Glycosylation is an enzymatic process that links
glycan sugars to other glycans, lipids or proteins and is essential
to virtually every biological process (1). The complete pattern of glycan
modifications in a cell or tissue (known as ‘the glycome’) is
assembled by the synchronised action of numerous glycosylation
enzymes on glycoproteins and/or lipids (2). Changes to the glycome are well
documented in cancer, and aberrant glycosylation is not just a
consequence, but also a driver of a malignant phenotype (3). The hallmarks of cancer were originally
described in 2000 and refer to capabilities acquired during the
multi-step development of cancer to enable cancer cells to survive,
proliferate and metastasise (4).
They include sustaining proliferative signalling, evading growth
suppressors, resisting cell death, enabling replicative
immortality, inducing angiogenesis, and activating invasion and
metastasis. Underpinning these hallmarks are genome instability and
inflammation which contribute to multiple hallmark capabilities
(4). In 2011 two next generation
cancer hallmarks were proposed (reprogramming of energy metabolism
and evading immune destruction) and the ‘tumour microenvironment’
was recognised as contributing to the acquisition of hallmark
traits (5).
Although not included in the original and next
generation hallmarks, aberrant glycosylation is now also widely
recognised as a new hallmark of cancer causally associated with all
of the hallmark capabilities (2,3,6). One of the most widely occurring cancer
associated changes in glycosylation is abnormal sialylation, which
is often driven by the altered expression of sialyltranserase
enzymes (7–9) and is linked to poor patient prognosis
and metastasis (10–13). Several sialyltransferase enzymes are
implicated in cancer, but in recent years ST6GAL1 (which catalyses
the addition of α2,6-linked sialic acids onto terminal N-glycans)
has become increasingly dominant in the literature. ST6GAL1 is
upregulated in numerous types of cancer, including pancreatic,
prostate, breast and ovarian cancer, and is has key roles in tumour
aggression and metastasis (14–18).
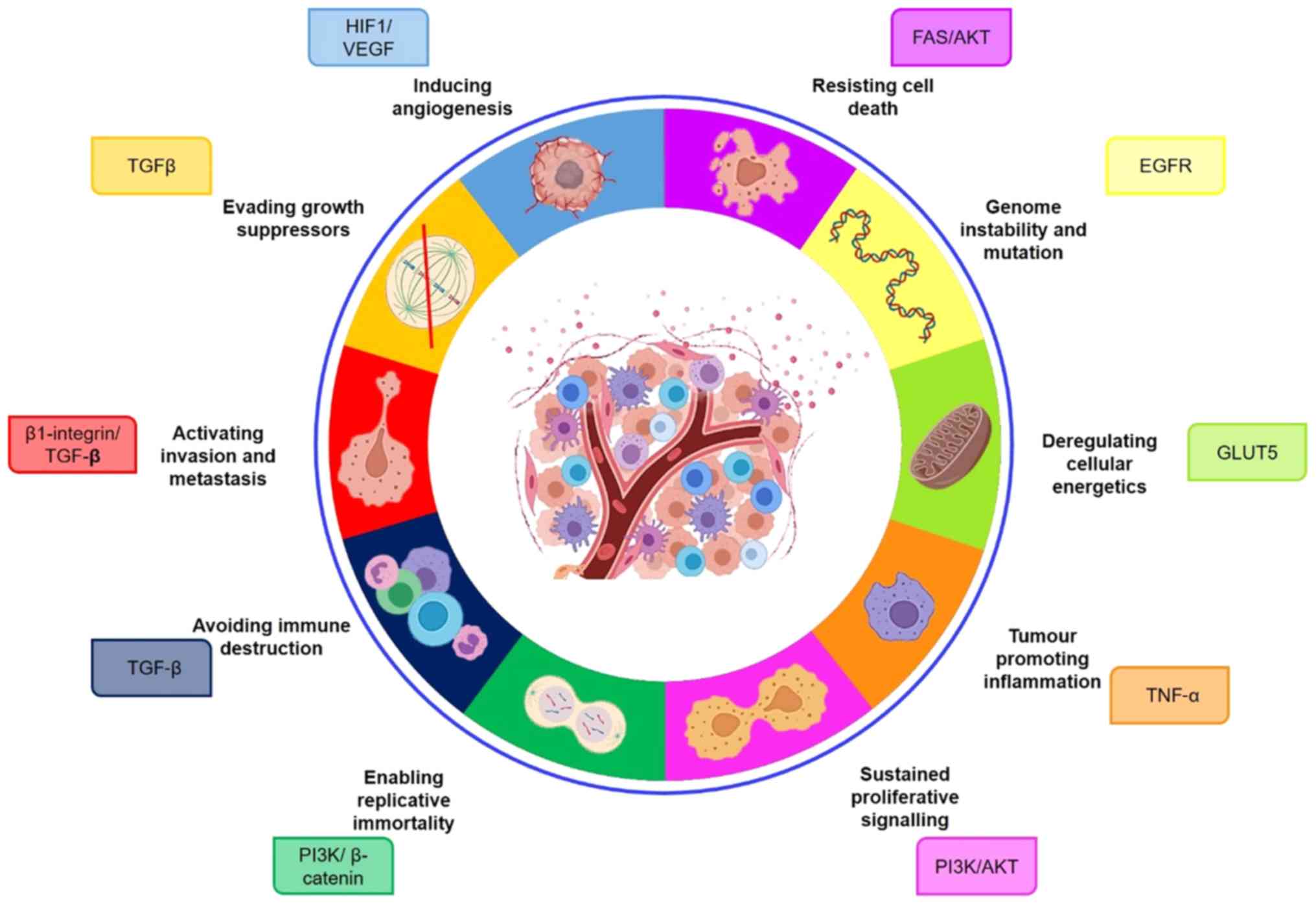
Here, we discuss ST6GAL in the context of the original and emerging
hallmarks of cancer, and highlight its role in pathways intrinsic
to tumour cell biology (Fig. 1).
