Introduction
A hypoxic microenvironment is a common
characteristic of solid tumors. It has been reported that hypoxia
can induce epithelial-to-mesenchymal transition (EMT) via
hypoxia-induced factor-1α (HIF-1α) in various types of cancer,
including gastric cancer, hepatoblastoma, pancreatic carcinoma,
colon carcinoma and mammary cancer (1–3). EMT is
an important process in cancer metastasis; during EMT, epithelial
cells lose their cell polarity and cell-cell adhesion, and gain
migratory and invasive properties to become mesenchymal stem cells.
To complete the metastatic process, cancer cells must migrate out
of the primary tumor. Studies in mammary cancer have shown that the
EMT phenomenon increases the migration and invasion of cancer
cells, and is closely associated with tumor occurrence,
infiltration and distant implantation (4,5). Cancer
cells must detach from the neighboring epithelial cells by reducing
E-cadherin expression to start invading the surrounding
extracellular matrix. It has been reported that HIF-1α expression
is rarely observed in normal tissues but is increased in various
cancer tissues or cells, and is closely associated with the
migration and invasion of tumor cells. Therefore, inhibition of
HIF-1α activation may reduce the development and progression of
tumors (6–8).
HIF-1α can directly or indirectly regulate EMT
regulators, including TWIST, Snail, carbonic anhydrase IX (CAIX),
glucose transporter protein-1 (GLUT-1), and other transcription
factors (9–11). These transcription factors then
transactivate EMT-related genes, including vimentin, E-cadherin and
N-cadherin, to regulate progression of the EMT (12,13).
HIF-1α levels in mammary cancer tissues are associated with
pathological stages, and high HIF-1α levels may result in high
multiplication rates and the formation of more aggressive tumors
(14).
The metastasis of triple-negative breast cancer
(TNBC) is associated with poor prognosis and high mortality due to
ineffective treatment. Fucoidan is a complex sulfated
polysaccharide extracted from brown seaweed. It has been reported
to exert antitumor activity in various types of cancer, including
acute prostate cancer cells, myeloid leukemia, lung cancer and
hepatocellular carcinoma cells (15–18). Our
previous study reported that fucoidan induces apoptosis of
different breast cancer cell lines, including mouse breast cancer
4T1 cells, human breast cancer MCF-7 cells and MDA-MB-231 cells
(19–21). However, the effects of fucoidan on
the metastasis of TNBC and the underlying mechanisms remain
elusive. The present study aimed to examine the effects of fucoidan
on EMT and the underlying molecular mechanisms in a TNBC cell line.
Since hypoxia may promote EMT in mammary cancer cells via
regulation of the expression of EMT regulators, the present study
also aimed to clarify whether fucoidan can directly affect HIF-1α
in a TNBC cell line.
Materials and methods
Materials
Fucoidan was obtained from Sigma-Aldrich (Merck
KGaA, Darmstadt, Germany). It was dissolved in normal saline at a
concentration of 20 mg/ml and then stored at −20°C. A human mammary
cancer cell line (MDA-MB-231) was purchased from the Cell Bank of
the Shanghai Institute of Cell Biology (Shanghai, China). RPMI-1640
medium was obtained from HyClone (GE Healthcare Life Sciences;
Logan, UT, USA) and was supplemented with 1%
penicillin/streptomycin and 10% fetal bovine serum (FBS; Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA). Antibodies
against CAIX (cat. no. A1463-100), HIF-1α (cat. no. TA301442),
vimentin (cat. no. 3634-100), N-cadherin (cat. no. 119-14215) and
histone H3 (cat. no. A310-257A) were obtained from Abcam
(Cambridge, MA, USA). Antibodies against zonula occludens-1 (ZO-1,
cat. no. sc-33725), E-cadherin (cat. no. sc-71008), TWIST (cat. no.
sc-81417), Snail (cat. no. sc-393172), GLUT-1 (cat. no. sc-377228),
Na, K-ATPase (cat. no. sc-71638) and β-actin (cat. no. sc-70319)
were purchased from Santa Cruz Biotechnology, Inc. (Dallas, TX,
USA). Secondary anti-rabbit (cat. no. sc-2491) and anti-mouse (cat.
no. sc-516102) antibodies were obtained from Santa Cruz
Biotechnology, Inc.
Cell culture
MDA-MB-231 cells were cultured in Roswell Park
Memorial Institute 1640 medium (Hyclone; GE Healthcare Life
Sciences, Logan, UT, USA), supplemented with 10% heat-inactivated
FBS (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) and
1% penicillin/streptomycin at 37°C in a humidified atmosphere
containing 5% CO2, and were allowed to attach onto the
dish. All cells were cultured to ~90% confluence in 6-well plates
prior to treatment. A hypoxic humidified incubator with a gas
mixture of 94% N2, 5% CO2 and 1%
O2 was used for hypoxia exposure. All cells were
incubated under hypoxia during or after fucoidan treatment.
MTT assay
MTT (Sigma-Aldrich; Merck KGaA) was used for the
cell proliferation assay. Each well of a 96-well plate was seeded
with 1×104 cells/ml. After overnight incubation at 37°C,
a total of 6.25, 12.5 or 25 µg/ml fucoidan was added to the breast
cancer cells. The plates were incubated in a hypoxic chamber at
37°C. Following incubation for 24, 48 or 72 h, 20 µl MTT reagent
was added to all wells at a concentration of 5 mg/ml at 37°C for 4
h, and the crystals were dissolved in DMSO. The absorbance was
detected at 490 nm on an ELISA reader (BioTek Instruments, Inc.,
Winooski, VT, USA).
Wound-healing assay
Breast cancer cells were grown until confluent in
6-well plates. Following treatment with fucoidan (0, 6.25, 12.5 or
25 µg/ml) for 24 h in a hypoxic chamber, the cell monolayers were
scratched using a sterile 200 µl plastic pipette tip. Following
aspiration of the medium, the displaced cells were washed with PBS.
Subsequently, the cells were continuely incubated in a hypoxic
chamber for 24 h. Five images were captured from randomly selected
microscope fields at different time points using an OLYMPUS CX31
microscope (Tokyo, Japan). The area of the wound was measured using
ImageJ 1.51p software (National Institutes of Health, Bethesda, MD,
USA).
Transwell invasion assay
For the invasion assay, 24-well Transwell units were
used. The lower units of a 6.5-mm Transwell filter with 0.4 µm pore
size were precoated with Matrigel (BD Biosciences, San Jose, CA,
USA). Following treatment with fucoidan (0, 6.25, 12.5 or 25 µg/ml)
for 24 h in a hypoxic chamber, the cancer cells were suspended in
serum-free medium, and 1×104 cells were seeded onto the
upper wells. The lower units were filled with medium containing 10%
FBS as a chemoattractant. After incubation in a hypoxic chamber for
12 h, the cancer cells on the upper units of the membrane were
removed using a cotton swab. The invading cells on the bottom
surface of the membrane were fixed with 70% ethanol for 10 min.
After staining with 0.1% crystal violet for 10 min at room
temperature, the cells were counted using an OLYMPUS CX31
microscope.
