Introduction
Cancer is one of the leading causes of human
mortality worldwide, with an estimated 14 million new cancer cases
projected for 2030 (1). In women,
breast cancer is quickly becoming the leading cause of mortality
worldwide, and novel therapeutic avenues are constantly being
explored (2). One such line of
investigation involves evaluating natural products extracted from
plants and endophytic fungi, such as vincristine and vinblastine,
which have been demonstrated to exhibit anticancer activities,
including the inhibition of breast cancer cell growth (3,4). Studies
are continuously being conducted in the search for novel effective
and nontoxic anticancer compounds from various medicinal
plants.
The genus Grangea belongs to the Compositae
(Asteraceae) family and comprises only six species, which are
mostly native and distributed throughout Africa, South Asia and
Southeast Asia (5,6). G. maderaspatana (L.) Poir.
(Phayaa Mutti) is one of the most common medicinal plants used in
traditional Thai medicine in various therapeutic approaches,
including ingestion of the whole plant to stimulate digestion,
reduce pain and inflammation, and regulate menses, while the leaf
is used for reducing spasms (7).
Although anesthetic, antioxidant, antibacterial and topoisomerase I
and II (Top I and II) inhibitory activities have been reported
previously for compounds derived from G. maderaspatana
(8–12), to the best of our knowledge, no
previous studies have assessed whether these compounds possess
anticancer activity.
A previous study in our laboratory revealed the
presence of sesquiterpene lactones (SLs) in G. maderaspatana
(12). SLs are compounds which have
several significant cancer-associated implications and are used in
targeted therapy against cancer cells, their stem cells and
specific signaling pathways (13).
Various SLs, including thapsigargin, artemisinin and parthenolide,
have been demonstrated to exhibit potent action against certain
types of cancer (13). SL extracts,
known as eudesmanolides, derived from frullanolide have been
demonstrated to possess anticancer activity in oral, non-small cell
lung and breast cancer cell lines (12). However, no scientific studies have
been conducted concerning frullanolide and its anticancer
activities or its cytotoxic mechanisms against breast cancer cells.
Therefore, in the present study, the cytotoxic effects of
frullanolide and its mode of action on breast cancer cell
inhibition were explored.
Materials and methods
Cell cultures
The breast cancer cell lines MCF-7 (HTB-22™),
MDA-MB-468 (HTB-132™) and MDA-MB-231 (HTB-26™), and a normal
epithelial breast cell line (MCF-12A; CRL-10782™) were purchased
from the American Type Culture Collection (ATCC, Manassas, VA,
USA), and maintained at the Department of Biomedical Sciences,
Faculty of Medicine, Prince of Songkla University (Hat Yai,
Thailand). The mouse fibroblast L-929 (CCL-1™; ATCC) cell line was
provided by Professor Teerapol Srichana, Department of
Pharmaceutical Technology, Faculty of Pharmaceutical Sciences,
Prince of Songkla University. The culture conditions of the cell
lines followed methods described previously (14). Briefly, MCF-7 cells were grown in
RPMI-1640 (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA), and the MDA-MB-468, MDA-MB-231 and L-929 cells were cultured
in Dulbecco's modified Eagle's medium (DMEM) (Gibco; Thermo Fisher
Scientific, Inc). The cultures were supplemented with 10% fetal
bovine serum (Gibco; Thermo Fisher Scientific, Inc.), with 100 U/ml
of penicillin and streptomycin. The MCF-12A cell line was cultured
in DMEM and Ham's F12 medium (GE Healthcare, Chicago, IL, USA),
supplemented with 100 ng/ml cholera toxin (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany), 500 ng/ml hydrocortisone (Sigma-Aldrich;
Merck KGaA), 0.01 mg/ml bovine insulin (Sigma-Aldrich; Merck KGaA),
20 ng/ml human epidermal growth factor (Invitrogen; Thermo Fisher
Scientific, Inc.) and 5% horse serum (Invitrogen; Thermo Fisher
Scientific, Inc.). All cultures were incubated at 37°C in a
humidified atmosphere containing 5% CO2.
Plant material and isolation
Dried G. maderaspatana (L.) Poir plants were
purchased from the Chaokromper Drug Store (Bangkok, Thailand).
Their identity was confirmed by Dr Nijsiri Ruangrungsri, Department
of Pharmacognosy and Pharmaceutical Botany, Faculty of
Pharmaceutical Sciences, Chulalongkorn University (Bangkok,
Thailand). A voucher specimen (no. 5182) was deposited at the
Museum of Natural Medicine, Chulalongkorn University. The dried
whole plant materials were ground to coarse powder using a mortar
and pestle. The powder was then stored at room temperature (RT)
prior to extraction. Crude compounds were extracted with
dichloromethane from a powder weight of 1,500 g. The extract was
evaporated under vacuum at 55°C, fractionated by silica gel column
chromatography (silica gel no. 9385) and then eluted with different
gradients of hexane-ethyl acetate 10:0 to 0:10 solvent systems,
resulting in 132.2 mg of a compound resembling white needles (Rf
0.85, silica gel CH2Cl2-Acetone 9:1). The
structure of the compound was elucidated by 1H,
13C nuclear magnetic resonance spectroscopy and mass
spectrometry. Its physical and spectral data were compared with
previous reports (9,12). The compound was identified as the
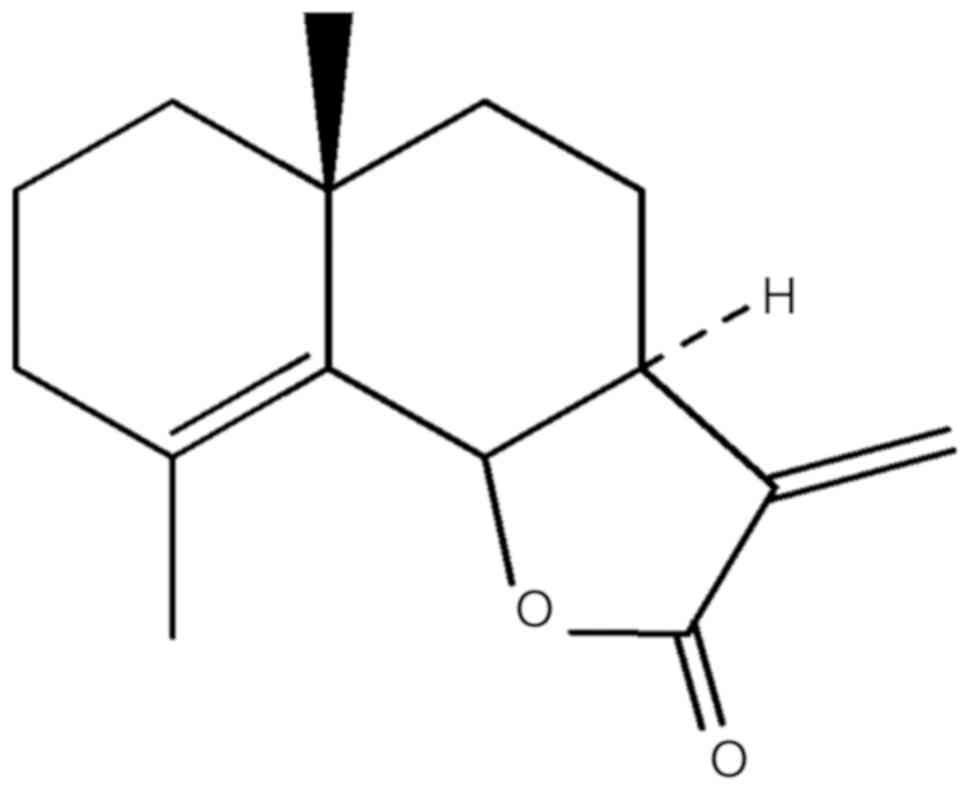
sesquiterpene lactone, frullanolide (Fig. 1), with the chemical formula
C15H20O2 (parent peak at m/z 232,
colorless solid). The compound was stored at −20°C prior to testing
with cancer cells.
