Introduction
A number of studies have indicated that non-coding
RNAs can serve as important regulators of biological processes
(1). MicroRNAs (miRNAs) are small
non-coding RNAs that are 18–25 nucleotides long, which are
important post-transcriptional regulators of gene expression by
inhibiting the stability or translational efficiency of target
genes (2). Recently studies have
demonstrated that miRNAs are aberrantly expressed in different
types of human cancer and serve important roles in regulating
cancer initiation and progression (3). miRNAs can regulate numerous biological
processes, including cancer cell proliferation, cycle arrest,
apoptosis, migration and invasion (4).
Gastric cancer (GC) is one of the most common
malignancies in China and exhibits a poor prognosis (5). A significant challenge for GC treatment
is the absence of early diagnostic biomarkers (6). Yuan et al (7), reported that the 5-year survival rate
of early GC is ~30-fold high compared with that of late-stage GC. A
number of studies have identified aberrantly expressed miRNAs in
GC, which serve regulatory roles in tumors (8). For example, miR-5590-3p has been
revealed to inhibit GC growth by targeting the DDX5/AKT/mammalian
target of rapamycin (mTOR) pathway (9). miRNA-223 promotes GC metastasis by
targeting EPB41L3 (10).
Additionally, miR-20a has been identified to be involved in the
resistance of GC by targeting CYLD (11). However, these studies only
investigated a small number of miRNAs and didn't perform a
systems-level identification of differentially expressed miRNAs
with a large sample size. Therefore, there is a requirement to
understand the molecular mechanisms of miRNAs that regulate GC
progression and identify miRNAs that may serve as biomarkers for
the detection of GC.
The present study identified differentially
expressed miRNAs in GC by analyzing four public datasets, including
GSE23739, GSE26295, GSE30070 and data from The Cancer Genome Atlas
(TCGA). In total, >800 GC samples were included in the present
study. Additionally, to investigate the molecular mechanisms of
aberrantly expressed miRNAs, gene ontology (GO) and Kyoto
Encyclopedia of Genes and Genomes (KEGG) analysis were
performed.
Materials and methods
Microarray data and data
preprocessing
The current study screened GC-associated miRNA
expression profiles in National Center for Biotechnology
Information (NCBI) Gene Expression Omnibus (GEO) datasets
(https://www.ncbi.nlm.nih.gov/gds/).
Candidate datasets with >50 samples were selected for further
analysis. Therefore, a total of four miRNA expression datasets of
GC were downloaded from the NCBI GEO and TCGA (https://portal.gdc.cancer.gov/) databases
including GSE23739, GSE30070, GSE26595 and data from TCGA. GSE23739
was submitted by Oh et al (12), and contained 40 GC samples and 40
normal samples obtained from the National Cancer Centre and the
Singhealth Tissue Repository (Singapore). GSE26595 was submitted by
Lim et al (13), and
contained 60 primary GC samples and 8 surrounding non-cancerous
samples from patients who underwent a curative gastrostomy as a
primary treatment method between 1999 and 2007 at Severance
Hospital and Gangnam Severance Hospital, Yonsei University College
of Medicine (Seoul, South Korea). GSE30070 was submitted by Kim
et al (14), and contained 90
GC samples and 34 normal samples collected at the Hospital of
Korean National Cancer Center by endoscopy between 2001 and 2006
following a protocol approved by the Institutional Review Board
(IRB) of the National Cancer Center Hospital in Goyang, Korea. All
patients and volunteers signed IRB-approved informed consent forms.
A total of 34 healthy volunteers underwent gastroscopy for routine
screening for GC and were confirmed to have normal gastric mucosa
by histology. No gastritis was identified among the 34 healthy
volunteers. TCGA data contained 389 GC samples and 41 normal
samples.
These datasets were based on different platforms.
For example, GSE23739 was based on Agilent Human miRNA Microarrays
and TCGA data were based on RNA-sequencing (RNA-seq). A previous
study demonstrated that RNA-seq and microarray-based methods do not
exhibit a high coincidence degree, although they are both good
technologies for measuring gene expression level (15). The present study used TCGA data and
three GEO datasets to identify common dysregulated miRNAs in GC. A
similar analysis has also been reported by a number of other groups
(16–19).
GO and KEGG pathway analysis
To predict the targets of the differentially
expressed miRNAs, four different databases were used, including
TargetScan (http://www.targetscan.org/), miRWALK (http://zmf.umm.uni-heidelberg.de/apps/zmf/mirwalk2/),
miRDB (http://www.mirdb.org/) and starbase
(http://starbase.sysu.edu.cn/). A total
of 605 target mRNAs were obtained. Accordingly, the network between
differentially expressed targets and miRNAs was constructed using
Cytoscape v3.2.1 (http://www.cytoscape.org/) software. Molecular
Annotation System 3.0 (http://bioinfo.capitalbio.com/mas3/) was used to
determine the biological roles of the target mRNAs. Gene functions
were classified into the following three subgroups: Biological
process, molecular function and cellular component. The enriched GO
terms were presented as enrichment scores. KEGG pathway analysis
(https://www.genome.jp/kegg/pathway.html) was performed
to determine the involvement of differentially expressed mRNAs in
different biological pathways using DAVID system (https://david.ncifcrf.gov/tools.jsp). P<0.05
was considered to indicate a statistically significant result.