HIF-1 activation assay
MDA-MB-231 cells treated with fucoidan (0, 6.25,
12.5 or 25 µg/ml) for 48 h in a hypoxic chamber were washed with
PBS. HIF-1α activation was evaluated using a HIF-1 activation assay
kit (cat. no. 47096; Active Motif, Carlsbad, CA, USA), according to
the manufacturer's protocols. Finally, the absorbance was detected
at 450 nm with a SpectraMax 190 plate reader (Molecular Devices,
LLC, Sunnyvale, CA, USA).
Overexpression of HIF-1α
Breast cancer cells were transfected with HIF-1α
overexpression vector (HIF-1α-pcDNA3.0; Shanghai Genechem Co.,
Ltd., Shanghai, China) or empty plasmid (pcDNA3.0; Shanghai
Genechem Co., Ltd.) using Lipofectamine® 2000
(Invitrogen, Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocols. Briefly, the cells were prepared into a
single cell suspension with 0.2–1 ×106/ml density by
trypsin digestion and then inoculated into the 6-well plate. The
transfection was conducted when the cells had reached 90–95%
confluence. Each well contained 4 µg plasmid and 10 µl
Lipofectamine® 2000. The follow-up experiments were
continued 24 h after transfection. The sequence information of
overexpression vector was shown in Fig.
1.
Western blotting
Nuclear extracts, cytosolic extracts and membrane
extracts were prepared using the Nuclear and Cytoplasmic Protein
Extraction Kit and Membrane and Cytosol Protein Extraction kit
(Beyotime Institute of Biotechnology, Shanghai, China). The
membrane protein samples were used to detect the levels of ZO-1 and
E-cadherin, and Na, K-ATPase was used as a reference for
determination. The cytoplasmic protein samples were used to detect
the expression levels of vimentin, TWIST, Snail, CAIX, N-cadherin
and GLUT-1, and β-actin was used as a reference for determination.
The nuclear protein samples were used to detect HIF-1α levels, and
histone H3 was used as a reference.
The total protein concentrations were determined
using a BCA protein assay kit (Beyotime Institute of
Biotechnology). Each protein sample (30 µg) was separated by
SDS-PAGE (9% gel) and then transferred to nitrocellulose (NC)
membranes, which were obtained from Beijing Solarbio Science &
Technology Co., Ltd. (Beijing, China). After blocking with 5%
non-fat dry milk in Tris-buffered saline with Tween-20 buffer
(TBST; 20 mM Tris-HCl, 120 mM NaCl, 0.1% Tween-20) at room
temperature for 1 h, the NC membranes were incubated with primary
antibodies at 4°C. The dilution ratio for anti-ZO-1, anit-vimentin
and anti-E-cadherin was 1:1,000. The dilution ratio for anti-TWIST,
anti-Snail, anti-CAIX, anti-N-cadherin and anti-GLUT-1 was 1:200.
The dilution ratio for anti-HIF-1α, anti-Na, K-ATPase, anti-β-actin
and anti-Histone H3 was 1:500. Following overnight incubation, the
NC membranes were washed three times with TBST for 10 min and were
then incubated with the corresponding secondary antibodies
(dilution, 1:80) at room temperature for 1 h. After washing,
protein bands were visualized by enhanced chemiluminescence with a
reagent containing the horseradish peroxidase (HRP) substrate
luminol, which was obtained from EMD Millipore (Billerica, MA,
USA). Protein expression was quantified using Gel-Pro Analyzer 3.1
software (Media Cybernetics, Inc., Rockville, MD, USA).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA in cells after transfection was extracted
using an RNAprep pure tissue kit (Tiangen Biotech Co., Ltd.,
Beijing, China). cDNA was synthesized by RT using 200 ng total RNA.
Each 20 µl RT reaction was performed using a miScript II Reverse
Transcriptase kit (Qiagen GmbH, Hilden, Germany) according the
manufacturer's protocol in a GeneAmp PCR system 9700 (Applied
Biosystems; Thermo Fisher Scientific, Inc.) for 60 min at 37°C,
followed by heat inactivation of the RT for 5 min at 95°C. The cDNA
was then subjected to RT-qPCR amplification on a LightCycler 480 RT
PCR system (Roche Diagnostics, Basel, Switzerland). The RT PCR
Master mix (Takara Biotechnology Co., Ltd., Dalian, China) and
specific primer/probe sets were used to amplify HIF-1α and GAPDH
(TaqMan Gene Expression Assays; Applied Biosystems; Thermo Fisher
Scientific, Inc.). The following primer pairs were used for qPCR:
HIF-1α, forward, 5′-CCTGAGCCTAATAGTCCC-3′, reverse,
5′-GCTGGCATTAGCAGTAGG-3′; and GAPDH, forward,
5′-ATTCCATGGCACCGTCAAGGCT-3′ and reverse,
5′-TCAGGTCCACCACTGACACGTT-3′ (designed by Takara Biotechnology Co.
Ltd.). The amplification consisted of 95°C for 30 sec and 40 cycles
of 95°C for 5 sec, 60°C for 30 sec, and 72°C for 2 min followed by
a final extension at 72°C for 5 min. The expression levels of each
gene were normalized to GAPDH expression levels using the
2−ΔΔCq method as previously described (22).
Statistical analysis
All results are expressed as the means ± standard
deviation. Each experiment was repeated three times. Statistical
analysis was performed using SPSS software (version 17; SPSS, Inc.,
Chicago, IL, USA). One-way analysis of variance was used to
determine the statistical significance and Tukey's post hoc test
was used to analyze the differences between two groups. P<0.05
was considered to indicate a statistically significant
difference.
Results
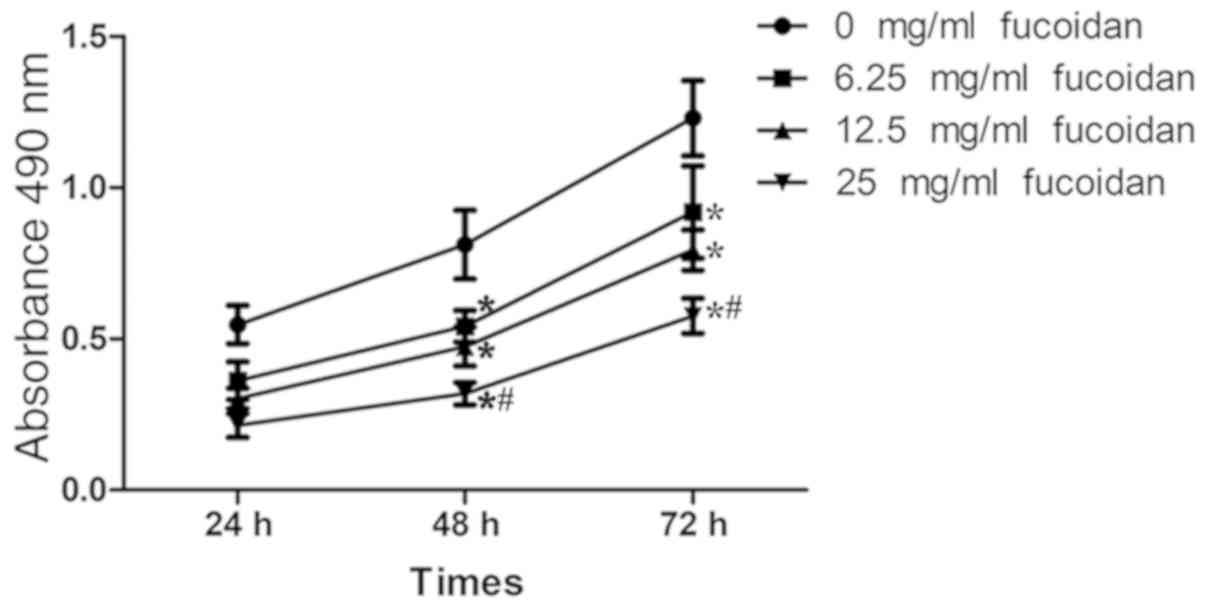
Fucoidan treatment inhibits the
proliferation of MDA-MB-231 cells under hypoxia
The effect of fucoidan on the proliferation of
breast cancer cells under hypoxia was determined using an MTT
assay. Fig. 2 indicated that
fucoidan treatment reduced the proliferation of breast cancer cells
(P<0.05). Following treatment with fucoidan at a concentration
of 25 µg/ml for 72 h, the cell proliferation had declined by
53.2%.
