Introduction
RNA-binding proteins (RBPs) can directly interact
with RNA or can be part of ribonucleoprotein complexes without
direct contact with RNA (1). The
presence of RNA-binding domains (RBDs) in proteins and AU-enriched
regions in target RNAs is essential for the synergistic function of
RBPs on their targets, which include mRNA, tRNA, rRNA, microRNA
(miRNA) and non-coding RNA (ncRNA). Transient or stable
interactions of RBPs with RNAs are crucial for RNA regulatory
processes. RBPs are involved in post-transcriptional gene
regulation (PTGR) primarily through the formation of functional
units termed ribonucleoprotein (RNP) complexes. mRNA-binding
proteins and corresponding messenger RNPs are directly responsible
for mRNA maturation and regulation; however, diverse ncRNA-targeted
RBPs are also involved in the process of PTGR (2,3). RBPs
can combine with miRNAs to form miRNPs, which are also involved in
the regulation of mRNA translation and stability. According to an
extensive classification of 1,542 human RBPs conducted by
Gerstberger et al (1),
approximately half of these RBPs can be grouped on the basis of
their mRNA targets, and others may interact with different types of
RNAs. Furthermore, approximately one-third of RBPs can bind to DNA
and RNA (4). The diverse binding
spectrum of RBPs indicates their diverse spectrum of function.
Recent studies have demonstrated that RBPs often associate with a
group of mRNAs encoding proteins with similar functions, forming an
RNA operon (5). The different types
of RBPs combine with the corresponding mRNAs at different times and
in different positions inside cells, thereby meeting the need of
the cell in a time- and location-dependent manner to adjust the
regulation of target molecules. A particular mRNA can be bound by
numerous different RBPs. RBPs can also act as a binding platform
for other factors and enzymes involved in mRNA regulation. RBPs are
considered the most important regulators of PTGR. In addition to
sustaining cellular metabolism, coordinating maturation,
transportation, stability and degradation of all classes of RNAs
through PTGR, RBPs serve a critical role in maintaining genome
integrity (6,7) and responding to a variety of cellular
stresses, thus ensuring cellular homeostasis (8,9).
Considering the multifaceted effects of RBPs, dysfunctional RBPs
can initiate various pathological changes, including
neurodegenerative disorders, cardiovascular diseases and certain
types of cancer (10–12).
Cancer is a complex and heterogeneous disease, and
is classically considered to be caused by genetic alterations that
result in the activation of oncogenic signaling pathways and/or
loss of tumor suppressor mechanisms (13). As RBPs serve a pivotal role in PTGR,
it is not surprising that abnormal changes to RBPs can cause
alterations of cancer-associated signaling pathways. Furthermore,
as RBPs regulate multiple targets in various PTGR steps, small
changes in their expression and/or activity can induce a
large-scale alteration of downstream regulatory networks,
potentially initiating cancer development (13). A wide range of mechanisms underlie
RBP alteration-induced oncogenesis, including changes to
alternative splicing and polyadenylation, and changes in RNA
stability, subcellular localization and translation (13). In order for tumor cells to achieve
survival, proliferation, metastasis and resistance to anticancer
therapeutics, they must make adaptive changes to the gene
expression. Regulating transcribed mRNA or PTGR is the most
effective and rapid mechanism for doing so, and it has a pivotal
role in tumorigenesis. Tumor cells can change the expression of
target mRNAs and feedback regulators (miRNA and ncRNA) through the
regulation of RBPs. The abnormal expression of RBPs has been
detected in numerous types of tumor, and different RBPs act at
different steps of mRNA metabolism. For example, Sam68 participates
in alternative splicing, producing a variety of tumor-promoting
mRNA variants (14); eukaryotic
translation initiation factor 4E is involved in directing ribosomes
to the 5′-cap of mRNAs and enhances the expression of specific
mRNAs that regulate certain tumorigenesis-associated processes,
such as proliferation [c-Myc and cyclin-dependent kinase (CDK)2],
metastasis (matrix metalloproteinase 9) and angiogenesis (vascular
endothelial growth factor) (15),
while embryonic lethal abnormal visual-like RNA binding protein 1
regulates the stability and translation efficiency of tumor-related
mRNAs (16). Cytoplasmic
activation/proliferation-associated protein-1 (caprin-1) is an RBP
that is essential for cell proliferation. As caprin-1 is closely
associated with control of the cell cycle, alteration of caprin-1
is involved in oncogenesis, which has been demonstrated in multiple
experimental cancer studies (17–20). It
is important for the study of cancer and its therapeutic
development to fully understand the biological function of caprin-1
and the association between its alteration and oncogenesis.