MTT assay
All cell lines were seeded in 96-well plates at a
density of 2×104 cells/well in 100 µl culture
medium/well. The cells were treated with different concentrations
of the dimethyl sulfoxide (DMSO)-dissolved frullanolide compound
(1.25, 2.5, 5.0, 10.0 and 20.0 µg/ml). After 72 h of incubation at
37°C, 100 µl MTT reagent (0.5 mg/ml) was added to each well and the
cultures were incubated for an additional 30 min. The MTT reagent
was removed and replaced by DMSO to ensure that solubilization was
complete. Absorbance at 570 and 650 nm (reference wavelengths) was
measured on a microplate reader. Half-maximal inhibitory
concentration (IC50) values were calculated from fitted
response curves of the concentration and viability (%).
Determination of cytotoxic activity followed the criterion; <5
µg/ml represented highly active; 5–10 µg/ml represented strongly
active; and >10 represented weak cytotoxicity (15,16).
Normal breast cells (MCF-12A) and L-929 fibroblasts were treated
using the aforementioned procedure. Selectivity index (SI) values,
indicating selectivity for tested cell lines, were calculated from
the ratio of IC50 values of the compounds obtained for
normal vs. cancer cells. An SI score >3 represented good
selectivity (17). This experiment
was performed in triplicate.
Flow cytometry for cell cycle analysis
and apoptosis detection
The MCF-7 and MDA-MB-468 cells were seeded at
densities of 4×105 cells/well in 12-well plates. The
MDA-MB-231 cells were seeded at 3×105 cells/well. The
cells were treated for 24 h with three concentrations of
frullanolide (0.5×, 1× and 2× IC50). Following removal
of the compound, the treated cells were fixed with 70% ethanol at
4°C for 4 h and washed three times with cold PBS. Harvesting and
fluorochrome binding of the cells were carried out as described
previously (2). For cell cycle
analysis, the fixed cells were resuspended in 400 µl propidium
iodide (PI; 50 µg/ml) solution/1×106 cells and incubated
at RT for 5–10 min in the dark. A total of 5,000 cells were
analyzed for each condition with a fluorescence-activated cell
sorting (FACS) Calibur flow cytometer (BD Biosciences, San Jose,
CA, USA), and histograms of cell population ratios in each phase of
the cell cycle were acquired. For apoptosis detection, the
apoptotic cells in the treated conditions were stained with Annexin
V-fluorescein isothiocyanate (FITC)/PI following the manufacturer's
protocol (FITC Annexin V Apoptosis Detection kit I; BD Pharmingen™,
BD Biosciences). Dot plot graphs of the apoptotic cell ratios were
created. All data were analyzed using WinMDI v.2.9 software (J.
Trotter, The Scripps Institute, La Jolla, CA, USA).
Western blot analysis
Untreated cells and cells treated with frullanolide
at 0.5× IC50, were harvested at 0, 12, 24 and 48 h. The
cells were lysed in radioimmunoprecipitation assay buffer (Thermo
Fisher Scientific, Waltham, MA, USA). Protein samples were
quantitated by Bradford Assay (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). Total proteins (50 µg) of each sample were run
separately on a 12% SDS-PAGE gel and transferred onto a
nitrocellulose membrane in glycine-methanol buffer at 4°C (100 V, 2
h). The membranes were blocked in 5% low-fat milk in TBS with 0.1%
Tween-20 (TBS-T) at RT for 1 h. Subsequently, the blots were
incubated at 4°C overnight with primary antibodies; B cell lymphoma
2-associated X protein (Bax; cat no. 5023; 1:1,000 dilution), p21
(cat no. 2947; 1:1,000 dilution), p53 (cat no. 9282; 1:1,000
dilution) and β-actin (cat no. 4967; 1:1,000 dilution), in 1%
low-fat milk in TBS-T. All antibodies were purchased from Cell
Signaling Technology, Inc. (Danvers, MA, USA). The blots were then
washed 3 times (5 min each) and incubated with enhanced
chemiluminescence (ECL) anti-rabbit immunoglobulin G horseradish
peroxidase (cat no. PKNA934; GE Healthcare Life Sciences, Little
Chalfont, UK) diluted to 1:5,000 in 1% low-fat milk in TBS-T at RT
for 1 h. The protein bands were detected using an ECL
chemiluminescent detection kit (Thermo Fisher Scientific, Inc.).
Due to the limited amount of frullanolide compound available, each
treatment was performed in triplicate. The triplicate samples were
pooled prior to protein lysate preparation and western
blotting.
Statistical analysis
The MTT results are presented as the mean ± standard
deviation. To evaluate the difference between the cell lines,
one-way analysis of variance (ANOVA) with Brown-Forsythe correction
was performed, then IC50 values of the cancerous cell
lines were compared with MCF-12A cells using one-sided Dunnett's
post hoc tests. Statistical analysis was performed with SPSS 20.0
software (IBM Corp., Armonk, NY, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
Cytotoxic activity of
frullanolide
The IC50 values of frullanolide for the
breast cancer cell lines MCF-7, MDA-MB-468 and MDA-MB-231 were
10.74±0.86, 8.04±2.69 and 12.36±0.31 µg/ml, respectively. The
IC50 value for normal breast cells (MCF-12A) was
28.65±6.57 and 19.07±7.16 µg/ml for the fibroblast L-929 cell line.
One-way ANOVA indicated significantly different values among the
groups. It was tested whether the compound had lower
IC50 values in the cancerous cell lines than when
applied to MCF-12A cells. In cancerous cell lines, frullanolide had
significantly lower IC50 values compared with MCF-12A,
whereas the L-929 fibroblast cell line did not exhibit a
significant difference. The results indicated a strong
anti-proliferative effect of frullanolide on the breast cancer cell
lines, but less of an effect on normal breast and fibroblast cell
lines (Table I). The SI, which
indicates the safety level of a compound toward normal breast
cells, indicated that frullanolide has high cytotoxic activity
against MDA-MB-468 (SI=3.56) but was less harmful to normal breast
cells.
 | Table I.Cytotoxic activity (IC50)
of frullanolide purified from whole plant of G.
maderaspatana exhibits differing dosage dependence in MCF-7,
MDA-MB-468, MDA-MB-231, MCF-12A and L929 cell lines. |
Table I.
