Introduction
Lung cancer is the principal cause of
cancer-associated mortality worldwide and is associated with high
morbidity. Non-small cell lung cancer (NSCLC) accounts for 80–85%
of all lung cancer cases (1).
According to a previous study, >234,030 new lung cancer cases
are predicted to occur every year, and 154,050 cases of lung
cancer-associated mortality are predicted in the USA (2). Furthermore, the World Health
Organization predicts that 1,000,000 cases of lung
cancer-associated mortality may be reached by 2025 in China
(3). Although advances have been
made in conventional cancer therapies, including diagnostic
imaging, surgery, radiotherapy and chemotherapy, the 5-year
survival rate of patients with NSCLC remains poor, mostly due to
drug resistance and tumor metastasis (4). Metastasis is a sign of malignancy and a
common characteristic of lung cancer, and is the major reason for
treatment failure and patient mortality. Approximately 30% of
patients present distant metastases at admission, 50–60% develop
distant metastases during treatment and 80–90% succumb to
metastases (5). Recently, specific
targeted drugs have been developed for lung cancer treatment,
including the epidermal growth factor receptor, and the tumor
angiogenesis inhibitors gefitinib and bevacizumab (6). However, the high recurrence and
metastasis rates seriously affect the quality of life and overall
survival of patients (7).
Understanding the molecular mechanisms underlying lung cancer may
provide novel strategies for lung cancer treatment, and improve the
survival rate and quality of life for patients with lung cancer;
however, such advances remain largely elusive. Understanding such
mechanisms may provide a foundation for the development of novel
therapeutic approaches.
Asiatic acid (AA) is a pentacyclic triterpenoid
extracted from the umbelliferous plant Centella asiatica
(L.) Urban. Previous studies have demonstrated that AA serves a
role in inhibiting lung cancer cell growth in vitro and
in vivo through mitochondrial damage (8,9). In
addition, it has been suggested that AA possesses pharmacological
activities, including inhibition of cancer proliferation,
apoptosis-inducing effects and anti-metastatic effects in various
types of tumor (10–12). Previous studies have suggested that
epithelial-mesenchymal transition (EMT) serves a crucial role in
primary invasion and secondary metastasis of various types of
cancer. EMT is characterized by reduced expression of the cell
adhesion molecule E-cadherin, increased expression of the
cytoskeletal component vimentin and enhanced mesenchymal cell
morphology (13–15). Tumor metastasis results from
molecular structure modifications that promote cell invasion and
diffusion to other areas. Identification of factors regulating EMT
would therefore be highly valuable for the treatment of tumor
metastasis. EMT is controlled by various transcription factors,
including transforming growth factor-β1 (TGF-β1). TGF-β1 is a
member of the TGF-β superfamily that contributes to EMT during
embryonic development and induces EMT during tumor progression
(16). AA has inhibitory effects on
various types of tumor; however, to the best of our knowledge, its
antitumor activity through EMT inhibition in cancer cells remains
unknown (17,18).
In the present study, the human alveolar epithelium
A549 cell line was used to study the anticancer effects and
underlying mechanisms of AA. To do so, the TGF-β1-induced EMT model
was used to explore the antitumor effects of AA on EMT and its
efficacy against lung cancer.
Materials and methods
Cells and reagents
The human A549 lung cancer cell line was purchased
from the Cell Bank, Shanghai Institute of Life Science, Chinese
Academy of Science (Shanghai, China). Cells were maintained in
Roswell Park Memorial Institute (RPMI)-1640 culture medium (Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with
10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.)
and 100 U/ml penicillin/streptomycin (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany), and were incubated at 37°C in a humidified
atmosphere containing 5% CO2. Cells in the exponential
growth phase (~80% confluence) were used in all experiments. AA was
purchased from Sigma-Aldrich; Merck KGaA.
Establishment of the EMT model of A549
cells
The A549 cells were washed with PBS and cultured
with 1 ml 0.25% trypsin. The trypsin was then removed and the cells
were resuspended in complete medium. After complete digestion,
cells in the logarithmic growth phase were harvested and seeded in
6-well plates at a density of 8×105 cells/well in 2 ml
medium. Following overnight incubation, cells were divided into
three groups, as follows: A negative control group, a
TGF-β1-treated group (10 ng/ml) and an AA + TGF-β1-treated group
(20 µmol/l AA + 10 ng/ml TGF-β1). Each condition was set up in
triplicate. Cells were treated for 24 h, after which, A549 cell
morphology and growth were observed and images were captured under
an inverted microscope (Leica Microsystems GmbH, Wetzlar,
Germany).
Cell viability assay
Cell viability was measured using the colorimetric
MTT assay as described previously (19). After complete digestion, cells in the
logarithmic growth phase were harvested and seeded in 96-well
plates at a density of 1×104 cells/well in 100 µl
medium, and incubated in serum-free medium for 24 h. Cells were
then treated with increasing concentrations of AA (5, 10, 20, 40
and 80 µmol/l) with or without TGF-β1 or medium for 24 h. MTT
reagent (100 µl, 1 mg/ml) was added to the cells for 4 h.
Eventually, the supernatant was removed and the purple formazan
crystals generated by viable cells were dissolved with 100 µl
dimethyl sulfoxide, prior to measuring absorbance at 570 nm with a
microplate reader (Multiskan Spectrum; Molecular Devices, LLC,
Sunnyvale, CA, USA).
Wound healing assay
A549 cell migration was assessed with the wound
healing assay as previously described (20). Briefly, A549 cells were seeded in
12-well plates at a density of 1×105 cells/well in 1 ml
medium and cultured in an incubator for 24 h. Subsequently, a
horizontal wound was generated with a 10-µl pipette tip, cells were
washed with PBS to remove cell debris, and were incubated with
culture medium (control) or TGF-β1 with or without AA (10, 20 or 40
µmol/l) for 24 h. The migration status was observed under a
fluorescence microscope at 0 and 24 h. The wound healing rate was
used to evaluate cell migration ability and quantified as follows:
(wound width at 0 h-wound width at 24 h)/wound width at 0 h with
ImageJ V1.8.0 software (National Institutes of Health, MD, USA)
(21). Experiments were performed
three times.
