Introduction
Acanthus ebracteatus Vahl. is a shrubby herb
commonly found in South Asia. It belongs in the Acanthaceae family
and is also known as sea holly or holly mangrove. Whole plants of
A. ebracteatus have been used as an astringent and
expectorant in Ayurveda (1), whereas
its leaves and seeds are usually boiled as a treatment for coughs
in Malaysia (2). In Thai traditional
medicine, the plant is consumed as an anti-inflammatory agent for
treating arthritis (3). The entire
plant is also boiled and used for treating skin diseases and
healing rash (4). For the skin
treatment, the combination of ethanol extract of A.
ebracteatus with collagen was reported to enhance skin
angiogenesis and also promoted wound closure in a mouse model
(5). Furthermore, hot water extract
of A. ebracteatus was revealed to inhibit cervical cancer
growth and angiogenesis in cell-implanted nude mice (6). However, little is known about the
biological effects of protein hydrolysates or peptides extracted
from A. ebracteatus.
According to a report from the World Health
Organization, the incidence rates of both non-melanoma and melanoma
skin cancer has been increasing over past years. Approximately 2–3
million cases of non-melanoma skin cancer and ~132,000 cases of
melanoma skin cancer are reported worldwide each year (7). Although non-melanoma skin cancer is not
always lethal, surgical treatment is often painful and disfiguring.
On the other hand, malignant melanoma is a major cause of skin
cancer-associated mortality and only early-stage melanomas can be
treated with surgery. More advanced types of melanoma skin cancer
require other treatments, such as immunotherapy, chemotherapy and
radiation therapy. However, these treatments often cause side
effects or development of multi-drug resistance (8), thus alternative or complementary
treatments from natural products are gaining more attention.
Natural products and compounds derived from plants
appear as a potential alternative treatment of cancers as they
possess inhibitory effects against various types of human cancer,
and can perturb cellular signaling pathways, particularly the
activation of apoptosis with minimal effects on normal cells
(9–13). The majority of reported
phytochemicals are flavonoids, carotenoids, terpenoids, vitamins
and certain polyphenoids (8).
Nevertheless, proteins or bioactive peptides containing 3–20 amino
acid units (14,15) have also been reported as potential
cancer treatments due to their high target specificity with low
toxicity, and they are easy to manipulate (16).
Therefore, the present study aimed to evaluate the
anti-non-melanoma skin cancer activity of protein hydrolysate,
partially purified peptides and purified peptides extracted from
the aerial parts of A. ebracteatus using an MTT assay. The
effects of plant extracts on the apoptosis pathway were
investigated using Annexin V. Finally, the effects of plant
extracts on the level of cellular protein expression [p53,
p53-serine 15 phosphorylation (Ser15P), RelA (p65) and Cyclin D1]
were also determined.
Materials and methods
Plant material
The aerial parts of A. ebracteatus in powder
form were purchased from a Thai traditional medicine shop
(Chao-krom-poe, Bangkok, Thailand).
Preparation of protein
hydrolysate
A. ebracteatus powder (250 g) was extracted
with 0.5% sodium dodecyl sulfate (SDS) (Ajax finechem) for 24 h at
37°C in a shaker (Gallenkamp). The extracts were then centrifuged
at 9,100 × g for 30 min at 4°C and protein was precipitated with
80% cold acetone (Avantor) in a ratio of 1:2 (v/v) at −20°C for 24
h. The supernatant was removed via centrifugation at 5,800 × g for
30 min at 4°C. Then, the protein precipitate was dissolved in 0.2 M
sodium acetate buffer (Ajax finechem), pH 4.5 and digested with
pepsin (Sigma-Aldrich; Merck KGaA) in a ratio of 1:25 (w/w) for 24
h at 37°C followed by heat inactivation for 10 min at 60–70°C.
Following this, this protein hydrolysate was centrifuged at 5,800 ×
g for 30 min at 4°C and pH was adjusted from 6.8 to 7.0 followed by
protein filtration through a membrane with a molecular weight
cut-off of 3 kDa (GE Healthcare). Finally, the protein content was
determined using Bradford assay (Bio-Rad Technologies) according to
the manufacturer's protocol.