Statistical analysis
Results were analyzed using SPSS software (version
15.0; SPSS, Inc.). Numerical data are presented as the mean ±
standard deviation. Statistical comparisons between two groups of
normalized data were performed using a student's t-test or
Mann-Whitney U-test according to the test condition. Statistical
comparisons among multiple groups of normalized data were performed
using one-way analysis of the variance followed by a Dunnett's post
hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Identification of the significantly
differentially expressed miRNAs in GC
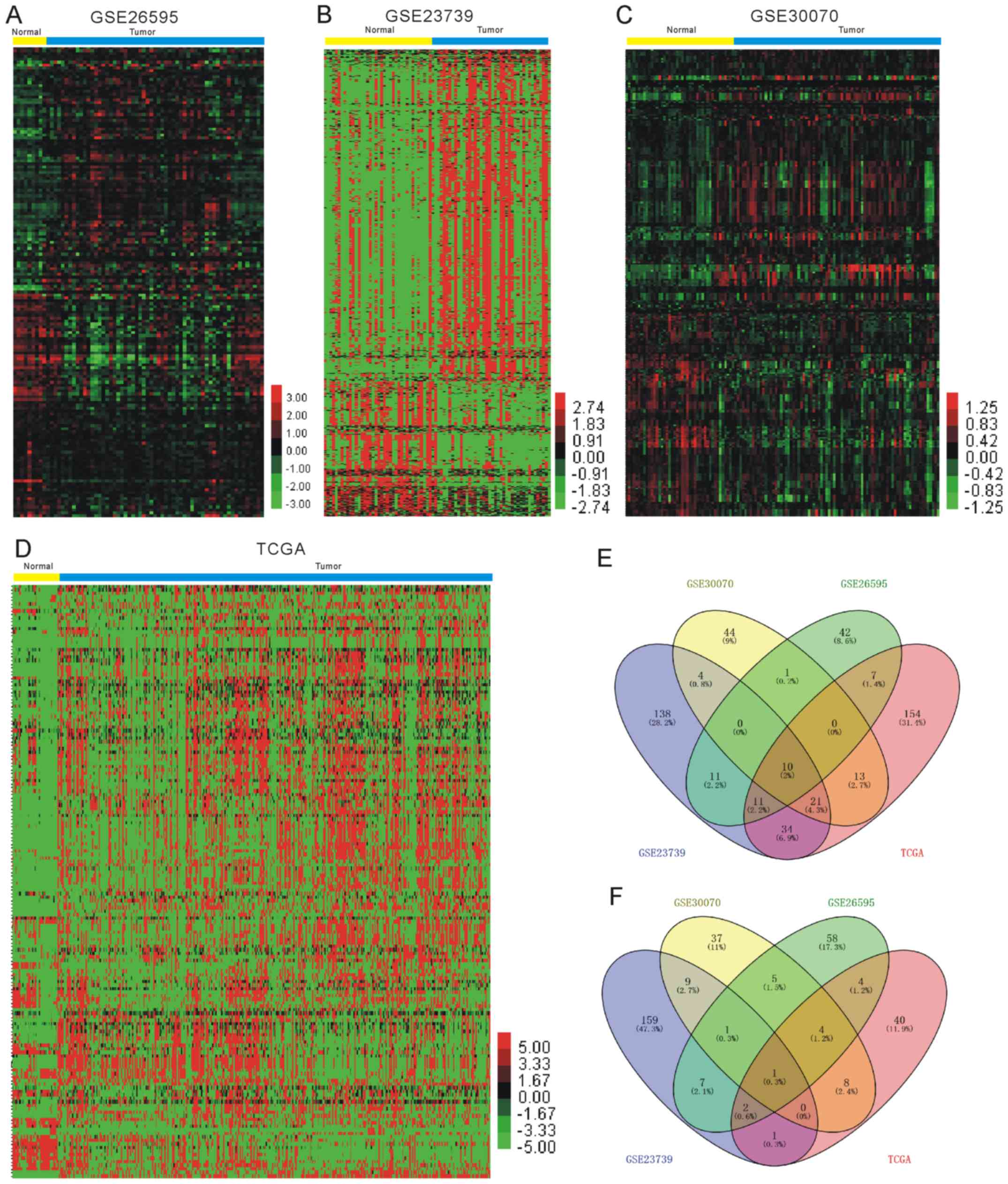
To identify the significantly differentially
expressed miRNAs in GC, the current study used four miRNA
expression profiles including 579 GC samples and 123 normal
samples, which were downloaded and analyzed from GEO and TCGA
datasets, as presented in Table I
and Fig. 1. A total of 10 miRNAs
(hsa-miR-15b-5p, hsa-miR-21-5p, hsa-miR-93-5p, hsa-let-7i-5p,
hsa-miR-25-3p, hsa-miR-185-5p, hsa-miR-181b-5p, hsa-miR-224-5p,
hsa-miR-196b-5p and hsa-miR-135b-5p) were identified as upregulated
and hsa-miR-204-5p was revealed as downregulated in GC in the four
datasets. Clustering analysis was subsequently performed for all
abnormally expressed miRNAs in GC (Fig.
1E and F; Table II).
 | Table I.Analysis of four miRNA expression
profiles, which include 579 gastric cancer samples and 123 normal
samples. |
Table I.
Analysis of four miRNA expression
profiles, which include 579 gastric cancer samples and 123 normal
samples.
| Dataset | Total cases | Normal cases | Tumor cases |
|---|
| GSE23739 | 80 | 40 | 40 |
| GSE30070 | 124 | 34 | 90 |
| GSE26595 | 68 | 8 | 60 |
| TCGA | 430 | 41 | 389 |
| Table II.Expression of miRNAs in gastric
cancer samples vs. normal tissue samples. |
Table II.
Expression of miRNAs in gastric
cancer samples vs. normal tissue samples.