Transwell assay
Cell migratory and invasive capacities were assessed
with specialized Transwell chambers (8-µm pores; BD Biosciences,
San Jose, CA, USA). For the migration assay, 150 µl cell suspension
(1×105 cells/ml) containing the appropriate drugs
[TGF-β1 (10 ng/ml) or AA + TGF-β1 (10, 20 or 40 µmol/l AA + 10
ng/ml TGF-β1] was added to the upper chamber of a Transwell system,
and 450 µl culture medium containing 15% FBS was added to the lower
chamber. After 48 h incubation, any non-migrating or non-invading
cells on the upper surface were removed. Cells in the lower chamber
were fixed with 4% paraformaldehyde for 30 min at room temperature,
and stained with the 0.1% crystal violet for 30 min at room
temperature. The number of migrated cells was counted randomly
under a light microscope (magnification, ×200, three fields/well).
For the invasion assay, the upper chamber was coated with 40 µl BD
Matrigel™ (BD Biosciences), which was initially dissolved in medium
(1:8) and allowed to adhere to the chamber at 37°C for 4 h, and 600
µl full medium containing 15% FBS was added to the lower chamber.
After 48 h, uninvaded cells in the upper chamber were wiped off
with cotton swabs, and invaded cells were fixed with 4%
paraformaldehyde for 30 min at room temperature, and stained with
0.1% crystal violet for 30 min at room temperature. The number of
invaded cells was counted under a light microscope (magnification,
×200).
Western blotting
After 24 h of treatment with AA (10, 20 and 40
µmol/l) with or without TGF-β1 (10 ng/ml), cells were washed twice
with ice-cold PBS, centrifuged at 12,000 × g for 10 min at 4°C,
lysed with radioimmunoprecipitation assay buffer (Beijing Solarbio
Science & Technology Co., Ltd., Beijing, China) and incubated
on ice for 20 min. The protein concentration was determined with a
bicinchoninic acid protein assay kit (Thermo Fisher Scientific,
Inc.). Total proteins (40 µg) were separated by 10% SDS-PAGE and
transferred onto a 0.45-µm polyvinylidene fluoride membrane (EMD
Millipore, Billerica, MA, USA). The membrane was blocked in a
buffer containing 0.3 g bovine serum albumin (Sigma-Aldrich; Merck
KGaA), 20 ml PBS with 20 µl 0.1% Tween-20 (PBST; Beijing Solarbio
Science & Technology Co., Ltd.) and 1 g non-fat milk at room
temperature for 2 h. The membrane was then incubated with the
primary antibodies against E-cadherin (1:1,000; cat. no. ab1416),
N-cadherin (1:1,000; cat. no. ab18203), β-catenin (1:1,000; cat.
no. ab16051), vimentin (1:1,000; cat. no. ab8978), snail family
transcriptional repressor (Snail; 1:1,000; cat. no. ab53519),
glycogen synthase kinase-3β (GSK-3β; 1:1,000; cat. no. ab93926) and
GAPDH (1:2,000; cat. no. ab8245) all from Abcam, Cambridge, MA,
USA, and phosphorylated-GSK-3β (p-GSK-3β; 1:1,000; cat. no. CST
#9336, Cell Signaling Technology, Inc., Danvers, MA, USA) overnight
at 4°C. The membrane was then washed three times for 15 min with
PBST, and incubated with a horseradish peroxidase (HRP)-conjugated
secondary antibody (1:1,500; Abcam; cat. no. ab9482) for 2 h at
room temperature. Enhanced chemiluminescence reagent (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) was used to detect the
signal on the membrane. The data were analyzed via densitometry
using the Gel Image Analysis system (Tanon 2500R, Shanghai, China)
and normalized to the expression of the internal control
(GAPDH).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated from A549 cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) as previously described (22).
cDNA was synthesized using a PrimeScript™ RT reagent kit (Takara
Bio, Inc., Otsu, Japan) according to the manufacturer's protocol.
cDNA was diluted 10 times and the SYBR-Green Supermix (Bio-Rad
Laboratories, Inc.) was used to perform PCR using the Opticon 2
Real-time PCR Detection system (Bio-Rad Laboratories, Inc.).
RT-qPCR reactions were performed as follows: Pre-denaturation at
95°C for 10 min, followed by 40 cycles of denaturation at 95°C for
10 sec, annealing at 58°C for 30 sec and extension at 72°C for 30
sec. The primers used for this experiment were designed as follows:
Snail, forward, 5′-GAGGACAGTGGGAAAGGCTC-3′, reverse,
5′-TGGCTTCGGATGTGCATCTT-3′; E-cadherin, forward,
5′-GGGGTCTGTCATGGAAGGTG-3′, reverse, 5′-CAAAATCCAAGCCCGTGGTG-3′;
N-cadherin, forward, 5′-GGGAAATGGAAACTTGATGGCA-3′, reverse,
5′-GGAGGGATGACCCAGTCTCT-3′; vimentin, forward,
5′-CTCTGGCACGTCTTGACCTT-3′, reverse, 5′-TTGCGCTCCTGAAAAACTGC-3′;
β-catenin, forward, 5′-GTGACTCTCGGAGCGGGA-3′, reverse,
5′-CAGGCAAACAGGTGCTCAAC-3′; GSK-3β, forward,
5′-GACTAAGGTCTTCCGACCCC-3′, reverse, 5′-TTAGCATCTGACGCTGCTGT-3′;
and GAPDH, forward, 5′-AATGGGCAGCCGTTAGGAAA-3′ and reverse,
5′-GCGCCCAATACGACCAAATC-3′. The relative mRNA expression levels
were analyzed using the comparative cycle quantification (Cq)
(2−∆∆Cq) method (23) and
normalized to the endogenous control. Each sample was set up in
triplicate and the experiments were repeated three times.
Statistical analysis
All experiments were performed three times and the
data are expressed as the means ± standard deviation. Unless stated
otherwise, statistical analysis was performed using GraphPad Prism
5.01 software (GraphPad Software, Inc., La Jolla, CA, USA).