Cytotoxic activity (IC50)
of frullanolide purified from whole plant of G.
maderaspatana exhibits differing dosage dependence in MCF-7,
MDA-MB-468, MDA-MB-231, MCF-12A and L929 cell lines.
| Cell line |
IC50a, µg/ml (µM) | Cytotoxic
activity | SIb | SI activity |
|---|
| MCF-7 |
10.74±0.86c (46.23) | Weak
cytotoxicity | 2.67 | Less
selectivity |
| MDA-MB-468 |
8.04±2.69c (34.61) | Strongly
active | 3.56 | High
selectivity |
| MDA-MB-231 |
12.36±0.31c (53.20) | Weak
cytotoxicity | 2.32 | Less
selectivity |
| MCF-12A |
28.65±6.57 (123.32) | Weak
cytotoxicity | None | None |
| L929 | 19.07±7.16
(82.08) | Weak
cytotoxicity | None | None |
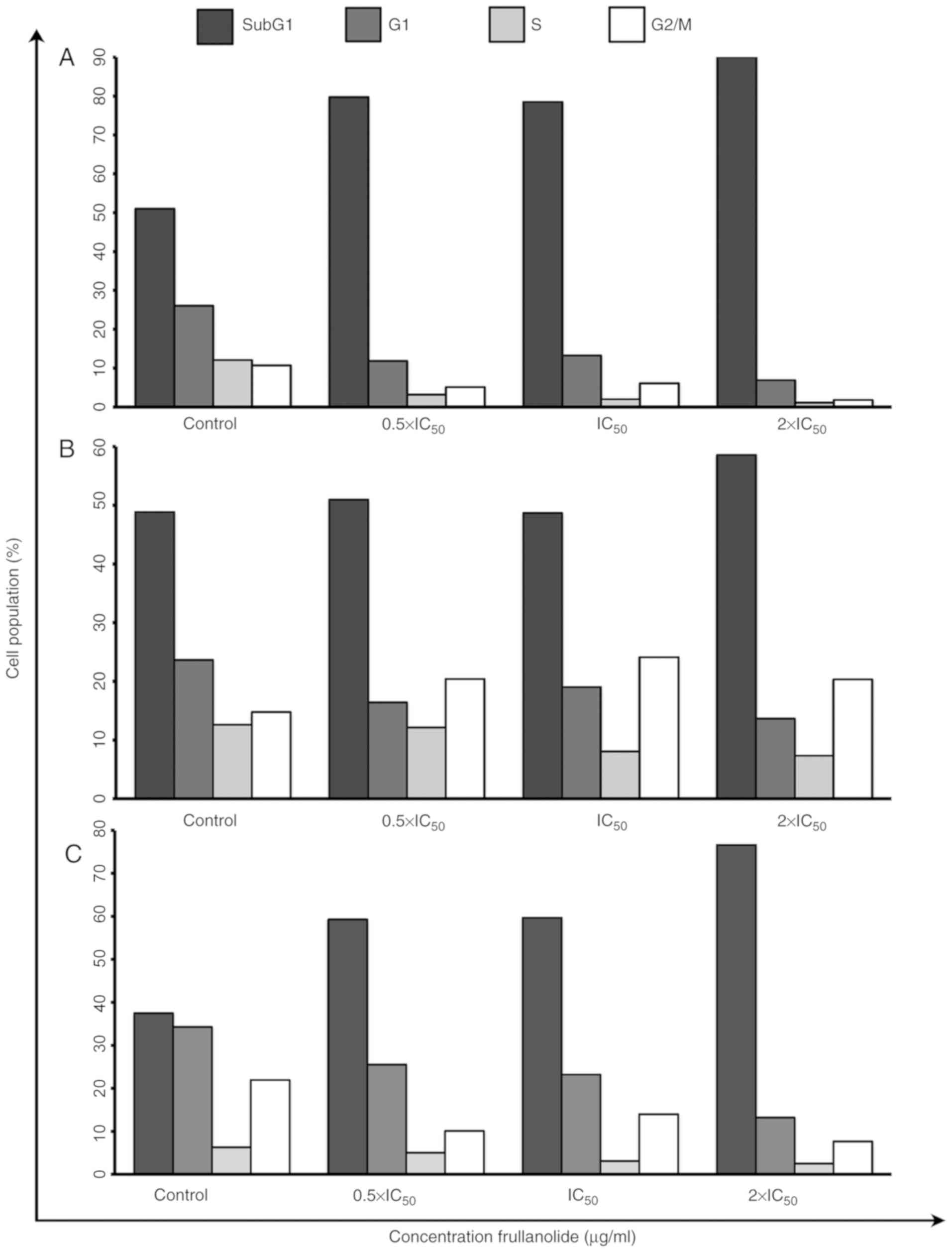
Cell cycle arrest of frullanolide
Cell cycle arrest was monitored using flow cytometry
(Figs. 2 and 3). Following staining of the nuclei with
PI, the FACS analysis revealed that frullanolide induced an
increased proportion of SubG1 cell debris for the MCF-7,
MDA-MB-468 and MDA-MB-231 cell lines in a dose-dependent manner.
The percentage of SubG1 cells in the MCF-7 cell line
increased from 79.77 at 0.5×IC50 to 90.10% at
2×IC50, compared with 51.05% in untreated cells
(Figs. 2A and 3A). There was also an increase in
SubG1 cells in MDA-MB-468, from 50.97 at
0.5×IC50 to 58.60% at 2×IC50 dosage (Figs. 2B and 3B). Additionally, the SubG1
proportion of the MDA-MB-231 cell line increased from 59.31 at
0.5×IC50 to 76.62% at 2×IC50 dose levels,
compared with 37.47% in untreated cells (Figs. 2C and 3C). The cell proportions in the
G1, S and G2/M phases in the histograms of
the MCF-7 treated cells tended to decrease in a dose-dependent
manner, from 11.87 to 6.91 (G1), 3.22 to 1.14 (S) and
5.14 to 1.85% (G2/M) (Figs.
2A and 3A). The distribution of
MDA-MB-468 cells among the phases also changed in a dose-dependent
manner (Fig. 2B). For MDA-MB-468,
the number of treated cells in the G1 and S phases
gradually decreased until the highest dosage (2×IC50).
Conversely, cells appeared to accumulate in the G2/M
phase in treated cells (20.43, 24.13 and 20.37% at 0.5×, 1× and 2×
IC50, respectively), compared with untreated cells
(14.81%; Fig. 3B). Notably, the
G1, S and G2/M phases of the MDA-MB-231 cell
cycles slightly decreased in a dose-dependent manner from 0.5× to
2×IC50 dose levels (G1, 25.53 to 13.19; S,
5.06 to 2.52; and G2/M, 10.10 to 7.68%) as shown in
Figs. 2C and 3C. These results indicated that
frullanolide was associated with cell cycle arrest at
G2/M in MDA-MB-468 but not in the MCF-7 and MDA-MB-231
cell lines.