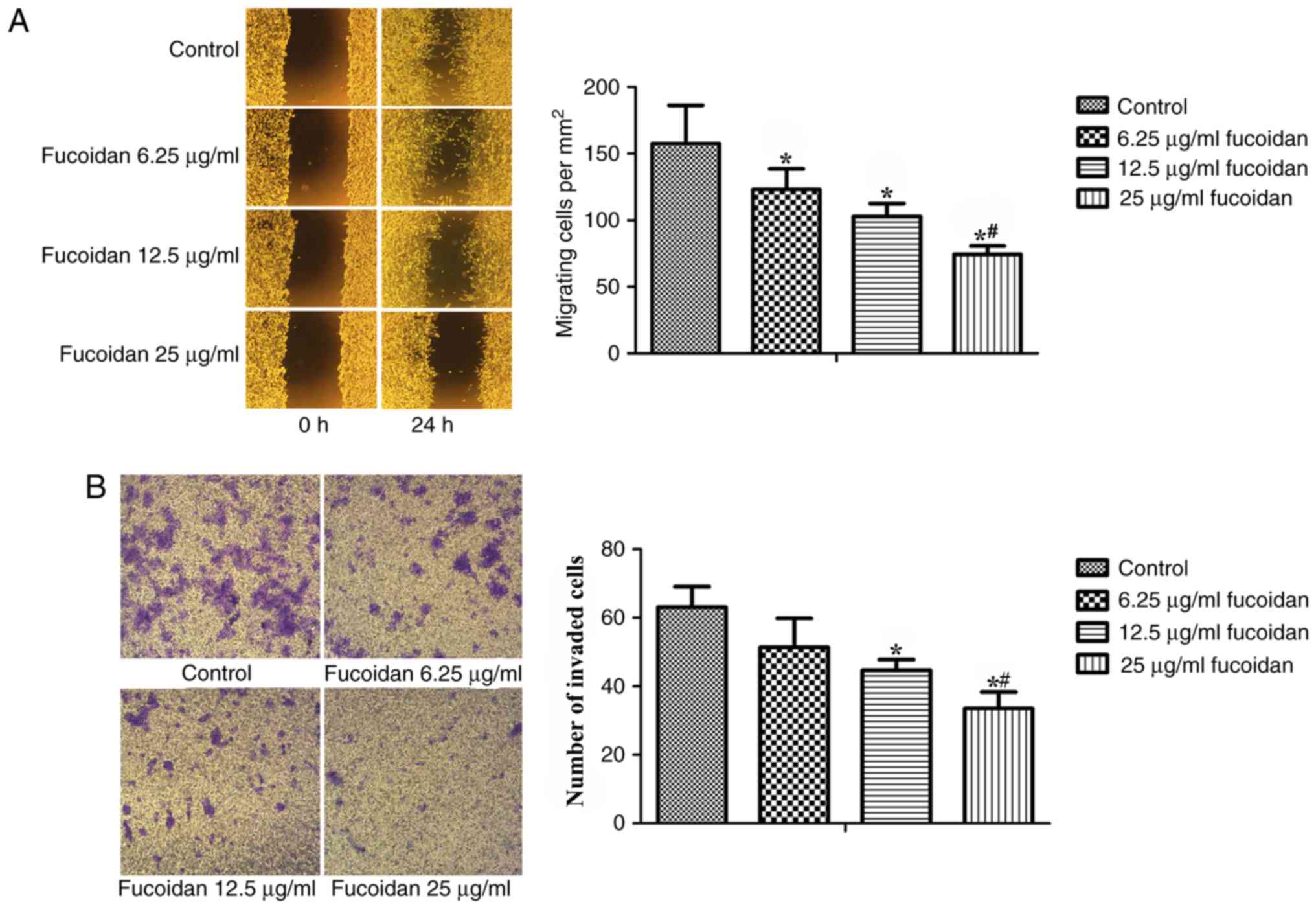
Fucoidan inhibits the migration and
invasion of breast cancer cells
Wound-healing and Matrigel-coated Transwell invasion
assays were used to detect whether fucoidan intervention exhibited
any effect on EMT in MDA-MB-231 cells. As shown in Fig. 3A, fucoidan treatment reduced the
migratory ability of breast cancer cells (P<0.001). Following
treatment with fucoidan at a concentration of 12.5 or 25 µg/ml for
24 h, cell invasion was also inhibited (Fig. 3B).