|
| GSE23739 | GSE26595 | GSE30070 | TCGA |
|---|
|
|
|
|
|
|
|---|
| miRNA | P-value | T | N | P-value | T | N | P-value | T | N | P-value | T | N |
|---|
| hsa-miR-15b-5p | 0.01 | 173.40 | 81.74 | 0.01 | 12.96 | 12.67 | 0.03 | 0.22 | 0.13 | <0.01 | 272.81 | 138.62 |
| hsa-miR-21-5p | <0.01 | 21106.46 | 5068.67 | <0.01 | 14.88 | 14.51 | 0.01 | −0.26 | −0.47 | <0.01 | 28,4397.89 | 61,871.29 |
| hsa-miR-93-5p | <0.01 | 481.12 | 177.25 | <0.01 | 12.84 | 12.43 | <0.01 | −0.22 | −0.39 | <0.01 | 6,766.76 | 1,685.58 |
| hsa-let-7i-5p | <0.01 | 1787.97 | 496.38 | <0.01 | 13.44 | 13.00 | 0.01 | −0.11 | −0.19 | <0.01 | 559.93 | 404.31 |
| hsa-miR-25-3p | 0.01 | 254.91 | 107.19 | <0.01 | 13.04 | 12.40 | <0.01 | −0.07 | −0.30 | <0.01 | 9,512.39 | 4104.21 |
| hsa-miR-185-5p | <0.01 | 179.77 | 92.53 | <0.01 | 11.25 | 10.46 | <0.01 | 0.04 | −0.06 | <0.01 | 75.59 | 28.86 |
|
hsa-miR-181b-5p | <0.01 | 109.79 | 67.29 | <0.01 | 10.90 | 9.77 | 0.03 | 0.15 | 0.03 | <0.01 | 279.87 | 83.34 |
| hsa-miR-224-5p | 0.03 | 29.75 | 2.54 | <0.01 | 9.89 | 7.99 | <0.01 | 0.70 | 0.18 | 0.04 | 36.37 | 13.50 |
|
hsa-miR-196b-5p | <0.01 | 30.49 | 7.97 | <0.01 | 10.42 | 8.34 | <0.01 | −0.20 | −0.35 | <0.01 | 637.67 | 26.86 |
|
hsa-miR-135b-5p | 0.01 | 32.91 | 2.34 | <0.01 | 11.85 | 9.36 | 0.01 | 0.30 | 0.05 | <0.01 | 66.81 | 6.74 |
| hsa-miR-204-5p | <0.01 | 3.61 | 11.63 | <0.01 | 8.25 | 10.73 | <0.01 | 0.06 | 0.52 | <0.01 | 7.31 | 21.90 |
Differentially expressed miRNAs are
dysregulated in all stages of GC
To evaluate the possible prognostic value of the 11
abnormally expressed miRNAs, the present study analyzed RNA-seq
data from TCGA, which included a cohort of 41 normal tissues, 7
grade I GC tissues, 140 grade II GC tissues and 232 grade III GC
tissues (20). All patients were
staged using the tumor-node-metastasis classification of the
American Joint Committee on Cancer/International Union Against
Cancer from 2009 (21). It was
identified that the expression levels of hsa-miR-15b-5p,
hsa-miR-21-5p, hsa-miR-93-5p, hsa-let-7i-5p, hsa-miR-25-3p,
hsa-miR-185-5p, hsa-miR-181b-5p, hsa-miR-224-5p, hsa-miR-196b-5p
and hsa-miR-135b-5p were significantly upregulated, and
hsa-miR-204-5p expression was significantly downregulated in grade
I, II and III 3 GC tissues compared with the normal controls
(Fig. 2A-K). However, differential
expression levels of the miRNAs were not observed between the
different GC stages. This analysis demonstrated that the abnormally
expressed miRNAs may be associated with GC progression and may
serve as early diagnostic biomarkers. However, the dysregulation of
these miRNAs could not predict the stage of GC.
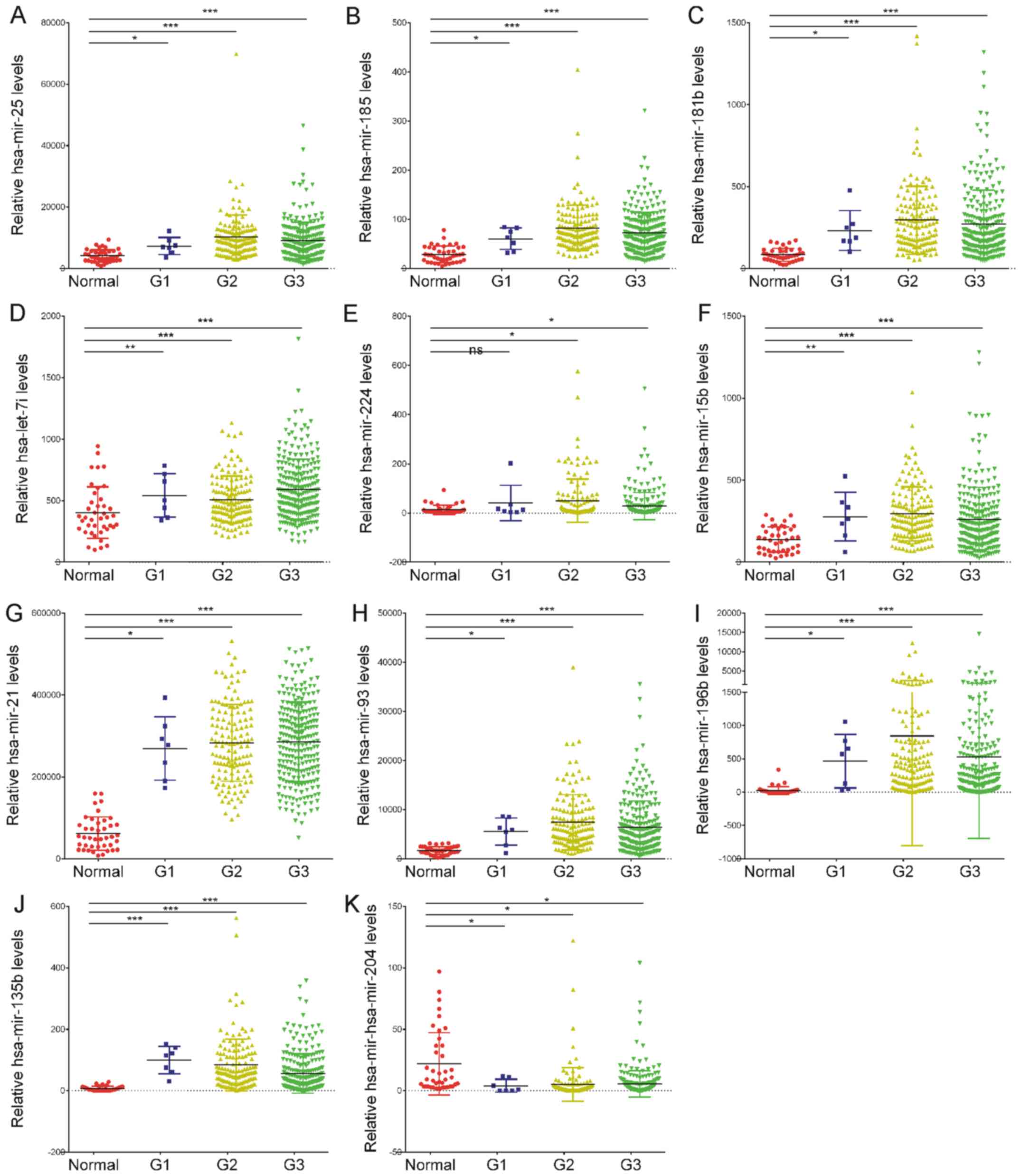 | Figure 2.Dot-plot assay for the expression
levels of the 11 abnormally expressed miRNAs in G1, G2 and G3
gastric cancer tissues compared with the normal controls. (A)
hsa-miR-25-3p, (B) hsa-miR-185-5p, (C) hsa-miR-181b-5p, (D)
hsa-let-7i-5p, (E) hsa-miR-224-5p, (F) hsa-miR-15b-5p, (G)
hsa-miR-21-5p, (H) hsa-miR-93-5p, (I) hsa-miR-196b-5p and (J)
hsa-miR-135b-5p were identified to be significantly upregulated in
G1, G2 and G3 gastric cancer tissues compared with the normal
samples. (K) hsa-miR-204-5p was revealed to be downregulated in G1,
G2 and G3 gastric cancer tissues compared with the normal samples.
