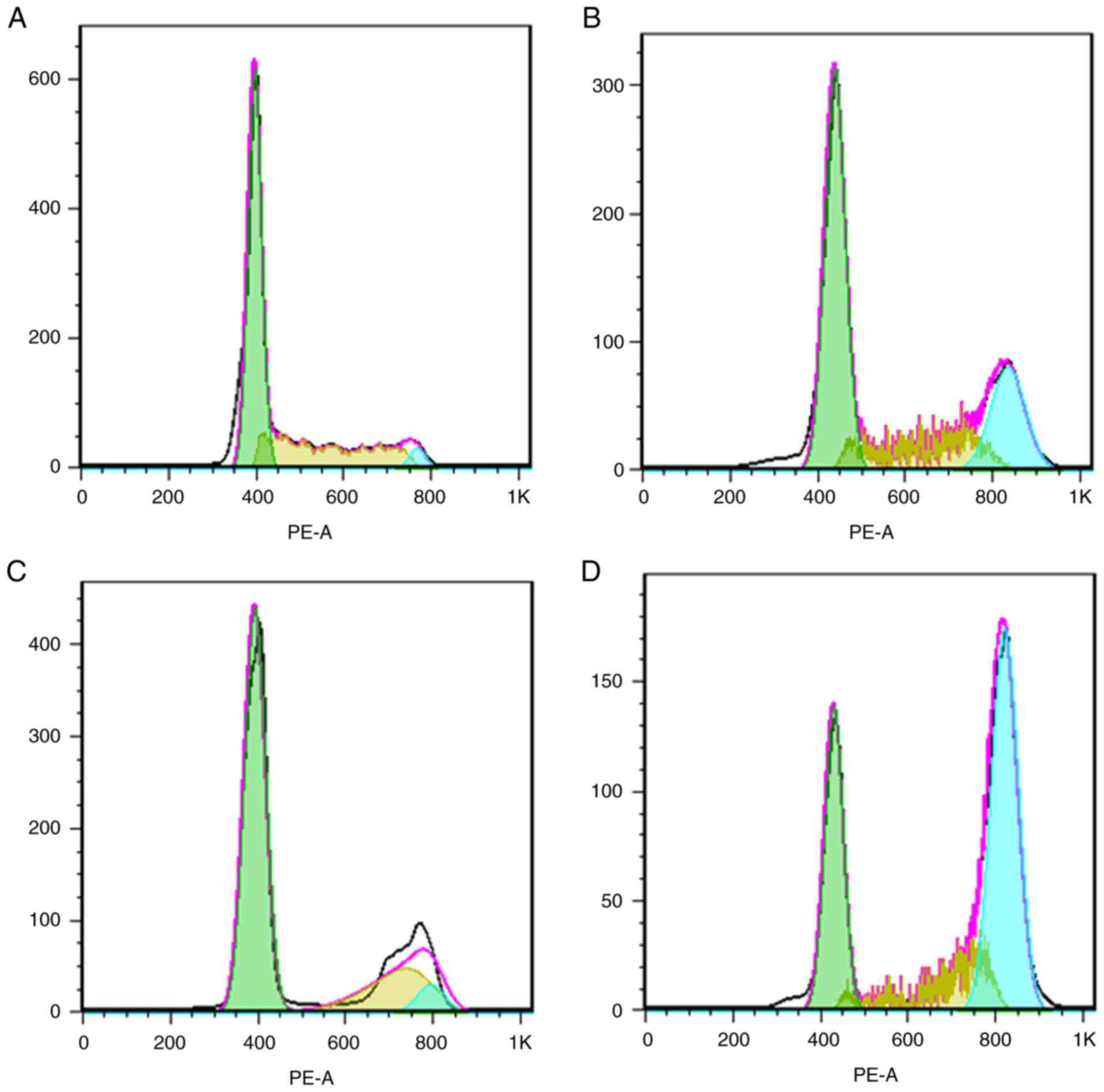
To further identify the mechanism underlying the
radiosensitization effect of apatinib, the cell cycle distribution
of KYSE-150 cells was analyzed by PI staining. As illustrated in
Fig. 6, the proportions of cells in
the G2/M phase within the apatinib, x-ray and combination groups
were 26.27±3.30, 68.79±2.77 and 47.27±3.59% respectively,
significantly higher compared with that of the control group
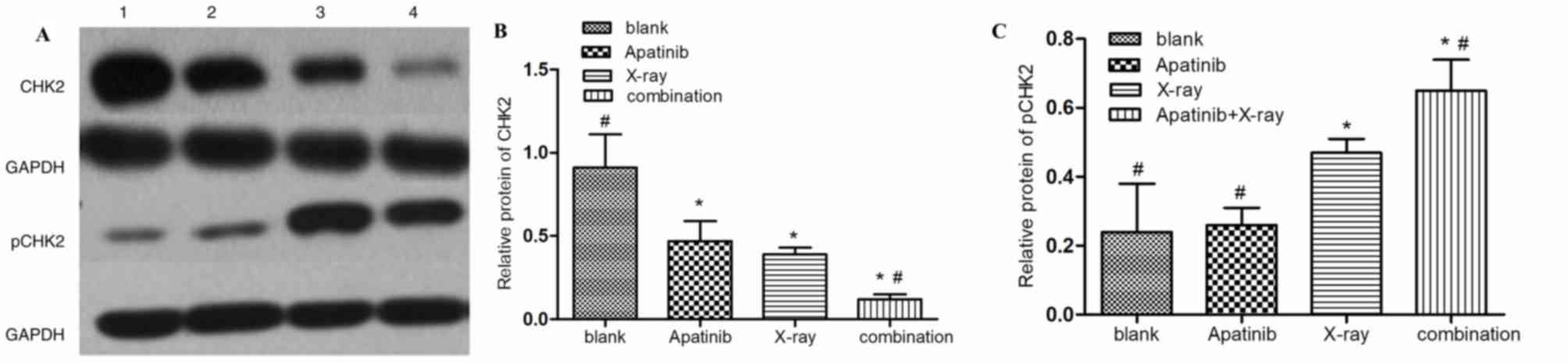
(12.14±2.13%). The expression of pCHK2, an important G2/M phase
checkpoint protein and a central signaling molecule of the DNA
damage response, was markedly higher in the combination group
compared with that in the control and apatinib group (Fig. 7). By contrast, the expression of pCHK2
in the combination group was lower compared with that in x-ray
alone group. This indicated that apatinib may not only increase the
population of KYSE-150 cells in the G2/M phase, but may also
inhibit the DNA repair response (Fig.
6). Data was not shown regarding the influence of Apatinib on
VEGFR-2 expression in KYSE-150 cells.
Discussion
High expression of VEGF is one of the primary causes
of radioresistance in esophageal cancer cells. Studies have
revealed that combination treatment with radiotherapy and anti-VEGF
compounds significantly reduces the radioresistance of esophageal
cancer cells and tissues, thereby enhancing radiosensitivity
(4). In order to investigate the
specific mechanism through which anti-VEGF drugs reduce
radioresistance, the effects of apatinib, a new molecular-targeted
compound, was investigated in combination with radiotherapy for its
ability to alter the radiosensitivity of esophageal cancer cells
in vitro.
Apatinib is a small molecule tyrosine kinase
inhibitor that targets VEGFR-2. The inhibitory effects of apatinib
have been reported in a number of human cancer types including
colon cancer, leukemia and intrahepatic bile duct cancer, wherein