Caprin-1 and RBPs
Caprin-1 is a ubiquitously expressed and highly
conserved cytoplasmic phosphoprotein. The gene is located on the
long arm of human chromosome 11 (11p13), encoding a 709-amino acid
protein, with a molecular weight of 116 kDa. Caprin-1 is part of
the conserved caprin family, which consists of two members,
caprin-1 and caprin-2, both of which contain two highly conserved
regions, homologous region-1 and −2 (21). High expression of caprin-1 was
initially identified in dividing cells of the thymus, and was also
known to be upregulated in activated T or B lymphocytes, and
hematopoietic progenitors. Caprin-1 expression has been reported to
be low in slowly dividing cells, such as those of the kidney or
muscles, but high levels have been detected in the brain (21). Caprin-1 is considered to be an RBP,
as it possesses RNA binding characteristics, i.e., the
arginine-glycine-glycine (RGG) motif and the RG enrichment region
(18). Using RNA-sequencing
technology, 6,064 mRNAs (>1,000 reads) were identified as
caprin-1 binding targets in the CD3 and CD28 antibody-activated
human T lymphoma cell line (Jurkat). Table I presents a partial list of caprin-1
target mRNAs divided into six categories according to their
biological function: Cell structure, RNA metabolism, RNA
translation, signaling transduction, ubiquitylation, and growth
factors and growth factor receptors (Wang et al, unpublished
results). Caprin-1 can affect cell survival and growth through
selectively binding a variety of mRNAs that are involved in cell
growth, differentiation and migration, including c-Myc and
cyclin-D2 (20,22–25).
c-Myc serves a central role in the transition from the
G1 to the S phase of the cell cycle, and cyclin D2
functions as a regulatory subunit of the kinases CDK4 or CDK6,
whose activity is required for the G1 to the S phase
transition (26,27). Cells without caprin-1 expression
exhibit delays in the transition from the G1 to the S
phase of the cell cycle (17).
Therefore, it is speculated that caprin-1 may serve an important
role in tumorigenesis.
 | Table I.Caprin-1-targeted mRNAs in activated
Jurkat cellsa. |
Table I.
Caprin-1-targeted mRNAs in activated
Jurkat cellsa.
| Function | Gene | Gene ID | Fold
enrichment |
|---|
| Cytoskeleton | Myosin heavy
chain | MYH9 | 10.60 |
|
| Talin-1 | TLN1 | 14.97 |
|
| Spectrin β
chain | SPTBN2 | 11.95 |
|
| Filamin-B | FLNB | 9.87 |
| Receptors | Interleukin-17
receptor A | IL17RA | 16.84 |
|
| Tetraspanin-27 | CD82 | 10.82 |
|
|
Interlekin-27-α | IL27RA | 9.04 |
|
| Fibroblast growth
factor receptor 4 | FGFR4 | 7.75 |
| Signaling | T-cell activation
NFKB-like protein | NP_640332 | 15.56 |
|
| TNFR1-associated
DEATH domain protein | TRADD | 11.07 |
|
| Phospholipase
C-β3 | PLCB3 | 10.81 |
|
| Protein phosphatase
1 regulatory 14B p21ras, PI3-kinase, MAPK | PPP1R114B | 9.48 |
| RNA
translation/metabolism | Eukaryotic
translation factor 4γ1 | eIF4-G1 | 10.83 |
|
| Pre-mRNA processing
splicing factor 8 | PRPFS | 8.89 |
|
| 40S ribosomal
protein S19-binding protein 1 | RPS19BP1 | 8.69 |
|
|
Polyadenylate-binding protein 1 G3BP1,
FXR1, FXR2 | PABPC1 | 5.02 |
| Growth
factor-associated | Wnt-8b | WNT8B | 15.56 |
|
| Fibroblast growth
factor-binding protein 3 | FGFBP3 | 12.21 |
|
| Insulin-like growth
factor II receptor | IGF2R | 8.45 |
Caprin-1 and cancer
Caprin-1 selectively binds to c-Myc and cyclin D2
mRNAs, which accelerates cell progression through the G1
phase into the S phase, enhances cell viability and promotes cell
growth, indicating that it may serve an important role in
tumorigenesis (17). This hypothesis
is supported by increasing experimental and clinical evidence.
Caprin-1 and associated regulatory factors (including miRNAs) are
abnormally expressed in tumor tissues and tumor cell lines. For
instance, miR-1 (24) and miR-223
(22) were revealed to be
downregulated in renal carcinoma cells and breast cancer cells,
respectively, resulting in a significant increase in the expression
level of caprin-1. The overexpression of caprin-1 may contribute to
the growth and invasion of tumor cells. Abnormal expression of
caprin-1 was also observed in patients with osteosarcoma and in
osteosarcoma cell lines (23).
Increased expression of caprin-1 can promote tumor growth and lung
metastasis from primary tumors, and shorten survival time (23). Tylophorine directly binds with
caprin-1 and accelerates degradation of the RNP formed by caprin-1
and RasGAP SH3-domain-binding protein (G3BP), downregulates c-Myc
and cyclins D1/D2, and ultimately inhibits the growth of the tumor
(20). Our recent studies revealed
the high expression of caprin-1 in clinical specimens from
glioblastoma patients. The degree of caprin-1 expression was
associated with clinical classification (28). Recently, a study by Casey et
al (29) that focused on c-Myc,
indirectly identified caprin-1 to be associated with certain immune
checkpoint proteins. c-Myc has been demonstrated to directly bind
to the promoters of two immune checkpoint protein genes, the innate
immune regulator CD47 and the adaptive immune checkpoint protein
programmed death-ligand 1 (PD-L1), thus regulating their expression
on the surface of tumor cells.