*P<0.05, **P<0.01, ***P<0.001. miRNA or miR, microRNA; G1,
grade I; G2, grade II; G3, grade III. |
Construction of a differentially
expressed miRNAs-mRNAs network for GC
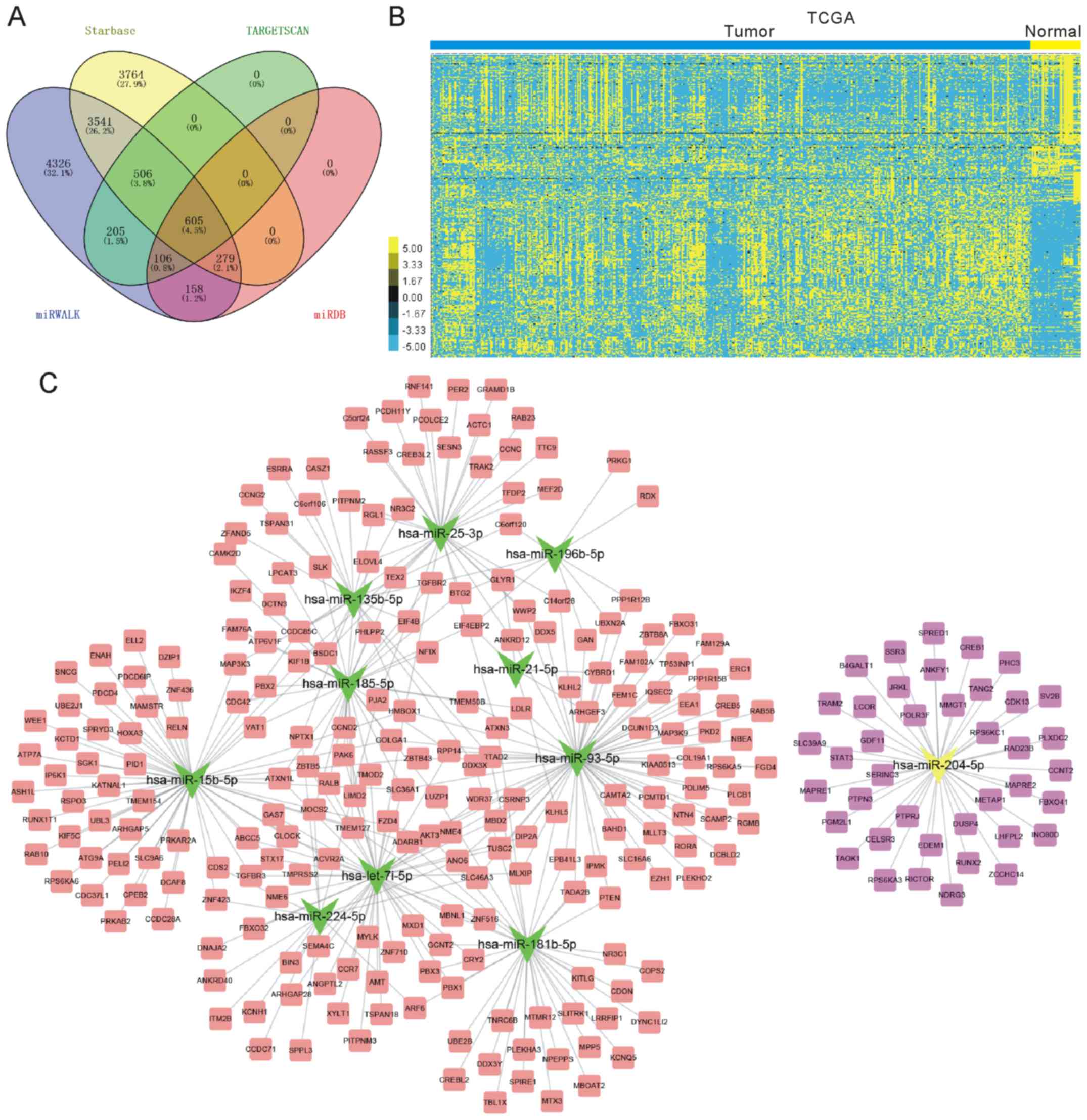
Four different databases were used, including
TargetScan, miRWALK, miRDB and starbase. A total of 605 target
mRNAs were obtained by combining the analyses of these databases
(Fig. 3A). Furthermore, the TCGA
dataset was analyzed to investigate the expression pattern of the
605 target mRNAs in GC (Fig. 3B). In
the present study, the downregulated genes were selected as
potential targets of the upregulated miRNAs and the upregulated
genes were selected as potential targets of the downregulated
miRNAs. As presented in Fig. 3C, a
total of 11 miRNAs and 267 mRNAs were included in this network.
Four miRNAs, including hsa-miR-93-5p, hsa-miR-15b-5p, hsa-let-7i-5p
and hsa-miR-204-5p were identified as key regulators in GC as they
were connected to >40 mRNAs in the network.