Multiples comparisons were performed by one-way analysis of
variance followed by Tukey's post hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Effect of TGF-β1 on inhibition of A549
cells viability
Cells were treated with increasing concentrations of
AA (0, 5, 10, 20, 40 and 80 µmol/l) with or without TGF-β1 (10
ng/ml) for 24 h. MTT assay was used to measure the effect of AA on
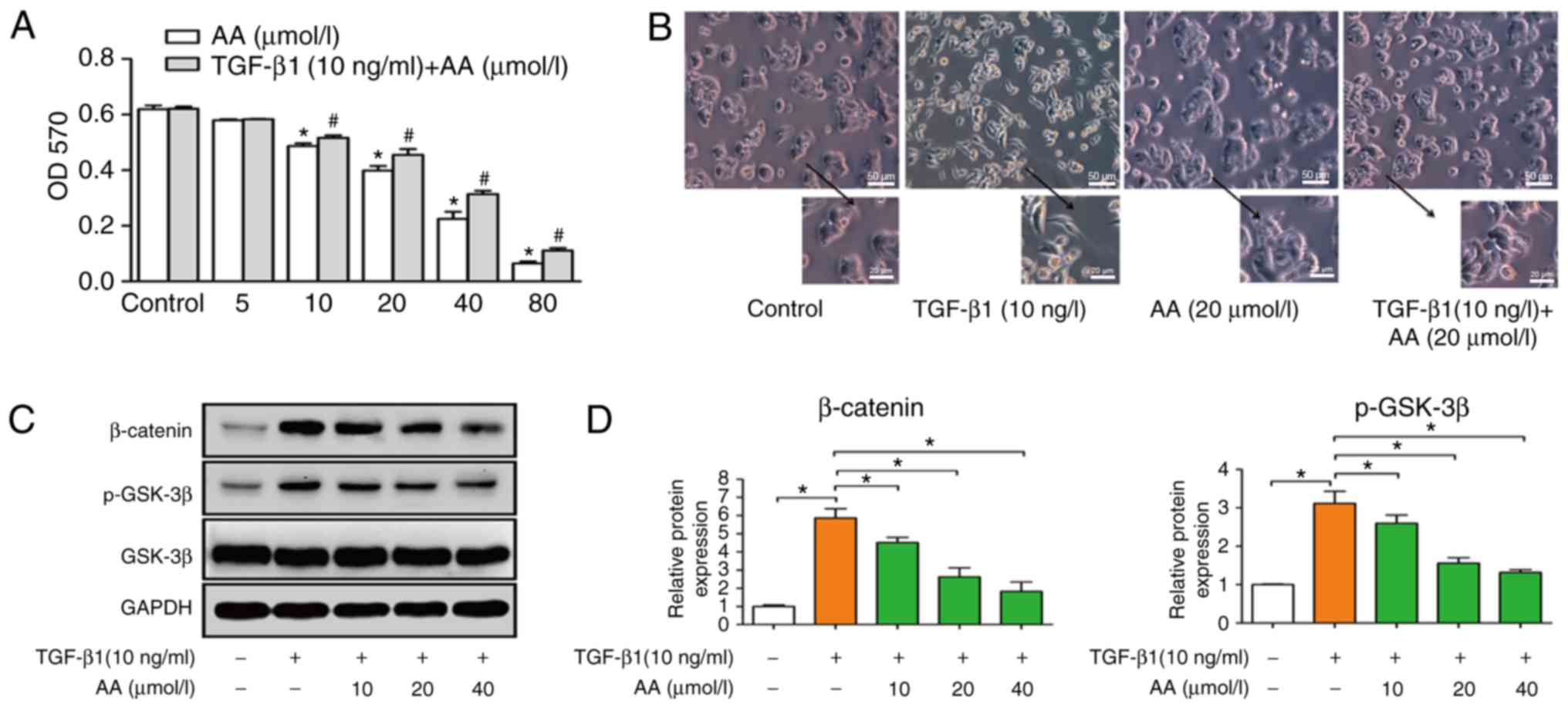
A549 cell viability (Fig. 1A).
Results demonstrated that AA significantly decreased cell viability
in a dose-dependent manner; notably, a high concentration of AA
inhibited the viability of A549 cells. Since the present study
aimed to investigate whether a low concentration of AA could
inhibit TGF-β1-induced EMT, the concentrations of AA used in
subsequent experiments were 10, 20 and 40 µmol/l. The viability of
A549 cells treated with TGF-β1 (10 ng/ml) was not effected.
TGF-βl induces EMT of A549 cells
A549 cells were treated with TGF-β1 (10 ng/ml) for
24 h to induce EMT. A549 cell morphology following TGF-β1 treatment
was assessed under an inverted microscope. Results demonstrated
that TGF-β1 induced a change in cell morphology, from the
cobblestone to the fusiform shape, similar to fibroid cells. These
results demonstrated that TGF-β1 allowed modification of the
epithelial A549 cells into a mesenchymal phenotype. As presented in
Fig. 1B, the control group exhibited
an epithelial cell morphology with strong intercellular adhesion;
however, following TGF-β1 treatment, cell morphology changed to
fusiform, exhibiting the morphological characteristics of
interstitial cells with few intercellular connections. Conversely,
AA inhibited TGF-β1-induced EMT, as observed by cells that retained
an epithelial morphology with few fusiform forms and increased
intercellular adhesion, which confirmed that EMT was induced
following 24 h treatment with TGF-β1 (Fig. 1B). In addition, AA treatment had no
effect on A549 cell morphology compared with the control group.
Furthermore, western blotting demonstrated that β-catenin and
p-GSK-3β protein expression levels were decreased by AA, while no
difference in the expression level of GSK-3β was observed (Fig. 1C and D).