apatinib acts to promote apoptosis and increase the sensitivity of
cancer cells to chemotherapy (8–10).
However, little is known about the effects of apatinib on the
radiosensitization of tumor cells, and its corresponding mechanism
of action. In the present study, analysis of the phosphorylation
level of VEGFR-2 was attempted by western blotting, however, this
was unsuccessful. A previous study suggested that VEGFR-2
expression relies on vascular endothelial cells, and that
experiments in vitro may be unable to detect VEGFR-2
expression (11). As a result, no
further experiments with VEGFR-2 were conducted in the present
study. VEGF is the upstream ligand of VEGFR-2, and it was observed
that apatinib significantly reduced the secretion of VEGF in
KYSE-150 esophageal cancer cells in a concentration-dependent
manner; this suggests that apatinib may have a potential
radiosensitization effect on these cells. Subsequently, a series of
experiments were conducted to determine the radiosensitization
effects of apatinib in KYSE-150 cells, and its potential mechanism
of action. It was indicated that apatinib significantly inhibited
the proliferation of KYSE-150 cells in a dose- and time-dependent
manner. Compared with 24 and 48 h treatment, the inhibitory effects
of a 72 h treatment with apatinib were markedly increased,
suggesting that apatinib acts in a time-dependent manner in
KYSE-150 cells. The survival fraction of KYSE-150 cells decreased
exponentially with increasing radiation exposure. The
radiobiological parameters D0, Dq and
SF2 of KYSE-150 cells gradually decreased with
increasing concentrations of apatinib, whereas the
radiosensitization ratio SERD0 of the cells increased;
the higher the radiosensitization ratio, the higher the
radiosensitivity. The results suggested that apatinib increases the
radiosensitivity of KYSE-150 cells by inhibiting the repair of
sublethal cell damage.