Cell culture and maintenance
Vero (normal kidney cells, ATCC® CCL-81™)
and A431 (skin cancer cells; ATCC® CRL-1555™) cells were
cultured in DMEM (Gibco; Invitrogen; Thermo Fisher Scientific,
Inc.) supplemented with 10% (v/v) heat inactivated fetal bovine
serum (Gibco; Invitrogen; Thermo Fisher Scientific, Inc.) and 1%
(v/v) antimycotic-antibiotic solution (Gibco; Invitrogen; Thermo
Fisher Scientific, Inc.). Both cell lines were cultured in 75
cm2 sterile flasks (SPL) at 37°C under 5% CO2
atmosphere (Heal force).
All cell lines were cultures in 75 cm2
sterile tissue culture flasks at 37°C with 5% CO2 in
DMEM supplemented with 10% heat inactivated FBS (10% v/v). Cells
were regularly passaged to maintain exponential growth. Basic cell
culture techniques were performed as previously described (17). All cells were checked for mycoplasma
contamination using PCR and staining with Hoechst 33258. All cell
lines were confirmed to be mycoplasma-free cultures.
Cell viability assay (MTT assay)
Cell viability was evaluated using MTT assay
(Invitrogen; Thermo Fisher Scientific, Inc.) as previously
described (18). Briefly,
1.5×103 cells were seeded in 96 well-microplate (SPL)
and incubated under 5% CO2 at 37°C for 24 h. Cells were
them treated with various final concentrations of A.
ebracteatus protein hydrolysate from 0.00 (water only), 3.33,
33.33, 333.33 and 1,666.67 ng protein/ml. For analyzing the
cytotoxicity of the partially purified protein hydrolysate, cells
were treated with 425.9 ng protein/ml [the half inhibitory
concentration (IC50) value of the A. ebracteatus
protein hydrolysate against A431] of each fraction. For evaluating
the cytotoxicity of the purified peptides, cells were treated with
different concentrations of single or combined peptides, ranging
from 100–500 µM. Following the addition of protein hydrolysates or
peptides, cells were incubated for 5 days without changing the
media nor substituting the protein hydrolysates or peptides.
Treated cells were then subjected to MTT assay according to the
manufacturer's protocol.
High performance liquid chromatography
(HPLC) analysis
HPLC analysis with a UV detector (RP-HPLC) was
performed to partially purified peptides from A. ebracteatus
protein hydrolysate. The column employed was an ODS-2 HYPERSIL C18;
250×4.6 mm, 5 µm particle size (Thermo Fisher Scientific, Inc.). A
gradient mobile phase of 0–50 min was applied, with HPLC-grade 0.1%
trifluoroacetic acid (TFA) (Thermo Fisher Scientific, Inc.),
acetonitrile (Thermo Fisher Scientific, Inc.) and water (90:10
v/v). The injected sample volume was 100 µl (17.05 µg protein/ml)
and the flow rate was 1.00 ml/min. Detection was performed at 220
nm. Each fraction was collected using a fraction collector
(Amersham Pharmacia Biotech). Finally, fractions were lyzed in 50
mM Tris/HCl buffer (Thermo Fisher Scientific, Inc.) and then
lyophilized using a freeze dryer (LaboGene).
Liquid chromatography tandem mass
spectrometry (LC-MS/MS) analysis
The peptide sequences were identified using
LC-MS/MS. Briefly, the peptides were acidified with 0.1% (v/v)
formic acid and subjected to an Ultimate 3000 capillary LC system
(Dionex). Peptide separation was performed using an Acclaim PepMap
RSLC 75 µm ×15 cm nanoviper C18 with a 2 µm particle sizes and a
100 Å pore size (Thermo Fisher Scientific, Inc.) at flow rate of
300 nl/min. Mobile phase A consisted of 2% acetonitrile and 0.1%
formic acid in HPLC grade water and mobile phase B consisted of
0.1% formic acid in HPLC grade acetonitrile. The instrument was
controlled by Hystar software. DataAnalysis™ and Biotools software
version 3.2 were used for the data interpretation and de
novo sequencing.
Peptide synthesis and bioinformatic
analysis
Four peptide sequences (Table I) were commercially synthesized by
GenScript® for >95% purity. The pI and molecular
weight (MW) of each peptide were calculated using a peptide
property calculator (19).
 | Table I.Peptide sequences identified from the
FR1 of A. ebracteatus by using LC-MS/MS. |
Table I.