Caprin-1 may function as a complex with
other RBPs
In biological microenvironments, protein molecules
do not act alone; they function through intracellular networks
formed by interactions with other proteins. According to published
experimental results, caprin-1 can directly bind with G3BP1
(18), fragile X mental retardation
protein (FMRP) (30) and Japanese
encephalitis virus core protein (31). G3BP is a classic RBP, containing one
nuclear transport factor 2-like function domain (NTF2), one acidic
amino acid region, one RBD and one C-terminal RGG/G-enriched
sequence. The direct combination of G3BP1 and caprin-1 is formed
through the NTF2 domain of G3BP1 interacting with amino acids
352–380 of caprin-1 (18). The RBD
region and RGG/G-enriched sequence can selectively bind mRNA. FMRP
is another important RBP, and its deficiency can induce fragile X
syndrome (32). FMRP contains two
Agenet-like functional regions, two heterogeneous nuclear
ribonucleoprotein K homology functional regions and one RGG region.
The FMRP 427–442 amino acid sequence can interact with the caprin-1
231–245 amino acid sequence (30),
while the Agenet-like region, KH functional region and RGG region
have the mRNA binding function. The existence of the
caprin-1/G3BP1/FMRP-containing RNP was further confirmed by the
recent crystallographic study by Wu et al (33). Crystal structures of a fragment of
caprin-1 (residues 132–251) revealed that tight homodimerization
can be formed through the combination of the HR1 regions of two
molecules of caprin-1, creating a large and highly negatively
charged concave surface, which acts as a protein-binding groove.
This dimeric caprin-1 structure can be used as a scaffold to
further combine with G3BP and FMRP to form a macromolecule polymer
platform for more complex functions. Theoretically, two molecules
of caprin-1 can combine with two molecules of G3BP1 and two
molecules of FMRP. The resulting macromolecule polymer does not
affect the integrity of the RNA-binding region of each component or
their ability to bind mRNAs. Further precise research on the
caprin-1/G3BP1/FMRP complex may have great potential to aid in
developing novel and more effective anticancer drugs.
Caprin-1 may initiate carcinogenesis via
stress granules
With increasing experimental evidence, the
importance of SG in cancer has been awarded unprecedented
attention. Our previous study demonstrated the presence of caprin-1
in RNA transport granules and stress granules (SGs) (18). These results indicated that caprin-1
may be involved in carcinogenesis through the formation of SGs. As
illustrated in Fig. 1, there are
numerous types of RNA-containing granule in the cell cytoplasm,
including RNA transport granules, SGs and processing bodies
(P-bodies). RNA transport granules are responsible for transporting
mRNA to specific subcellular structures. For example, actin and
microtubules are located at the leading edge of the cell membrane
in the direction of fibroblast and tumor cell movement, which is
closely associated with the invasive growth and metastasis of tumor
cells. Caprin-1 is present in RNA transport granules (18), which are located at tubulin-enriched
sites, such as mid-bodies, actin-enriched podosomes (34,35), the
leading edge membrane of moving T lymphocytes (Wang et al,
unpublished data) and migrating fibroblasts (36). Caprin-1 and G3BP1 can combine with
FMRP and coexist in RNA transport granules. P-bodies are the site
of RNA degradation; they contain multiple molecules that are
responsible for mRNA degradation, surveillance, translation
inhibition and RNA-mediated gene suppression. P-bodies also include
marker proteins decapping mRNA, RNA degradation enzyme 1 and RNA
degradation enzyme 2, GW182 (another RBP), argonaute 2, RNA-induced
silencing complex and miRNAs. SGs are formed in response to cells
undergoing different stress stimuli and are a conserved mechanism
to reduce stress-associated damage and promote cell survival. SGs
contain translationally stalled mRNAs, translation initiation
factors, 40S and 60S ribosomal subunits, translation of suspended
enzymes, specific RBPs and signaling molecules (37,38). The
majority of classic SGs are produced by stress-induced
phosphorylation of eukaryotic initiation factor 2α (eIF2α).
Caprin-1, G3BP, T-cell-restricted intracellular antigen-1 (TIA-1),
TIA-1-related protein, tristetraprolin (TTP) and FMRP can all
induce the biogenesis of SGs. Caprin-1 combines with G3BP1 and/or
TIA-1 and coexists in the classic SGs. However, high expression of
caprin-1, G3BP, TIA-1, TTP or FMRP alone can induce the formation
of SGs, even when there is no stress stimulus signal (18,39–43). SGs
function to preserve RNAs against harmful conditions; they also
serve a decisive role in facilitating further storage, translation
reprogramming or degradation of untranslated mRNAs (38,44,45). The
association between RNA granules and caprin-1-associated
tumorigenesis is illustrated in Fig.
1.
