Introduction
Over the past few decades, male infertility has
increased (1), a phenomenon that is
closely associated with environmental pollution (2). Particularly, cadmium is a highly toxic
heavy metal that has been reported to induce male infertility
(3,4). With the rise of the incidence rate of
infertility, the adverse effects of environmental factors are
becoming an increasing concern. Particularly, environmental
pollution has posed a serious threat in recent years. Guangxi is an
area with high cadmium, which has an adverse effect on the safety
of life (1,3,4).
Previous studies have demonstrated that the reproductive toxicity
of cadmium is the highest, particularly in the male reproductive
toxicity system (4–8). Human intake of cadmium primarily
originates from the consumption of contaminated water, crops,
cigarettes or other sources (9).
Cadmium deposition in the kidney, bones, liver, lung and
reproductive organs (10,11) leads to severe organ impairment
(12). Three mechanisms of
cadmium-induced cellular injury have been proposed (5,13–16). The
first concerns cadmium-calcium interactions, in which cadmium may
enter cells through calcium channels and compete with calcium to
bind calmodulin, interfering therefore with calmodulin and with
calmodulin-dependent physiological and biochemical processes
(1,3). The second mechanism involves protein
alterations. Particularly, cadmium may interact with the hydroxyl,
mercapto and amino groups of proteins to generate cadmium-protein
complexes, which can suppress or inactivate multiple enzyme
systems, which can have a deleterious impact on biological
activities (4). In the third
mechanism, cadmium can influence the expression of apoptotic genes
(5,17). Cadmium can induce abnormal gene
expression, inhibit DNA damage repair and cell apoptosis. Cadmium
exposure increases the mRNA expression level of the pro-apoptotic
Bax gene in the small intestine of rats and decreases the mRNA
expression level of anti-apoptotic Bcl-2 gene. Thereby, it
increases the Bax/Bcl-2 ratio and induces apoptosis of cells
(18,19). Testicular stromal cells are
essentially responsible for testosterone synthesis (20). Testosterone secreted by Leydig cells
is transported to target organs throughout the body via the blood
circulation, and promotes differentiation and development of
reproductive organs, development and maintenance of male secondary
sexual characteristics, and sperm by binding to androgen receptors.
It serves an important role in maintaining sexual function and
promoting metabolism, including promotion of protein synthesis,
bone growth and erythropoiesis (21,22).
Additionally, testosterone can enter Sertoli cells in a paracrine
manner, and is involved in the regulation of spermatogenesis. When
testosterone enters Sertoli cells, it serves the following four
functions during spermatogenesis: i) Maintenance of blood-testis
barrier; ii) regulation of meiosis; iii) maintenance of the
adhesion between spermatozoa and Sertoli cells; and iv) regulation
of the release of mature sperm (23,24).
Testosterone and androgen receptor serve an important role in male
reproduction. When the androgen receptor gene is knocked out,
testosterone secretion is reduced, testis is markedly atrophied,
spermatogenesis is blocked in pre-meiotic diploid phase and
spermatozoa are hardly detected in the epididymis. Infertility may
occur in mice, and serum testosterone concentrations in mice are
reduced when infertility occurs (25). Cholesterol undergoes a series of
reactions in testicular stromal cells to produce certain crucial
compounds that are associated with testosterone synthesis,
including luteinizing hormone receptor (LHR) and 17α-hydroxylase
(20). Furthermore, nitric oxide
(NO) is an important second messenger. The half-life of NO is very
short. Nitric oxide synthase (NOS) is a necessary enzyme in the
process of NO synthesis. Therefore, the concentration of NOS is
frequently used to indirectly represent the level of NO (26). This has been reported to affect
testosterone biogenesis and sperm production when expressed in
testicular stromal cells (27–31).
Previous studies have suggested that the content of NO in infertile
sperm is associated with the germ cell apoptosis rate (32,33).
These results also indirectly suggested that an increased amount of
NO may enhance the ability of antioxidant damage and fight against
the damage of harmful factors to germ cells or other cells in the
process of cadmium induced male infertility. Additionally,
excessive NO may be a prerequisite for oxidative stress, since
excessive NO causes semen nitrification stress reaction, and
eventually male infertility (32,34).
However, it remains unclear whether NO is involved in cadmium
toxicity in testicular stromal cells or whether its role is
associated with LHR and 17α-hydroxylase. Subsequently, the present
study aimed to investigate the mechanisms of cadmium-induced
reproductive toxicity.
Materials and methods
Materials, equipment and reagents
The present study used an upright light microscope
(Olympus Corporation) and a chemiluminescence imager (Tanon Science
and Technology Co., Ltd.). In addition, the current study utilized
various reagents as follows: LHR (Boster Biological Technology),
17α-hydroxylase (Affinity, Inc.) endothelial NOS (eNOS; Affinity,
Inc.), diaminobenzidine tetrahydrochloride (DAB) kit (Origene
Technologies, Inc.), streptavidin-biotin complex
immunohistochemical kit (Fuzhou Maixin Biotech Co., Ltd.), terminal
deoxynucleotidyl transferase dUTP nick end labeling (TUNEL)
apoptosis kit (Roche Diagnostics), mouse androgen ELISA kit (cat.
no. E03A0019; BlueGene Biotech Co., Ltd.) and cadmium chloride
(analytical purity; Sinopharm Chemical Reagent Co., Ltd.).