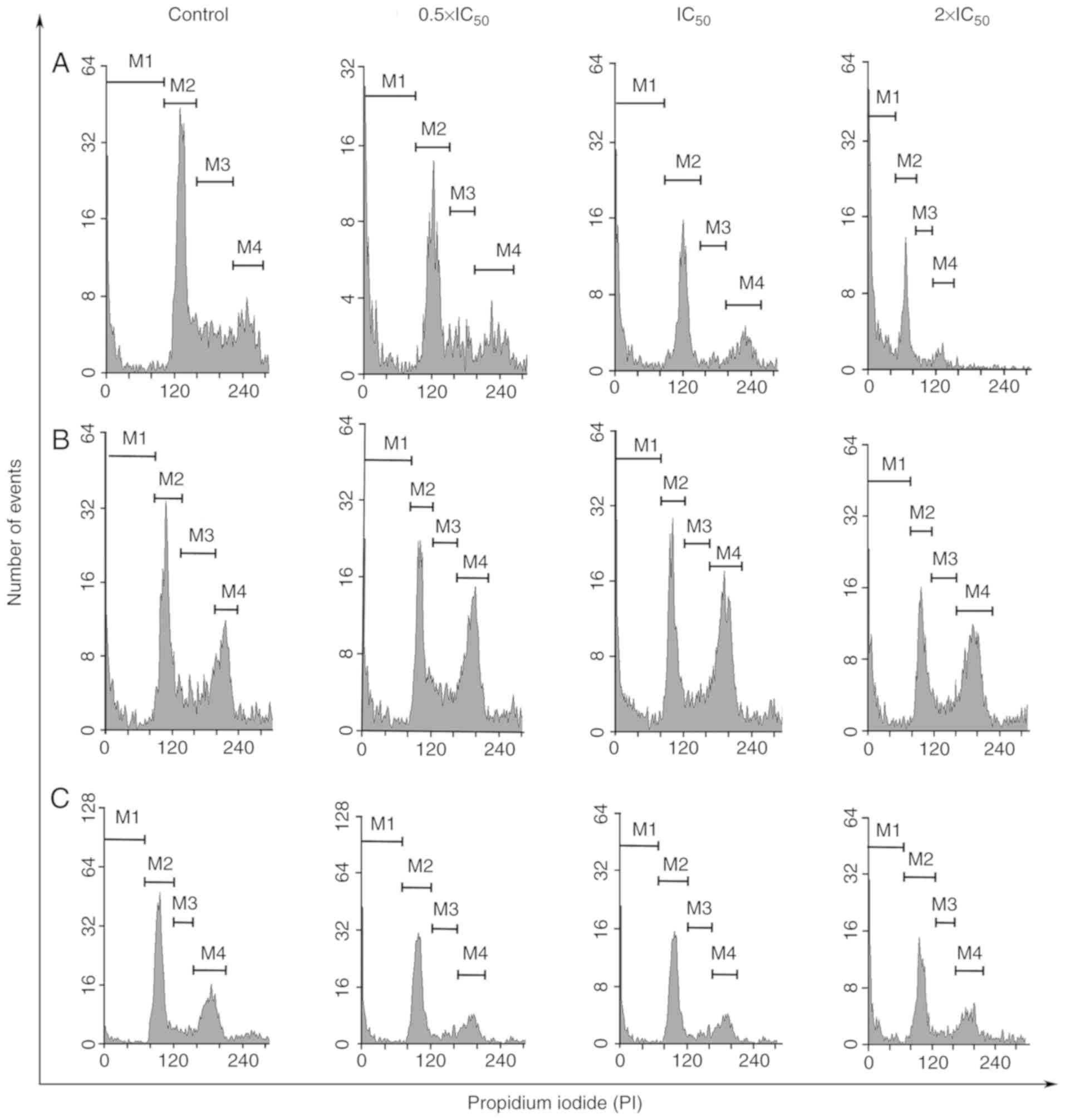 | Figure 2.Cell cycle distribution of breast
cancer cell lines following frullanolide treatments (0.5×, 1× and
2×IC50) using flow cytometry analysis. Histograms of
PI-labelled DNA content of each cell cycle phase in untreated and
treated (A) MCF-7, (B) MDA-MB-468 and (C) MDA-MB-231 cells,
respectively. M1, M2, M3, M4 in each histogram designate the number
of cells in the respective SubG1, G1, S and
G2/M phases. IC50, half-maximal inhibitory
concentration; PI, propidium iodide. |
Apoptosis induction of
frullanolide
To assess the apoptotic action of frullanolide,
Annexin V-FITC/PI was used to stain treated cells. A density plot
was created based on the data obtained from the FACS analysis. The
plot was divided into four quadrants (Fig. 4A-C), with the lower left quadrant
presenting viable cells (Annexin V-negative, PI-negative), the
lower right quadrant presenting cells that underwent early
apoptosis (Annexin V-positive, PI-negative), the upper right
quadrant presenting cells which underwent late apoptosis (Annexin
V-positive, PI-positive), and the upper left presenting necrotic
cells (Annexin V-negative, PI-positive). After 24 h of treatment of
MCF-7 cells, the lowest viable counts along with the highest
apoptotic counts (early and late apoptosis) were identified at the
0.5×IC50 dose level (14.97 and 84.38%, respectively;
Fig. 4A and D). These results
indicated that frullanolide may be effective in MCF-7 cells at
different dose levels, particularly at a low dose level
(0.5×IC50). Conversely, frullanolide treatment of
MDA-MB-468 cells resulted in the lowest number of viable cells
(11.96%) and the highest number of apoptotic cells (86.38%) at the
highest dosage (2×IC50; Fig.
4B and E). Although no significant differences were observed
for the MDA-MB-231 cells regarding the number of viable and
apoptotic cells among all treatment conditions, treatment exhibited
effective cytotoxic activity compared with the control (Fig. 4C and F). Therefore, the data
indicated that frullanolide may induce apoptosis in breast cancer
cells at all tested concentrations.