Effect of AA on migration and invasion
of TGF-β1-treated A549 cells
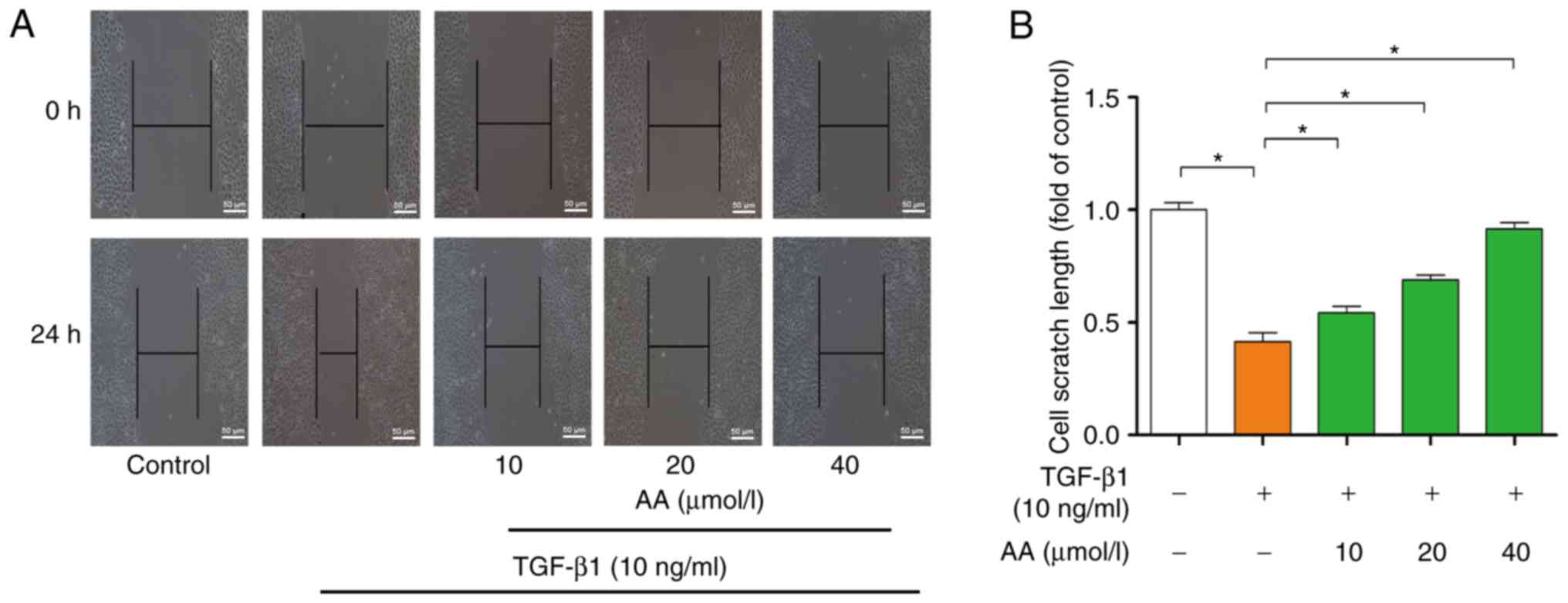
The effects of AA on cell migration are presented in
Fig. 2A. Compared with in the
control group, 24 h TGF-β1 treatment significantly promoted A549
cell migration (P<0.05). However, AA treatment significantly
(P<0.05) inhibited TGF-β1-induced A549 cell migration following
24 h of AA treatment. The effects of AA on scratch length are
presented in Fig. 2B. Results
indicated that AA significantly inhibited TGF-β1-induced A549 cell
migration in a dose-dependent manner.
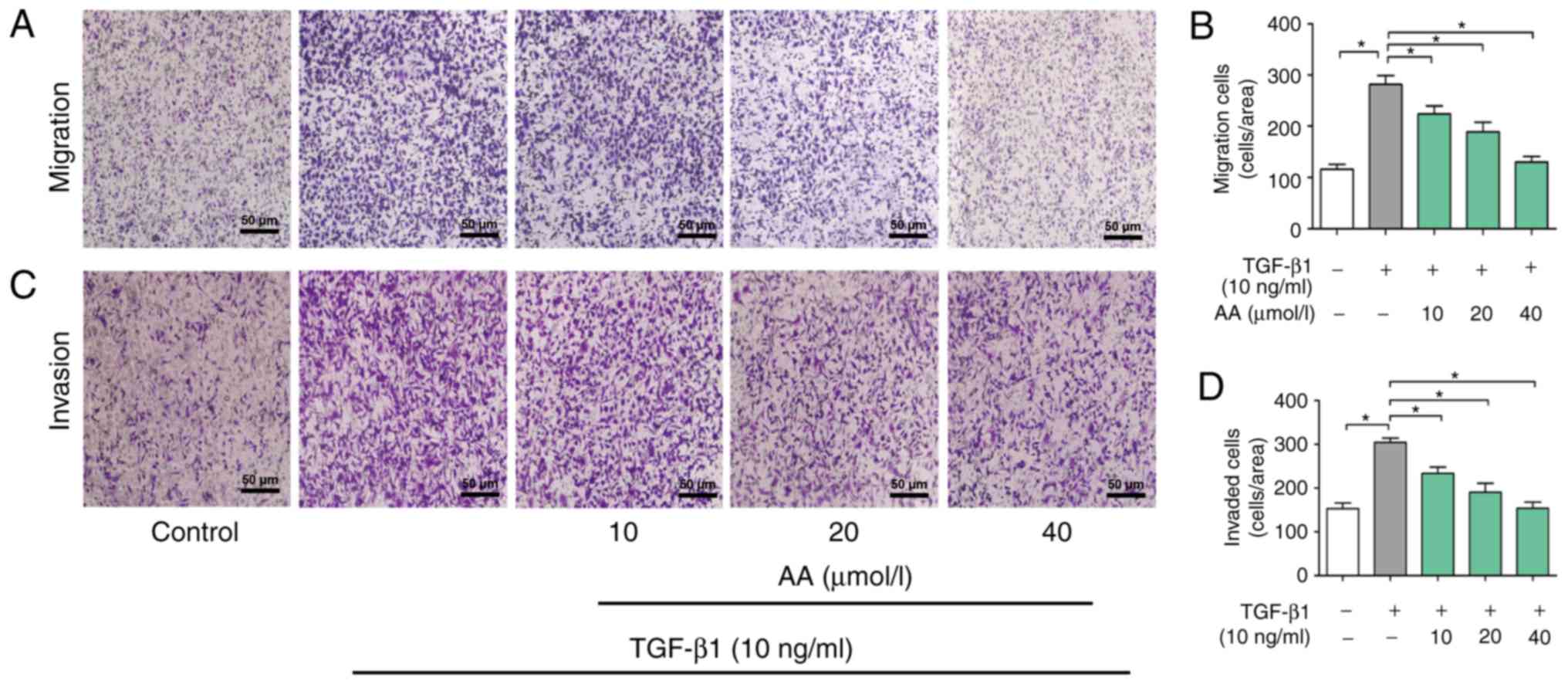
The role of AA on invasion of A549 cells was then
determined. As presented in Fig. 3A and
B, the migration of cells in the AA+ TGF-β1 treatment group was
significantly inhibited compared with the cells in the TGF-β1
treatment group (P<0.05). Furthermore, TGF-β1 treatment
significantly promoted A549 cell invasion in a dose-dependent
manner (P<0.05). Conversely, 24 h treatment with AA
significantly inhibited TGF-β1-induced cell invasion in a
dose-dependent manner (P<0.05; Fig.