ST6GAL1 levels are upregulated in several
carcinomas, as is the degree of α2,6-sialylation (15–17,19–26)
(Table I). In particular, elevated
ST6GAL1 is often correlated with high tumour grade, metastasis and
reduced patient prognosis. In both prostate and breast cancer,
ST6GAL1 expression correlates with a more aggressive tumour grade
(16,27), and in ovarian cancer levels increase
in advanced stage disease (18).
Tumours with elevated ST6GAL1 expression are thought to be more
invasive and metastatic, evidenced by increased lymphovascular
invasion, deep stromal invasion, distant metastasis and
neighbouring vesicle invasion (16,28,29).
ST6GAL1 is also associated with reduced recurrence-free intervals
and a poorer overall survival in ovarian, prostate and pancreatic
cancer (16,18,28,30). The
only exception is bladder cancer, where ST6GAL1 is believed to have
a tumour suppressive role (31).
The ability of cancer cells to invade and spread is
central to the development of an invasive, malignant tumour. This
development, which promotes local invasion and distant metastasis
is a multistep process referred to as the invasion-metastasis
cascade (5). The process is
regulated via complex crosstalk across several signalling pathways
and transcription factors, resulting in an epithelial to
mesenchymal transition (EMT) of cancer cells (32). EMT is an example of cellular
phenotype switching, characterised by a loss of epithelial markers
in favour of the migratory phenotype of mesenchymal cells (33,34). The
literature demonstrating a relationship between sialylation and the
acquisition of invasive and metastatic phenotypes is extensive
(13,35–41).
Further to this, ST6GAL1 upregulation has been shown to induce a
more invasive, migratory cell phenotype in gastric, colon, liver,
prostate, ovarian, breast and cervical cancers (16,17,42,43).
ST6GAL1 gene knockdown and over expression experiments in
vitro have demonstrated its capacity to regulate the invasive
and metastatic features of cancer cells in several cell types
(16,24,35,42–44). In
2017, a prominent glyco-oncology study identified elevated
ST6GAL1 as part of a pro-metastatic gene signature in
melanoma tumours (29).
ST6GAL1 is also increased in patients with metastatic
cervical cancer (7,43), where levels correlate with stromal
invasion, metastatic spread to the lymph nodes and poor patient
prognosis (43). Similarly, in
triple-negative breast cancer, ST6GAL1 levels are linked to
metastasis and reduced survival times (19), and high ST6Gal1 in ovarian cancer is
associated with lymphovascular invasion and distant metastasis
(7). In breast cancer cells,
overexpression of ST6GAL increases the turnover of cell surface
E-cadherin and promotes TGF-β-induced EMT providing a potential
mechanistic link between ST6GAL1-mediated sialylation and
metastasis (17).
Central to cancer cell biology is the excessive
capacity to proliferate, and to do so in the absence of
proliferative stimuli (4,45). Sialylation has been shown to alter
proliferative signalling cascades in different cancers (24,46).
ST6GAL1 in particular can regulate cellular proliferation in
hepatocellular carcinoma (HCC), as shown through gene knockdown and
overexpression studies carried out in the MHCC97L HCC cell line.
The HCC in vitro studies suggested that overexpression of
ST6GAL1 in HCC cells increased activation of the PI3K/Akt
signalling pathway (42).
Hyper-activation of the PI3K signalling cascade is well documented
in several different cancer subtypes as regulating and promoting
hyper-proliferation of oncogenic cells (47–50). If
ST6GAL1 can activate PI3K/Akt signalling, as suggested by Zhao
et al (42) this may point
towards mechanistic link between ST6GAL1 upregulation and
increased cellular proliferation. Similar effects on proliferation
and PI3K/Akt signalling have also been observed following
ST6GAL1 gene silencing in the DU145 and PC-3 prostate cancer
cell lines. Wei et al (16)
observed approximately a two-fold decrease in cancer cell
proliferation following ST6GAL1 gene silencing. It is
important to note that this relationship has only been observed in
HCC and prostate cancer, and is contradicted by findings generated
using glioma cells, assessing the relationship between ST6GAL1 and
proliferative capacity. In this instance, ST6GAL1 overexpression
did not perpetuate hyper-proliferation (23). This contradiction could suggest that
the effect of ST6GAL1 on proliferative signalling may be exclusive
to specific cancer subtypes.
In the seminal 2000 ‘hallmarks of cancer’ paper,
replicative immortality was outlined as being essential to develop
and sustain macroscopic tumours (4,5). This
innate replicative potential, without the threat of cellular
senescence has been termed immortalization and is underpinned by
telomere abnormalities or oncogenic induced cellular senescence.
Several key oncogene or tumour suppressor genes have a role in
promoting oncogenic induced cellular senescence (including Tp53,
RAS, c-MYC and PTEN) and some have been shown to be
targets for glycosylation (51–56). As
outlined previously, ST6GAL1 can regulate PI3K/AKT signalling, a
known RAS effector cascade (16,57).
ST6GAL1 gene knockdown in prostate cancer cells results in
decreased levels of PI3K/AKT/GSK-3β and β-catenin signalling
molecules (16). Β-catenin, a member
of the Wnt signalling pathway, has a well-defined oncogenic
activity and has been well characterised as an enabler of
replicative immortality through direct activation of telomerase
reverse transcriptase (TERT) (58,59).
Sialylation by ST6GAL1 upregulates several oncogenes crucial for
the immortalization of cancer cells, interacting with RAS effector
pathways, and Wnt/β-catenin signalling. Known downstream effects of
these pathways result in telomerase repression and oncogenic
induced stress, thereby indicating a role for ST6GAL1 in enabling
replicative immortality (52).