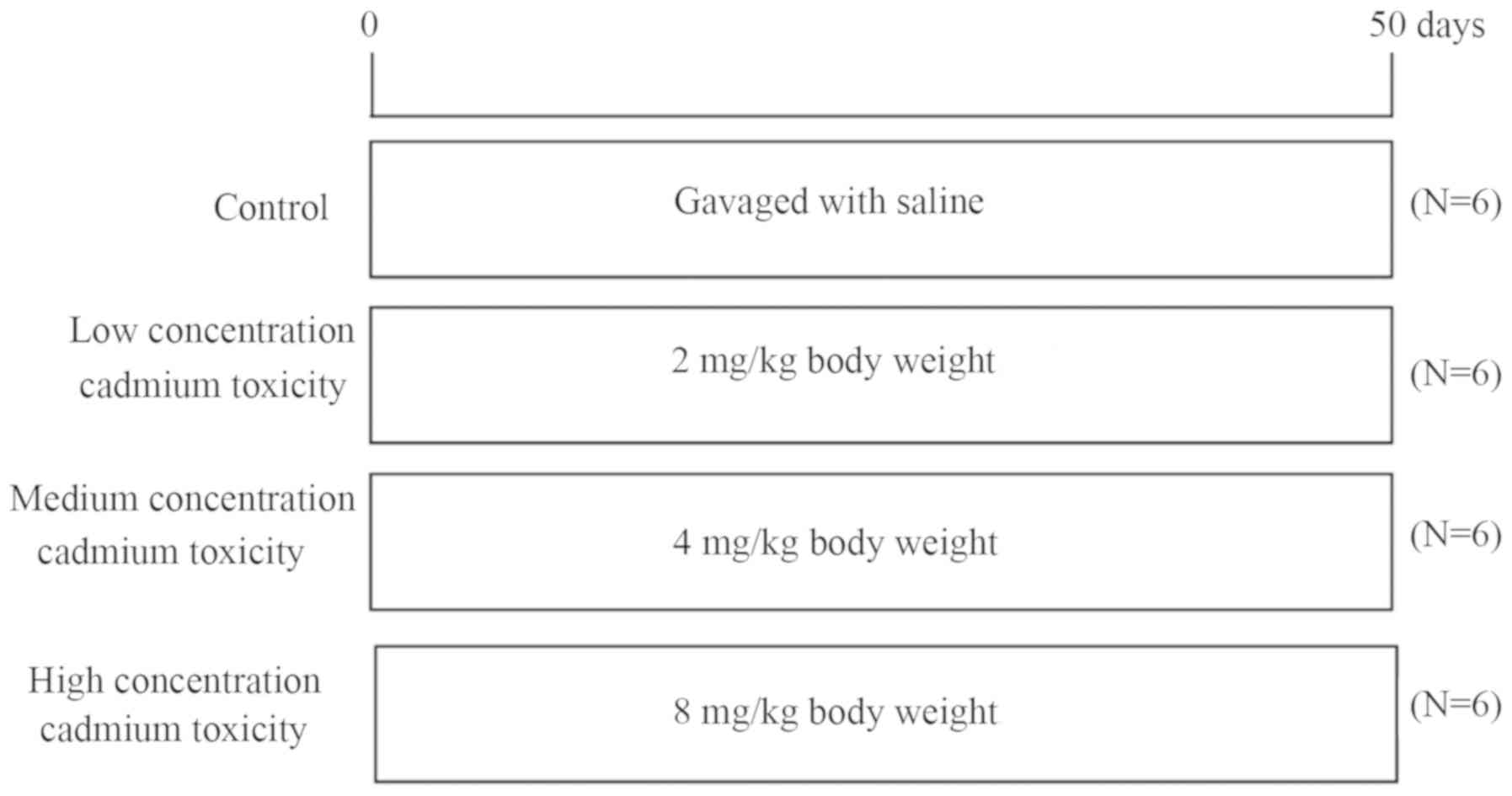
Animal grouping
A total of 24 Institute of Cancer Research male
specific pathogen-free mice with an average weight of 23 g were
obtained from Hunan SJA Laboratory Animal Co., Ltd. (Hunan, China)
and housed in the Animal Experimental Center of Guilin Medical
College. All experiments were approved by the Ethical Committees of
Guilin Medical University (Guilin, China). All mice were housed at
a controlled temperature of 25°C, 55–65% humidity and with a 12-h
light/dark cycle. The food and water provided were sterilized and
mice had free access to food and water. Mice were randomly divided
into four groups as determined by preliminary experiments (Fig. 1; n=6 animals/group), including the
normal control group (gavage-fed with saline), the
low-concentration cadmium toxicity group [fed with 2 mg/kg (BW)
cadmium], the medium-concentration cadmium toxicity group (fed with
4 mg/kg BW cadmium) and the high-concentration cadmium toxicity
group (fed with 8 mg/kg BW cadmium). Gavage was performed once per
day for 8 consecutive weeks and the volume administration each time
was ≤0.2 ml. Mice were subsequently sacrificed by cervical
dislocation and according to the guidelines given in ‘The
Laboratory Mouse’ (35). A midline
incision was made on the abdomen using a scalpel, and the organs
and testicular tissues were identified and removed. One half of the
tissue was paraffin embedded for immunohistochemistry and the other
half was frozen for subsequent protein analysis. In addition, blood
samples were taken to determine androgen levels.
Immunochemical detection
Testicular tissues were fixed with 10% neutral
formaldehyde for 48 h at room temperature and paraffin-embedded,
sectioned, heated to 60°C, dewaxed in xylene and rehydrated
(alcohol series, 100, 95, 85 and 75%). Subsequently, they were
washed in PBS for 5 min. Sections (4-µm thick) were subsequently
incubated with 3% hydrogen peroxide at room temperature for 30 min.
Sections were incubated with mouse monoclonal primary antibodies
against eNOS (cat. no. ab76198; 1:100; Abcam), LHR (cat. no.
RPU51114; 1:250; OriGene Technologies, Inc.) and 17α-hydroxylase
(cat. no. RPU51114; 1:250; Biomatik) overnight at 4°C. The sections
were then incubated with secondary antibodies [undiluted; cat. no.
PV-6000; Universal kit (mouse/rabbit polymer detection system);
cat. no. PV-6000; OriGene Technologies, Inc.] for 30 min at room
temperature. Sections were subsequently incubated with DAB and
stained with hematoxylin and eosin (H&E). Hematoxylin staining
was performed for 2 min at room temperature, and eosin staining was
performed for 30 sec at room temperature. The sections were
dehydrated, clarified and mounted. Images were captured with an
upright light microscope (magnification, ×400) and analyzed using
AxioVision software (version 4.8.2; Carl Zeiss AG). Two independent
pathologists evaluated the slides. The immunostaining intensity was
scored as 0, 1, 2 and 3 for negative, weak, moderate and strong
staining, respectively. In addition, immunoreactivity was scored as
follows: Strong (3), moderate
(2), mild (1) or negative (0) for >60, 21–60, 5–20
or <5% stained cells, respectively.
Western blotting
Proteins were extracted from testicular tissues
using RIPA buffer (Beijing Solarbio Science & Technology Co.,
Ltd.). Protein concentration was determined using a Bicinchoninic
Acid Protein Concentration Assay kit (Enhanced) purchased from
Beyotime Institute of Biotechnology (cat. no. P0010S). Proteins (20
µg/lane) were separated by 10% SDS-PAGE and transferred onto a
nitrocellulose membrane. Membranes were blocked in 5% non-fat milk
in TBS with 0.05% Tween-20 for 2 h at room temperature. Membranes
were incubated with mouse monoclonal primary antibodies against
eNOS, LHR and 17α-hydroxylase as aforementioned, and β-actin (cat.
no. TA-09; 1:1,000; OriGene Technologies, Inc.) at room temperature
for 2 h and 4°C overnight. Membranes were then incubated with the
goat anti-mouse IgG-horseradish peroxidase (HRP; cat. no. sc-2004;
1:5,000; Santa Cruz Biotechnology, Inc.) and goat anti-rabbit
IgG-HRP (cat. no. sc-2005; 1:2,000 Santa Cruz Biotechnology, Inc.)
secondary antibodies at 37°C for 1 h. Enhanced chemiluminescence
reagent (SuperSignal West Femto Substrate Trial kit; cat. no. D046;
Bridgen Biotech Co., Ltd.) was used to detect the signal on the
membrane and images were captured using the Bio-Rad Gel Image
Chemidoc™ XRS system (Bio-Rad Laboratories, Inc.). The data were
analyzed via densitometry using Bio-Rad Image Lab software (version
6.0.1; Bio-Rad Laboratories, Inc.) and normalized to the expression
of the internal control (β-actin).