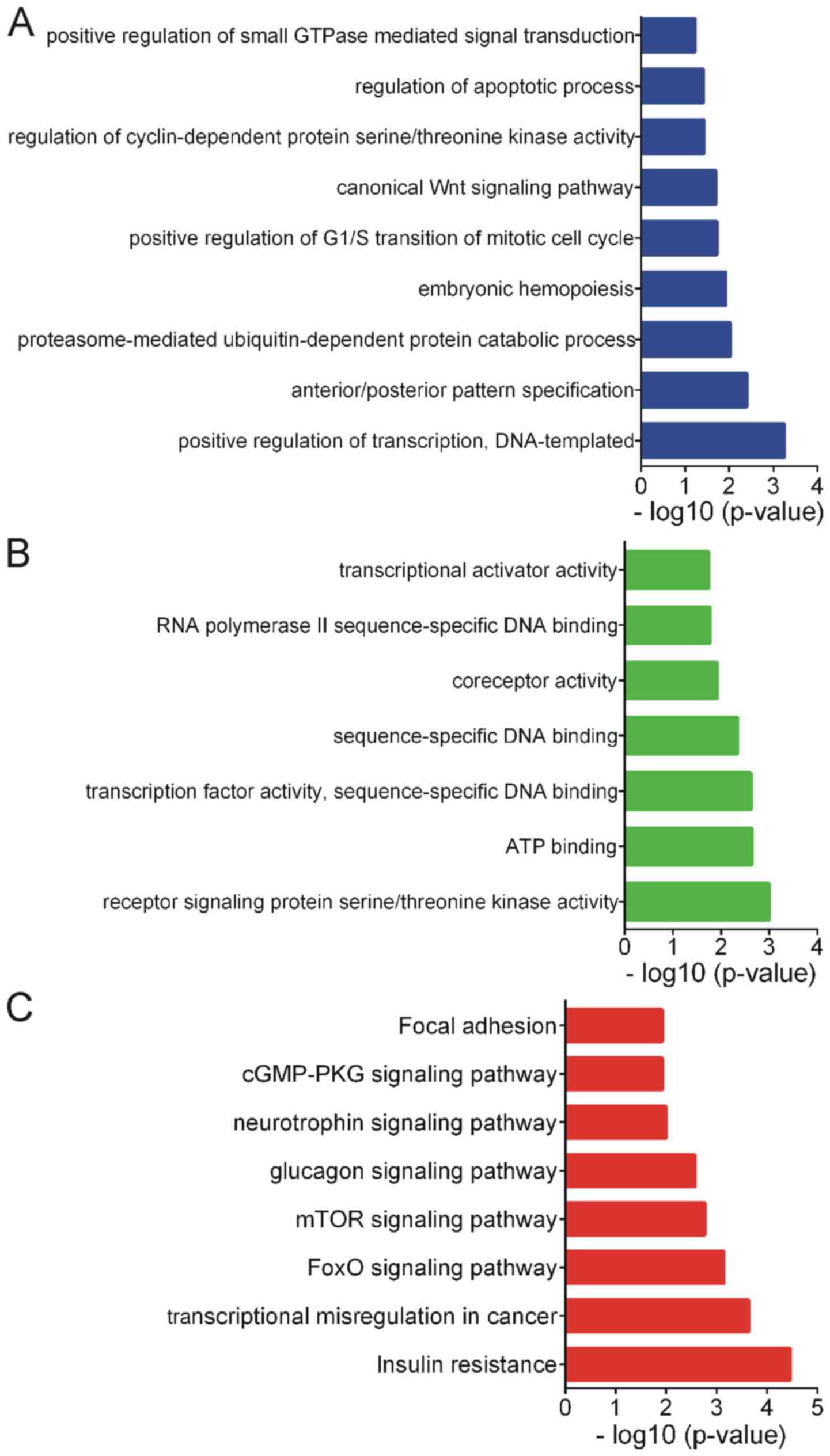
GO analysis of differentially
expressed miRNAs
GO analysis was performed for the differentially
expressed miRNAs using the target mRNAs. We hypothesized that the
potential functions of target mRNAs could reflect the possible
roles of differentially expressed miRNAs. The present study only
presented the GO analysis results for the biological processes and
molecular functions (Fig. 4).
According to the GO analysis, differentially expressed miRNAs were
enriched in ‘positive regulation of transcription’,
‘anterior/posterior pattern specification’, ‘proteasome-mediated
ubiquitin-dependent protein catabolic process’, ‘embryonic
hemopoiesis’, ‘positive regulation of G1/S transition of mitotic
cell cycle’, ‘canonical Wnt signaling pathway’, ‘regulation of
cyclin-dependent protein serine/threonine kinase activity’,
‘regulation of apoptotic process’ and ‘positive regulation of small
GTPase mediated signal transduction’ (Fig. 4A).
In addition, the differentially expressed miRNAs
were involved in regulating several molecular functions, including
‘receptor signaling protein serine/threonine kinase activity’, ‘ATP
binding’, ‘transcription factor activity’, ‘sequence-specific DNA
binding’, ‘sequence-specific DNA binding’ and ‘coreceptor activity’
(Fig. 4B).
KEGG analysis of differentially
expressed miRNAs
KEGG pathway analysis revealed that the
differentially expressed miRNAs predominantly participate in the
regulation of ‘Insulin resistance’, ‘Transcriptional misregulation
in cancer’, ‘FoxO signaling pathway’, ‘mTOR signaling pathway’,
‘Glucagon signaling pathway’, ‘Neurotrophin signaling pathway’ and
‘cGMP-PKG signaling pathway’ (Fig.
4C).