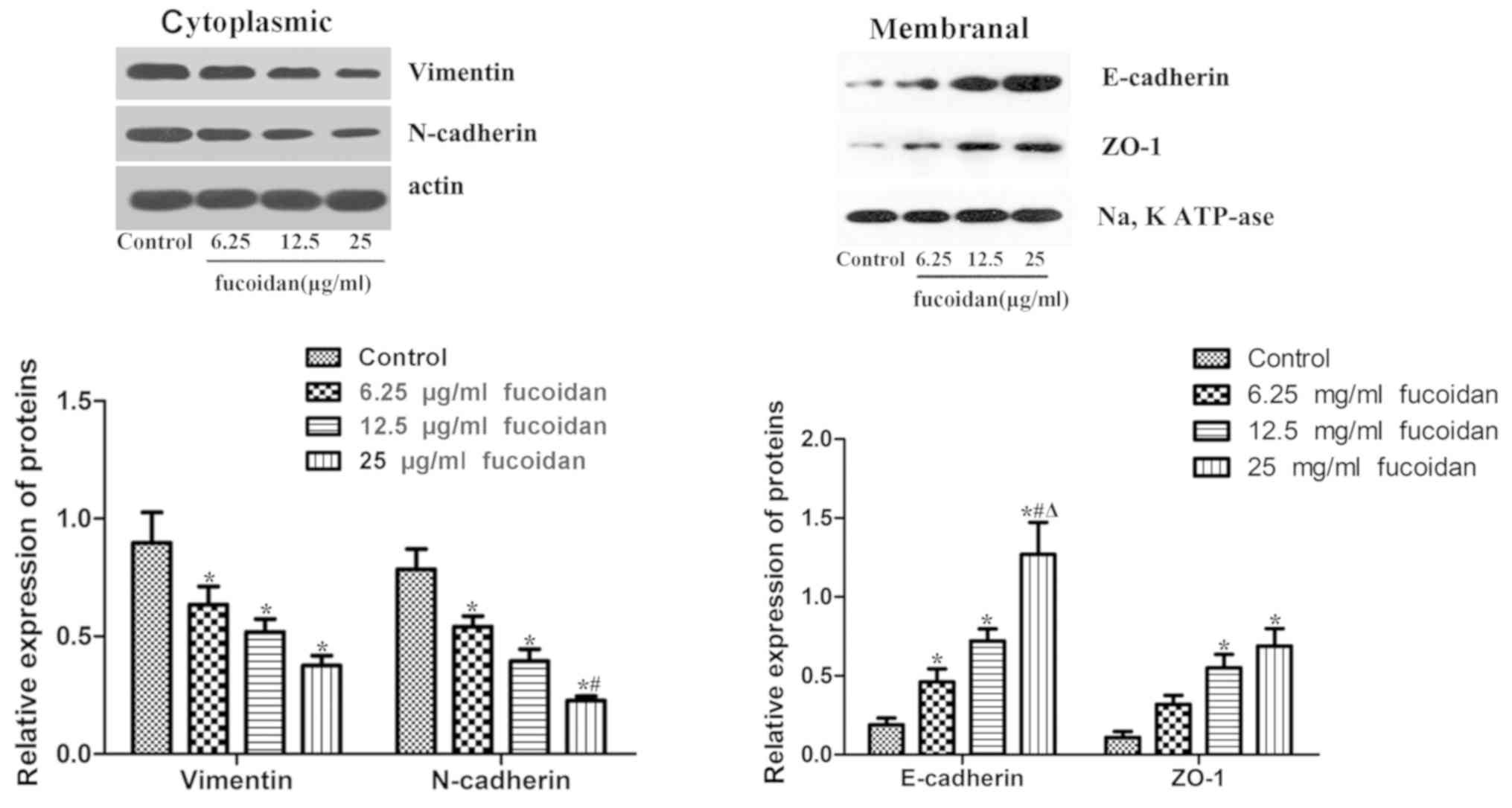
Fucoidan affects the expression levels
of EMT markers
The results revealed that fucoidan treatment
downregulated the expression levels of the mesenchymal markers
N-cadherin and vimentin (Fig. 4). In
the membranes of MDA-MB-231 cells, the epithelial markers ZO-1 and
E-cadherin were rarely expressed, but following treatment with
fucoidan, the expression levels of ZO-1 and E-cadherin were
significantly increased.
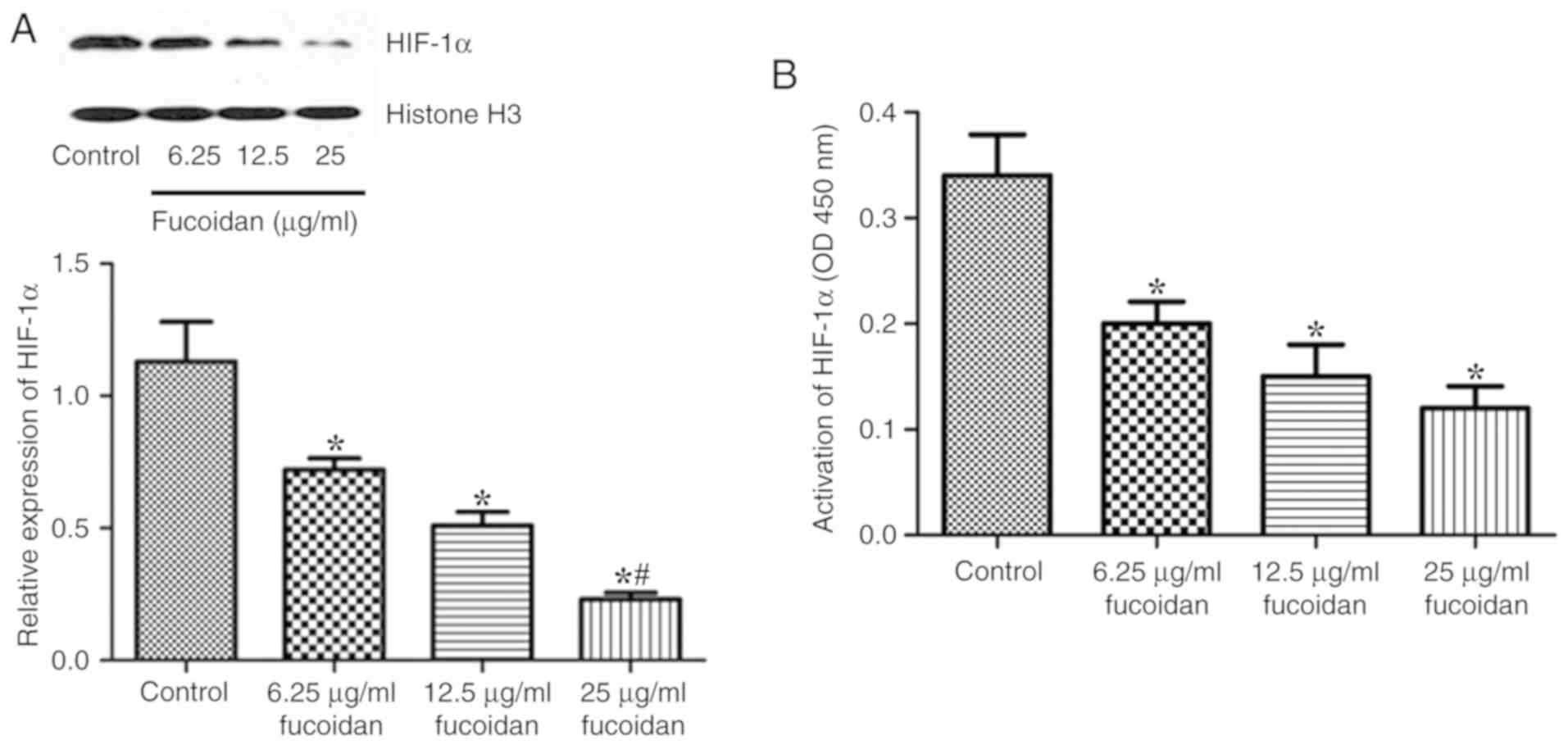
Fucoidan inhibits protein accumulation
in the nucleus and activation of HIF-1α
The effects of fucoidan on the expression and
nuclear translocation of HIF-1α were assessed by western blotting.
The nuclear protein levels of HIF-1α were decreased following
treatment with fucoidan under hypoxic conditions (Fig. 5A). The activation of HIF-1α was also
assessed. As shown in Fig. 5B,
fucoidan treatment decreased the activation of HIF-1α.