For a cancer to sustain macroscopic tumour
development and promote metastasis, the formation of neovasculature
is necessary for the supply of nutrients and oxygen. This process,
known as angiogenesis, is tightly regulated by opposing factors;
stimulating and inhibiting the receptors displayed on the surface
of vascular endothelial cells (4,5,60,61).
Although key molecular regulators of angiogenesis have been
identified, such as VEGF and TSP-1, it is now widely accepted that
regulation of angiogenesis is a highly complex process, heavily
influenced by the tumour microenvironment, gaining influence from
things such as tumour metabolism, immune infiltrate and
cancer-associated fibroblasts (CAFs) (62). Abnormal glycosylation changes have
been identified throughout several pro-angiogenic pathways in
cancers, and recently it was found that VEGF-induced angiogenesis
was dependent upon sialylation of the VEGF-receptor 2 (VEGFR2)
(63–68). A high profile Cell paper, published
in 2014, concluded that cancer cells can undergo hypoxia induced
glycan remodelling which can confer tumour resistance to anti-VEGF
treatment. Levels of ST6GAL1 were elevated in tumours sensitive to
anti-VEGF treatment, and ST6GAL1 knockdown protected tumours
from anti-VEGF treatment (22,67). In
line with this, knockdown of ST6GAL1 in an osteosarcoma cell
line also reduced levels of VEGF (22), suggesting a major role for ST6GAL1 in
cancer associated angiogenesis.
Tumour metabolism and hypoxia play a key part in
promoting the formation of new blood vessels through the activation
of several pro-angiogenic factors (62,69). The
major determinant of hypoxia mediated angiogenesis is HIF1, a
protein capable of upregulating VEGF and PI3K signalling in the
absence of oxygen to promote the growth of new vessels (69,70).
Hypoxia experiments carried out in ovarian and pancreatic cancer
cell lines indicate increased ST6GAL1 expression can lead to
an accumulation of HIF1α under hypoxic conditions, as well as
increases in HIF1α transcriptional targets (20). These increases suggest upregulated
ST6GAL1 confers pro-survival characteristics under hypoxic
conditions. Taken alongside evidence of ST6GAL1 as a major
regulator of VEGF signalling, this suggests ST6GAL1 is an important
sialyltransferase critical for angiogenesis.
It is well established that for a cancer to develop
it must evade and overcome cellular apoptosis (71–73).
Apoptosis is the programmed death of a cell, central for ensuring
the correct cell turnover and development, so much so that aberrant
apoptosis has been implicated in several human diseases, including
cancer (74). For this reason,
research into the mechanisms which underpin cell death has exploded
over the past two decades, and we now have a fair understanding of
the prominent molecular processes which regulate apoptosis
(75). Glycan changes have been
linked to apoptosis since the late 1990s (76–80), and
sialylation (including ST6GAL1 mediated sialylation) has been
functionally associated with the programmed cell death of several
different cell types (21,81–83). The
TNF family of death receptors regulate programmed cell death, and
include proteins such as DR4, DR5 and FAS. In a colon cancer model,
ST6GAL1 upregulation can decrease levels of FAS mediated cell death
(independent of both DR4 and DR5) through direct sialylation of the
FAS protein (84). As mentioned
earlier, elevated levels of ST6GAL1 drive hyper-activated AKT
signalling in several cancer models, a key pathway which can be
upregulated to enable tumour cells to evade apoptosis (85,86). Due
to the large number of downstream targets of PI3K signalling, the
effect of ST6GAL sialylation on this pathway may result in changes
in both cell survival and cellular proliferation. This, taken with
evidence that a key TNF death receptor is a direct target of
ST6GAL1, indicates that ST6GAL1 upregulation can confer
anti-apoptotic characteristics.
Tumour suppressor genes negatively regulate cell
proliferation and tumour growth and are vital ‘gate-keepers’ of the
genome (87). A key feature of
cancer cells is their ability to inactivate or avoid these growth
suppressing signals to continue hyper-proliferation (4,5). Several
known tumour suppressor genes have been identified as targets of
abnormal glycosylation, thereby promoting tumourigenesis (2,88).
Although in all other cancers sialylation by ST6GAL1 appears to be
a pro-oncogenic event, upregulation of the sialyltransferase in
bladder cancer appears to have a tumour suppressive role, with low
ST6GAL1 expression being a feature of more advanced invasive
disease, and upregulation of ST6GAL1 a hallmark of non-invasive
disease (31). This obvious
contradiction with evidence from other cancer types highlights the
heterogeneous role of ST6GAL1 activity in cancer. It is useful to
note the functional association between ST6GAL1 directed
sialylation and TGFβ signalling, as TGFβ has been identified as
both a tumour suppressor gene and a cytokine capable of promoting
oncogenic events (89). The complex
nature of cancer disease biology and of sialylation may mean that
ST6GAL1 has dual roles in both promoting and inhibiting cancer
progression.
In 2011, Hanahan and Weinberg revisited their
original hallmarks of cancer and proposed two emerging hallmarks,
‘deregulating cellular energetics’ and ‘avoiding immune
destruction’. They also identified two enabling characteristics,
‘promoting inflammation’ and ‘genome instability and mutation’ as
crucial in the acquisition of the cancer hallmarks (5). Consistent with the previous hallmarks
of cancer, ST6GAL1 appears to have important interactions with
pathways important in the ‘next generation’ hallmarks of cancer. An
anomaly of cancer cell biology is that even in the presence of
oxygen, cancer cells will fuel themselves using aerobic
glycolysis-a phenomenon now termed the ‘Warburg effect’ (90). This metabolic reprograming allows
cancer cells to thrive and meet the energetic demands of their
proliferative capacity, and has been shown to be associated with
changes in glycosylation (91,92).