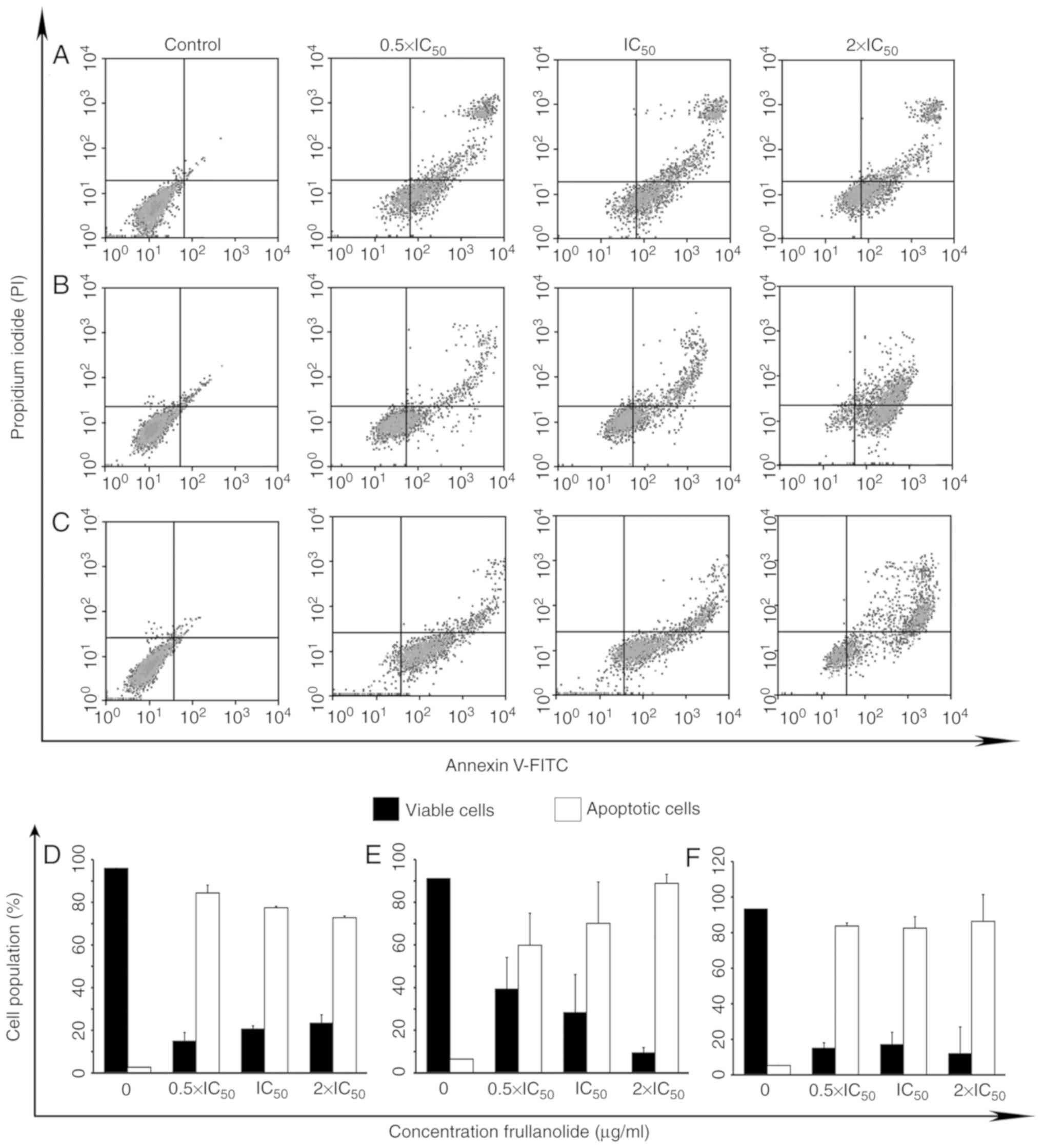 | Figure 4.Density plots of frullanolide-induced
apoptosis. (A) MCF-7, (B) MDA-MB-468 and (C) MDA-MB-231 cells
treated with frullanolide at 0.5×, 1× and 2×IC50. The
density plots show four quadrants; the lower left, lower right,
upper right and upper left quadrants represent viable, early
apoptotic, late apoptotic and necrotic cells, respectively. The
percentages of cell populations indicate viable cells (black
column) and total apoptotic cells (white column) for the (D) MCF-7,
(E) MDA-MB-468 and (F) MDA-MB-231 cell lines. Error bars represent
the data obtained from treated breast cancer cells in two
independent experiments. FITC, fluorescein isothiocyanate;
IC50, half-maximal inhibitory concentration; PI,
propidium iodide. |
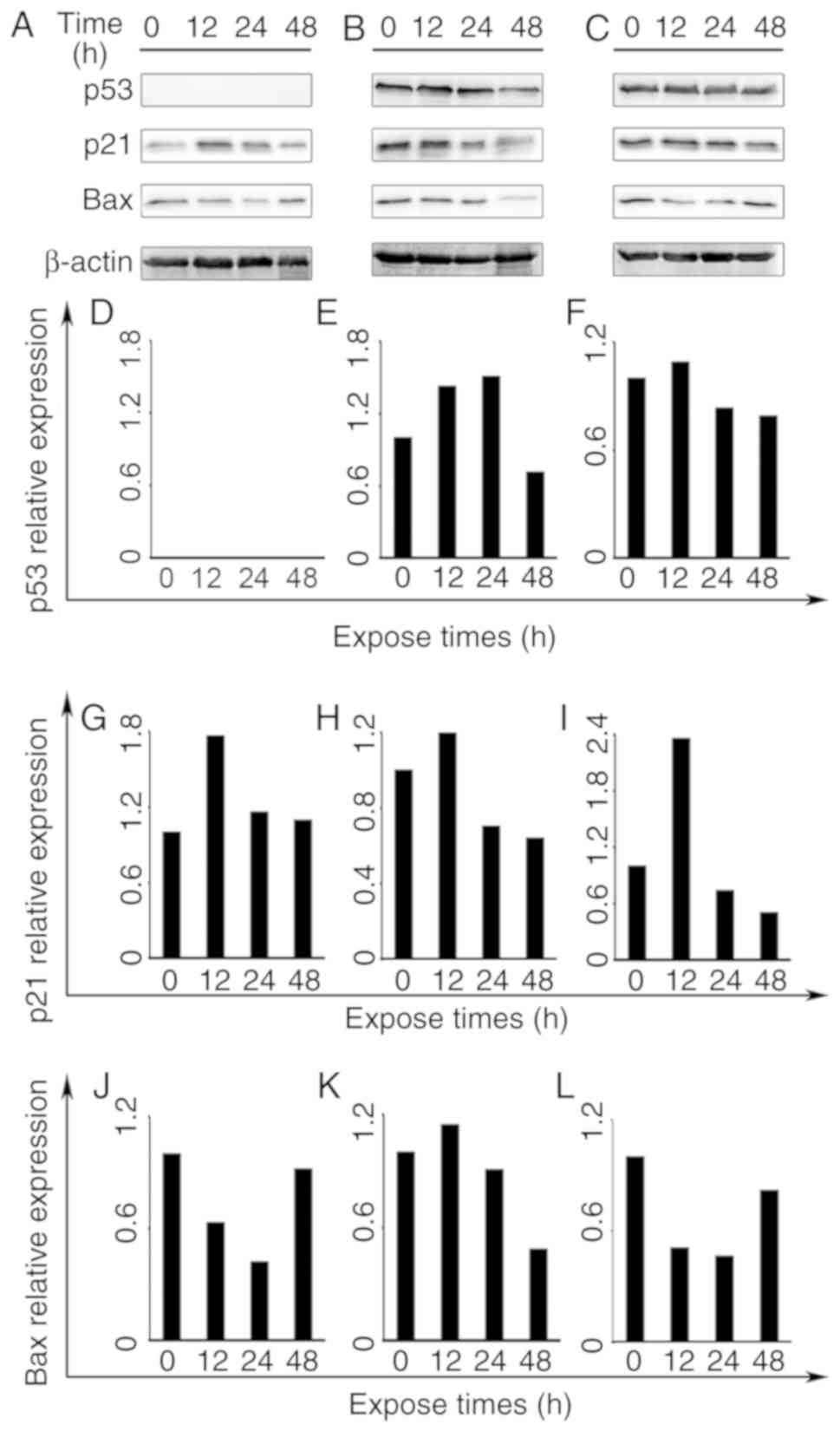
Protein expression by western blot
analysis
To elucidate the possible mechanisms of apoptotic
induction by frullanolide, the expression levels of three proteins
(Bax, p21 and p53) were investigated by western blot analysis at
0.5×IC50, for 12, 24 and 48 h in the three breast cancer
cell lines. The overall effect of the frullanolide treatment in
these breast cancer cell lines on detected protein expression is
presented in Fig. 5. For all cell
types treated with 0.5×IC50 frullanolide, Bax protein
expression decreased between 12 and 24 h. Its expression was prone
to accumulation in treated MCF-7 and MDA-MB-231 cells at 48 h
(Fig. 5A, C, J and L). However,
expression levels of Bax protein increased at 12 h in treated
MDA-MB-468 cells and its expression gradually reduced at 24–48 h
(Fig. 5K). Elevated p21 expression
was noted at 12 h, and gradually decreased between 24 and 48 h in
all three breast cancer cell lines. The p21 expression was
upregulated when compared with the controls (Fig. 5G-I). Notably, p53 protein was
expressed differentially depending on the cell type. p53 expression
was undetectable in MCF-7 cells (Fig. 5A
and D), while distinct adverse alterations were identified in
the treated MDA-MB-468 and MDA-MB-231 cells (Fig. 5E and F). The p53 protein levels in
frullanolide-treated MDA-MB-468 cells increased at 12–24 h, and
immediately declined by ~0.80-fold until the end of exposure (24–48
h) when compared with the control group (Fig. 5E). Therefore, it appeared conclusive
that the frullanolide mechanism was involved in apoptotic
pathways.