Peptide sequences identified from the
FR1 of A. ebracteatus by using LC-MS/MS.
| Peptide name | Sequence | pI | Molecular weight
(g.mol−1) |
|---|
| AE1 | KVGNAADRV | 9.9 | 929.03 |
| AE2 | NDLSGDNSVRW | 3.71 | 1,262.29 |
| AE3 | VLNPNLEPPPNP | 1.1 | 1,300.46 |
| AE4 | WAGEEVKAPHW | 5.26 | 1,309.43 |
Apoptosis study
Cells (1×105 cells) were seeded in 24
well-microplates (SPL) and incubated in the presence of 5%
CO2 at 37°C for 24 h. Cells were then treated with 425.9
ng protein/ml of the partially purified peptide. Cells were
incubated in the presence of 5% CO2 at 37°C for 3 days.
Following incubation, cells were washed twice with cold phosphate
buffer saline (PBS) and then harvested. For the positive control,
cells were induced with 50 µg/ml dihydrochloride hydrate
(Sigma-Aldrich; Merck KgaA) for 30 min. Cells were then stained
using Annexin V and dead cells assay kit (Merck Millipore) in the
dark for 15 min. Finally, cells were analyzed using a flow
cytometer (Merck Millipore).
Western blot analysis
Cells (1×105 cells) were seeded in
24-well microplates and incubated at 5% CO2, 37°C for 24
h. Cells were then treated with 425.9 ng protein/ml of the
partially purified peptides and incubated in the presence of 5%
CO2, 37°C for 5 days. Protein extraction was performed
using passive lysis buffer (PLB) (Promega), and the protein
concentration was determined using a Bradford assay. For the
immunoblotting step, the protein was firstly separated using
NuPAGE® Bis-Tris gel (Invitrogen; Thermo Fisher
Scientific, Inc.) and transferred on to PVDF membrane (Merck
Millipore). Then, the membrane was blocked with 5% skimmed milk in
TBS-T for 1 h. Next, the membrane was washed with TBS-T buffer and
then incubated overnight using primary antibodies that were diluted
in 0.01% BSA in TBS-T. The dilution ratios for anti-p53
(Invitrogen; Thermo Fisher Scientific, Inc.), anti-RelA (p65),
anti-Cyclin D1 (Santa Cruz Biotechnology) were equal to 1:1,000
(v/v); for anti-alpha-tubulin (Invitrogen; Thermo Fisher
Scientific, Inc.) was equal to 1:1,500 (v/v); and anti-Ser15P was
equal to 1:300 (Abcam). The membrane was then washed and incubated
with horseradish peroxidase-conjugated secondary antibodies (Abcam)
for 1 h. Finally, the ECL substrate (GE Healthcare) was added onto
a membrane and analyzed using a molecular imager (Bio-Rad
Technologies).
Statistical analysis
An unpaired Student's t-test was performed using
GraphPad QuickCalcs (GraphPad software). Unless any statistically
significant differences between the mean of three or more
independent groups were compared, one-way ANOVA with Dunnett's T3
analysis was performed using IBM SPSS software. P<0.05 was
considered to indicate a statistically significant result.
Results
Protein hydrolysate of A. ebracteatus
significantly inhibits the cell viability of non-melanoma skin
cancer cells in a dose dependent manner
Natural bioactive peptides are primarily released
from their precursor proteins by digestive enzymes during
gastrointestinal digestion (14).