This need to proliferative often leaves cells in a glucose deficit,
at which point other sugar substrates are utilised to sustain
tumour growth (93). High dietary
intake of fructose has been linked to increased risk of pancreatic
cancer, and also linked to metastatic pancreatic cancer (15). In the same study, ST6GAL1 was found
to be increased in metastatic disease, in a fructose dependant
manner and through regulation by the GLUT5 fructose receptor. This
link with GLUT5 suggests a possible link to sialylation by ST6GAL1
and metabolic reprogramming of cancer cells.
The ability of cancer cells to avoid immune
destruction, through activation of immune suppressors allows for
uninterrupted tumour growth and progression (94). As mentioned earlier, there is a
mechanistic link between TGF-β signalling and ST6GAL1 directed
sialylation. TGF-β is a known immunosuppresor gene, important in
the regulation of helper T-cells and regulatory T-cells, inhibiting
cytokine production and suppressing macrophages, dendritic cells
and natural killer cells (95,96).
Although the association between TGF-β and ST6GAL1 is better
understood in the context of EMT, there may be a role for
sialylation in allowing cancer cells to evade immune destruction
through this TGF-β interaction. At odds with the idea that immune
cells seek to destroy cancer cells, the evidence now suggests that
some infiltrating immune cells promote tumourigenesis, contributing
growth factors, survival factors and pro-angiogenic factors which
all help to sustain the tumour microenvironment (97–99).
Glycosylation, and in particular sialylation is known to play an
important role in regulation of the immune response (100,101).
ST6GAL1-null mice exhibit a widespread immunodeficient phenotype,
indicating that ST6GAL1 sialylation is integral to regulation of
the immune system (102). ST6GAL1
has been shown to promote B cell activation. IgG has also been
shown to be a direct target for ST6GAL1 sialylation in an estrogen
dependant manner in rheumatoid arthritis (103). It is evident that immune signalling
is a substrate for regulation by sialylation, however much more
needs to be done to characterise the effect of ST6GAL1 on the
immune system (104). Regulation of
the immune response through aberrant sialylation could result in
tumour promoting inflammation.
Genomic instability and an accumulation of genomic
mutations underpin carcinogenesis in all cancer subsets. In an
attempt to avoid instability, DNA damage sensors actively survey
the genome for DNA damage, and upon recognition of mutations
activate DNA repair pathways. Mutation or inactivation of these
repair pathways, such as the p53 signalling pathway, will
inevitably result in mutational accumulation. An upstream regulator
of DNA repair pathways is EGFR, known to regulate DNA repair, DNA
replication and maintenance of genome stability when found in the
nucleus (105). A study of ST6GAL1
sialylation in ovarian cancer, suggested that increased ST6GAL1
sialylation confers resistance to chemotherapeutic intervention
(106). Of interest, they
postulated that this chemoresistance was through direct sialylation
of EGFR by ST6GAL1, resulting in heightened activation of EGFR.
This suggests a direct mechanistic link between ST6GAL1-sialylation
and DNA damage repair and implicates ST6GAL1 in the maintenance of
genome stability.
The hallmarks of cancer are crucial to our
understanding of cancer cell biology, and in guiding our efforts to
identify novel biomarkers and develop new therapeutic strategies.
Aberrant glycosylation is a universal feature of cancer cells, and
glycans can modulate several of the pathways intrinsic to tumour
cell biology. Here, we suggest that the sialyltransferase enzyme
ST6GAL1 has widespread applications in the study of cancer, and
importantly is implicated in all of the recognised cancer hallmarks
(Fig. 1). Given the widespread
impact of sialylation in cancer, and the evident prognostic value
of ST6GAL1 levels, an improved understanding of how ST6GAL1
mediated sialylation sustains cancer cell biology may open the door
to a new range of cancer therapeutics.
Not applicable.
The present study was supported by Prostate Cancer
UK through a Research Innovation Award (grant no.
RIA16-ST2-011).
Not applicable.
RG, ES, KEL and JM jointly conceived and designed
the review, researched the literature and wrote the manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interest.
|
1
|
Varki A: Biological roles of glycans.
Glycobiology. 27:3–49. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pinho SS and Reis CA: Glycosylation in
cancer: Mechanisms and clinical implications. Nat Rev Cancer.
15:540–555. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Munkley J and Elliott DJ: Hallmarks of
glycosylation in cancer. Oncotarget. 7:35478–35489. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Vajaria BN and Patel PS: Glycosylation: A
hallmark of cancer? Glycoconj J. 34:147–156. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang PH, Lee WL, Juang CM, Yang YH, Lo WH,
Lai CR, Hsieh SL and Yuan CC: Altered mRNA expressions of
sialyltransferases in ovarian cancers. Gynecol Oncol. 99:631–639.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bhide GP and Colley KJ: Sialylation of
N-glycans: Mechanism, cellular compartmentalization and function.
Histochem Cell Biol. 147:149–174. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Munkley J, Oltean S, Vodak D, Wilson BT,
Livermore KE, Zhou Y, Star E, Floros VI, Johannessen B, Knight B,
et al: The androgen receptor controls expression of the
cancer-associated sTn antigen and cell adhesion through induction
of ST6GalNAc1 in prostate cancer. Oncotarget. 6:34358–34374. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vajaria BN, Patel KR, Begum R and Patel
PS: Sialylation: An Avenue to Target Cancer Cells. Pathol Oncol
Res. 22:443–447. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Munkley J: The role of Sialyl-Tn in
cancer. Int J Mol Sci. 17:2752016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Scott E and Munkley J: Glycans as
biomarkers in prostate cancer. Int J Mol Sci. 20:E13892019.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang Z, Wuhrer M and Holst S: Serum
sialylation changes in cancer. Glycoconj J. 35:139–160. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dall'Olio F: The
sialyl-alpha2,6-lactosaminyl-structure: Biosynthesis and functional
role. Glycoconj J. 17:669–676. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hsieh CC, Shyr YM, Liao WY, Chen TH, Wang
SE, Lu PC, Lin PY, Chen YB, Mao WY, Han HY, et al: Elevation of
β-galactoside α2,6-sialyltransferase 1 in a fructoseresponsive
manner promotes pancreatic cancer metastasis. Oncotarget.