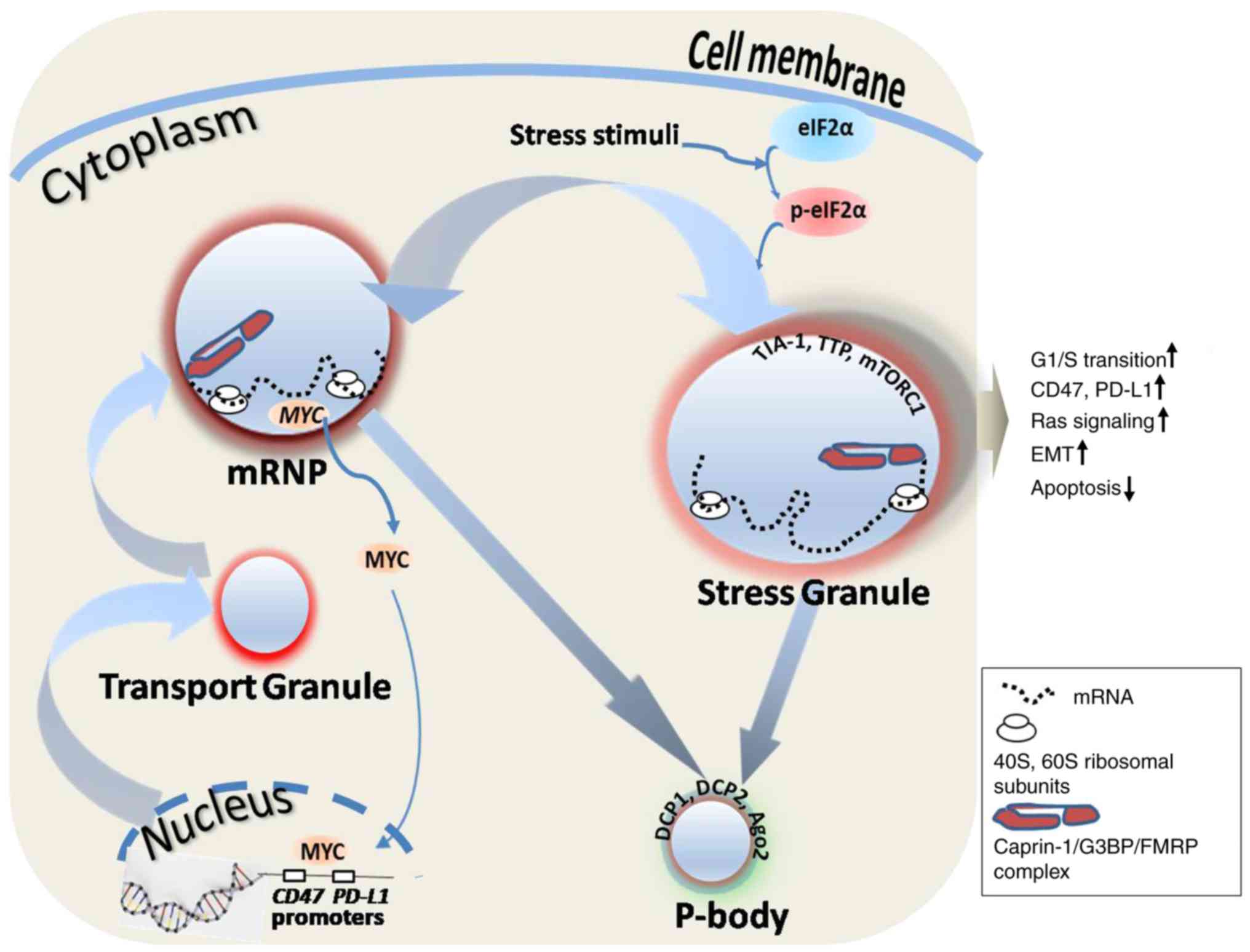 | Figure 1.RNA granules and caprin-1-associated
tumorigenesis. Caprin-1 mRNA is produced in the nucleus and
transported into mRNP or the SG through the transport granule, and
finally degraded in the P-body. Caprin-1-associated tumorigenesis
takes place primarily through affecting SG-mediated tumorigenic
characteristics and the MYC-mediated immune checkpoint. eIF2α can
be activated by various stress stimuli and induces SG formation,
thereby leading to tumorigenic characteristics, such as increased
transition from the G1 to the S phase of the cell cycle,
increased Ras signaling pathway, EMT and decreased apoptosis.
Caprin-1 can also indirectly enhance the expression of immune
checkpoint proteins, such as CD47 and PD-L1, through MYC, which is
one of the regulatory targets of caprin-1. Caprin-1, cytoplasmic
activation/proliferation-associated protein-1; mRNP, mRNA protein
complexes; SG, stress granule; P-body, processing body; eIF2α,
eukaryotic translation initiation factor 2α; EMT, epithelial to
mesenchymal transition; PD-L1, programmed death-ligand 1; TIA-1,
T-cell-restricted intracellular antigen-1; TTP, tristetraprolin;
mTORC1, mammalian target of rapamycin complex 1; DCP, decapping
enzyme; Ago2, argonaute 2; G3BP, GTPase-activating protein (Src
homology 3 domain) binding protein; FMRP, fragile X mental
retardation 1. |
SG formation is part of the integrated stress
response, which is a process that receives information from
different ‘stress sensors’ and acts with cells to adapt to stress
stimuli (46). These sensors are
usually kinases that can phosphorylate serine/threonine residues of
eIF2α. The tumor microenvironment is characterized by tissue
hypoxia, high levels of reactive oxygen species and insufficient
nutrients. These conditions can activate the cell stress response
and promote SG formation, inducing tumor cells to adapt to the
environment quickly and to alter metabolic pathways, thus
maintaining their rapid growth. The existence of this mechanism has
been validated in various clinical investigations. The presence of
SGs has been verified in numerous types of human cancer, including
glioblastoma, hepatocellular carcinoma and osteosarcoma (47–50). In
addition, radiation therapy and chemotherapy can stimulate tumor
cells to form SGs, which can result in resistance to treatment
(46). A variety of approved
chemotherapeutic agents have been indicated to induce SG, resulting
in reduced therapeutic efficacy (46,51).