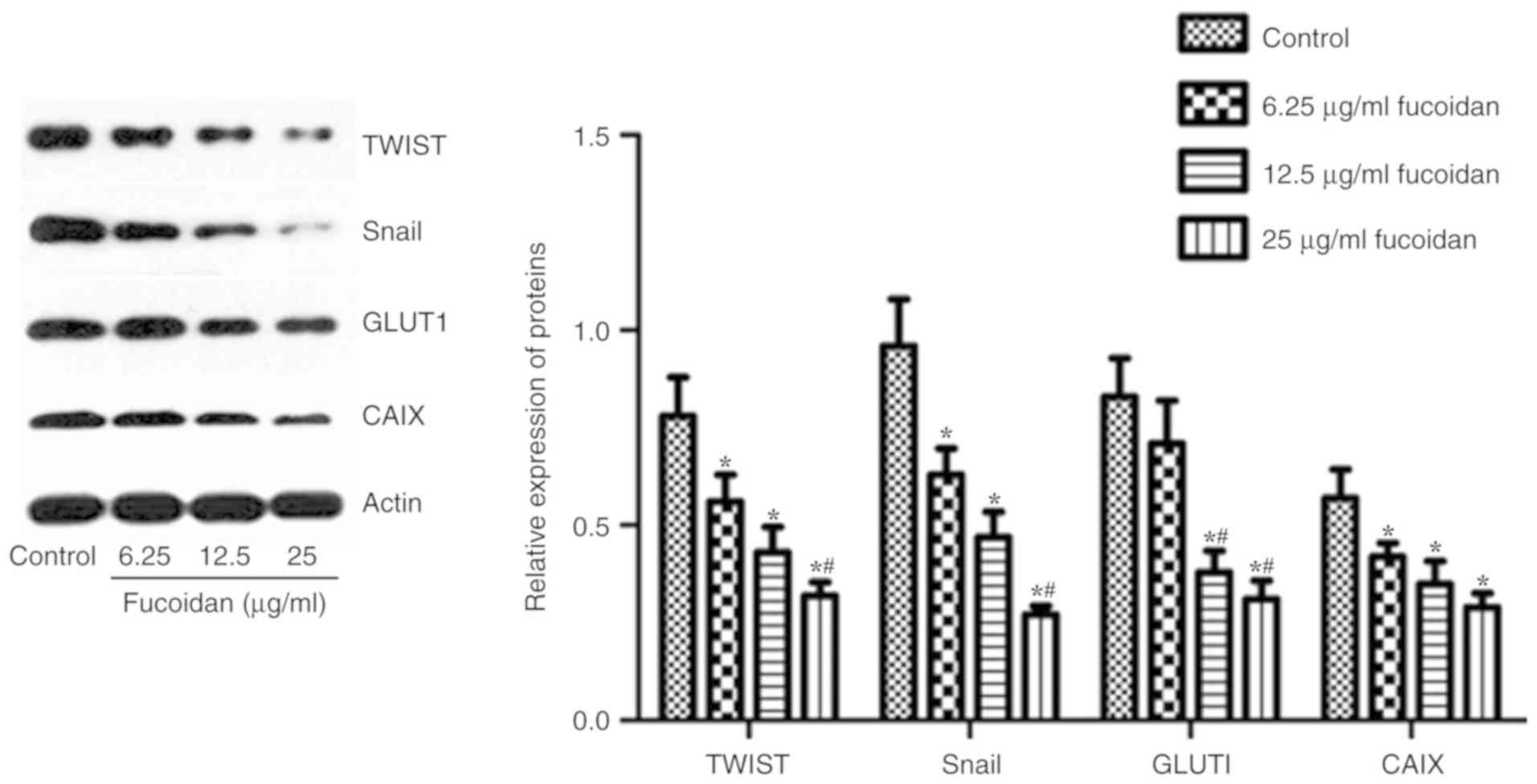
Fucoidan downregulates the expression
of HIF-1α target genes
HIFs can regulate the expression of numerous genes
involved in tumor growth, metastasis and metabolic reprogramming
(23,24). HIF-1α can directly or indirectly
regulate the EMT regulators TWIST, Snail, CAIX and GLUT-1 (9–11). The
expression levels of TWIST, Snail, CAIX and GLUT-1 were detected by
western blotting. As shown in Fig.
6, following treatment with fucoidan, the expression levels of
TWIST-1, Snail, CAIX and GLUT-1 were inhibited.
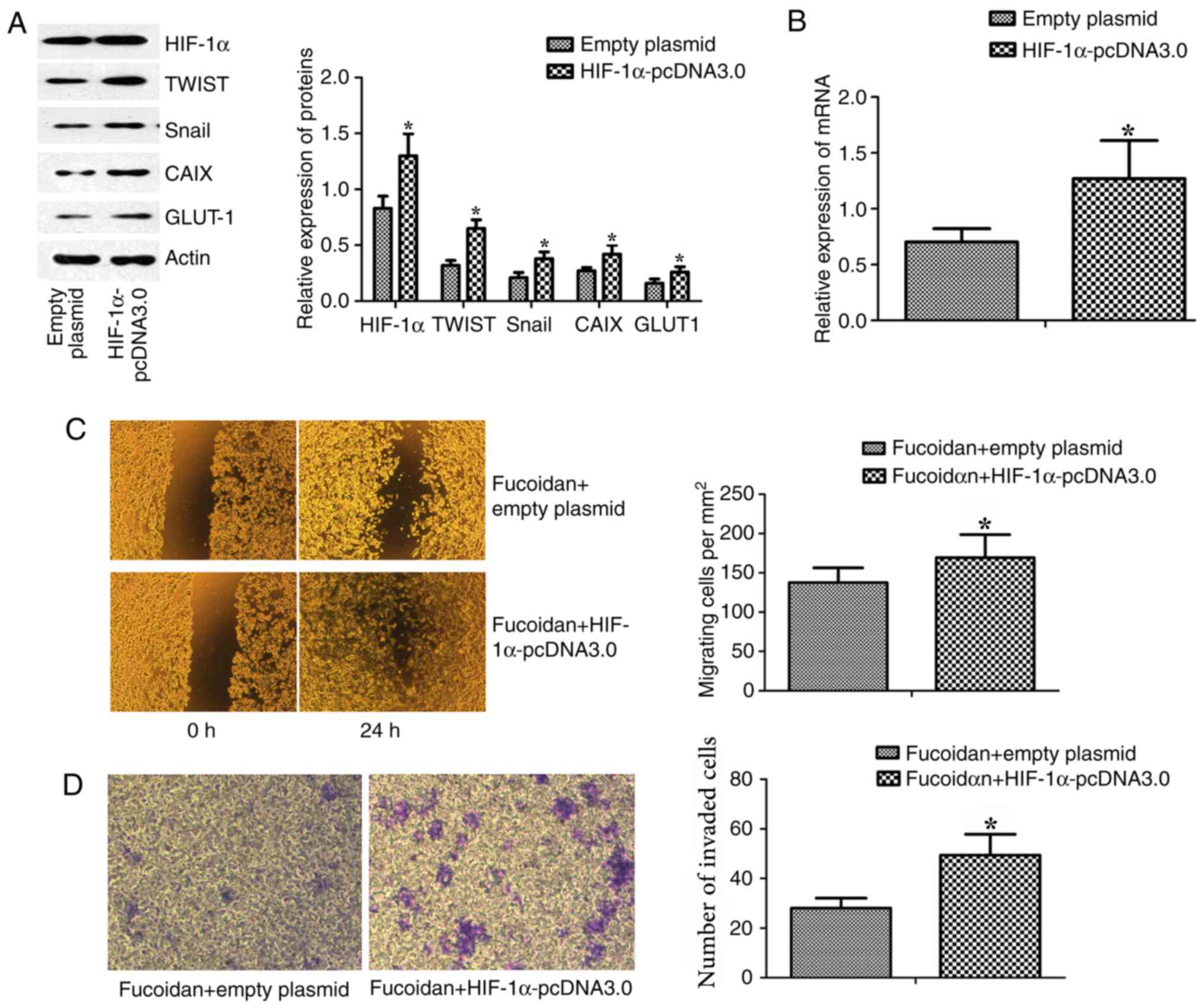
Overexpression of HIF-1α results in
the reversal of fucoidan-mediated suppression of cell migration and
invasion
MDA-MB-231 cells were transfected with HIF-1α
overexpression plasmid (HIF-1α-pcDNA3.0) to determine whether the
anti-EMT effects of fucoidan depended on HIF-1α; empty plasmid
(pcDNA3.0) was used as a control. The expression of HIF-1α and its
target genes (TWIST and Snail) in MDA-MB-231 cells was increased by
HIF-1α-pcDNA3.0 transfection (Fig.