Determination of testis cell apoptosis
with the TUNEL assay
Mice testis cell apoptosis was assessed by TUNEL
assay (Roche Diagnostics) (36).
Sections were fixed in 4% paraformaldehyde for 20 min at room
temperature. Paraffin-embedded sections were dewaxed in xylene and
rehydrated using an ethanol gradient (100, 100, 95, 85 and 75%).
Sections were incubated with proteinase K at 37°C for 30 min and
with 3% hydrogen peroxide at 37°C for 10 min and washed with PBS.
Section were incubated with 50 µl TUNEL assay substrate at 37°C in
the dark for 1 h. Sections were washed with PBS and incubated with
50 µl converter-POD (included in the TUNEL kit) at 37°C for 30 min.
Sections were then washed with PBS and treated with DAB for color
development. Sections were observed under an upright light
microscope (magnification, ×400) and particles with brown or yellow
nuclei were considered as positively stained and counted. At least
three fields were analyzed for each mouse. Sections were graded
according to the percentage of positive cells as follows: Strong
(3), moderate (2), mild (1)
or negative (0) for >60, 21–60, 5–20 or <5% stained
cells.
Determination of serum androgen levels
in mice
Following mice sacrifice, 0.8 ml blood was collected
from the heart. Blood was placed on ice for 30 min and centrifuged
at 1,000 g/min for 20 min. The resulting supernatant was obtained
and stored at −20°C. Androgen levels were determined using an ELISA
kit. The ELISA assay was conducted according to the manufacturer's
protocol. Briefly, the blood sample was first brought to room
temperature. Following standard product preparation and sample
addition, the antibody was added for 1 h at 37°C. Subsequent to
washing, A and B agent were gently agitated for blending, followed
by incubation at 37°C in the dark for 6 min. Termination solution
was added and measurements were taken sequentially at a wavelength
of 450 nm.
Statistical analysis
SPSS 18.0 software (SPSS, Inc.) was used for
statistical analysis. Data are expressed as the mean ± standard
deviation. One-way ANOVA followed by Fisher's least significant
difference test was used for comparison between the experimental
groups and the control group.
Results
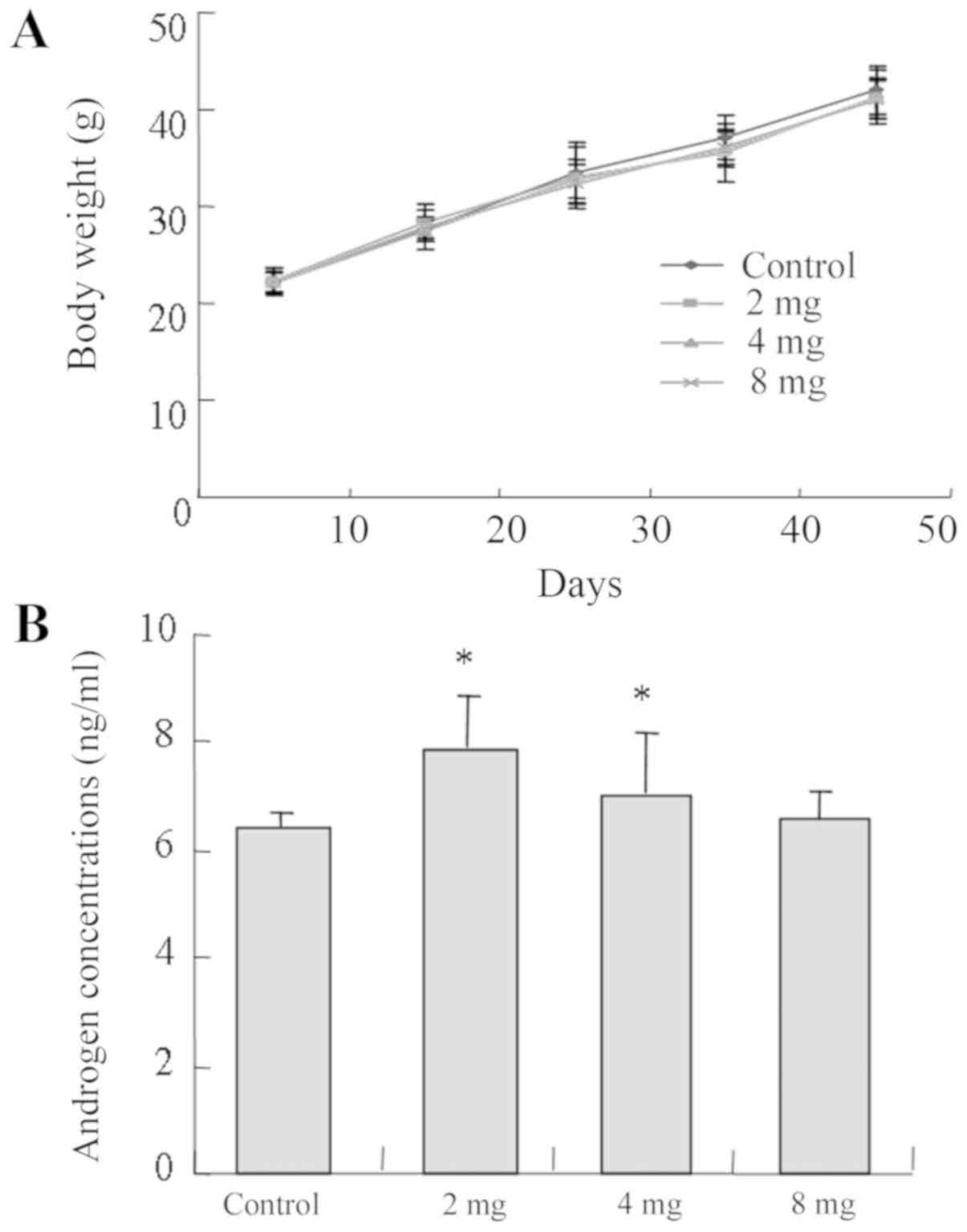
Effect of cadmium on mice weight and
serum androgen level
There was no significant difference identified
between the body weights of mice in the experimental and control
groups (P>0.05; Fig. 2A). In
addition, mice from the low and medium groups demonstrated higher
androgen concentrations compared with the control group and the
low-cadmium group (2 mg) presented the highest androgen
concentration (P<0.05; Fig.
2B).