Therefore, it can be hypothesized that blocking SG production may
effectively kill tumor cells. Experimental evidence supports the
hypothesis that the inhibition of SG formation can enhance cell
mortality and inhibit tumor growth (52).
In addition to roles in cell mortality and
tumorigenesis, SGs are also associated with tumor metastasis. In
tumor xenograft animal experiments, SG-producing osteosarcoma cells
were more resistant to treatment and were able to migrate to the
lungs with greater ease, while inhibiting SG formation via G3BP1
limited tumor invasion and prevented metastasis in vivo
(49). Dissemination of tumor cells
can enhance metastasis. Bone marrow-derived tumor cells from
patients with breast cancer have been revealed to contain SGs, and
these SGs contribute to cell survival of disseminated tumor cells
(45).
Clinical perspectives on caprin-1
As demonstrated in Table
II, caprin-1 has been reported to be involved in almost all
types of human malignancy, although the majority of the evidence
obtained thus far comes from in vitro settings. Given its
pivotal role in the PTGR of cell cycle control-associated genes,
caprin-1 and/or caprin-1-containing complex act as an amplifier to
drive tumorigenesis (17,18). The clinical significance of caprin-1
will be elucidated along with the progress of the corresponding
investigations.
| Table II.Experimental evidence of
caprin-1-associated human malignancies. |
Table II.
Experimental evidence of
caprin-1-associated human malignancies.
| Type of cancer | Type of
evidence | (Refs.) |
|---|
| Hepatocellular | Tissue microarray
and IHC of specimens from 239 patients | (53) |
|
| Human HepG2 and
Hep3B cell lines | (20,47) |
| Osteosarcoma | Tissue microarray
and IHC of specimens from 59 patients | (23) |
|
| Human SaOS-2 and
U2OS xenograft model | (23,49) |
| Breast | PET imaging study
of SaOS-2/Caprin-1 vs. 143B model | (54) |
| Melanoma | Human MCF-7, T-47D
and MDA-MB-231 vs. MCF-10A | (20,22,47) |
|
| Human BT-474 and
MDA-MB-453 ×enograft model | (45) |
|
| Proteomic analysis
of human BLM melanoma cells | (55) |
| Glioblastoma | Human C6 and U87MG
glioma cell lines | (48) |
| Prostate | Human PC-3 and
LnCaP cell lines | (20,47) |
| Cervical | Human HeLa cell
line | (20,47) |
| Gastric | Tissue microarray
and IHC of specimens from 262 patients | (56) |
|
| Human NUGC-3,
HGC-27 and MGC80-3 cell lines | (20,56) |
| Pancreatic | Human H1299
×enograft model | (57) |
| Colon | Human PANC-1,
BxPC3, S2-013, SUIT-2, COLO357, HPAF, MIA-Paca2, Capan2 and AsPC-1
cell lines | (20,58) |
| Leukemia | Human DLD-1, SW480
and HCT116 ×enograft model | (20,59) |
|
| Human U-937 and
human leukemic Jurkat T cells | (20) |
Although being able to use caprin-1 as a therapeutic
target in clinical practice may be a distant prospect, there are a
number of established associations between caprin-1 and clinical
cancer science: i) Caprin-1 acts as a biomarker for the clinical
diagnosis of certain malignancies (53,54).
Considering the close association between the expression level of
caprin-1 and tumorigenesis, high expression levels of caprin-1 can
be used as an indicator of certain malignancies. Caprin-1 mRNA
and/or protein levels can be determined using molecular biology and
liquid biopsy techniques in tissue or peripheral blood samples
(28,55). ii) The degree of caprin-1 expression
can be used to predict the prognosis of malignancies (23). iii) Caprin-1 and complexes can be
used as a target for the development of cancer therapeutic drugs,
such as specific short hairpin RNAs to knockdown caprin-1
expression, or specific peptides to block the formation of
caprin-1/G3BP1/FMRP complexes (20,56–59).
Conclusions
Caprin-1 forms part of RNP complexes and directly
interacts with mRNAs, and is therefore defined as an RBP. Caprin-1
acts alone or combines with other RBPs to participate in the
regulation of cell cycle control-associated gene expression.
Caprin-1 selectively interacts with c-Myc and cyclin D1/D2 mRNAs
activating cell proliferation and upregulating the expression of
immune checkpoint proteins CD47 and PD-L1. Through the formation of
SGs, caprin-1 is also involved in the process of tumor cell
adaption to adverse conditions, such as radiation and chemotherapy.