8:7691–7709. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wei A, Fan B, Zhao Y, Zhang H, Wang L, Yu
X, Yuan Q, Yang D and Wang S: ST6Gal-I overexpression facilitates
prostate cancer progression via the PI3K/Akt/GSK-3β/β-catenin
signaling pathway. Oncotarget. 7:65374–65388. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lu J, Isaji T, Im S, Fukuda T, Hashii N,
Takakura D, Kawasaki N and Gu J: β-Galactoside
α2,6-sialyltranferase 1 promotes transforming growth
factor-β-mediated epithelial-mesenchymal transition. J Biol Chem.
289:34627–34641. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wichert B, Milde-Langosch K, Galatenko V,
Schmalfeldt B and Oliveira-Ferrer L: Prognostic role of the
sialyltransferase ST6GAL1 in ovarian cancer. Glycobiology.
28:898–903. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hebbar M, Krzewinski-Recchi MA, Hornez L,
Verdière A, Harduin-Lepers A, Bonneterre J, Delannoy P and Peyrat
JP: Prognostic value of tumoral sialyltransferase expression and
circulating E-selectin concentrations in node-negative breast
cancer patients. Int J Biol Markers. 18:116–122. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jones RB, Dorsett KA, Hjelmeland AB and
Bellis SL: The ST6Gal-I sialyltransferase protects tumor cells
against hypoxia by enhancing HIF-1α signaling. J Biol Chem.
293:5659–5667. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu Z, Swindall AF, Kesterson RA, Schoeb
TR, Bullard DC and Bellis SL: ST6Gal-I regulates macrophage
apoptosis via α2-6 sialylation of the TNFR1 death receptor. J Biol
Chem. 286:39654–39662. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Meng Q, Ren C, Wang L, Zhao Y and Wang S:
Knockdown of ST6Gal-I inhibits the growth and invasion of
osteosarcoma MG-63 cells. Biomed Pharmacother. 72:172–178. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yamamoto H, Oviedo A, Sweeley C, Saito T
and Moskal JR: Alpha2,6-sialylation of cell-surface N-glycans
inhibits glioma formation in vivo. Cancer Res. 61:6822–6829.
2001.PubMed/NCBI
|
|
24
|
Zhao Y, Wei A, Zhang H, Chen X, Wang L,
Zhang H, Yu X, Yuan Q, Zhang J and Wang S: α2,6-Sialylation
mediates hepatocellular carcinoma growth in vitro and in vivo by
targeting the Wnt/β-catenin pathway. Oncogenesis. 6:e3432017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Munkley J: Glycosylation is a global
target for androgen control in prostate cancer cells. Endocr Relat
Cancer. 24:R49–R64. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Munkley J, Vodak D, Livermore KE, James K,
Wilson BT, Knight B, Mccullagh P, Mcgrath J, Crundwell M, Harries
LW, et al: Glycosylation is an androgen-regulated process essential
for prostate cancer cell viability. EBioMedicine. 8:103–116. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Recchi MA, Hebbar M, Hornez L,
Harduin-Lepers A, Peyrat JP and Delannoy P: Multiplex reverse
transcription polymerase chain reaction assessment of
sialyltransferase expression in human breast cancer. Cancer Res.
58:4066–4070. 1998.PubMed/NCBI
|
|
28
|
Bassaganas S, Allende H, Cobler L, Ortiz
MR, Llop E, de Bolós C and Peracaula R: Inflammatory cytokines
regulate the expression of glycosyltransferases involved in the
biosynthesis of tumor-associated sialylated glycans in pancreatic
cancer cell lines. Cytokine. 75:197–206. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Agrawal P, Fontanals-Cirera B, Sokolova E,
Jacob S, Vaiana CA, Argibay D, Davalos V, McDermott M, Nayak S,
Darvishian F, et al: A Systems biology approach identifies FUT8 as
a driver of melanoma metastasis. Cancer Cell. 31:804–819.e7. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Munkley J: The glycosylation landscape of
pancreatic cancer. Oncol Lett. 17:2569–2575. 2019.PubMed/NCBI
|
|
31
|
Antony P, Rose M, Heidenreich A, Knuchel
R, Gaisa NT and Dahl E: Epigenetic inactivation of ST6GAL1 in human
bladder cancer. BMC Cancer. 14:9012014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jin K, Li T, van Dam H, Zhou F and Zhang
L: Molecular insights into tumour metastasis: Tracing the dominant
events. J Pathol. 241:567–577. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Montanari M, Rossetti S, Cavaliere C,
D'Aniello C, Malzone MG, Vanacore D, Di Franco R, La Mantia E,
Iovane G, Piscitelli R, et al: Epithelial-mesenchymal transition in
prostate cancer: An overview. Oncotarget. 8:35376–35389. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Goossens S, Vandamme N, Van Vlierberghe P
and Berx G: EMT transcription factors in cancer development
re-evaluated: Beyond EMT and MET. Biochim Biophys Acta Rev Cancer.
1868:584–591. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Park JJ and Lee M: Increasing the α 2, 6
sialylation of glycoproteins may contribute to metastatic spread
and therapeutic resistance in colorectal cancer. Gut Liver.
7:629–641. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ju T, Wang Y, Aryal RP, Lehoux SD, Ding X,
Kudelka MR, Cutler C, Zeng J, Wang J, Sun X, et al: Tn and
sialyl-Tn antigens, aberrant O-glycomics as human disease markers.