7A). The mRNA expression levels of HIF-1α in breast cancer
cells were measured by RT-qPCR. Post-transfection with
HIF-1α-pcDNA3.0, the mRNA expression levels of HIF-1α were
increased (Fig. 7B).
Post-transfection with HIF-1α-pcDNA3.0, breast cancer cells were
incubated with 25 µg/ml fucoidan, and cell invasion and migration
were detected. Wound-healing and Matrigel-coated Transwell invasion
assays revealed that overexpression of HIF-1α resulted in the
reversal of fucoidan-mediated suppression of cell migration and
invasion (Fig. 7C and D).
Discussion
The present study demonstrated that fucoidan
treatment inhibited the proliferation of breast cancer cells, and
suppressed the migration and invasion of mammary cancer cells in a
hypoxic microenvironment. The nuclear translocation and activity of
HIF-1α were also reduced by fucoidan. In addition, fucoidan
significantly downregulated the expression levels of N-cadherin and
vimentin, but upregulated ZO-1 and E-cadherin expression.
During the process of EMT, cancer cells lose the
characteristics of epithelial cells, such as polarity and cell
adhesion. Subsequently, cancer cells acquire a mesenchymal
cell-like morphology and a migratory ability. Once cancer cells
cease to express epithelial markers, including E-cadherin (an
adherens junction protein), they initiate the expression of
mesenchymal markers, including N-cadherin and vimentin (25). ZO-1 is another epithelial marker and
is a submembrane scaffolding protein. When ZO-1 relocates into the
cell from tight junctions, it may promote the invasion activity
(26).
HIFs are critical transcription factors that can
regulate adaptive cellular responses to low O2
concentrations in Metazoa. HIFs have been reported to be
upregulated in various cancer cells under hypoxia, which is
commonly found in tumor microenvironments (23,24).
HIF-1 has two subunits, the HIF-1α and HIF-1β
subunits; these two subunits form a heterodimeric protein. The
partial pressure of oxygen can regulate the expression of the
HIF-1α subunit; however, HIF-1β is constitutively expressed
(27,28). At normal partial pressure of oxygen,
HIF-1α can be hydroxylated by O2 and
α-ketoglutarate-dependent prolyl hydroxylases. Hydroxylated HIF-1α
can be recognized by the von Hippel-Lindau tumor suppressor
protein, which targets HIF-1α to be degraded via ubiquitination.
Therefore, HIF-1α levels are kept low in the cytoplasm. However,
under hypoxic conditions, HIF-1α cannot be hydroxylated, resulting
in accumulation of HIF-1α in the cytoplasm (29,30).
Once the presence of HIF-1α is stable in the cytoplasm, HIF-1α can
be transported into the nucleus. Subsequently, the two subunits,
HIF-1α and HIF-1β, can form a heterodimer. The HIF-1α/1β
heterodimer can bind to hypoxia-response elements (HREs). HREs
exist in hypoxia target genes, and these genes are involved in
tumor growth, metastasis, metabolic reprogramming, chemoresistance
and radioresistance (23,24,31)
It has been reported that HIF-1α is hyperactivated
in TNBC (32). High HIF-1α levels in
cancer tissues are associated with high mortality in several tumor
types, including mammary cancer. Furthermore, elevated HIF-1α
levels in the mammary tumor tissues of patients have been
associated with high rates of metastasis and mortality (27). The present results showed that
fucoidan treatment in highly metastatic TNBC cell lines may inhibit
the nuclear accumulation and activation of HIF-1α.
HIF-1α is an important transcription factor that can
induce the expression of ~40 genes in hypoxic stromal and cancer
cells, including glycolytic enzymes, glucose transporters,
erythropoietin, and vascular endothelial growth factors. In
addition, HIF-1α can increase the expression of some genes, and the
protein products of these genes can enhance the delivery of oxygen
or promote metabolic adaptation to hypoxia, as well as increase
metastasis and tumor invasion (33,34).
HIF-1α can directly or indirectly regulate EMT
regulators, and these transcription factors can transactivate
EMT-related genes, including N-cadherin, vimentin and E-cadherin
(12,13). It has been reported that hypoxia and
continuously high levels of HIF-1α can result in high levels of
certain regulators, including TWIST, zinc finger E-box-binding
homeobox (ZEB)2, Snail, transcription factor 3, ZEB1, CAIX and
GLUT-1, to moderate EMT and metastasis (9–11,35,36).
TWIST is an important factor that induces EMT and
metastasis in cancer. HIF-1α can directly increase TWIST levels via
the response element of HIF1 located in the proximal promoter of
the TWIST gene (9,37). TWIST can bind to the E-box in the
promoter of E-cadherin and downregulate the expression of
E-cadherin (38). Downregulation of
TWIST is important for invasion in TNBC (39). Snail is an important target gene of
HIF, and overexpression of Snail is involved in EMT. Furthermore,
high levels of Snail are associated with highly aggressive types of
cancer in humans and mice (40).
Snail overexpression is associated with a high rate of metastasis
in mammary tumors, whereas Snail silencing decreases the
invasiveness and cell motility of mammary cancer (41).
HIF-1 can result in metabolic adaptation (or
‘reprogramming’) to hypoxic and energy-deprived conditions, and can
enable tumor cells to survive. HIF-1 upregulates glucose influx
through GLUT-1 and increases proton efflux via membranous CAIX.
CAIX is involved in the hydration of carbon dioxide to form
carbonic acid, which suggests a putative role of this enzyme in the
acidification of the tumor environment. A high level of CAIX in
patients with mammary cancer has been associated with a short
disease-free survival time (42).