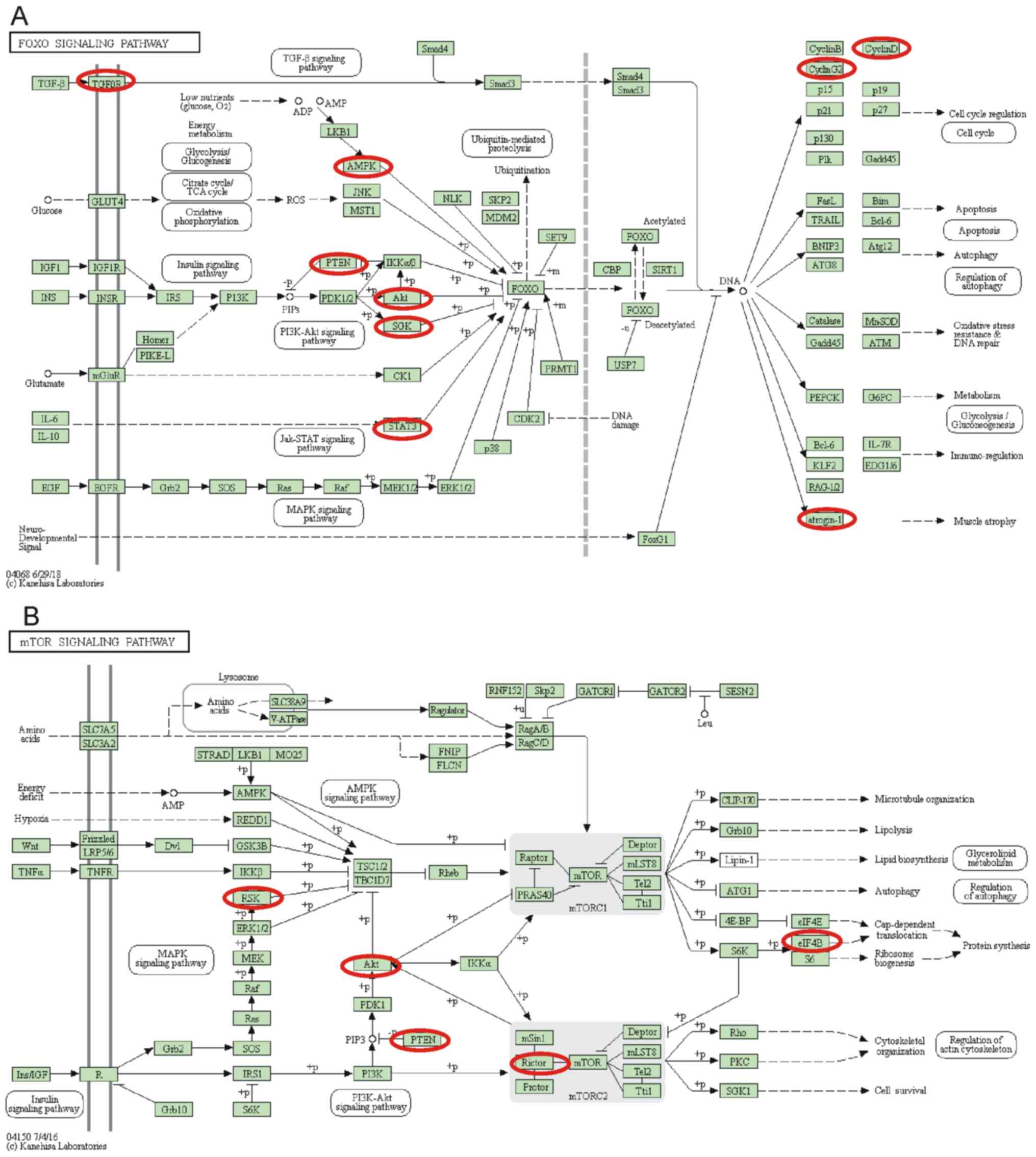
Notably, it was identified that the differentially
expressed miRNAs serve crucial roles in regulating the forkhead box
protein O (FoxO) signaling pathway and the mTOR signaling pathway.
Nine genes in the FoxO signaling pathway, including AKT3, FBXO32,
CCND2, CCNG2, PTEN, PRKAB2, SGK1, STAT3 and TGFBR2, and six genes
in the mTOR signaling pathway, including AKT3, RICTOR, EIF4B, PTEN,
RPS6KA3 and RPS6KA6, were identified to be targets of the
differentially expressed miRNAs (Fig.
5).
Discussion
The most significant challenge for the treatment of
GC is the absence of early diagnostic biomarkers (1). miRNAs are small non-coding RNAs that
demonstrate clinical value in a number of types of cancer (2). Numerous studies have identified certain
differentially expressed miRNAs in GC (22–24);
however, these studies only investigated a small number of miRNAs
and lacked large sample sizes to systemically identify numerous
differentially expressed miRNAs.
Previous studies regarding GC have indicated that
hsa-miR-93 (25), hsa-miR-19a
(26), hsa-miR-20a (27) and hsa-miR-221 (28) might act as oncogenic miRNAs. To
identify differentially expressed miRNAs in GC, the present study
analyzed four public datasets, including GSE23739, GSE26595,
GSE30070 and data from TCGA. A total of 10 miRNAs (hsa-miR-15b-5p,
hsa-miR-21-5p, hsa-miR-93-5p, hsa-let-7i-5p, hsa-miR-25-3p,
hsa-miR-185-5p, hsa-miR-181b-5p, hsa-miR-224-5p, hsa-miR-196b-5p
and hsa-miR-135b-5p) were revealed to be upregulated in GC and
hsa-miR-204-5p was identified to be downregulated in GC, which
suggests these miRNAs serve key roles in GC. To evaluate the
possible prognostic value of these abnormally expressed miRNAs, the
current study analyzed RNA-seq data from TCGA and identified that
the expression levels of the miRNAs were significantly upregulated
or downregulated in grade I, II and III GC tissues compared with
the normal samples. However, differential expression levels of the
miRNAs were not observed between the different GC stages. This
analysis demonstrated that these abnormally expressed miRNAs may be
associated with GC progression and may serve as diagnostic
biomarkers. Furthermore, previous studies have indicated that hsa-
hsa-miR-93 (25), hsa-miR-19a
(26), hsa-miR-20a (27) and hsa-miR-221 (28) may act as oncogenic miRNAs in GC,
which is consistent with the current analysis. The current study
performed a comprehensive analysis to identify key miRNAs in GC
progression by using a series of public datasets. The results may
assist with the identification of biomarkers for the prognosis of
GC.
Considering the important roles of miRNAs in human
disease, a number of studies have investigated the functions and
molecular mechanisms of miRNAs by performing loss or gain of
function assays (29–32). However, these experimental
validations are expensive and time-consuming. Therefore, a number
of effective and feasible computational methods have been developed
to assist with the prediction of potential associations between
disease and miRNAs. For example, Chen et al (33) developed the Path-Based miRNA-Disease
Association model for miRNA-disease association prediction. In
addition, Chen et al (33,34)
developed the Bipartite Network Projection for miRNA-Disease
Association and Hybrid Approach for miRNA-Disease Association
prediction models for miRNA-disease prediction. It can be suggested
that a combination of experimental validations, and effective and
feasible computational prediction methods is a useful strategy to
investigate the potential roles of miRNAs in human disease. The
present study constructed a differently expressed miRNAs-mRNAs
network, and performed GO and KEGG analysis with the target mRNAs.
A total of 11 miRNAs and 267 mRNAs were included in this network.
Four miRNAs, including hsa-miR-93-5p, hsa-miR-15b-5p, hsa-let-7i-5p
and hsa-miR-204-5p were identified as key regulators in GC as there
were connected with >40 mRNAs in the network.