Discussion
SLs, derived from a natural product and a subfamily
of terpenoids, have been reported to exhibit numerous potential
medicinal properties, including anti-inflammatory, antimicrobial
and anticancer actions, in in vitro and in vivo
studies, as well as clinical trials (13,18–20).
More than 5,000 SLs have been identified from Asteraceae
spp. (18), and ~1,500
publications have reported on their anti-inflammatory and
anticancer properties (13). In
vitro cytotoxicity screening for antitumor agents has been
widely employed in various types of cancer cell lines in the search
for novel anticancer drugs. A number of potentially active
compounds have been identified through a large-scale screening
program based on the US National Cancer Institute criteria
(21). Purified and crude compounds
of one of these anticancer agents were reported to possess strong
antitumor activity, with IC50 values of ≤4 and ≤20
µg/ml, respectively (21). Several
studies have identified specific SLs and derivatives which exhibit
broad-spectrum antitumor activity towards several cancer types,
including non-small cell lung cancer, colorectal cancer, leukemia,
laryngeal cancer, gynecological cancer and breast cancer (13,22–27). The
different anticancer abilities of SLs are associated with their
carbocyclic skeleton classification (13,28).
Eudesmanolides (6/6-bicyclic compounds) a subgroup of SLs, have
been reported in several studies to exhibit anticancer action
(29–34). For example, the eudesmane skeleton
(santamarine) has been revealed to exhibit potential anticancer
properties toward the L1210 murine (IC50=0.41 µg/ml),
CCRF-CEM human leukemia (IC50=0.59 µg/ml), KB human
nasopharyngeal (IC50=0.16 µg/ml), LS174T human colon
carcinoma (IC50=0.92 µg/ml) and MCF-7 breast
adenocarcinoma (IC50=0.53 µg/ml) cell lines (29). Li et al (30), studied the efficacy of
eudesmane-based santamarine against a number of gynecological
cancer cell lines, and revealed that HeLa ovarian cancer and SHIN3
cervical cancer cell line viabilities are decreased by 50%
following respective treatments of 2.60 and 3.08 µg/ml santamarine
for 48 h, while the HOC-21 and HAC-2 ovarian cancer, and HEC-1
endometrial cancer cell lines had some tolerance
(IC50>10 µg/ml) (31).
Similarly, the cytotoxic activity of a eudesmanolide compound
[15-hydroxy-eudesm-4,11(13)-diene-12-oic acid] is potent toward a
panel of human cancer cell lines (PC prostate cancer, HT29 colon
cancer, MCF-7 breast cancer and A549 lung cancer) with
IC50 of 5.8±0.2, 5.8±0.2, 6.8±0.4 and 69.6±7.1 µM,
respectively (31). Five compounds
isolated from the flowers of Tanacetum vulgare exhibit
degrees of anticancer activity between 15.3 and 60.0 µM against
A549 lung cancer cells (32).
Notably, a novel 12,8-eudesmanolide extract from Eutypella
spp. exhibits cytotoxicity against lymphoma, hepatocarcinoma
and myeloid leukemia cell lines (33–34).
These and other studies indicate that eudesmanolide-based SLs are
potentially strong anticancer compounds for the treatment of
various types of cancer, including breast cancer.
In the present study, frullanolide (a eudesmanolide)
was assessed by MTT assay, and exhibited potent anti-breast cancer
activity in breast cancer cell lines, including MCF-7, MDA-MB-468
and MDA-MB-231. The IC50 values were measured after 72
h. In comparison, the cytotoxicity of frullanolide was strongest
for MDA-MB-468, a triple negative breast cancer (TNBC) cell line
which lacks estrogen receptor, progesterone receptor and human
epidermal growth factor receptor 2 (35). This TNBC cell line is an important
model for breast cancer responsiveness due to the limited choices
of chemotherapeutic treatment for this disease (35–37).
Notably, in the present study, frullanolide exhibited potent
activity against MDA-MB-468 cells, which possess a unique targeted
receptor, the epidermal growth factor receptor (EGFR), on the cell
membrane (35). This receptor may be
involved in the specific interactions with frullanolide, leading to
its toxicity in these cells. Using in silico screening,
Sawatdichaikul et al (38)
reported a list of plant compounds that bind to EGFR. Two promising
compounds from the medicinal plants list are exiguaflavanone A and
exiguaflavanone B, which are compounds purified from Artemisia
indica Willd and Sophora exigua, respectively. These
belong to the Asteraceae family, the same family as G.
maderaspatana (L.) Poir. The structure of these two compounds
has approximately the same carbon skeleton orientation as
frullanolide (38). However, further
experiments are required to confirm any interactions between
frullanolide and EGFR. Additionally, Maldonado et al
(27) reported on two novel
eudesmanolide structures (C17H25O5
and C17H22O4) extracted from the
small genus Kaunia (Asteraceae family) which exhibited
strong anti-breast cancer activity toward five breast cancer cell
lines; HCC1937 (TNBC), JIMT-1, L56Br-C1, MCF-7 and SK-BR-3. The
IC50 values of eudesmanolide
C17H25O5 and
C17H22O4 ranged between 9.3 and
27.0 µM (L56Br-C1 >SK-BR-3 >JIMT-1>HCC1937>MCF-7) and
3.2–11.0 µM (HCC1937>SK-BR-3>JIMT-1>L56Br-C1>MCF-7)
(27). A previous computational
structure-based screening method to identify natural compounds that
specifically target the mouse double minute 2 homolog protein
revealed that, out of the 35 top candidates, 8 eudesmanolide SLs
(IJ-1, IJ-3, IJ-5, IJ-6, IJ-9, IJ-11, IH-45 and IH-49) exerted more
potent anti-TNBC activity (MDA-MB-231) than anti-non-TNBC (MCF-7)
activity at 72 h (39). The
IC50 values ranged between 10 and 50 µM (39). In the present study, the anti-breast
cancer activity of frullanolide in the MDA-MB-468, MCF-7 and
MDA-MB-231 cell lines exhibited similar IC50 levels of
34.61, 46.23 and 53.20 µM, respectively. In comparison, the
anti-breast cancer activity of frullanolide in certain cell lines
(specifically TNBC) exhibited moderately strong activity, but
exhibited high selectivity for breast cancer cells (less harmful to
normal breast cells).