Given its role in various clinical malignancies, caprin-1 holds the
potential to be used as a biomarker and target for the development
of novel therapeutics.
Acknowledgements
Not applicable.
Funding
This study was funded by the Key Projects of
Precision Medicine (grant no. 2016JZ01) and the Education
Department of Hubei Province (grant no. Q20162102).
Availability of data and material
The datasets used/analyzed in the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
ZSY, HQ, HG, JL and BW collected data. LJD and BW
contributed to the writing of the manuscript. All the authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
caprin-1
|
cytoplasmic
activation/proliferation-associated protein-1
|
|
eIF2α
|
eukaryotic initiation factor 2α
|
|
FMRP
|
fragile X mental retardation
protein
|
|
G3BP1
|
RasGAP-SH3-domain binding protein
1
|
|
miRNA
|
microRNA
|
|
ncRNA
|
non-coding RNA
|
|
NTF2
|
nuclear transport factor 2
|
|
PD-L1
|
programmed death-ligand 1
|
|
PTGR
|
post-transcriptional gene
regulation
|
|
RBD
|
RNA-binding domain
|
|
RBP
|
RNA-binding protein
|
|
RNP
|
ribonucleoprotein
|
|
SGs
|
stress granules
|
|
TIA-1
|
T-cell-restricted intracellular
antigen-1
|
References
|
1
|
Gerstberger S, Hafner M and Tuschl T: A
census of human RNA-binding proteins. Nat Rev Genet. 15:829–845.
2014. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gerstberger S, Hafner M, Ascano M and
Tuschl T: Evolutionary conservation and expression of human
RNA-binding proteins and their role in human genetic disease. Adv
Exp Med Biol. 825:1–55. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mori T, Ngouv H, Hayashida M, Akutsu T and
Nacher JC: ncRNA-disease association prediction based on sequence
information and tripartite network. BMC Syst Biol. 12 (Suppl
1):S372018. View Article : Google Scholar
|
|
4
|
Hudson WH and Ortlund EA: The structure,
function and evolution of proteins that bind DNA and RNA. Nat Rev
Mol Cell Biol. 15:749–760. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Keene JD: RNA regulons: Coordination of
post-transcriptional events. Nat Rev Genet. 8:533–543. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nishida K, Kuwano Y, Nishikawa T, Masuda K
and Rokutan K: RNA binding proteins and genome integrity. Int J Mol
Sci. 18:E13412017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kai M: Roles of RNA-binding proteins in
DNA damage response. Int J Mol Sci. 17:3102016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Harvey R, Dezi V, Pizzinga M and Willis
AE: Post-transcriptional control of gene expression following
stress: The role of RNA-binding proteins. Biochem Soc Trans.
45:1007–1014. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sheinberger J and Shav-Tal Y: mRNPs meet
stress granules. FEBS Lett. 591:2534–2542. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Maziuk B, Balance HI and Wolozin B:
Dysregulation of RNA binding protein aggregation in
neurodegenerative disorders. Front Mol Neurosci. 10:892017.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Geuens T, Bouhy D and Timmerman V: The
hnRNP family: Insights into their role in health and disease. Hum
Genet. 135:851–867. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wurth L and Gebauer F: RNA-binding
proteins, multifaceted translational regulators in cancer. Biochim
Biophys Acta. 1849:881–886. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pereira B, Billaud M and Almeida R:
RNA-binding proteins in cancer: Old players and new actors. Trends
Cancer. 3:506–528. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Paronetto MP, Cappellari M, Busa R,
Pedrotti S, Vitali R, Comstock C, Hyslop T, Knudsen KE and Sette C:
Alternative splicing of the cyclin D1 proto-oncogene is regulated
by the RNA-binding protein Sam68. Cancer Res. 70:229–239. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hsieh AC and Ruggero D: Targeting
eukaryotic translation initiation factor 4E (eIF4E) in cancer. Clin
Cancer Res. 16:4914–4920. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Abdelmohsen K and Gorospe M:
Posttranscriptional regulation of cancer traits by HuR. Wiley
Interdiscip Rev RNA. 1:214–229. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang B, David MD and Schrader JW: Absence
of caprin-1 results in defects in cellular proliferation. J
Immunol. 175:4274–4282. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Solomon S, Xu Y, Wang B, David MD,
Schubert P, Kennedy D and Schrader JW: Distinct structural features
of caprin-1 mediate its interaction with G3BP-1 and its induction
of phosphorylation of eukaryotic translation initiation factor
2alpha, entry to cytoplasmic stress granules, and selective
interaction with a subset of mRNAs. Mol Cell Biol. 27:2324–2342.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Reich J and Papoulas O: Caprin controls
follicle stem cell fate in the Drosophila ovary. PLoS One.
7:e353652012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Qiu YQ, Yang CW, Lee YZ, Yang RB, Lee CH,
Hsu HY, Chang CC and Lee SJ: Targeting a ribonucleoprotein complex
containing the caprin-1 protein and the c-Myc mRNA suppresses tumor
growth in mice: An identification of a novel oncotarget.