Proteomics. Proteomics Clin Appl. 7:618–631. 2013.PubMed/NCBI
|
|
37
|
Schultz MJ, Swindall AF and Bellis SL:
Regulation of the metastatic cell phenotype by sialylated glycans.
Cancer Metastasis Rev. 31:501–518. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kannagi R, Izawa M, Koike T, Miyazaki K
and Kimura N: Carbohydrate-mediated cell adhesion in cancer
metastasis and angiogenesis. Cancer Sci. 95:377–384. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Irimura T, Nakamori S, Matsushita Y,
Taniuchi Y, Todoroki N, Tsuji T, Izumi Y, Kawamura Y, Hoff SD,
Cleary KR, et al: Colorectal cancer metastasis determined by
carbohydrate-mediated cell adhesion: Role of sialyl-LeX antigens.
Semin Cancer Biol. 4:319–324. 1993.PubMed/NCBI
|
|
40
|
Ugorski M and Laskowska A: Sialyl
Lewis(a): A tumor-associated carbohydrate antigen involved in
adhesion and metastatic potential of cancer cells. Acta Biochim
Pol. 49:303–311. 2002.PubMed/NCBI
|
|
41
|
Murugaesu N, Iravani M, van Weverwijk A,
Ivetic A, Johnson DA, Antonopoulos A, Fearns A, Jamal-Hanjani M,
Sims D, Fenwick K, et al: An in vivo functional screen identifies
ST6GalNAc2 sialyltransferase as a breast cancer metastasis
suppressor. Cancer Discov. 4:304–317. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhao Y, Li Y, Ma H, Dong W, Zhou H, Song
X, Zhang J and Jia L: Modification of sialylation mediates the
invasive properties and chemosensitivity of human hepatocellular
carcinoma. Mol Cell Proteomics. 13:520–536. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang PH, Lee WL, Lee YR, Juang CM, Chen
YJ, Chao HT, Tsai YC and Yuan CC: Enhanced expression of alpha
2,6-sialyltransferase ST6Gal I in cervical squamous cell carcinoma.
Gynecol Oncol. 89:395–401. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lin S, Kemmner W, Grigull S and Schlag PM:
Cell surface alpha 2,6 sialylation affects adhesion of breast
carcinoma cells. Exp Cell Res. 276:101–110. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Feitelson MA, Arzumanyan A, Kulathinal RJ,
Blain SW, Holcombe RF, Mahajna J, Marino M, Martinez-Chantar ML,
Nawroth R, Sanchez-Garcia I, et al: Sustained proliferation in
cancer: Mechanisms and novel therapeutic targets. Semin Cancer
Biol. 35 (Suppl):S25–S54. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ma X, Dong W, Su Z, Zhao L, Miao Y, Li N,
Zhou H and Jia L: Functional roles of sialylation in breast cancer
progression through miR-26a/26b targeting ST8SIA4. Cell Death Dis.
7:e25612016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Luo J, Manning BD and Cantley LC:
Targeting the PI3K-Akt pathway in human cancer: Rationale and
promise. Cancer Cell. 4:257–262. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Tokunaga E, Kimura Y, Mashino K, Oki E,
Kataoka A, Ohno S, Morita M, Kakeji Y, Baba H and Maehara Y:
Activation of PI3K/Akt signaling and hormone resistance in breast
cancer. Breast Cancer. 13:137–144. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Deying W, Feng G, Shumei L, Hui Z, Ming L
and Hongqing W: CAF-derived HGF promotes cell proliferation and
drug resistance by up-regulating the c-Met/PI3K/Akt and GRP78
signalling in ovarian cancer cells. Biosci Rep. 37:BSR201604702017.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
De Marco C, Laudanna C, Rinaldo N,
Oliveira DM, Ravo M, Weisz A, Ceccarelli M, Caira E, Rizzuto A,
Zoppoli P, et al: Specific gene expression signatures induced by
the multiple oncogenic alterations that occur within the
PTEN/PI3K/AKT pathway in lung cancer. PLoS One. 12:e01788652017.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yaswen P, MacKenzie KL, Keith WN, Hentosh
P, Rodier F, Zhu J, Firestone GL, Matheu A, Carnero A, Bilsland A,
et al: Therapeutic targeting of replicative immortality. Semin
Cancer Biol. 35 (Suppl):S104–S128. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Roninson IB: Tumor cell senescence in
cancer treatment. Cancer Res. 63:2705–2715. 2003.PubMed/NCBI
|
|
53
|
Braig M and Schmitt CA: Oncogene-induced
senescence: Putting the brakes on tumor development. Cancer Res.
66:2881–2884. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Lin AW and Lowe SW: Oncogenic ras
activates the ARF-p53 pathway to suppress epithelial cell
transformation. Proc Natl Acad Sci USA. 98:5025–5030. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Courtois-Cox S, Jones SL and Cichowski K:
Many roads lead to oncogene-induced senescence. Oncogene.
27:2801–2809. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Chou TY, Hart GW and Dang CV: c-Myc is
glycosylated at threonine 58, a known phosphorylation site and a
mutational hot spot in lymphomas. J Biol Chem. 270:18961–18965.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Castellano E and Downward J: RAS
interaction with PI3K: More than just another effector pathway.
Genes Cancer. 2:261–274. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Delmas V, Beermann F, Martinozzi S,
Carreira S, Ackermann J, Kumasaka M, Denat L, Goodall J, Luciani F,
Viros A, et al: Beta-catenin induces immortalization of melanocytes
by suppressing p16INK4a expression and cooperates with N-Ras in
melanoma development. Genes Dev. 21:2923–2935. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Zhang Y, Toh L, Lau P and Wang X: Human
telomerase reverse transcriptase (hTERT) is a novel target of the
Wnt/β-catenin pathway in human cancer. J Biol Chem.