Influence of cadmium on testicular
morphology and apoptosis
As presented in Fig.
3A, H&E staining demonstrated that mice from the control
group presented testicular morphological characteristics indicative
of sexual maturity as follows: i) Testicular seminiferous tubules
with thick walls; ii) spermatogenic cells and support cells at
various developmental stages with swirling contours and on
thickened spermatogenic epithelia; iii) sperm cells in the
seminiferous tubules; and iv) plump testicular stromal cells. By
contrast, mice in the low-cadmium group demonstrated decreased
thickness of the testicular seminiferous tubule walls, less
apparent swirling contours and no obvious changes in the appearance
of testicular stromal cells. However, sperm cells were observed on
the walls of the seminiferous tubules. Mice in the medium-cadmium
group presented the following features: i) A thin germinal
epithelium; ii) sporadic bleeding in the testicular stroma; iii)
cells with aberrant swirling contours; and iv) decreased
spermatogenesis. Mice from the high-cadmium group exhibited the
following features: i) A very thin germinal epithelium; ii)
seminiferous tubules with aberrant morphology; iii) a markedly low
level of normal spermatogenesis; iv) no swirling contours; v)
apparent abnormalities of the testicular stroma (stromal vacancy);
and vi) spots of bleeding. The expression levels of LHR and
17α-hydroxylase were higher in the low groups compared with in the
control groups (P<0.05), but less high in the high cadmium group
(P>0.05). eNOS and 17-α-hydroxylase exhibited consistent
alterations. The low-cadmium group and the medium-cadmium group
exhibited higher expression levels than the control group, while
the expression levels in the high-cadmium group were slightly
lower. eNOS expression was the highest in the medium-cadmium group
and relatively low in the low-cadmium group (Fig. 3A and C; Table I). In addition, the number of
TUNEL-positive nuclei significantly increased with cadmium in a
concentration-dependent manner, which suggested elevated apoptosis
rates (Fig. 3B and C; Table I).
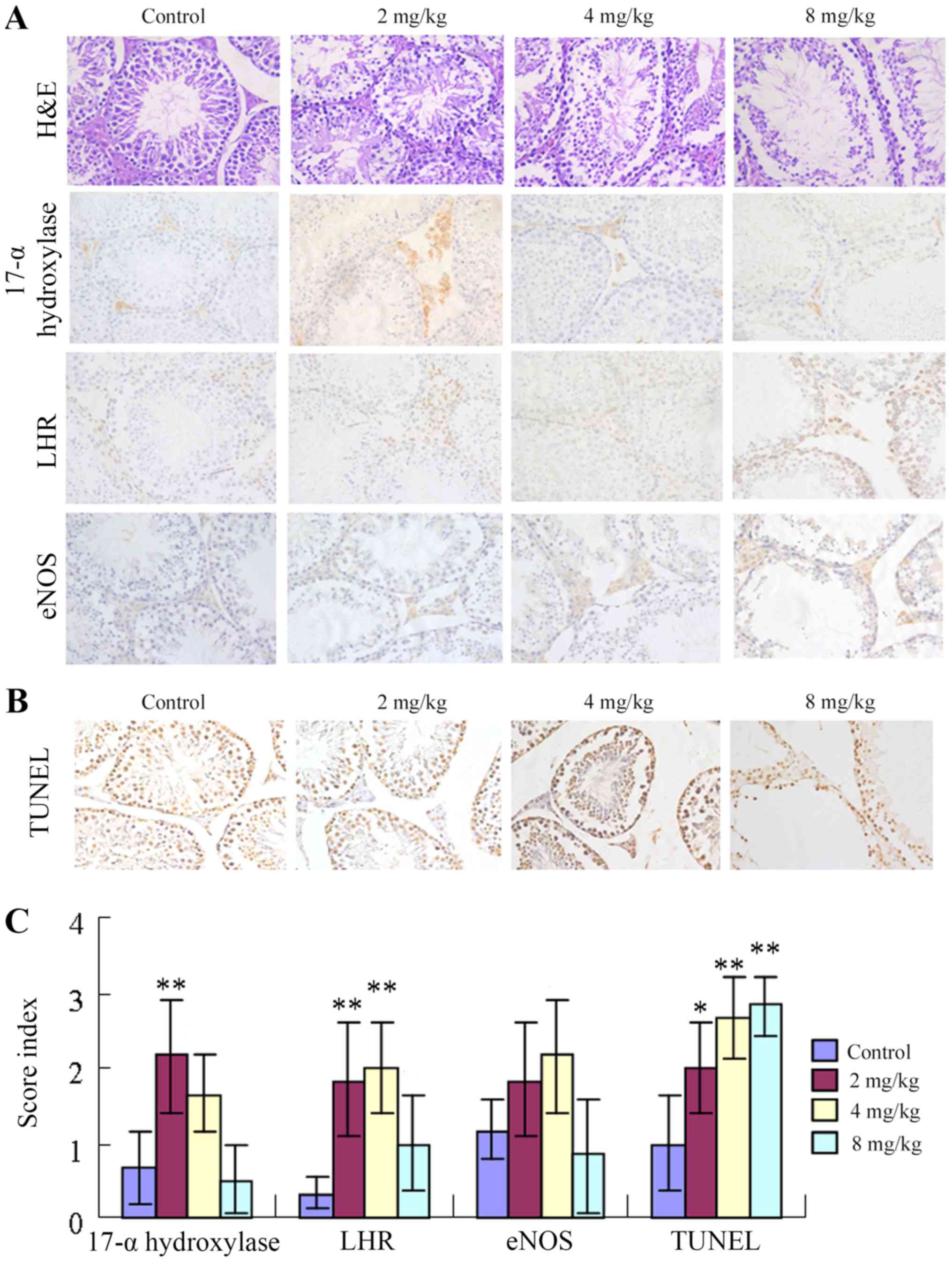 | Figure 3.Influence of cadmium on testicular
tissue protein levels and apoptosis of testicular cells. (A)
H&E staining revealed testicular morphology.
Immunohistochemical staining demonstrated that the testicular
morphology and expression levels of LHR, 17α-hydroxylase and eNOS
were altered in testicular tissues following administration of
different concentrations of cadmium. Magnification, ×400. (B) The
effect of cadmium on testicular cell apoptosis was measured by the
TUNEL assay. The number of TUNEL-positive nuclei following cadmium
treatment was increased in a concentration-dependent manner, which
indicated elevated rates of apoptosis. Magnification, ×400. (C)
Expression levels of LHR, 17α-hydroxylase, eNOS and TUNEL in
testicular tissues following administration of different
concentrations of cadmium. Data are presented as the mean ±
standard deviation. *P<0.05, **P<0.01 vs. control group.
eNOS, endothelial nitric oxide synthase; LHR, luteinizing hormone
receptor; TUNEL, terminal deoxynucleotidyl transferase dUTP nick
end labeling; H&E, hematoxylin and eosin. |
 | Table I.Expression levels of LHR,
17α-hydroxylase, eNOS and TUNEL in testicular tissues. |
Table I.