Hypoxia in tumor tissues can markedly induce the expression of CAIX
and has been proposed to be involved in acidification of the tumor
microenvironment. In addition, hypoxia can increase cell adhesion
and invasiveness. It has been reported that high levels of CAIX are
linked to poor prognosis via regulation of the EMT (43). CAIX has the capacity to modulate cell
adhesion mediated by E-cadherin through interactions with β-catenin
(44). GLUT-1 levels are positively
associated with the expression of vimentin and N-cadherin in tumor
tissues but are negatively associated with the levels of
E-cadherin. Compared to the survival rates of patients with low
levels of GLUT-1, vimentin and N-cadherin, those of patients with
high levels of GLUT-1, vimentin and N-cadherin are much lower
(45). In the present study,
fucoidan treatment decreased the expression of HIF-1α target genes,
including TWIST, Snail, CAIX and GLUT-1, in cells in a hypoxic
environment.
The mechanism by which fucoidan downregulates the
HIF-1α signaling pathway will be the next question explored by this
research team. In vivo experiments are required to further
confirm the effects of downregulation of this pathway on inhibition
of migration and invasion. In addition, the present study suggested
that HIF-1α may be a possible novel target for the treatment of
TNBC. In animal experiments, we aim to further determine the
antitumor effect of fucoidan on TNBC, and focus on the preventative
and inhibitory effects of fucoidan and its mechanism on tumor
metastasis. Once the inhibitory effects and target of action of
fucoidan on tumor metastasis have been confirmed and the
appropriate dosage of fucoidan has been clarified, further clinical
trials may be conducted. This would help to develop fucoidan into a
natural antitumor drug.
In conclusion, to the best of our knowledge, the
present study identified a novel mechanism by which fucoidan may
inhibit EMT and metastasis through the HIF-1α signaling pathway in
mammary cancer. Fucoidan inhibited the activation and nuclear
accumulation of HIF-1α. EMT regulators downstream of HIF-1α,
including TWIST, Snail, CAIX and GLUT-1, were also downregulated.
Subsequently, the expression levels of N-cadherin and vimentin,
which are typical markers of EMT, were decreased, whereas the
levels of ZO-1 and E-cadherin were increased, leading to the
inhibition of EMT and migration in TNBC cells under hypoxic
conditions. Therefore, the present study provides novel insight
into the mechanisms of fucoidan in mammary cancer treatment.
Acknowledgements
The authors wish to thank Professor Yinlin Ge and Dr
Zheng Zheng (Department of Medicine, Qingdao University, Qingdao,
Shandong, China) for their helpful comments and discussion on this
study.
Funding
The present work was supported by the National
Nature Science Foundation of China (grant no. 81502298), the
Shandong Provincial Natural Science Foundation (grant no.
ZR2014JL056), the Qingdao Postdoctoral Application Research Project
(grant no. 2015165) and the Qingdao People's Livelihood Science and
Technology Project (grant no. 18-6-1-70-nsh).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
MX and HL analyzed and interpreted the data. WL, JZ
and TS performed the cell culture, proliferation assay,
overexpression of HIF-1α and examination of protein factors. DX and
YL performed migration and invasion assays. MX was a major
contributor in writing the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Matsuoka J, Yashiro M, Doi Y, Fuyuhiro Y,
Kato Y, Shinto O, Noda S, Kashiwagi S, Aomatsu N, Hirakawa T, et
al: Hypoxia stimulates the EMT of gastric cancer cells through
autocrine TGFβ signaling. PLoS One. 8:e623102013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cannito S, Novo E, Compagnone A, Valfrè di
Bonzo L, Busletta C, Zamara E, Paternostro C, Povero D, Bandino A,
Bozzo F, et al: Redox mechanisms switch on hypoxia-dependent
epithelial-mesenchymal transition in cancer cells. Carcinogenesis.
29:2267–2278. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lester RD, Jo M, Campana WM and Gonias SL:
Erythropoietin promotes MCF-7 breast cancer cell migration by an
ERK/mitogen-activated protein kinase-dependent pathway and is
primarily responsible for the increase in migration observed in
hypoxia. J Biol Chem. 280:39273–39277. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tomaskovic-Crook E, Thompson EW and Thiery
JP: Epithelial to mesenchymal transition and breast cancer. Breast
Cancer Res. 11:2132009. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kotiyal S and Bhattacharya S: Breast
cancer stem cells, EMT and therapeutic targets. Biochem Biophys Res
Commun. 453:112–116. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Majmundar AJ, Wong WJ and Simon MC:
Hypoxia-inducible factors and the response to hypoxic stress. Mol
Cell. 40:294–309. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Luo D, Wang Z and Wu J, Jiang C and Wu J:
The role of hypoxia inducible factor-1 in hepatocellular carcinoma.
Biomed Res Int. 2014:4092722014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ji RC: Hypoxia and lymphangiogenesis in
tumor microenvironment and metastasis. Cancer Lett. 346:6–16. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang MH, Wu MZ, Chiou SH, Chen PM, Chang
SY, Liu CJ, Teng SC and Wu KJ: Direct regulation of TWIST by
HIF-1alpha promotes metastasis. Nat Cell Biol. 10:295–305. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tsai YP and Wu KJ: Hypoxia-regulated
target genes implicated in tumor metastasis. J Biomed Sci.
19:1022012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lendahl U, Lee KL, Yang H and Poellinger
L: Generating specificity and diversity in the transcriptional
response to hypoxia. Nat Rev Genet. 10:821–832. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
de Herreros AG, Peiró S, Nassour M and
Savagner P: Snail family regulation and epithelial mesenchymal
transitions in breast cancer progression. J Mammary Gland Biol
Neoplasia. 15:135–147. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Evans AJ, Russell RC, Roche O, Burry TN,
Fish JE, Chow VW, Kim WY, Saravanan A, Maynard MA, Gervais ML, et
al: VHL promotes E2 box-dependent E-cadherin transcription by
HIF-mediated regulation of SIP1and snail. Mol Cell Biol.
27:157–169. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bos R, van der Groep P, Greijer AE,
Shvarts A, Meijer S, Pinedo HM, Semenza GL, van Diest PJ and van
der Wall E: Levels of hypoxia-inducible factor-1α independently
predict prognosis in patients with lymph node negative breast
carcinoma. Cancer. 97:1573–1581. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wei C, Xiao Q, Kuang X, Zhang T, Yang Z
and Wang L: Fucoidan inhibits proliferation of the SKM-1 acute
myeloid leukaemia cell line via the activation of apoptotic
pathways and production of reactive oxygen species. Mol Med Rep.