3C and D).
AA regulates EMT-associated protein
expression in A549 cells
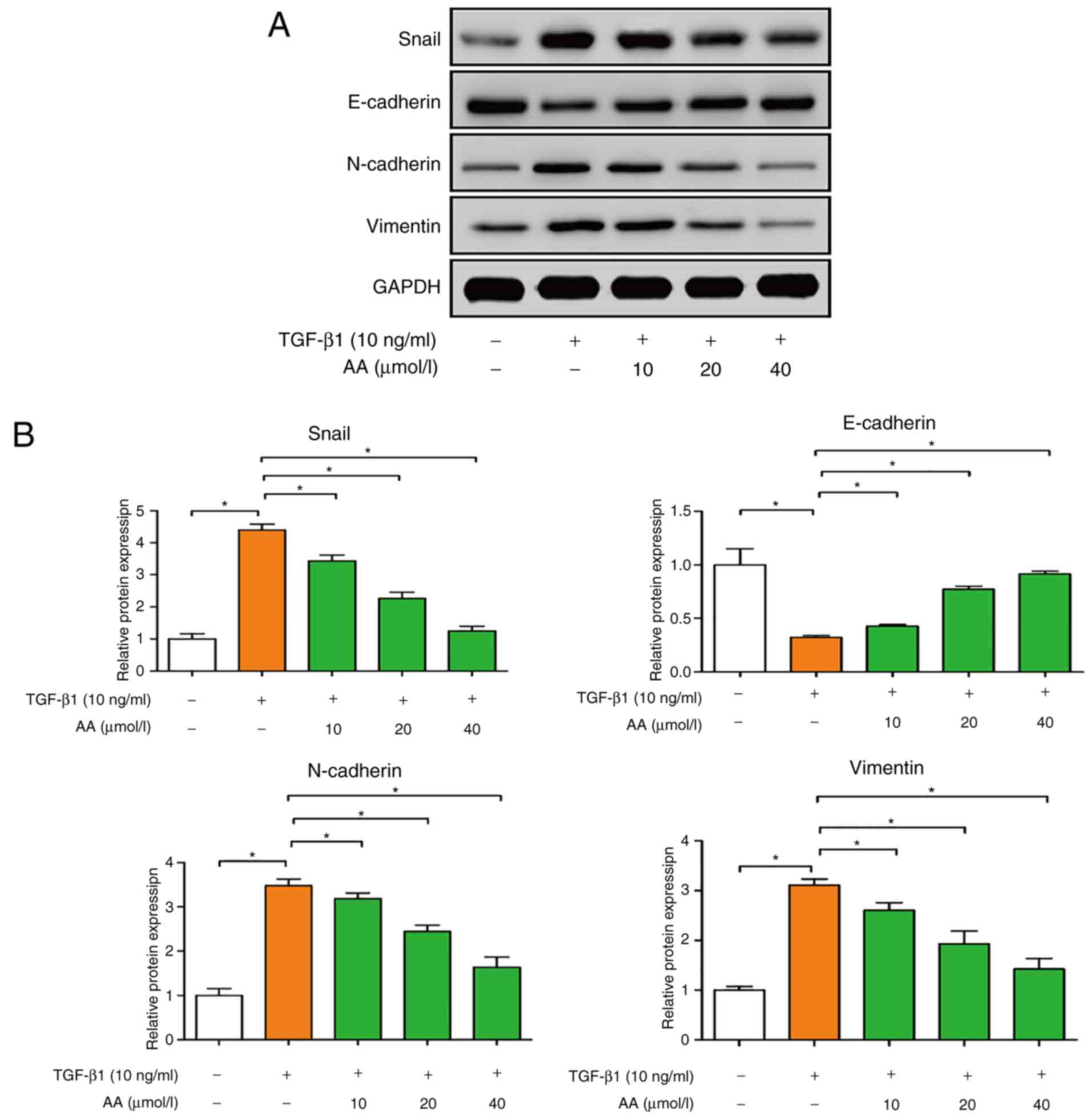
The effects of AA on the expression of EMT protein
markers in A549 cells were examined by western blotting. As
presented in Fig. 4A and B,
TGF-β1-treated cells exhibited significantly decreased E-cadherin
expression (P<0.05), and increased N-cadherin, vimentin and
Snail expression (P<0.05), compared with the control group.
Conversely, AA-treated cells presented a significant increase in
E-cadherin expression (P<0.05), and a significant decrease in
N-cadherin, vimentin and Snail expression (P<0.05). These
results suggested that AA may significantly inhibit EMT of A549
cells.