Oncotarget. 6:2148–2163. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Grill B, Wilson GM, Zhang KX, Wang B,
Doyonnas R, Quadroni M and Schrader JW: Activation/division of
lymphocytes results in increased levels of cytoplasmic
activation/proliferation-associated protein-1: Prototype of a new
family of proteins. J Immunol. 172:2389–2400. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gong B, Hu H, Chen J, Cao S, Yu J, Xue J,
Chen F, Cai Y, He H and Zhang L: Caprin-1 is a novel microRNA-223
target for regulating the proliferation and invasion of human
breast cancer cells. Biomed Pharmacother. 67:629–636. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sabile AA, Arlt MJ, Muff R, Husmann K,
Hess D, Bertz J, Langsam B, Aemisegger C, Ziegler U, Born W and
Fuchs B: Caprin-1, a novel Cyr61-interacting protein, promotes
osteosarcoma tumor growth and lung metastasis in mice. Biochim
Biophys Acta. 1832:1173–1182. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xiao H, Zeng J, Li H, Chen K, Yu G, Hu J,
Tang K, Zhou H, Huang Q, Li A, et al: MiR-1 downregulation
correlates with poor survival in clear cell renal cell carcinoma
where it interferes with cell cycle regulation and metastasis.
Oncotarget. 6:13201–13215. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Teng Y, Ren Y, Hu X, Mu J, Samykutty A,
Zhuang X, Deng Z, Kumar A, Zhang L, Merchant ML, et al:
MVP-mediated exosomal sorting of miR-193a promotes colon cancer
progression. Nat Commun. 8:144482017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Matsumura I, Tanaka H and Kanakura Y: E2F1
and c-Myc in cell growth and death. Cell Cycle. 2:333–338. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pardee AB: G1 events and regulation of
cell proliferation. Science. 246:603–608. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang L, Gui H, Tang XJ, Yang ZS, Zou DD,
Lu JT, Yan LD, Dai LJ, Luo J and Wang B: Expression and
tumor-Promoting effects of caprin-1 in human glioma. Glioma.
1:136–141. 2018. View Article : Google Scholar
|
|
29
|
Casey SC, Tong L, Li Y, Do R, Walz S,
Fitzgerald KN, Gouw AM, Baylot V, Gütgemann I, Eilers M and Felsher
DW: MYC regulates the antitumor immune response through CD47 and
PD-L1. Science. 352:227–231. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
El Fatimy R, Tremblay S, Dury AY, Solomon
S, De Koninck P, Schrader JW and Khandjian EW: Fragile X mental
retardation protein interacts with the RNA-binding protein caprin1
in neuronal ribonucleoprotein complexes [corrected]. PLoS One.
7:e393382012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Katoh H, Okamoto T, Fukuhara T, Kambara H,
Morita E, Mori Y, Kamitani W and Matsuura Y: Japanese encephalitis
virus core protein inhibits stress granule formation through an
interaction with Caprin-1 and facilitates viral propagation. J
Virol. 87:489–502. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jin P, Zarnescu DC, Ceman S, Nakamoto M,
Mowrey J, Jongens TA, Nelson DL, Moses K and Warren ST: Biochemical
and genetic interaction between the fragile X mental retardation
protein and the microRNA pathway. Nat Neurosci. 7:113–117. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wu Y, Zhu J, Huang X and Du Z: Crystal
structure of a dimerization domain of human Caprin-1: Insights into
the assembly of an evolutionarily conserved ribonucleoprotein
complex consisting of Caprin-1, FMRP and G3BP1. Acta Crystallogr D
Struct Biol. 72:718–727. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mingle LA, Okuhama NN, Shi J, Singer RH,
Condeelis J and Liu G: Localization of all seven messenger RNAs for
the actin-polymerization nucleator Arp2/3 complex in the
protrusions of fibroblasts. J Cell Sci. 118:2425–2433. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Carson JH and Barbarese E: Systems
analysis of RNA trafficking in neural cells. Biol Cell. 97:51–62.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Copsey AC, Cooper S, Parker R, Lineham E,
Lapworth C, Jallad D, Sweet S and Morley SJ: The helicase, DDX3X,
interacts with poly(A)-binding protein 1 (PABP1) and caprin-1 at
the leading edge of migrating fibroblasts and is required for
efficient cell spreading. Biochem J. 474:3109–3120. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Thomas MG, Loschi M, Desbats MA and
Boccaccio GL: RNA granules: The good, the bad and the ugly. Cell
Signal. 23:324–334. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kedersha N, Ivanov P and Anderson P:
Stress granules and cell signaling: More than just a passing phase?
Trends Biochem Sci. 38:494–506. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gilks N, Kedersha N, Ayodele M, Shen L,
Stoecklin G, Dember LM and Anderson P: Stress granule assembly is
mediated by prion-like aggregation of TIA-1. Mol Biol Cell.
15:5383–5398. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Tourriere H, Chebli K, Zekri L, Courselaud
B, Blanchard JM, Bertrand E and Tazi J: The RasGAP-associated
endoribonuclease G3BP assembles stress granules. J Cell Biol.