Ionizing radiation-induced cell killing is
predominantly associated with DNA double-strand breaks (DSBs) and
cell cycle redistribution. Apoptosis is regulated by a series of
signaling pathways, including the caspase-9/caspase-3/PARP pathway
(10). Pro-apoptotic signaling
promotes the relocation of cytochrome c oxidase from the
mitochondria to the cytoplasm, and activates cytoplasmic caspase-9,
which cleaves and activates downstream proteins such as caspase-3
(8). Activated caspase-3 cleaves
PARP, which consequently causes the separation of two catalytic
domains at the PARP-carboxy terminus, and a subsequent reduction in
function, which leads to DNA fragmentation and the induction of
apoptosis (8). Studies have suggested
that apoptosis is associated with the radiosensitivity of cells,
and the extent of apoptosis may be used as a measure of
radiosensitivity (9). In the present
study, it was found that apatinib and x-ray promote the apoptosis
of esophageal cancer cells, although the apoptotic effect of
combination treatment was markedly higher compared with that of
apatinib or x-ray alone. Western blotting results further confirmed
that apatinib increased the cleavage and inactivation of the
apoptosis-regulatory protein PARP, accelerating radiation-induced
apoptosis. The exact apoptosis regulatory signaling pathway in
esophageal cancer cells remains unclear, and further research is
required for clarification.
Following irradiation, cancer cells activate DNA
damage response pathways to repair DSBs. Furthermore, cell cycle
checkpoints remove damaged cells from the actively proliferating
population, and halt the cell cycle to temporarily allow for the
repair of DSBs, another primary reason for radioresistance in
cancer cells (12). CHK2 is a key
kinase in this signaling pathway and is an important checkpoint
protein of the G2/M phase. Its activation promotes the repair of
DNA damage and prevents the entry of DNA into mitosis (13). It is therefore a key target in
radiobiology. The present study illustrated that following x-ray
irradiation, the proportion of cells in the G2/M phase was
significantly increased, and pretreatment with apatinib prior to
irradiation significantly increased radiation-induced G2/M arrest.
Meanwhile, apatinib considerably downregulated the
radiation-induced phosphorylation of CHK2. This further confirmed
that apatinib promotes cell cycle arrest in esophageal cancer cells
by inhibiting CHK2 and through the subsequent repair of DNA damage,
thereby accelerating apoptosis.
Apatinib is a novel drug developed in China, which
has been widely used for the treatment of advanced gastric cancer,
colorectal cancer, non-small cell lung cancer and breast cancer
(10,14,15).
However, its specific mechanism of action and target when combined
with radiotherapy remain unclear. In the present study, it was
demonstrated that apatinib markedly increased radiosensitivity by
inhibiting the secretion of VEGF and the proliferation of KYSE-150
cells. When combined with x-ray irradiation, apatinib markedly
inhibited cancer cell survival, and induced apoptosis by activating
PARP-mediated apoptotic signaling pathways and cell cycle
redistribution, which corresponded to the reduced level of pCHK2.
Therefore an association between VEGF and CHK2 is hypothesized,
although the specific mechanism is unclear. This provides scope for
further studies on the radiosensitizing mechanism of apatinib for
the treatment of esophageal cancer, and provides a theoretical
basis for novel strategies for esophageal cancer combination
therapy. As a small molecular tyrosine kinase inhibitor, it is also
important to verify the effects of apatinib on the expression level
of the receptor tyrosine kinase c-Kit, and the proto-oncogene
tyrosine-protein kinase c-SRC, which is a limitation of the present
study that requires further investigation.
Acknowledgements
Not applicable.