287:32494–32511. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Eelen G, Dubois C, Cantelmo AR, Goveia J,
Brüning U, DeRan M, Jarugumilli G, van Rijssel J, Saladino G,
Comitani F, et al: Role of glutamine synthetase in angiogenesis
beyond glutamine synthesis. Nature. 561:63–69. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Hanahan D and Folkman J: Patterns and
emerging mechanisms of the angiogenic switch during tumorigenesis.
Cell. 86:353–364. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
De Palma M, Biziato D and Petrova TV:
Microenvironmental regulation of tumour angiogenesis. Nat Rev
Cancer. 17:457–474. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Chiodelli P, Rezzola S, Urbinati C,
Federici Signori F, Monti E, Ronca R, Presta M and Rusnati M:
Contribution of vascular endothelial growth factor receptor-2
sialylation to the process of angiogenesis. Oncogene. 36:6531–6541.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Cheng WK and Oon CE: How glycosylation
aids tumor angiogenesis: An updated review. Biomed Pharmacother.
103:1246–1252. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Croci DO, Cerliani JP, Pinto NA, Morosi LG
and Rabinovich GA: Regulatory role of glycans in the control of
hypoxia-driven angiogenesis and sensitivity to anti-angiogenic
treatment. Glycobiology. 24:1283–1290. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Lynch TP, Ferrer CM, Jackson SR, Shahriari
KS, Vosseller K and Reginato MJ: Critical role of O-Linked
beta-N-acetylglucosamine transferase in prostate cancer invasion,
angiogenesis, and metastasis. J Biol Chem. 287:11070–11081. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Croci DO, Cerliani JP, Dalotto-Moreno T,
Méndez-Huergo SP, Mascanfroni ID, Dergan-Dylon S, Toscano MA,
Caramelo JJ, García-Vallejo JJ, Ouyang J, et al:
Glycosylation-dependent lectin-receptor interactions preserve
angiogenesis in anti-VEGF refractory tumors. Cell. 156:744–758.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Croci DO and Rabinovich GA: Linking tumor
hypoxia with VEGFR2 signaling and compensatory angiogenesis:
Glycans make the difference. Oncoimmunology. 3:e293802014.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
LaGory EL and Giaccia AJ: The
ever-expanding role of HIF in tumour and stromal biology. Nat Cell
Biol. 18:356–365. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Zhang W, Xiong Z, Wei T, Li Q, Tan Y, Ling
L and Feng X: Nuclear factor 90 promotes angiogenesis by regulating
HIF-1α/VEGF-A expression through the PI3K/Akt signaling pathway in
human cervical cancer. Cell Death Dis. 9:2762018. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Evans EK and Kornbluth S: Regulation of
apoptosis in Xenopus egg extracts. Adv Enzyme Regul. 38:265–280.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Lowe M, Lane JD, Woodman PG and Allan VJ:
Caspase-mediated cleavage of syntaxin 5 and giantin accompanies
inhibition of secretory traffic during apoptosis. J Cell Sci.
117:1139–1150. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Kerr JF: History of the events leading to
the formulation of the apoptosis concept. Toxicology 181–182.
471–474. 2002. View Article : Google Scholar
|
|
74
|
Fernald K and Kurokawa M: Evading
apoptosis in cancer. Trends Cell Biol. 23:620–633. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Adams JM and Cory S: Bcl-2-regulated
apoptosis: Mechanism and therapeutic potential. Curr Opin Immunol.
19:488–496. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Walker BK, Lei H and Krag SS: A functional
link between N-linked glycosylation and apoptosis in Chinese
hamster ovary cells. Biochem Biophys Res Commun. 250:264–270. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Zhang D, Liu X, Gao J, Sun Y, Liu T, Yan Q
and Yang X: The role of epithelial cell adhesion molecule
N-glycosylation on apoptosis in breast cancer cells. Tumour Biol.
39:10104283176959732017.PubMed/NCBI
|
|
78
|
Rapoport E and Pendu JL: Glycosylation
alterations of cells in late phase apoptosis from colon carcinomas.
Glycobiology. 9:1337–1345. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Gwak H, Kim S, Dhanasekaran DN and Song
YS: Resveratrol triggers ER stress-mediated apoptosis by disrupting
N-linked glycosylation of proteins in ovarian cancer cells. Cancer
Lett. 371:347–353. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Seyrek K, Richter M and Lavrik IN:
Decoding the sweet regulation of apoptosis: The role of
glycosylation and galectins in apoptotic signaling pathways. Cell
Death Differ. 26:981–993. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Suzuki O, Abe M and Hashimoto Y:
Caspase-dependent drug-induced apoptosis is regulated by cell
surface sialylation in human B-cell lymphoma. Oncol Lett.
10:687–690. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Meesmann HM, Fehr EM, Kierschke S,
Herrmann M, Bilyy R, Heyder P, Blank N, Krienke S, Lorenz HM and
Schiller M: Decrease of sialic acid residues as an eat-me signal on
the surface of apoptotic lymphocytes. J Cell Sci. 123:3347–3356.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Peter ME, Hellbardt S, Schwartz-Albiez R,
Westendorp MO, Walczak H, Moldenhauer G, Grell M and Krammer PH:
Cell surface sialylation plays a role in modulating sensitivity
towards APO-1-mediated apoptotic cell death. Cell Death Differ.
2:163–171. 1995.PubMed/NCBI
|
|
84
|
Swindall AF and Bellis SL: Sialylation of
the Fas death receptor by ST6Gal-I provides protection against
Fas-mediated apoptosis in colon carcinoma cells. J Biol Chem.
286:22982–22990. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Yao R and Cooper GM: Requirement for
phosphatidylinositol-3 kinase in the prevention of apoptosis by
nerve growth factor. Science. 267:2003–2006. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Franke TF, Kaplan DR and Cantley LC: PI3K:
Downstream AKTion blocks apoptosis. Cell. 88:435–437. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Burkhart DL and Sage J: Cellular
mechanisms of tumour suppression by the retinoblastoma gene. Nat
Rev Cancer. 8:671–682. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Masuda M, Yageta M, Fukuhara H, Kuramochi
M, Maruyama T, Nomoto A and Murakami Y: The tumor suppressor
protein TSLC1 is involved in cell-cell adhesion. J Biol Chem.