160:823–831. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kedersha N, Cho MR, Li W, Yacono PW, Chen
S, Gilks N, Golan DE and Anderson P: Dynamic shuttling of TIA-1
accompanies the recruitment of mRNA to mammalian stress granules. J
Cell Biol. 151:1257–1268. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Mazroui R, Huot ME, Tremblay S, Filion C,
Labelle Y and Khandjian EW: Trapping of messenger RNA by fragile X
mental retardation protein into cytoplasmic granules induces
translation repression. Hum Mol Genet. 11:3007–3017. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Stoecklin G, Stubbs T, Kedersha N, Wax S,
Rigby WF, Blackwell TK and Anderson P: MK2-induced
tristetraprolin:14-3-3 complexes prevent stress granule association
and ARE-mRNA decay. EMBO J. 23:1313–1324. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Buchan JR and Parker R: Eukaryotic stress
granules: The ins and outs of translation. Mol Cell. 36:932–941.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gupta N, Badeaux M, Liu Y, Naxerova K,
Sgroi D, Munn LL, Jain RK and Garkavtsev I: Stress
granule-associated protein G3BP2 regulates breast tumor initiation.
Proc Natl Acad Sci USA. 114:1033–1038. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Anderson P, Kedersha N and Ivanov P:
Stress granules, P-bodies and cancer. Biochim Biophys Acta.
1849:861–870. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Adjibade P, St-Sauveur VG, Quevillon
Huberdeau M, Fournier MJ, Savard A, Coudert L, Khandjian EW and
Mazroui R: Sorafenib, a multikinase inhibitor, induces formation of
stress granules in hepatocarcinoma cells. Oncotarget.
6:43927–43943. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Vilas-Boas Fde A, da Silva AM, de Sousa
LP, Lima KM, Vago JP, Bittencourt LF, Dantas AE, Gomes DA, Vilela
MC, Teixeira MM and Barcelos LS: Impairment of stress granule
assembly via inhibition of the eIF2alpha phosphorylation sensitizes
glioma cells to chemotherapeutic agents. J Neurooncol. 127:253–260.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Somasekharan SP, El-Naggar A, Leprivier G,
Cheng H, Hajee S, Grunewald TG, Zhang F, Ng T, Delattre O,
Evdokimova V, et al: YB-1 regulates stress granule formation and
tumor progression by translationally activating G3BP1. J Cell Biol.
208:913–929. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Leprivier G, Rotblat B, Khan D, Jan E and
Sorensen PH: Stress-mediated translational control in cancer cells.
Biochim Biophys Acta. 1849:845–860. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Szaflarski W, Fay MM, Kedersha N, Zabel M,
Anderson P and Ivanov P: Vinca alkaloid drugs promote
stress-induced translational repression and stress granule
formation. Oncotarget. 7:30307–30322. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Fournier MJ, Coudert L, Mellaoui S,
Adjibade P, Gareau C, Côté MF, Sonenberg N, Gaudreault RC and
Mazroui R: Inactivation of the mTORC1-eukaryotic translation
initiation factor 4E pathway alters stress granule formation. Mol
Cell Biol. 33:2285–2301. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Tan N, Dai L, Liu X, Pan G, Chen H, Huang
J and Xu Q: Upregulation of caprin1 expression is associated with
poor prognosis in hepatocellular carcinoma. Pathol Res Pract.
213:1563–1567. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Campanile C, Arlt MJ, Kramer SD, Honer M,
Gvozdenovic A, Brennecke P, Fischer CR, Sabile AA, Müller A,
Ametamey SM, et al: Characterization of different osteosarcoma
phenotypes by PET imaging in preclinical animal models. J Nucl Med.
54:1362–1368. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Kedracka-Krok S, Jankowska U, Elas M, Sowa
U, Swakon J, Cierniak A, Olko P, Romanowska-Dixon B and Urbanska K:
Proteomic analysis of proton beam irradiated human melanoma cells.
PLoS One. 9:e846212014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Min L, Ruan Y, Shen Z, Jia D, Wang X, Zhao
J, Sun Y and Gu J: Overexpression of Ras-GTPase-activating protein
SH3 domain-binding protein 1 correlates with poor prognosis in
gastric cancer patients. Histopathology. 67:677–688. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhang H, Zhang SH, He HW, Zhang CX, Yu DK
and Shao RG: Downregulation of G3BPs inhibits the growth, migration
and invasion of human lung carcinoma H1299 cells by suppressing the
Src/FAK-associated signaling pathway. Cancer Gene Ther. 20:622–629.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Taniuchi K, Nishimori I and Hollingsworth
MA: The N-terminal domain of G3BP enhances cell motility and
invasion by posttranscriptional regulation of BART. Mol Cancer Res.
9:856–866. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Zhang H, Zhang S, He H, Zhao W, Chen J and
Shao RG: GAP161 targets and downregulates G3BP to suppress cell
growth and potentiate cisplaitin-mediated cytotoxicity to colon
carcinoma HCT116 cells. Cancer Sci. 103:1848–1856. 2012. View Article : Google Scholar : PubMed/NCBI
|