Bioinformatics analysis revealed that the
differentially expressed miRNAs are associated with ‘positive
regulation of transcription’, ‘positive regulation of G1/S
transition of mitotic cell cycle’, ‘canonical Wnt signaling
pathway’ and ‘regulation of apoptotic process’. KEGG pathway
analysis identified that the differentially expressed miRNAs
predominantly participate in regulation of ‘Insulin resistance’,
‘Transcriptional misregulation in cancer’, ‘FoxO signaling
pathway’, ‘mTOR signaling pathway’, ‘Glucagon signaling pathway’,
‘Neurotrophin signaling pathway’ and ‘cGMP-PKG signaling pathway’.
Notably, nine genes associated with the FoxO signaling pathway,
including AKT3, FBXO32, CCND2, CCNG2, PTEN, PRKAB2, SGK1, STAT3 and
TGFBR2, and six genes associated with the mTOR signaling pathway,
including AKT3, RICTOR, EIF4B, PTEN, RPS6KA3 and RPS6KA6, were
identified as targets of the differentially expressed miRNAs.
Previous studies have demonstrated the important roles of FoxO and
mTOR signaling in GC progression. For example, downregulation of
FOXO4 was revealed to inhibit tumor proliferation and metastasis in
GC (35). In addition, mTOR
signaling serves crucial roles in regulating GC proliferation,
apoptosis and metastasis (36,37).
Nadauld et al (38)
identified that TGFBR2 acts an oncogene in diffuse GC. Furthermore,
PTEN has been demonstrated to be involved in regulating apoptotic
cell death, metastasis, angiogenesis and chemoresistance in GC
(39). These results suggest that
dysregulation of these miRNAs have important roles in GC
progression.
In conclusion, the present study identified 11
miRNAs that are significantly differently expressed in GC by
analyzing 1,000 GC samples from four public datasets, including
GSE23739, GSE26595, GSE30070 and data from TCGA. hsa-miR-93-5p,
hsa-miR-15b-5p, hsa-let-7i-5p and hsa-miR-204-5p were revealed as
key regulators in GC. GO analysis and KEGG analysis demonstrated
that the differentially expressed miRNAs are involved in the
positive regulation of transcription, positive regulation of the
G1/S transition in the mitotic cell cycle, the canonical Wnt
signaling pathway, the FoxO signaling pathway and the mTOR
signaling pathway. In summary, the current study may provide
potential new therapeutic and prognostic targets for GC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
CY and YG conceived and designed the study. CY, YZ
and WT developed the methodology. CY and YZ analyzed and
interpreted the data. CY, YZ, WT and YG drafted, reviewed and
revised the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hudler P: Outlook on epigenetic
therapeutic approaches for treatment of gastric cancer. Current
Cancer Drug Targets. 18:65–88. 2018.PubMed/NCBI
|
|
2
|
Ondracek J, Fadrus P, Sana J, Besse A,
Loja T, Vecera M, Radova L, Smrcka M, Slampa P and Slaby O: Global
MicroRNA expression profiling identifies unique MicroRNA pattern of
radioresistant glioblastoma cells. Anticancer Res. 37:1099–1104.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wan X, Pu H, Huang W, Yang S, Zhang Y,
Kong Z, Yang Z, Zhao P, Li A, Li T and Li Y: Androgen-induced
miR-135a acts as a tumor suppressor through downregulating RBAK and
MMP11, and mediates resistance to androgen deprivation therapy.
Oncotarget. 7:51284–51300. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wu S, Gu Y, Huang Y, Wong TC, Ding H, Liu
T, Zhang Y and Zhang X: Novel biomarkers for non-functioning
invasive pituitary adenomas were identified by using analysis of
microRNAs expression profile. Biochem Genet. 55:253–267. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Oze I, Shimada S, Nagasaki H, Akiyama Y,
Watanabe M, Yatabe Y, Matsuo K and Yuasa Y: Plasma microRNA-103,
microRNA-107, and microRNA-194 levels are not biomarkers for human
diffuse gastric cancer. J Cancer Res Clin Oncol. 143:551–554. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yuan M, Wang Z, Hu G, Yang Y, Lv W, Lu F
and Zhong H: A retrospective analysis of hyperthermic
intraperitoneal chemotherapy for gastric cancer with peritoneal
metastasis. Mol Clin Oncol. 5:395–399. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yuan X, Wang S, Liu M, Lu Z, Zhan Y, Wang
W and Xu AM: Histological and pathological assessment of miR-204
and SOX4 levels in gastric cancer patients. Biomed Res Int.
2017:68946752017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wu N, Han Y, Liu H, Jiang M, Chu Y, Cao J,
Lin J, Liu Y, Xu B and Xie X: miR-5590-3p inhibited tumor growth in
gastric cancer by targeting DDX5/AKT/m-TOR pathway. Biochem Biophys
Res Commun. 503:1491–1497. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li X, Zhang Y, Zhang H, Liu X, Gong T, Li
M, Sun L, Ji G, Shi Y, Han Z, et al: miRNA-223 promotes gastric
cancer invasion and metastasis by targeting tumor suppressor
EPB41L3. Mol Cancer Res. 9:824–833. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhu M, Zhou X, Du Y, Huang Z, Zhu J, Xu J,
Cheng G, Shu Y, Liu P, Zhu W and Wang T: miR-20a induces cisplatin
resistance of a human gastric cancer cell line via targeting CYLD.
Mol Med Rep. 14:1742–1750. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Oh HK, Tan ALK, Das K, Ooi CH, Deng NT,
Tan IB, Beillard E, Lee J, Ramnarayanan K, Rha SY, et al: Genomic
loss of miR-486 regulates tumor progression and the OLFM4
antiapoptotic factor in gastric cancer. Clin Cancer Res.
17:2657–2667. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lim JY, Yoon SO, Seol SY, Hong SW, Kim JW,
Choi SH, Lee JS and Cho JY: Overexpression of miR-196b and HOXA10
characterize a poor-prognosis gastric cancer subtype. World J
Gastroenterol. 19:7078–7088. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim CH, Kim HK, Rettig RL, Kim J, Lee ET,
Aprelikova O, Choi IJ, Munroe DJ and Green JE: miRNA signature
associated with outcome of gastric cancer patients following
chemotherapy. BMC Med Genomics. 4:792011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li J, Hou R, Niu X, Liu R, Wang Q, Wang C,
Li X, Hao Z, Yin G and Zhang K: Comparison of microarray and
RNA-Seq analysis of mRNA expression in dermal mesenchymal stem
cells. Biotechnol Lett. 38:33–41. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang J, Meng F, Dai E, Yang F, Wang S,
Chen X, Yang L, Wang Y and Jiang W: Identification of associations
between small molecule drugs and miRNAs based on functional
similarity. Oncotarget. 7:38658–38669. 2016.PubMed/NCBI
|
|
17
|
Liu H, Zhang Q, Lou Q, Zhang X, Cui Y,
Wang P, Yang F, Wu F, Wang J, Fan T3 and Li S: Differential
analysis of lncRNA, miRNA and mRNA expression profiles and the
prognostic value of lncRNA in esophageal cancer. Pathol Oncol Res.