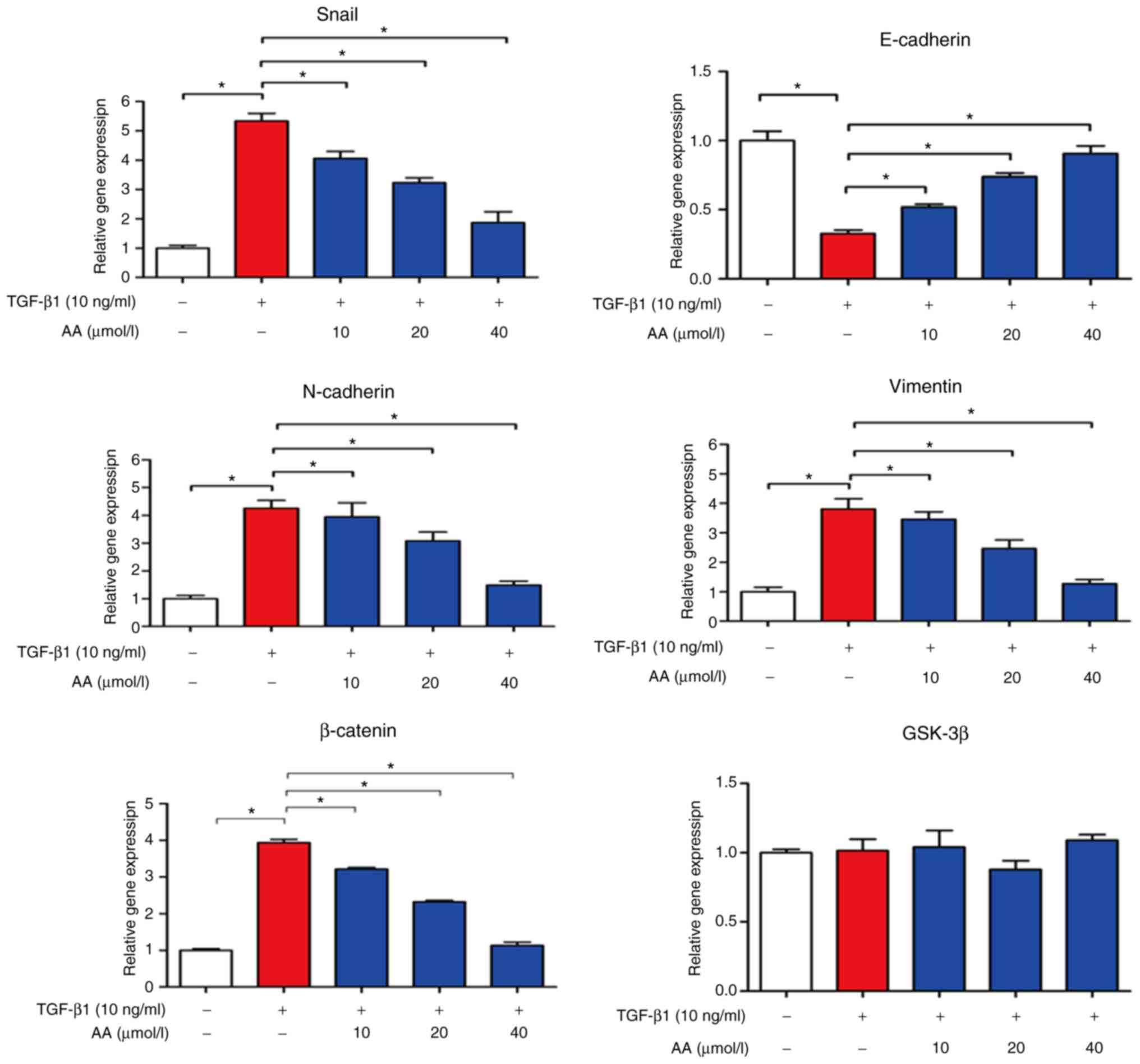
AA regulates EMT-associated mRNA
expression levels in A549 cells
The effects of AA on EMT-associated mRNA expression
levels in A549 cells were evaluated by RT-qPCR, as presented in
Fig. 5. In TGF-β1-treated cells,
E-cadherin expression levels were significantly decreased
(P<0.05), whereas N-cadherin (P<0.05), Snail (P<0.05),
vimentin and β-catenin expression levels were markedly increased
compared with in the control group (P<0.05). However, combined
cell treatment with TGF-β1 and AA significantly increased
E-cadherin expression levels (P<0.05), but decreased the
expression levels of N-cadherin, vimentin, Snail and β-catenin
compared with in TGF-β1-treated cells (P<0.05). No change in
GSK-3β expression level was observed in A549 cells treated with
TGF-β1 with or without AA. These results suggested that AA may
significantly inhibit EMT of A549 cells.
Discussion
Lung cancer is one of the most common types of
cancer worldwide and is one of the leading causes of
cancer-associated mortality. In China, the increasing incidence of
lung cancer and lung-cancer-associated mortality have become
serious health issues. At present, the 5-year survival rate remains
poor (~18%), since >57% of lung cancer cases are diagnosed at a
distant stage (2). Understanding the
biological characteristics of lung cancer and the molecular
mechanism underlying invasion and metastasis is therefore crucial.
Metastasis is the main biological process that distinguishes benign
from malignant tumors, and is an underlying cause of mortality in
patients with malignant tumors; however, the mechanism of tumor
metastasis is very complex and remains incompletely understood. EMT
has been reported as an important biological process of epithelial
cell-derived tumor cells that undergo migration and invasion
(24,25).
EMT involves the transformation of epithelial cells
into interstitial cells and serves crucial roles in embryonic
development, chronic inflammation, tissue reconstruction and cancer
metastasis (26). EMT is associated
with decreased expression of the cell adhesion molecule E-cadherin,
increased expression of the cytoskeletal component vimentin and by
enhanced characteristic morphology of mesenchymal cells (27–29).
TGF-β1 is an EMT-inducible factor that activates the TGF-β1
signaling pathway in epithelial cells and promotes EMT occurrence,
reduces tumor cell adhesion and enhances invasion in certain types
of cancer, including breast, lung, liver, colon and pancreatic
cancer (30–32). β-catenin is the main structural
component of intercellular adhesion, and abnormal alterations in
its expression are associated with tumor infiltration and
metastasis (33). The protein
p-GSK-3β is a multifunctional serine threonine kinase, which
regulates the activation of various signaling pathways and serves
important roles in protein expression, and tumor cell proliferation
and apoptosis. GSK-3β is considered to be the major negative
regulatory factor for β-catenin, and its inactivation can cause
cell cycle disturbance and accelerate tumor proliferation (34). In the present study, an EMT model of
lung cancer was established through the treatment of A549 cells
with TGF-β1. Following 24 h TGF-β1 treatment, A549 cells became
fusiform and presented interstitial cell features. In addition, the
treatment of A549 cells with TGF-β1 increased β-catenin and
p-GSK-3β protein expression levels. These results were consistent
with previous studies, which suggested that TGF-β1 may induce EMT
transformation of lung cancer cells (35,36).
EMT is a dynamic multi-step process that occurs
prior to tumor cell metastasis. These cells acquire the migratory
and invasive abilities of mesenchymal cells, which allow their
entry into the bloodstream and lymph channels for distant
metastasis (37). In the present
study, AA significantly inhibited invasion and migration of
TGF-β1-treated cells, which suggested that AA may prevent cell
metastasis by inhibiting EMT of A549 cells.
E-cadherin and N-cadherin belong to the cadherin
family; the extracellular domains of cadherin proteins share
immunoglobulin-like structure with the N-terminal domain.