12:6649–6655. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Boo HJ, Hong JY, Kim SC, Kang JI, Kim MK,
Kim EJ, Hyun JW, Koh YS, Yoo ES and Kwon JM: The anticancer effect
of fucoidan in PC-3 prostate cancer cells. Mar Drugs. 11:2982–2999.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen H, Cong Q, Du Z, Liao W, Zhang L, Yao
Y and Ding K: Sulfated fucoidan FP08S2 inhibits lung cancer cell
growth in vivo by disrupting angiogenesis viatargeting VEGFR2/VEGF
and blocking VEGFR2/Erk/VEGF signaling. Cancer Lett. 382:44–52.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yan MD, Yao CJ, Chow JM, Chang CL, Hwang
PA, Chuang SE, Whang-Peng J and Lai GM: Fucoidan elevates
MicroRNA-29b to regulate DNMT3B MTSS1 axis and inhibit EMT in human
hepatocellular carcinoma cells. Mar Drugs. 13:6099–6116. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xue M, Ge Y, Zhang J, Liu Y, Wang Q, Hou L
and Zheng Z: Fucoidan inhibited 4T1 mouse breast cancer cell growth
in vivo and in vitro via downregulation of Wnt/β-catenin signaling.
Nutr Cancer. 65:460–468. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xue M, Ge Y, Zhang J, Wang Q, Hou L, Liu
Y, Sun L and Li Q: Anticancer properties and mechanisms of fucoidan
on mouse breast cancer in vitro and in vivo. PLoS One.
7:e434832012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xue M, Ji X, Xue C, Liang H, Ge Y, He X
and Zhang L, Bian K and Zhang L: Caspase-dependent and
caspase-independent induction of apoptosis in breast cancer by
fucoidan via the PI3K/AKT/GSK3β pathway in vivo and in vitro.
Biomed Pharmacother. 94:898–908. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang SW, Zhang ZG, Hao YX, Zhao YL, Qian
F, Shi Y, Li PA, Liu CY and Yu PW: HIF-1α induces the
epithelial-mesenchymal transition in gastric cancer stem cells
through the Snail pathway. Oncotarget. 8:9535–9545. 2017.PubMed/NCBI
|
|
24
|
Melillo G: Targeting hypoxia cell
signaling for cancer therapy. Cancer Metastasis Rev. 26:341–352.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hay ED: An overview of
epithelio-mesenchymal transformation. Acta Anat (Basel). 154:8–20.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lesage J, Suarez-Carmona M,
Neyrinck-Leglantier D, Grelet S, Blacher S, Hunziker W, Birembaut
P, Noël A, Nawrocki-Raby B, Gilles C and Polette M: Zonula
occludens-1/NF-κB/CXCL8: A new regulatory axis for tumor
angiogenesis. FASEB J. 31:1678–1688. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Semenza GL: Cancer-stromal cell
interactions mediated by hypoxia-inducible factors promote
angiogenesis, lymphangiogenesis, and metastasis. Oncogene.
32:4057–4063. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang GL, Jiang BH, Rue EA and Semenza GL:
Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS
heterodimer regulated by cellular O2 tension. Proc Natl Acad Sci
USA. 92:5510–5514. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Salceda S and Caro J: Hypoxia-inducible
factor 1alpha (HIF-1alpha) protein is rapidly degraded by the
ubiquitin-proteasome system under normoxic conditions. Its
stabilization by hypoxia depends on redox-induced changes. J Biol
Chem. 272:22642–22647. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Semenza GL: HIF-1: Upstream and downstream
of cancer metabolism. Curr Opin Genet Dev. 20:51–56. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shannon AM, Bouchier-Hayes DJ, Condron CM
and Toomey D: Tumor hypoxia, chemotherapeutic resistance and
hypoxia-related therapies. Cancer Treat Rev. 29:297–307. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Montagner M, Enzo E, Forcato M, Zanconato
F, Parenti A, Rampazzo E, Basso G, Leo G, Rosato A, Bicciato S, et
al: SHARP1 suppresses breast cancer metastasis by promoting
degradation of hypoxia-inducible factors. Nature. 487:380–384.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wigerup C, Påhlman S and Bexell D:
Therapeutic targeting of hypoxia and hypoxia-inducible factors in
cancer. Pharmacol Ther. 164:152–169. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Finger EC and Giaccia AJ: Hypoxia,
inflammation, and the tumor microenvironment in metastatic disease.
Cancer Metastasis Rev. 29:285–293. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Peinado H, Olmeda D and Cano A: Snail, Zeb
and bHLH factors in tumour progression: An alliance against the
epithelial phenotype? Nat Rev Cancer. 7:415–428. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chu CY, Jin YT, Zhang W, Yu J, Yang HP,
Wang HY, Zhang ZJ, Liu XP and Zou Q: CA IX is upregulated in
CoCl2-induced hypoxia and associated with cell invasive potential
and a poor prognosis of breast cancer. Int J Oncol. 48:271–280.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chen D, Dang BL, Huang JZ, Chen M, Wu D,
Xu ML, Li R and Yan GR: MiR-373 drives the
epithelial-to-mesenchymal transition and metastasis via the
miR-373-TXNIP-HIF1α-TWIST signaling axis in breast cancer.
Oncotarget. 6:32701–32712. 2015.PubMed/NCBI
|
|
38
|
Lopez D, Niu G, Huber P and Carter WB:
Tumor-induced upregulation of Twist, Snail, and Slug represses the
activity of the human VE-cadherin promoter. Arch Biochem Biophys.
482:77–82. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Montserrat N, Gallardo A, Escuin D,
Catasus L, Prat J, Gutiérrez-Avignó FJ, Peiró G, Barnadas A and
Lerma E: Repression of E-cadherin by SNAIL, ZEB1, and TWIST in
invasive ductal carcinomas of the breast: A cooperative effort? Hum
Pathol. 42:103–110. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cano A, Pérez-Moreno MA, Rodrigo I,
Locascio A, Blanco MJ, del Barrio MG, Portillo F and Nieto MA: The
transcription factor snail controls epithelial-mesenchymal
transitions by repressing E-cadherin expression. Nat Cell Biol.
2:76–83. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang A, Wang Q, Han Z, Hu W, Xi L, Gao Q,
Wang S, Zhou J, Xu G, Meng L, et al: Reduced expression of Snail
decreases breast cancer cell motility by downregulating the
expression and inhibiting the activity of RhoA GTPase. Oncol Lett.
6:339–346. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Tan EY, Yan M, Campo L, Han C, Takano E,
Turley H, Candiloro I, Pezzella F, Gatter KC, Millar EK, et al: The
key hypoxia regulated gene CAIX is upregulated in basal-like breast
tumours and is associated with resistance to chemotherapy. Br J
Cancer. 100:405–411. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hyuga S, Wada H, Eguchi H, Otsuru T,
Iwgami Y, Yamada D, Noda T, Asaoka T, Kawamoto K, Gotoh K, et al:
Expression of carbonic anhydrase IX is associated with poor
prognosis through regulation of the epithelial-mesenchymal
transition in hepatocellular carcinoma. Int J Oncol. 51:1179–1190.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Svastová E, Zilka N, Zat'ovicová M,
Gibadulinová A, Ciampor F, Pastorek J and Pastoreková S: Carbonic
anhydrase IX reduces E-cadherin-mediated adhesion of MDCK cells via
interaction with beta-catenin. Exp Cell Res. 290:332–345. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zuo J, Wen J, Lei M, Wen M, Li S, Lv X,
Luo Z and Wen G: Hypoxia promotes the invasion and metastasis of
laryngeal cancer cells via EMT. Med Oncol. 33:152016. View Article : Google Scholar : PubMed/NCBI
|