Expression levels of LHR,
17α-hydroxylase, eNOS and TUNEL in testicular tissues.
| Group |
17-α-hydroxylase | LHR | eNOS | TUNEL |
|---|
| Control | 0.67±0.47 | 0.33±0.18 | 1.17±0.41 | 1.00±0.63 |
| 2 mg/kg |
2.17±0.75b |
1.83±0.75b | 1.83±0.75 |
2.00±0.63a |
| 4 mg/kg | 1.67±0.52 |
2.00±0.63b | 2.17±0.75 |
2.67±0.52b |
| 8 mg/kg | 0.50±0.45 | 1.00±0.63 | 0.83±0.75 |
2.83±0.41b |
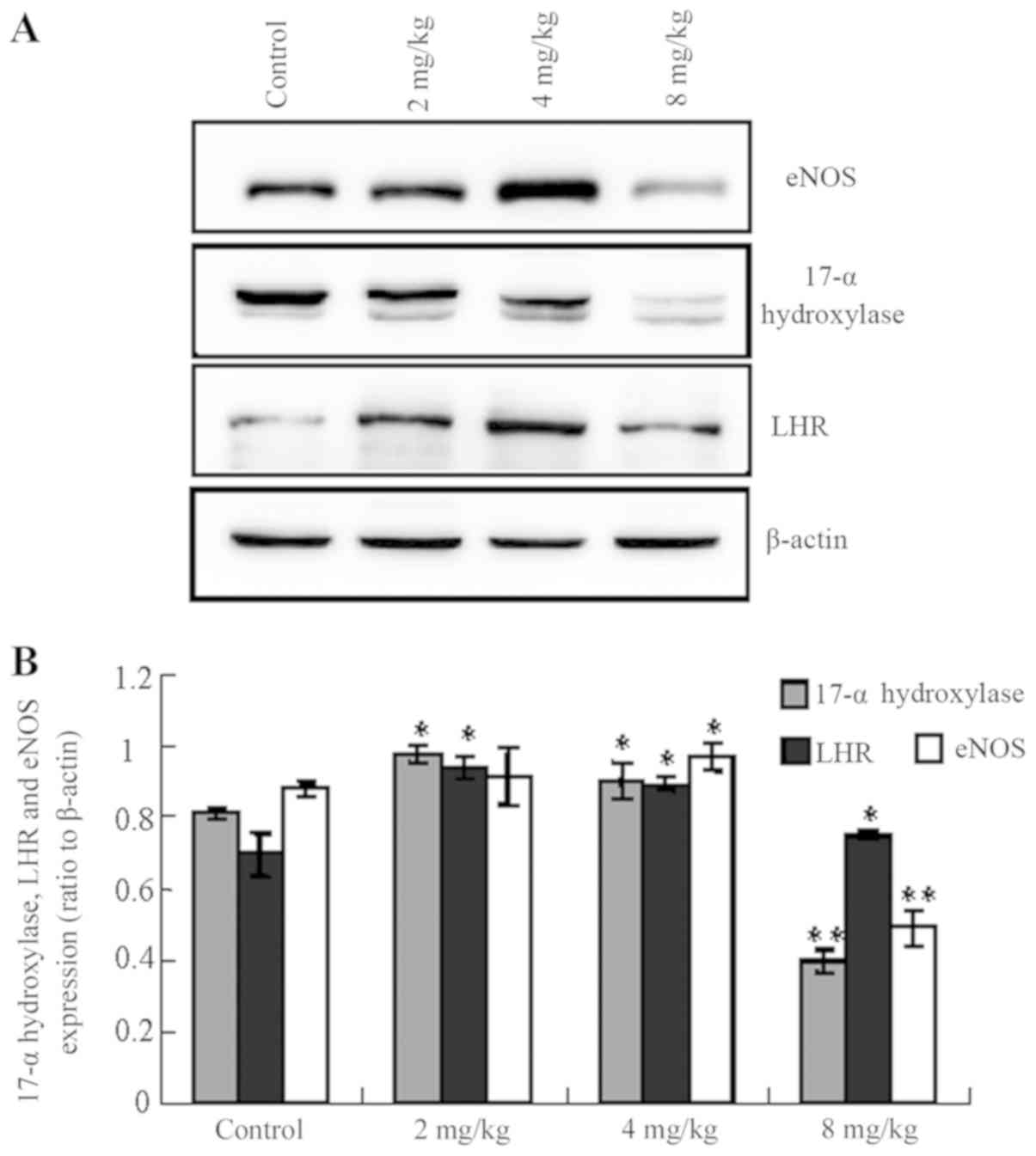
Influence of cadmium on protein levels
in testicular tissue
The expression levels of LHR, 17α-hydroxylase and
eNOS were altered following the administration of different
concentrations of cadmium. Notably, the levels of LHR,
17α-hydroxylase and eNOS were increased in the low- and
medium-cadmium groups, whereas they were significantly decreased in
the high-cadmium group compared with the control group (Fig. 4; 17-α-hydroxylase control vs.
low-/medium-/high-cadmium, P<0.05/<0.05/<0.01; LHR control
vs. low-/medium-/high-cadmium, P<0.05/<0.05/<0.05; eNOS
control vs. low-/medium-/high-cadmium,
P<0.05/<0.05/<0.01).
Discussion
The present study investigated the impact of
intragastric administration of cadmium on mice testicular features.
There was no significant difference identified in body weight
between the experimental and control groups. Cadmium dose and drug
delivery time may have affected the digestive system in mice,
although these factors were not taken into consideration.
Alternatively, perhaps due to the gavage time not being of a
sufficiently long duration, the observed cadmium dose-effect
differences were small and consequently no major effects were
observed in the BW of the mice. The histopathological results
demonstrated that the high-dose of cadmium induced seminiferous
tubule wall thinning, an increase in interstitial hyperemia level,
and lower numbers of sertoli and sperm cells. These findings
suggest that the blood-testis barrier in the mice may have been
destroyed, since the mice in the low-dose group exhibited no
changes. A previous study also demonstrated that mice exposure to
low doses of cadmium does not cause any physiological processes
disorder, whereas exposure to higher doses of cadmium affects the
autoimmune reaction of testicular tissue and leads to
microcirculation impairment, blood-testis barrier damage, which
therefore disturb spermatogenesis (37). The TUNEL results also demonstrated
that apoptosis was significantly higher in all cadmium-treated
groups compared with the control group.