E-cadherin forms an E-cadherin/β-catenin complex through connection
of β-catenin and cytoskeleton protein (38,39). The
E-cadherin/β-catenin complex interferes with cell adhesion, and
inhibits tumor cell invasion and metastasis (30). N-cadherin induces EMT by affecting
E-cadherin-associated cell adhesion, which gives cancer cells an
invasive and malignant phenotype (40). Vimentin is an important cytoskeletal
protein that is mainly present in mesenchymal tissues and cells. A
previous study reported that vimentin is closely associated with
tumor metastasis (41). Furthermore,
Snail is a transcriptional regulator and an upstream regulator of
E-cadherin, which negatively regulates E-cadherin expression. When
EMT occurs, Snail is upregulated and E-cadherin is downregulated,
which promotes the transition of epithelial cells to a mesenchymal
phenotype to promote tumor metastasis (42). In the present study, AA markedly
upregulated E-cadherin mRNA and protein expression levels; however,
Snail, N-cadherin, vimentin and β-catenin expression levels were
downregulated by AA in TGF-β1-treated cells. These results
suggested that AA may inhibit EMT of A549 cells.
In conclusion, the present study suggested that AA
may inhibit migration and invasion of A549 cells through EMT
inhibition. This finding provided novel foundations for the
treatment of lung cancer, and supplied theoretical and experimental
evidence for the use of AA as an EMT inhibitor.
Acknowledgements
Not applicable.
Funding
This study was supported by The Traditional Chinese
Medicine in Henan Province (grant no. 2017ZY2063).
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
QC and QZ designed the experiments. QC, JR, QY and
BL performed the experiments. QC, JR, QY and BL analyzed the data.
QC and JR wrote the manuscript. QC and JR revised the manuscript.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patients consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
AA
|
asiatic acid
|
|
EMT
|
epithelial-mesenchymal transition
|
|
TGF-β1
|
transforming growth factor-β1
|
References
|
1
|
Feng B, Zhang K, Wang R and Chen L:
Non-small-cell lung cancer and miRNAS: Novel biomarkers and
promising tools for treatment. Clin Sci (Lond). 128:619–634. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
World Health Organization. 2002,
https://www.who.int/bulletin/archives/80(1)78.pdfJune
18–2018
|
|
4
|
Smith CB, Kelley AS and Meier DE: Evidence
for new standard of care in non-small cell lung cancer patients.
Semin Thorac Cardiovasc Surg. 22:193–194. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Stuschke M, Eberhardt W, Pöttgen C,
Stamatis G, Wilke H, Stüben G, Stöblen F, Wilhelm HH, Menker H,
Teschler H, et al: Prophylactic cranial irradiation in locally
advanced non-small-cell lung cancer after multimodality treatment:
Long-term follow-up and investigations of late neuropsychologic
effects. J Clin Oncol. 17:2700–2709. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Minguet J, Smith KH and Bramlage P:
Targeted therapies for treatment of non-small cell lung
cancer-recent advances and future perspectives. Int J Cancer.
138:2549–2561. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xu JH, Yang HP, Zhou XD, Wang HJ, Liang G
and Tang CL: Role of Wnt inhibitory factor-1 in inhibition of
bisdemethoxycurcumin mediated epithelial-to-mesenchymal transition
in highly metastatic lung cancer 95D cells. Chin Med J (Engl).
128:1376–1383. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wu T, Geng J, Guo W, Gao J and Zhu X:
Asiatic acid inhibits lung cancer cell growth in vitro and in vivo
by destroying mitochondria. Acta Pharm Sin B. 7:65–72. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lu Y, Liu S, Wang Y, Wang D, Gao J and Zhu
L: Asiatic acid uncouples respiration in isolated mouse liver
mitochondria and induces HepG2 cells death. Eur J Pharmacol.
786:212–223. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ren L, Cao QX, Zhai FR, Yang SQ and Zhang
HX: Asiatic acid exerts anticancer potential in human ovarian
cancer cells via suppression of PI3K/Akt/mTOR signalling. Pharm
Biol. 54:2377–2382. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Park BC, Paek SH, Lee YS, Kim SJ, Lee ES,
Choi HG, Yong CS and Kim JA: Inhibitory effects of asiatic acid on
7,12-dimethylbenz[a]anthracene and 12-O-tetradecanoylphorbol
13-acetate-induced tumor promotion in mice. Biol Pharm Bull.
30:176–179. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tang XL, Yang XY, Jung HJ, Kim SY, Jung
SY, Choi DY, Park WC and Park H: Asiatic acid induces colon cancer
cell growth inhibition and apoptosis through mitochondrial death
cascade. Biol Pharm Bull. 32:1399–1405. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yilmaz M and Christofori G: EMT, the
cytoskeleton, and cancer cell invasion. Cancer Metastasis Rev.
28:15–33. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ke Y, Zhao W, Xiong J and Cao R: miR-149
inhibits non-small-cell lung cancer cells EMT by targeting FOXM1.
Biochem Res Int. 2013:5067312013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Roy BC, Kohno T, Iwakawa R, Moriguchi T,
Kiyono T, Morishita K, Sanchez-Cespedes M, Akiyama T and Yokota J:
Involvement of LKB1 in epithelial-mesenchymal transition (EMT) of
human lung cancer cells. Lung Cancer. 70:136–145. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shen SJ, Zhang YH, Gu XX, Jiang SJ and Xu
LJ: Yangfei Kongliu formula, a compound Chinese herbal medicine,
combined with cisplatin, inhibits growth of lung cancer cells
through transforming growth factor-β1 signaling pathway. J Integr
Med. 15:242–251. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Al-Saad S, Al-Shibli K, Donnem T, Persson
M, Bremnes RM and Busund LT: The prognostic impact of NF-kappaB
p105, vimentin, E-cadherin and Par6 expression in epithelial and
stromal compartment in non-small-cell lung cancer. Br J Cancer.