Apr 10–2019.(Epub ahead of print). View Article : Google Scholar
|
|
18
|
He S, Shi J, Mao J, Luo X, Liu W, Liu R
and Yang F: The expression of miR-375 in prostate cancer: A study
based on GEO, TCGA data and bioinformatics analysis. Pathol Res
Pract. 1523752019.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
19
|
He K, Li W, Guan D, Gong M, Ye S, Fang Z,
Huang JF and Lu A: Regulatory network reconstruction of five
essential microRNAs for survival analysis in breast cancer by
integrating miRNA and mRNA expression datasets. Funct Integr
Genomics. Mar 12–2019.(Epub ahead of print). View Article : Google Scholar
|
|
20
|
Yu H, Ye L, Mansel RE, Zhang Y and Jiang
WG: Clinical implications of the influence of Ehm2 on the
aggressiveness of breast cancer cells through regulation of matrix
metalloproteinase-9 expressio. Mol Cancer Res. 8:1501–1512. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Edge SB and Compton CC: The American joint
committee on cancer: The 7th edition of the AJCC cancer staging
manual and the future of TNM. Ann Surg Oncol. 17:1471–1474. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lu R, Zhao G, Yang Y, Jiang Z, Cai J,
Zhang Z and Hu H: Long noncoding RNA HOTAIRM1 inhibits cell
progression by regulating miR-17-5p/PTEN axis in gastric cancer. J
Cell Biochem. 120:4952–4965. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Magalhães L, Quintana L, Lopes DCF, Vidal
AF, Pereira AL, D'Araujo Pinto LC, de Jesus Viana Pinheiro J,
Khayat AS, Goulart LR, Burbano R, et al: APC gene is modulated by
hsa-miR-135b-5p in both diffuse and intestinal gastric cancer
subtypes. BMC Cancer. 18:10552018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu R, Zhang C, Hu Z, Li G, Wang C, Yang
C, Huang D, Chen X, Zhang H, Zhuang R, et al: A five-microRNA
signature identified from genome-wide serum microRNA expression
profiling serves as a fingerprint for gastric cancer diagnosis. Eur
J Cancer. 47:784–791. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liang H, Wang F, Chu D, Zhang W, Liao Z,
Fu Z, Yan X, Zhu H, Guo W, Zhang Y, et al: miR-93 functions as an
oncomiR for the downregulation of PDCD4 in gastric carcinoma. Sci
Rep. 6:237722016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wu Q, Yang Z, An Y, Hu H, Yin J, Zhang P,
Nie Y, Wu K, Shi Y and Fan D: MiR-19a/b modulate the metastasis of
gastric cancer cells by targeting the tumour suppressor MXD1. Cell
Death Dis. 5:e11442014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang Y, Han T, Wei G and Wang Y:
Inhibition of microRNA-17/20a suppresses cell proliferation in
gastric cancer by modulating UBE2C expression. Oncol Rep.
33:2529–2536. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shi J, Zhang Y, Jin N, Li Y, Wu S and Xu
L: MicroRNA-221-3p plays an oncogenic role in gastric carcinoma by
inhibiting PTEN expression. Oncol Res. 25:523–536. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang Z, Li Z, Gao C, Chen P, Chen J, Liu
W, Xiao S and Lu H: miR-21 plays a pivotal role in gastric cancer
pathogenesis and progression. Lab Invest. 88:1358–1366. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wu H, Lin W and Tsai K: Advances in
molecular biomarkers for gastric cancer: miRNAs as emerging novel
cancer markers. Expert Rev Mol Med. 16:e12014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shin VY and Chu K: MiRNA as potential
biomarkers and therapeutic targets for gastric cancer. World J
Gastroenterol. 20:10432–10439. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tchernitsa OI, Kasajima A, Schäfer R,
Kuban RJ, Ungethüm U, Györffy B, Neumann U, Simon E, Weichert W,
Ebert MP and Röcken C: Systematic evaluation of the miRNA-ome and
its downstream effects on mRNA expression identifies gastric cancer
progression. J Pathol. 222:310–319. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen X, Xie D, Wang L, Zhao Q, You Z and
Liu H: BNPMDA: Bipartite network projection for MiRNA-disease
association prediction. Bioinformatics. 34:3178–3186. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen X, Niu Y, Wang G and Yan G: HAMDA:
Hybrid approach for MiRNA-disease association prediction. J Biomed
Inform. 76:50–58. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Su L, Liu X, Chai N, Lv L, Wang R, Li X,
Nie Y, Shi Y and Fan D: The transcription factor FOXO4 is
down-regulated and inhibits tumor proliferation and metastasis in
gastric cancer. BMC Cancer. 14:3782014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Matsuoka T and Yashiro M: The role of
PI3K/Akt/mTOR signaling in gastric carcinoma. Cancers (Basel).
6:1441–1463. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wu YJ, Wong BS, Yea SH, Lu CI and Weng SH:
Sinularin induces apoptosis through mitochondria dysfunction and
inactivation of the pI3K/Akt/mTOR pathway in gastric carcinoma
cells. Mar Drugs. 14(pii): E1422016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Nadauld LD, Garcia S, Natsoulis G, Bell
JM, Miotke L, Hopmans ES, Xu H, Pai RK, Palm C, Regan JF, et al:
Metastatic tumor evolution and organoid modeling implicate TGFBR2
as a cancer driver in diffuse gastric cancer. Genome Biol.
15:4282014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Xu W, Yang Z and Lu N: Roles of PTEN
(Phosphatase and Tensin Homolog) in gastric cancer development and
progression. Asian Pac J Cancer Prev. 15:17–24. 2014. View Article : Google Scholar : PubMed/NCBI
|