Gonadotropin regulates testosterone synthesis by
Leydig cells through the gonadal axis and testosterone stimulates
spermatogenesis by binding to the androgen receptor (38–40). The
two key factors involved in testosterone synthesis by Leydig cells
are LHR and 17α-hydroxylase. In the present study, the results from
ELISA and immunochemistry demonstrated that these two proteins were
highly expressed in the low- and medium-dose cadmium groups
compared with the control group. In addition, the low-dose group
exhibited the highest level of serum androgen. These results
indicate that, in the low- and medium-cadmium groups, the gonadal
axis of the body regulates testosterone synthesis. Furthermore, the
synthesis of testosterone can be completed through feedback
mechanisms (41). The present
results also indicate that medium levels of cadmium may promote
androgen synthesis in males. It has been reported that exposure to
low doses of cadmium may increase the androgen level in males
(42–44). It is possible that there is a
critical point between the medium and high concentrations of
cadmium beyond which the compensatory activities of androgen cannot
keep the testicular structure intact. The current results
demonstrated that at a high cadmium concentration, the testicular
structure was severely impaired to a degree that may have been
beyond the body's ability to compensate. Furthermore, the
expression levels of serum androgen and proteins were decreased in
the high concentration group. What factors are involved in a
compensatory mechanism requires further investigation.
In addition, results from the present study
demonstrated that eNOS was positively expressed in testicular cells
and that eNOS expression level was the highest in the medium-dose
cadmium group. NO is an important effector molecule in the
regulation of spermatogenesis. A large quantity of NO has been
reported to exert toxic effects in sperm, whereas lower
concentrations of NO are conducive to fertility and spermatogenesis
(45,46). The present study revealed that the
highest eNOS levels were observed in the medium-cadmium
concentration group, and the serum androgen levels were highest in
the low- and medium-cadmium groups. These results suggest that NO
may be a downstream messenger that affects testosterone synthesis
or a stress-response molecule in the compensatory mechanism of
testosterone synthesis. According to the results of the present
study, the peak of eNOS was later than that of serum androgen
concentration. The former appeared in the medium concentration
group, the latter appeared in the low concentration group, and the
increase in NO concentration in this feedback mechanism was lagging
behind the synthesis of androgen. This may explain the delayed peak
in NO levels with respect to cadmium levels. Furthermore, cadmium
toxicity is associated with increased blood pressure (47). NO is a vasodilation factor that
stimulates the relaxation of endothelium-smooth muscle cells,
participating therefore in the regulation of vascular function
(48). In the present study, the
observation that eNOS was at its highest levels in the
medium-cadmium group was not compatible with the expression profile
of the two testosterone synthesis enzymes. One possibility is that
at the high cadmium concentrations, the mice experienced elevated
blood pressure as well as NO synthesis; NO can relax blood vessels
to antagonize the increased blood pressure. As higher levels of NO
stimulate relaxation of the blood vessels, an additional input of
energy would be required to maintain the body's blood pressure.
Alternatively, a medium dose of cadmium was capable of leading to
the destruction of a large number of Leydig cells. To facilitate
the functionality of the remaining testicular stromal cells, a
higher expression level of NO, leading to blood vessel dilation,
would ensure increased delivery of oxygen and nutrients to the
cells and/or extraction of metabolic waste.
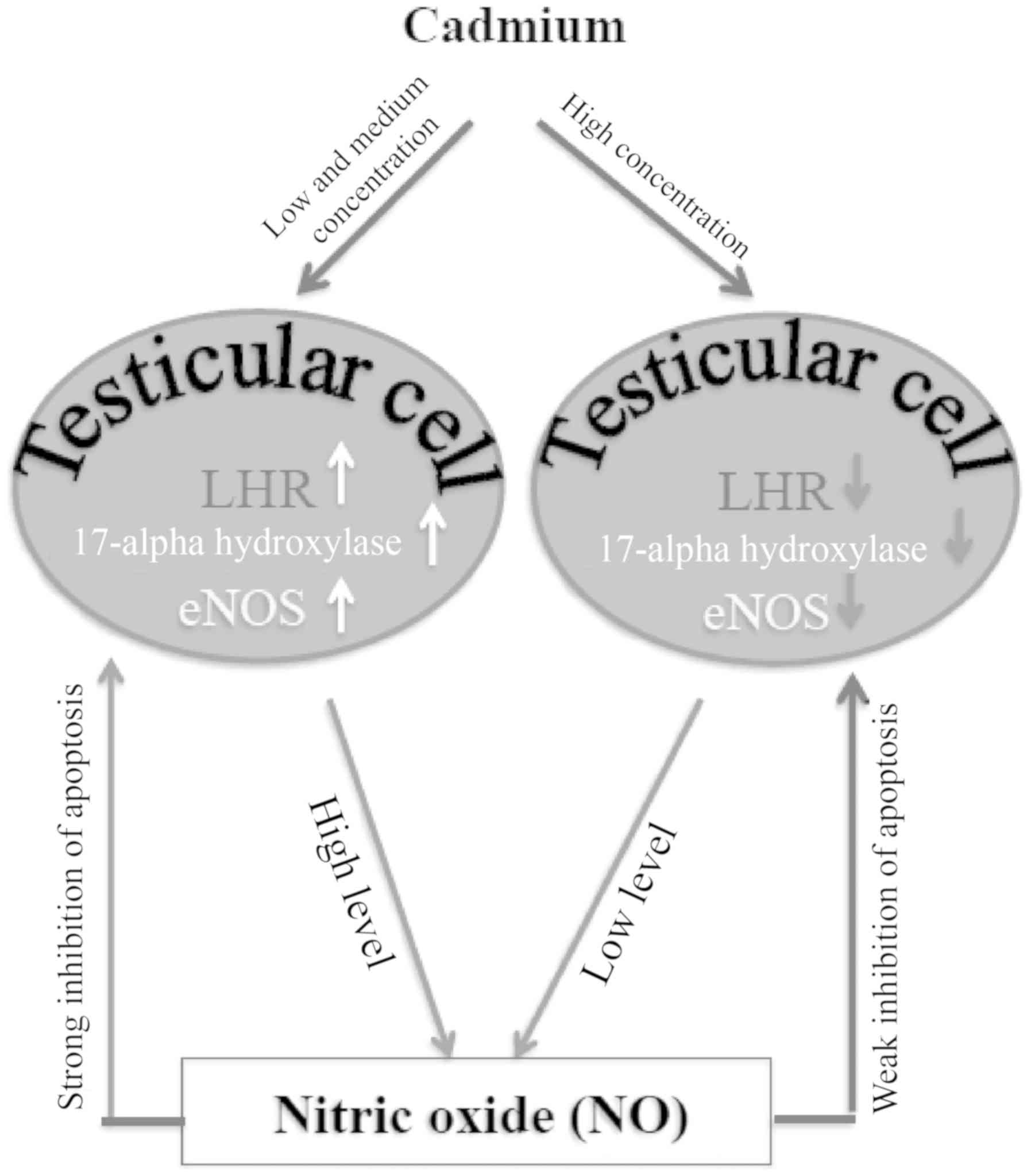
In conclusion, up to medium concentration trace
amounts of cadmium had no effect on the testicular structure of
male mice, although the number of apoptotic cells was increased.