277:31014–31019. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Bierie B and Moses HL: TGF-beta and
cancer. Cytokine Growth Factor Rev. 17:29–40. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Warburg O, Wind F and Negelein E: The
metabolism of tumors in the body. J Gen Physiol. 8:519–530. 1927.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Ferrer CM, Lynch TP, Sodi VL, Falcone JN,
Schwab LP, Peacock DL, Vocadlo DJ, Seagroves TN and Reginato MJ:
O-GlcNAcylation regulates cancer metabolism and survival stress
signaling via regulation of the HIF-1 pathway. Mol Cell.
54:820–831. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Ferrer CM and Reginato MJ: Sweet
connections: O-GlcNAcylation links cancer cell metabolism and
survival. Mol Cell Oncol. 2:e9618092014. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Chen WL, Wang YY, Zhao A, Xia L, Xie G, Su
M, Zhao L, Liu J, Qu C, Wei R, et al: Enhanced fructose utilization
mediated by SLC2A5 is a unique metabolic feature of acute myeloid
leukemia with therapeutic potential. Cancer Cell. 30:779–791. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Vinay DS, Ryan EP, Pawelec G, Talib WH,
Stagg J, Elkord E, Lichtor T, Decker WK, Whelan RL, Kumara HMCS, et
al: Immune evasion in cancer: Mechanistic basis and therapeutic
strategies. Semin Cancer Biol. 35 (Suppl):S185–S198. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Wrzesinski SH, Wan YY and Flavell RA:
Transforming growth factor-beta and the immune response:
Implications for anticancer therapy. Clin Cancer Res. 13:5262–5270.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Yoshimura A and Muto G: TGF-β function in
immune suppression. Curr Top Microbiol Immunol. 350:127–147.
2011.PubMed/NCBI
|
|
97
|
DeNardo DG, Andreu P and Coussens LM:
Interactions between lymphocytes and myeloid cells regulate
pro-versus anti-tumor immunity. Cancer Metastasis Rev. 29:309–316.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Qian BZ and Pollard JW: Macrophage
diversity enhances tumor progression and metastasis. Cell.
141:39–51. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Colotta F, Allavena P, Sica A, Garlanda C
and Mantovani A: Cancer-related inflammation, the seventh hallmark
of cancer: Links to genetic instability. Carcinogenesis.
30:1073–1081. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Dube DH and Bertozzi CR: Glycans in cancer
and inflammation-potential for therapeutics and diagnostics. Nat
Rev Drug Discov. 4:477–488. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Marth JD and Grewal PK: Mammalian
glycosylation in immunity. Nat Rev Immunol. 8:874–887. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Hennet T, Chui D, Paulson JC and Marth JD:
Immune regulation by the ST6Gal sialyltransferase. Proc Natl Acad
Sci USA. 95:4504–4509. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Engdahl C, Bondt A, Harre U, Raufer J,
Pfeifle R, Camponeschi A, Wuhrer M, Seeling M, Mårtensson IL,
Nimmerjahn F, et al: Estrogen induces St6gal1 expression and
increases IgG sialylation in mice and patients with rheumatoid
arthritis: A potential explanation for the increased risk of
rheumatoid arthritis in postmenopausal women. Arthritis Res Ther.
20:842018. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Perdicchio M, Ilarregui JM, Verstege MI,
Cornelissen LA, Schetters ST, Engels S, Ambrosini M, Kalay H,
Veninga H, den Haan JM, et al: Sialic acid-modified antigens impose
tolerance via inhibition of T-cell proliferation and de novo
induction of regulatory T cells. Proc Natl Acad Sci USA.
113:3329–3334. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Chou RH, Wang YN, Hsieh YH, Li LY, Xia W,
Chang WC, Chang LC, Cheng CC, Lai CC, Hsu JL, et al: EGFR modulates
DNA synthesis and repair through Tyr phosphorylation of histone H4.
Dev Cell. 30:224–237. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Britain CM, Holdbrooks AT, Anderson JC,
Willey CD and Bellis SL: Sialylation of EGFR by the ST6Gal-I
sialyltransferase promotes EGFR activation and resistance to
gefitinib-mediated cell death. J Ovarian Res. 11:122018. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Chiricolo M, Malagolini N, Bonfiglioli S
and Dall'Olio F: Phenotypic changes induced by expression of
beta-galactoside alpha2,6 sialyltransferase I in the human colon
cancer cell line SW948. Glycobiology. 16:146–154. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Schultz MJ, Holdbrooks AT, Chakraborty A,
Grizzle WE, Landen CN, Buchsbaum DJ, Conner MG, Arend RC, Yoon KJ,
Klug CA, et al: The tumor-associated glycosyltransferase ST6Gal-I
regulates stem cell transcription factors and confers a cancer stem
cell phenotype. Cancer Res. 76:3978–3988. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Gretschel S, Haensch W, Schlag PM and
Kemmner W: Clinical relevance of sialyltransferases ST6GAL-I and
ST3GAL-III in gastric cancer. Oncology. 65:139–145. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Ma H, Zhou H, Song X, Shi S, Zhang J and
Jia L: Modification of sialylation is associated with multidrug
resistance in human acute myeloid leukemia. Oncogene. 34:726–740.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Pousset D, Piller V, Bureaud N, Monsigny M
and Piller F: Increased alpha2,6 sialylation of N-glycans in a
transgenic mouse model of hepatocellular carcinoma. Cancer Res.
57:4249–4256. 1997.PubMed/NCBI
|