Funding
This work was supported by The Natural Science
Foundation of China (grant no. 11705095), The Key Project of
Science and Technology Development Fund of Nanjing Medical
University (grant no. 2017NJMUZD039), The Changzhou Science and
Technology Support-Social Development Project (grant no.
CE20165024); The Changzhou High-level Health Talent Project (grant
no. 2016C2BJ007); and The Applied Basic Research Project of
Changzhou Science and Technology Bureau (grant no. CJ20159050).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FS conducted the majority of the experiments; ZS, SN
and JW assisted with the experiments; LH and JW conducted
statistical analysis and guided the research process; LH drafted
the manuscript and proposed revision suggestions; MZ, YF, ZK and QH
assisted in the completion of experiments; JY participated in the
development of the research idea and experimental design, and
proposed revision suggestions.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Welsh J, Settle SH, Amini A, Xiao L,
Suzuki A, Hayashi Y, Hofstetter W, Komaki R, Liao Z and Ajani JA:
Failure patterns in patients with esophageal cancer treated with
definitive chemoradiation. Cancer. 118:2632–2640. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cellini F and Valentini V: Targeted
therapies in combination with radiotherapy in oesophageal and
gastroesophageal carcinoma. Curr Med Chem. 21:990–1004. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yu J, Liu F, Sun Z, Sun M and Sun S: The
enhancement of radiosensitivity in human esophageal carcinoma cells
by thalidomide and its potential mechanism. Cancer Biother
Radiopharm. 26:219–227. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yu JP, Sun SP, Sun ZQ, Ni XC, Wang J, Li
Y, Hu LJ and Li DQ: Clinical trial of thalidomide combined with
radiotherapy in patients with esophageal cancer. World J
Gastroenterol. 20:5098–5103. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tian S, Quan H, Xie C, Guo H, Lu F, Xu Y,
Li J and Lou L: YN968D1 is a novel and selective inhibitor of
vascular endothelial growth factor receptor-2 tyrosine kinase with
potent activity in vitro and in vivo. Cancer Sci. 102:1374–1380.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Leith JT, Davis PJ, Mousa SA and Hercbergs
AA: In vitro effects of tetraiodothyroacetic acid combined with
X-irradiation on basal cell carcinoma cells. Cell Cycle.
16:367–373. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ghorai A, Sarma A, Bhattacharyya NP and
Ghosh U: Carbon ion beam triggers both caspase-dependent and
caspase-independent pathway of apoptosis in HeLa and status of
PARP-1 controls intensity of apoptosis. Apoptosis. 20:562–580.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rahmanian N, Hosseinimehr SJ and Khalaj A:
The paradox role of caspase cascade in ionizing radiation therapy.
J Biomed Sci. 23:882016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lin Y, Zhai E, Liao B, Xu L, Zhang X, Peng
S, He Y, Cai S, Zeng Z and Chen M: Autocrine VEGF signaling
promotes cell proliferation through a PLC-dependent pathway and
modulates apatinib treatment efficacy in gastric cancer.
Oncotarget. 8:11990–12002. 2017.PubMed/NCBI
|
|
11
|
Lin YC, Liu CY, Kannagi R and Yang RB:
Inhibition of endothelial SCUBE2 (Signal Peptide-CUB-EGF
Domain-Containing Protein 2), a novel VEGFR2 (Vascular Endothelial
Growth Factor Receptor 2) coreceptor, suppresses tumor
angiogenesis. Arterioscler Thromb Vasc Biol. 38:1202–1215. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Morgan MA and Lawrence TS: Molecular
pathways: Overcoming radiation resistance by targeting DNA damage
response pathways. Clin Cancer Res. 21:2898–2904. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Manic G, Obrist F, Sistigu A and Vitale I:
Trial watch: Targeting atm-chk2 and atr-chk1 pathways for
anticancer therapy. Mol Cell Oncol. 2:e10129762015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhou N, Liu C, Hou H, Zhang C, Liu D, Wang
G, Liu K, Zhu J, Lv H, Li T and Zhang X: Response to apatinib in
chemotherapy-failed advanced spindle cell breast carcinoma.
Oncotarget. 7:72373–72379. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Song Z, Yu X, Lou G, Shi X and Zhang Y:
Salvage treatment with apatinib for advanced non-small-cell lung
cancer. Onco Targets Ther. 10:1821–1825. 2017. View Article : Google Scholar : PubMed/NCBI
|