However, increased doses of cadmium induced testicular structure
damage. The results from the present study also revealed that
cadmium toxicity altered the expression levels of LHR and
17α-hydroxylase in testicular cells, which may alter testosterone
synthesis. In addition, based on the results of the present study,
one may hypothesize that NO was involved in the repair or
compensation mechanism(s) following cadmium-induced testicular
injury and this effect was mediated by eNOS expression regulation
in Leydig cells (Fig. 5). Future
study should investigate whether other interconnected signaling
pathways are involved in this compensation mechanism. Due to
limited funds, the present study did not investigate the effect of
relevant drugs on cadmium-induced testicular injury. Therefore,
future studies should focus on this aspect.
Acknowledgements
Not applicable.
Funding
This study was supported by the Guangxi Natural
Science Foundation Project (grant no. 2016 GXNSFAA 380319) and the
Guilin Scientific Research and Technology Development project
(grant no. 20130120-8).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding authors on reasonable
request.
Authors' contributions
YR, WS, LJZ, WZ, HQ, YH, DL, CM, SZ and LZ were
responsible for research creation and design, data acquisition,
analysis and interpretation of data, statistical analysis,
manuscript drafting, and critical revision of the manuscript for
important intellectual content. All authors read and approved the
final version of manuscript.
Ethics approval and consent to
participate
All experiments were approved by the Ethical
Committees of Guilin Medical University (Guilin, China).
Patients consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Vandenberg LN, Colborn T, Hayes TB,
Heindel JJ, Jacobs DR Jr, Lee DH, Shioda T, Soto AM, vom Saal FS,
Welshons WV, et al: Hormones and endocrine-disrupting chemicals:
Low-dose effects and nonmonotonic dose responses. Endocr Rev.
33:378–455. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Medina MF, Arrieta MC, Villafañe MN,
Klyver SMR, Odstrcil IMA and González ME: Early signs of toxicity
in testes and sperm of rats exposed to low cadmium doses. Toxicol
Ind Health. 33:576–587. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Akinloye O, Arowojolu AO, Shittu OB and
Anetor JI: Cadmium toxicity: A possible cause of male infertility
in Nigeria. Reprod Biol. 6:17–30. 2006.PubMed/NCBI
|
|
4
|
Luevano J and Damodaran C: A review of
molecular events of cadmium-induced carcinogenesis. J Environ
Pathol Toxicol Oncol. 33:183–194. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Niknafs B, Salehnia M and Kamkar M:
Induction and determination of apoptotic and necrotic cell death by
cadmium chloride in testis tissue of mouse. J Reprod Infertil.
16:24–29. 2015.PubMed/NCBI
|
|
6
|
De Angelis C, Galdiero M, Pivonello C,
Salzano C, Gianfrilli D, Piscitelli P, Lenzi A, Colao A and
Pivonello R: The environment and male reproduction: The effect of
cadmium exposure on reproductive function and its implication in
fertility. Reprod Toxicol. 73:105–127. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Marettoá E, Maretta M and Leqáth J: Toxic
effects of cadmium on testis of birds and mammals: A review. Anim
Reprod Sci. 155:1–10. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Verqilio CS, Moreira RV, Carvalho CE and
Melo EJ: Evolution of cadmium effects in the testis and sperm of
the tropical fish Gymnotus carapo. Tissue Cell. 47:132–139. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang Q, Li P, Wen Y, Li S, Chen J, Liu X,
Wang L and Li X: Cadmium inhibits lysine acetylation and
succinylation inducing testicular injury of mouse during
development. Toxicol Lett. 291:112–120. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ali I, Damdimopoulou P, Stenius U,
Adamsson A, Mäkelä SI, Åkesson A, Berglund M, Håkansson H and
Halldin K: Cadmium-induced effects on cellular signaling pathways
in the liver of transgenic estrogen reporter mice. Toxicol Sci.
127:66–75. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Schneider SN, Liu Z, Wang B, Miller ML,
Afton SE, Soleimani M and Nebert DW: Oral cadmium in mice carrying
5 versus 2 copies of the Slc39a8 gene: Comparison of uptake,
distribution, metal content, and toxicity. Int J Toxicol. 33:14–20.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hu H, Lu X, Cen X, Chen X, Li F and Zhong
S: RNA-Seq identifies key reproductive gene expression alterations
in response to cadmium exposure. Biomed Res Int. 2014:5292712014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Angenard G, Muczynski V, Coffigny H,
Pairault C, Duquenne C, Frydman R, Habert R, Rouiller-Fabre V and
Livera G: Cadmium increases human fetal germ cell apoptosis.
Environ Health Perspect. 118:331–337. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Singh KP, Kumari R, Pevey C, Jackson D and
DuMond JW: Long duration exposure to cadmium leads to increased
cell survival, decreased DNA repair capacity, and genomic
instability in mouse testicular Leydig cells. Cancer Lett.
279:84–92. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ji YL, Wang H, Zhao XF, Wang Q, Zhang C,
Zhang Y, Zhao M, Chen YH, Meng XH and Xu DX: Crosstalk between
endoplasmic reticulum stress and mitochondrial pathway mediates
cadmium-induced germ cell apoptosis in testes. Toxicol Sci.
124:446–459. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Oliveira H, Lopes T, Almeida T, Pereira
Mde L and Santos C: Cadmium-induced genetic instability in mice
testis. Hum Exp Toxicol. 31:1228–1236. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Veeriah V, Saran U, Swaminathan A,
Balaguru UM, Thangaraj P, Nagarajan S, Rajendran VK and Chatterjee
S: Cadmium-induced embryopathy: Nitric oxide rescues teratogenic
effects of Cadmium. Toxicol Sci. 144:90–104. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Seid Alian N, Khodarahmi P and Naseh V:
The effect of cadmium on apoptotic genes mRNA expression of Bax and
Bcl-2 in small intestine of rats. Iran J Pathol. 13:408–414.
2018.PubMed/NCBI
|
|
19
|
Breton J, Le Clère K, Daniel C, Sauty M,
Nakab L, Chassat T, Dewulf J, Penet S, Carnoy C, Thomas P, et al:
Chronic ingestion of cadmium and lead alters the bioavailability of
essential and heavy metals, gene expression pathways and
genotoxicity in mouse intestine. Arch Toxicol. 87:1787–1795. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Uno Y, Hosaka S and Yamazaki H:
Identification and analysis of CYP7A1, CYP17A1, CYP20A1, CYP27A1
and CYP51A1 in cynomolgus macaques. J Vet Med Sci. 76:1647–1650.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen H, Ge RS and Zirkin BR: Leydig cells:
From stem cells to aging. Mol Cell Endocrinol. 306:9–16. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ge R, Chen G and Hardy MP: The role of the
Leydig cell in spermatogenic function. Adv Exp Med Biol.