To evaluate the mechanism of apoptosis following
frullanolide treatment, flow cytometry analysis was selected for
measurement of quantitative DNA content (PI staining) and apoptotic
cells (double-staining fluorescent PI and annexin V-FITC).
Apoptosis-associated protein (p53, p21 and Bax) expression levels
were analyzed by western blotting. Notably, apoptotic induction by
the appropriate frullanolide treatment occurred in all three breast
cancer cell lines, but G2/M arrest only occurred in
frullanolide-treated MDA-MB-468 cells. Niculescu et al
(40), reported that the level of
p21 accumulation serves a role in negatively regulating the
G2/M transition. The same study revealed that the
expression of p21 is associated with endoreduplication in
pRb-negative cells by inhibiting cyclin-dependent kinases leading
to G2 arrest (40).
The findings of the present study supported the
observation of cell cycle arrest in MDA-MB-468, in that an increase
of p21 expression after 12 h of treatment was observed. The
expression of p21 also induces the apoptotic pathway in
p53-dependent and -independent pathways (41). Overall, after 12 h of treatment,
apoptosis was induced by increasing the expression of p21 and Bax
at various time points in the MDA-MB-468, MDA-MB-231 and MCF-7 cell
lines.
In the present study, p53 expression in treated
MCF-7 cells was not detectable due to its short half-life, caused
by proteasomal degradation (42,43).
This effect may be regulated by p53-independent p21 activation
molecules, including transforming growth factor β, tumor necrosis
factor α, histone deacetylase inhibitors, interferon γ and
interleukin 6 (41).
In conclusion, the results of the present study
suggested that the apoptotic pathway was involved in
frullanolide-induced cell death via p21 induction and
p53-independent pathways in the MCF-7 cell line and p53-dependent
pathways in the MDA-MB-468 and MDA-MB-231 breast cancer cell lines.
Further experiments are required to clarify the molecular mechanism
involved in the anti-breast cancer activity of frullanolide.
Acknowledgements
The authors would like to thank Mr. David Patterson
(International Affairs Office, Faculty of Medicine, Prince of
Songkla University, Hat Yai, Thailand) for English
proofreading.
Funding
The present study was financially supported by
grants from the Faculty of Medicine, Prince of Songkla University
(grant no. REC57-0162-04-2) and Prince of Songkla University
Funding (grant no. MED560604S).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
SC performed the experiments, acquired and analyzed
the data, and was a major contributor in the manuscript writing. PG
was responsible for the cytotoxic activity assay and interpreted
the data. TS provided the protocol and guidance for FACS analysis.
SS extracted and purified the frullanolide compound. RB performed
the western blotting. KK designed all the experiments, analyzed the
data and was a major contributor in editing the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Stewart BW and Wild CP: World cancer
Report 2014 Lyon. International Agency for Research on Cancer
(IARC). pp16–53. 2014.
|
|
2
|
Srisawat T, Sukpondma Y, Chimplee S,
Kanokwiroon K, Tedasen A and Graidist P: Extracts from vatica
diospyroides type SS fruit show low dose activity against
MDA-MB-468 breast cancer cell line via apoptotic action. Biomed Res
Int. 2014:4796022014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Noble RL: The discovery of the vinca
alkaloids-chemotherapeutic agents against cancer. Biochem Cell Bio.
68:1344–1351. 1990. View
Article : Google Scholar
|
|
4
|
Kumar A, Patil D, Rajamohanan PR and Ahmad
A: Isolation, purification and characterization of vinblastine and
vincristine from endophytic fungus Fusarium oxysporum
isolated from Catharanthus roseus. PLoS One. 8:e718052013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Patel V, Shukla S and Patel S: Free
radical scavenging activity of Grangea maderaspatana Poir.
Pharmacogn Mag. 5:381–387. 2009. View Article : Google Scholar
|
|
6
|
Rao VM, Damu GLV, Sudhakar D and Rao CV:
Two new bio-active flavones from Grangea maderaspatana
(Artemisia maderaspatana). Asian J Chem. 21:1552–1558.
2009.
|
|
7
|
Chaturvedi D: Sesquiterpene lactones:
Structural diversity and their biological activities. In: Tiwari VK
and Mishra BB (eds.): Opportunity, Challenge and Scope of Natural
Products in Medicinal Chemistry. Research Signpost. (Kerala,
India). 313–334. 2011.
|
|
8
|
Ahmed M, Islam MM, Hossain CF and Khan OF:
A preliminary study on the analgesic activity of Grangea
maderaspatana. Fitoterapia. 72:553–554. 2011. View Article : Google Scholar
|
|
9
|
Ruangrungsi N, Kasiwong S and
Likhitwitayawuid K: Constituents of Grangea maderaspatana a
new Eudesmanolide. J Nat Prod. 52:130–134. 1989. View Article : Google Scholar
|
|
10
|
Sangmalee S, Laorpaksa A and Sukrong S: A
topoisomerase II poison screen of ethnomedicinal Thai plants using
a yeast cell-based assay. J Ethnopharmacol. 142:432–437. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Singh D, Mathela CS, Pande V and Panwar A:
Antioxidant and antimicrobial activity of Grangea
maderaspatana (L.) Poir. Extract. JDDT. 1:46–52. 2013.
|
|
12
|
Uppatanpreecha P: Topoisomerase I
inhibitory activity from Thai medicinal plants in yeast cell-based
assay. Dissertation. (Bangkok, Chulalongkorn University). 2009.
|
|
13
|
Ghantous A, Gali-Muhtasib H, Vuorela H,
Saliba NA and Darwiche N: What made sesquiterpene lactones reach
cancer clinical trials? Drug Discov Today. 15:668–678. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Srisawat T, Chumkaew P, Heed-Chim W,
Sukpondma Y and Kanokwiroon K: Phytochemical screening and
cytotoxicity of crude extracts of Vatica diospyroides
Symington type LS. Trop J Pharm Res. 12:71–76. 2013.
|
|
15
|
Alitheen NB, Mashitoh AR, Yeap SK,
Shuhaimi M, Manaf AA and Nordin L: Cytotoxic effect of
damnacanthal, nordamnacanthal, zerumbone and betulinic acid
isolated from Malaysian plant sources. Int Food Res J. 17:711–719.