99:1476–1483. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang HJ, Wang HY, Zhang HT, Su JM, Zhu J,
Wang HB, Zhou WY, Zhang H, Zhao MC, Zhang L and Chen XF:
Transforming growth factor-β1 promotes lung adenocarcinoma invasion
and metastasis by epithelial-to-mesenchymal transition. Mol Cell
Biochem. 355:309–314. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zang QQ, Lu Z, Ning G and Cheng H:
Ophiopogonin D inhibits cell proliferation, causes cell cycle
arrest at G2/M, and induces apoptosis in human breast carcinoma
MCF-7 cells. J Integr Med. 14:51–59. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu JJ, Liu JY, Chen J, Wu YX, Yan P, Ji
CD, Wang YX, Xiang DF, Zhang X, Zhang P, et al: Scinderin promotes
the invasion and metastasis of gastric cancer cells and predicts
the outcome of patients. Cancer Lett. 376:110–117. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Uygun K, Bilici A, Kaya S, Oven
Ustaalioglu BB, Yildiz R, Temiz S, Seker M, Aksu G, Cabuk D and
Gumus M: XELIRI plus bevacizumab compared with FOLFIRI plus
bevacizumab as first-line setting in patients with metastatic
colorectal cancer: Experiences at two-institutions. Asian Pac J
Cancer Prev. 14:2283–2288. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Beekman JM, Reischl J, Henderson D, Bauer
D, Ternes R, Peña C, Lathia C and Heubach JF: Recovery of
microarray-quality RNA from frozen EDTA blood samples. J Pharmacol
Toxicol Methods. 59:44–49. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang J and Weinberg RA:
Epithelial-mesenchymal transition: At the crossroads of development
and tumor metastasis. Dev Cell. 14:818–829. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nagathihalli NS, Massion PP, Gonzalez AL,
Lu P and Datta PK: Smoking induces epithelial-to-mesenchymal
transition in Non-small cell lung cancer through HDAC-mediated
downregulation of E-cadherin. Mol Cancer Ther. 11:2362–2372. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Aref AR, Huang RYJ, Kang W, Jing SW,
Thiery JP and Kamm RD: Transitions between epithelial and
mesenchymal states in microfluidic platform. Acquisition of
malignant and stem cell traits. 2013.
|
|
27
|
Kiesslich T, Pichler M and Neureiter D:
Epigenetic control of epithelial-mesenchymal-transition in human
cancer. Mol Clin Oncol. 1:3–11. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhao R, Gong L, Li L, Guo L, Zhu D, Wu Z
and Zhou Q: nM23-H1 is a negative regulator of TGF-β1-dependent
induction of epithelial-mesenchymal transition. Exp Cell Res.
319:740–749. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Reinhold WC, Reimers MA, Lorenzi P, Ho J,
Shankavaram UT, Ziegler MS, Bussey KJ, Nishizuka S, Ikediobi O,
Pommier YG and Weinstein JN: Multifactorial regulation of
E-cadherin expression: An integrative study. Mol Cancer Ther.
9:1–16. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cho KH, Yu SL, Cho DY, Chang GP and Hoi
YL: Breast cancer metastasis suppressor 1 (BRMS1) attenuates
TGF-β1-induced breast cancer cell aggressiveness through
downregulating HIF-1α expression. BMC Cancer. 15:8292015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Toba-Ichihashi Y, Yamaoka T, Ohmori T and
Ohba M: Up-regulation of Syndecan-4 contributes to TGF-β1-induced
epithelial to mesenchymal transition in lung adenocarcinoma A549
cells. Biochem Biophys Rep. 5:1–7. 2015.PubMed/NCBI
|
|
32
|
Li Y, Zhu G, Zhai H, Jia J, Yang W, Li X
and Liu L: Simultaneous stimulation with tumor necrosis factor-α
and transforming growth factor-β1 induces epithelial-mesenchymal
transition in colon cancer cells via the NF-κB pathway. Oncol Lett.
15:6873–6880. 2018.PubMed/NCBI
|
|
33
|
Ding Y, Shen S, Lino AC, Curotto de
Lafaille MA and Lafaille JJ: Beta-catenin stabilization extends
regulatory T cell survival and induces anergy in nonregulatory T
cells. Nat Med. 14:162–169. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shakoori A, Ougolkov A, Yu ZW, Zhang B,
Modarressi MH, Billadeau DD, Mai M, Takahashi Y and Minamoto T:
Deregulated GSK3beta activity in colorectal cancer: Its association
with tumor cell survival and proliferation. Biochem Biophys Res
Commun. 334:1365–1373. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Singh A and Settleman J: EMT, cancer stem
cells and drug resistance: An emerging axis of evil in the war on
cancer. Oncogene. 29:4741–4751. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lin XL, Liu M, Liu Y, Hu H, Pan Y, Zou W,
Fan X and Hu X: Transforming growth factor β1 promotes migration
and invasion in HepG2 cells: Epithelial-to-mesenchymal transition
via JAK/STAT3 signaling. Int J Mol Med. 41:129–136. 2018.PubMed/NCBI
|
|
37
|
Talmadge JE and Fidler IJ: AACR centennial
series: The biology of cancer metastasis: Historical perspective.
Cancer Res. 70:5649–5669. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Macdonald PR, Progias P, Ciani B, Patel S,
Mayer U, Steinmetz MO and Kammerer RA: Structure of the
extracellular domain of Tie receptor tyrosine kinases and
localization of the angiopoietin-binding epitope. J Biol Chem.
281:28408–28414. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Abuetabh Y, Persad S, Nagamori S, Huggins
J, Al-Bahrani R and Sergi C: Expression of E-cadherin and β-catenin
in two cholangiocarcinoma cell lines (OZ and HuCCT1) with different
degree of invasiveness of the primary tumor. Ann Clin Lab Sci.
41:217–223. 2011.PubMed/NCBI
|
|
40
|
Chunhacha P, Sriuranpong V and
Chanvorachote P: Epithelial-mesenchymal transition mediates anoikis
resistance and enhances invasion in pleural effusion-derived human
lung cancer cells. Oncol Lett. 5:1043–1047. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang H, Liu J, Yue D, Gao L, Wang D,
Zhang H and Wang C: Clinical significance of E-cadherin, β-catenin,
vimentin and S100A4 expression in completely resected squamous cell
lung carcinoma. J Clin Pathol. 66:937–945. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Batlle E, Sancho E, Francí C, Domínguez D,
Monfar M, Baulida J and García De Herreros A: The transcription
factor snail is a repressor of E-cadherin gene expression in
epithelial tumour cells. Nat Cell Biol. 2:84–89. 2000. View Article : Google Scholar : PubMed/NCBI
|