636:255–269. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Smith LB and Walker WH: The regulation of
spermatogenesis by androgens. Semin Cell Dev Biol. 30:2–13. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rey RA, Musse M, Venara M and Chemes HE:
Ontogeny of the androgen receptor expression in the fetal and
postnatal testis: Its relevance on Sertoli cell maturation and the
onset of adult spermatogenesis. Microsc Res Tech. 72:787–95. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chang C, Chen YT, Yeh SD, Xu Q, Wang RS,
Guillou F, Lardy H and Yeh S: Infertility with defective
spermatogenesis and hypotestosteronemia in male mice lacking the
androgen receptor in Sertoli cells. Proc Natl Acad Sci USA.
101:6876–6881. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang F and Liao L: Role and significance
of nitric oxide neurochemical remodeling in lower urinary tract
dysfunction after spinal cord injury. Chin J Urol. 332012.
|
|
27
|
Ren YP, Sun L, Shao XY, Chen J, Xiong B
and Nong LL: Expressions of eNOS and cytochrome P450 in the testis
of sexually mature SD rats and their significance. Zhonghua Nan Ke
Xue. 15:911–914. 2009.(In Chinese). PubMed/NCBI
|
|
28
|
Lim KH, Ancrile BB, Kashatus DF and
Counter CM: Tumour maintenance is mediated by eNOS. Nature.
452:646–649. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Alpcan S, Başar H, Aydos TR, Kul O, Kısa Ü
and Başar MM: Apoptosis in testicular tissue of rats after
vasectomy: Evaluation of eNOS, iNOS immunoreactivities and the
effects of ozone therapy. Turk J Urol. 40:199–206. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kondo Y, Ishikawa T, Yamaguchi K, Yada T
and Fujisawa M: Oral administration of tetrahydrobiopterin
attenuates testicular damage by reducing nitric oxide synthase
activity in a cryptorchid mouse model. J Androl. 29:153–163. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Guo J, Jia Y, Tao SX, Li YC, Zhang XS, Hu
ZY, Chiang N, Lue YH, Hikim AP, Swerdloff RS, et al: Expression of
nitric oxide synthase during germ cell apoptosis in testis of
cynomolgus monkey after testosterone and heat treatment. J Androl.
30:190–199. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zheng LP, Zhu X, Yang L, et al: Analysis
and correlation study of trace elements in blood and semen of male
sterile patients. Mod Prev Med. 1464–1466. 2012.
|
|
33
|
Cheng DZ, Du AL, Li CS, et al: Scavenging
of superoxide free radicals generated by autooxidation of
pyrogallol with ginger extract. Chin Spices. 35–39. 2014.
|
|
34
|
Saito H, Good S, Sato S, Ito A, Ikumi Y,
Tanaka S, Ida T, Fujii S, Akaike T and Shimokawa H: Important role
of endothelial caveolin-1 in the protective role of
endothelium-dependent hyperpolarization against nitric
oxide-mediated nitrative stress in microcirculation in mice. J
Cardiovasc Pharmacol. 71:113–126. 2018.PubMed/NCBI
|
|
35
|
Vladimir K: Gross anatomy, anatomy and
normative biology, The Laboratory Mouse second edition, Chapter
2.2. 145–149. 2012.
|
|
36
|
Ohtani K, Yanagiba Y, Ashimori A, Takeuchi
A, Takada N, Togawa M, Hasegawa T, Ikeda M and Miura N: Influence
of injection timing on severity of cadmium-induced testicular
toxicity in mice. J Toxicol Sci. 38:145–150. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ogawa Y, Itoh M, Hirai S, Suna S, Naito M,
Qu N, Terayama H, Ikeda A, Miyaso H, Matsuno Y, et al: Cadmium
exposure increases susceptibility to testicular autoimmunity in
mice. J Appl Toxicol. 33:652–660. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wu S, Chen Y, Fajobi T, DiVall SA, Chang
C, Yeh S and Wolfe A: Conditional knockout of the androgen receptor
in gonadotropes reveals crucial roles for androgen in gonadotropin
synthesis and surge in female mice. Mol Endocrinol. 28:1670–1681.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Denolet E, De Gendt K, Allemeersch J,
Engelen K, Marchal K, Van Hummelen P, Tan KA, Sharpe RM, Saunders
PT, Swinnen JV and Verhoeven G: The effect of a Sertoli
cell-selective knockout of the androgen receptor on testicular gene
expression in prepubertal mice. Mol Endocrinol. 20:321–334. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Welsh M, Saunders PT, Atanassova N, Sharpe
RM and Smith LB: Androgen action via testicular peritubular myoid
cells is essential for male fertility. FASEB J. 23:4218–4230. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Iwasa T, Matsuzaki T, Yano K, Yanagihara
R, Mayila Y and Irahara M: The effects of chronic testosterone
administration on hypothalamic gonadotropin-releasing hormone
regulatory factors (Kiss1, NKB, pDyn and RFRP) and their receptors
in female rats. Gynecol Endocrinol. 34:437–441. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Telisman S, Colak B, Pizent A, Jurasović J
and Cvitković P: Reproductive toxicity of low-level lead exposure
in men. Environ Res. 105:256–266. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Meeker JD, Rossano MG, Protas B,
Padmanahban V, Diamond MP, Puscheck E, Daly D, Paneth N and Wirth
JJ: Environmental exposure to metals and male reproductive
hormones: Circulating testosterone is inversely associated with
blood molybdenum. Fertil Steril. 93:130–140. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Jurasović J, Cvitković P, Pizent A, Colak
B and Telisman S: Semen quality and reproductive endocrine function
with regard to blood cadmium in Croatian male subjects. Biometals.
17:735–743. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Weinberg JB, Doty E, Bonaventura J and
Haney AF: Nitric oxide inhibition of human sperm motility. Fertil
Steril. 64:408–413. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hellstrom WJ, Bell M, Wang R and Sikka SC:
Effect of sodium nitroprusside on sperm motility, viability, and
lipid peroxidation. Fertil Steril. 61:1117–1122. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Prozialeck WC, Edwards JR, Nebert DW,
Woods JM, Barchowsky A and Atchison WD: The vascular system as a
target of metal toxicity. Toxicol Sci. 102:207–218. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Chen CA, Wang TY, Varadharaj S, Reyes LA,
Hemann C, Talukder MA, Chen YR, Druhan LJ and Zweier JL:
S-glutathionylation uncouples eNOS and regulates its cellular and
vascular function. Nature. 468:1115–1118. 2010. View Article : Google Scholar : PubMed/NCBI
|