2010.
|
|
16
|
Wibowo A, Ahmat N, Hamzah AS, Sufian AS,
Ismail NH, Ahmad R, Jaafar FM and Takayama H: Malaysianol A, a new
trimer resveratrol oligomer from the stem bark of Dryobalanops
aromatica. Fitoterapia. 82:676–681. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bézivin C, Tomasi S, Lohézic-Le
Dévéhat F and Boustie J: Cytotoxic activity of some lichen
extracts on murine and human cancer cell lines. Phytomedicine.
10:499–503. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chadwick M, Trewin H, Gawthrop F and
Wagstaff CF: Sesquiterpenoid lactones: Benefits to plants and
people. Int J Mol Sci. 14:12780–12805. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kreuger MR, Grootjans S, Biavatti MW,
Vandenabeele P and D'Herde K: Sesquiterpene lactones as drugs with
multiple targets in cancer treatment: Focus on parthenolide.
Anticancer Drugs. 23:883–896. 2012.PubMed/NCBI
|
|
20
|
Matejić J, Šarac Z and Ranđelović V:
Pharmacological activity of sesquiterpene lactones. Biotechnol
Biotechnol Equip. 24 (Suppl 1):S95–S100. 2010. View Article : Google Scholar
|
|
21
|
Cordell GA, Kinghorn D and Pezzuto JM:
Separation, structure elucidation, and bioassay of cytotoxic
natural products. Colegate SM and Molyneux RJ: Bioactive natural
products, Boca raton, CRC press. 195–216. 1993.
|
|
22
|
Wu C, Chen F, Rushing JW, Wang X, Kim HJ,
Huang G, Haley-Zitlin V and He G: Antiproliferative activities of
parthenolide and golden feverfew extract against three human cancer
cell lines. J Med Food. 9:55–61. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
van Haaften C, Duke CC, Weerheim AM, Smit
NP, van Haard PM, Darroudi F and Trimbos BJ: Potent cytotoxic
effects of Calomeria amaranthoides on ovarian cancers. J Exp
Clin Cancer Res. 30:1–6. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fischedick JT, Pesic M, Podolski-Renic A,
Bankovicc J, de Vosd RCH, Perić M, Todorovićg S and Tanicc N:
Cytotoxic activity of sesquiterpene lactones from Inula
britannica on human cancer cell lines. Phytochem Lett.
6:246–252. 2012. View Article : Google Scholar
|
|
25
|
Kabeer FA, Sreedevi GB, Nair MS,
Rajalekshmi DS, Gopalakrishnan LP, Kunjuraman S and Prathapan R:
Antineoplastic effects of deoxyelephantopin, a sesquiterpene
lactone from Elephantopus scaber, on lung adenocarcinoma
(A549) cells. J Integr Med. 11:269–277. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Costantino VV, Mansilla SF, Speroni J,
Amaya C, Cuello-Carrión D, Ciocca DR, Priestap HA, Barbieri MA,
Gottifredi V and Lopez LA: The sesquiterpene lactone
dehydroleucodine triggers senescence and apoptosis in association
with accumulation of DNA damage markers. PLoS One. 8:e531682013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Maldonado EM, Svensson D, Oredsson SM and
Sterner O: Cytotoxic sesquiterpene lactones from Kauna
lasiophthalma Griseb. Sci Pharm. 82:147–160. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Babaeia G, Aliarab A, Abroon S, Rasmi Y
and Gholizadeh-Ghaleh Aziz S: Application of sesquiterpene lactone:
A new promising way for cancer therapy based on anticancer
activity. Biomed Pharmacother. 106:239–246. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ma G, Chong L, Li Z, Cheung AH and
Tattersall MH: Anticancer activities of sesquiterpene lactones from
Cyathocline purpurea in vitro. Cancer Chemother Pharmacol.
64:143-152. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li Y, Ni ZY, Zhuc MC, Dong M, Wang SW, Shi
QW, Zhang ML, Wang YF, Huo CH, Kiyota H and Cong B: Antitumour
activities of sesquiterpene lactones from Inula helenium and
Inula japonica. Z Naturforsch C. 67:375–380. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mahmoud AA, AlFredan MA and El-Sayed WM:
Isolation, characterization and anticancer activity of seven
compounds from the aerial parts of Conyza triloba. Int J
Pharmacognosy and Phytochem Res. 8:2071–2079. 2016.
|
|
32
|
Rosselli S, Bruno M, Raimondo FM, Spadaro
V, Varol M, Koparal AT and Maggio A: Cytotoxic effect of
eudesmanolides isolated from flowers of Tanacetum vulgare ssp.
Siculum. Molecules. 17:8186–8195. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Rivero A, Quintana J, Eiroa JL, Lo´pez M,
Triana J, Bermejo J and Este´vez F: Potent induction of apoptosis
by germacranolide sesquiterpene lactones on human myeloid leukemia
cells. Eur J Pharmacol. 482:77–84. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang Y, Wang Y, Wu AA, Zhang L, Hu Z,
Huang H, Xu Q and Deng X: New 12,8-eudesmanolides from Eutypella
sp. 1–15. J Antibiot (Tokyo). 70:1029–1032. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Holliday DL and Speirs V: Choosing the
right cell line for breast cancer research. Breast Cancer Res.
13:2152011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hudisa CA and Gianni L: Triple-negative
breast cancer: An unmet medical need. Oncologist. 16 (Suppl
1):S1–S11. 2011. View Article : Google Scholar
|
|
37
|
Kandula M, Ch KK and Ys AR: Molecular
mechanism and targeted therapy options of triple-negative (ER, PgR,
HER−2/neu) breast cancer: Review. World J Oncol.
4:137–141. 2013.PubMed/NCBI
|
|
38
|
Sawatdichaikul O, Hannongbua S, Sangma C,
Wolschann P and Choowongkomon K: In silico screening of
epidermal growth factor receptor (EGFR) in the tyrosine kinase
domain through a medicinal plant compound database. J Mol Model.
18:1241–1254. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Qin JJ, Wang W, Voruganti S, Wang H, Zhang
WD and Zhang R: Identification of a new class of natural product
MDM2 inhibitor: In vitro and in vivo anti-breast
cancer activities and target validation. Oncotarget. 6:2623–2640.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Niculescu III AB, Chen X, Smeets M, Hengst
L, Prives C and Reed SI: Effects of p21Cip1/Waf1 at both
the G1/S and the G2/M cell cycle transitions:
pRb ss a critical determinant in blocking DNA replication and in
preventing endoreduplication. Mol Cell Biol. 18:629–643. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Gartel ALand Tyner AL: The role of the
cyclin-dependent kinase inhibitor p21 in apoptosis. Mol Cancer
Ther. 8:639–649. 2002.
|
|
42
|
Moll UM and Petrenko O: The MDM2-p53
interaction. Mol Cancer Res. 1:1001–1008. 2003.PubMed/NCBI
|
|
43
|
Tsvetkov P, Reuven N and Shaul Y:
Ubiquitin-independent p53 proteasomal degradation. Cell Death
Differ. 17:103–108. 2010. View Article : Google Scholar : PubMed/NCBI
|