Therefore, the production of bioactive peptides in the present
study mimicked the human digestive system in the body by digesting
the extracted lysates with the enzyme pepsin. The MTT assay was
then performed to evaluate the anti-non-melanoma skin cancer
activity of A. ebracteatus protein hydrolysate. The
non-melanoma skin cancer cells (A431) were treated with various
concentrations of protein hydrolysate ranging from 0–1,666.7 ng
protein/ml. Notably, Vero cells, an epithelial cell line from
African green monkeys were used to represent normal cells in the
present study. As a result, A. ebracteatus protein
hydrolysate significantly inhibited the cell viability of A431 in a
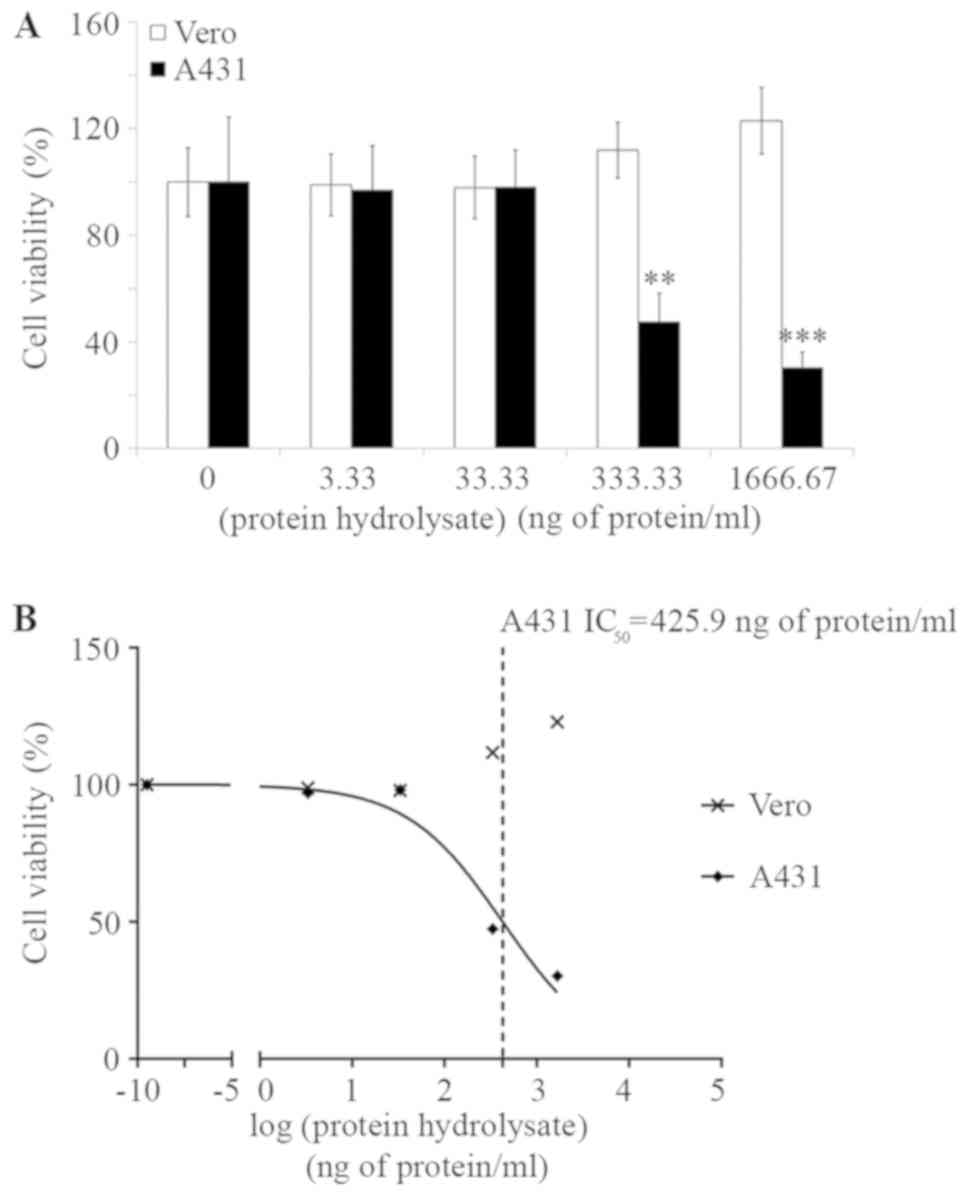
dose dependent manner (Fig. 1A). By
treating cells with 333.3 ng protein/ml, the cell viability of A431
was only 47.5±10.8% (P<0.001) without affecting the Vero cells
(Fig. 1A). The IC50 of
A. ebracteatus protein hydrolysate against A431 cells
calculated by GraphPad was equal to 425.9 ng protein/ml (Fig. 1B). Furthermore, A. ebracteatus
protein hydrolysate was revealed to induce anticancer activities
against a board range of cancer cell lines (Fig. S1).
Subsequently, partially purified peptides of the
A. ebracteatus protein hydrolysate were then investigated in
order to identify bioactive fractions.
In vitro identification of partially
purified peptides against non-melanoma skin cancer from A.
ebracteatus
To partially purify protein hydrolysate and identify
fractions that possess anti-epidermoid cancer activities, a reverse
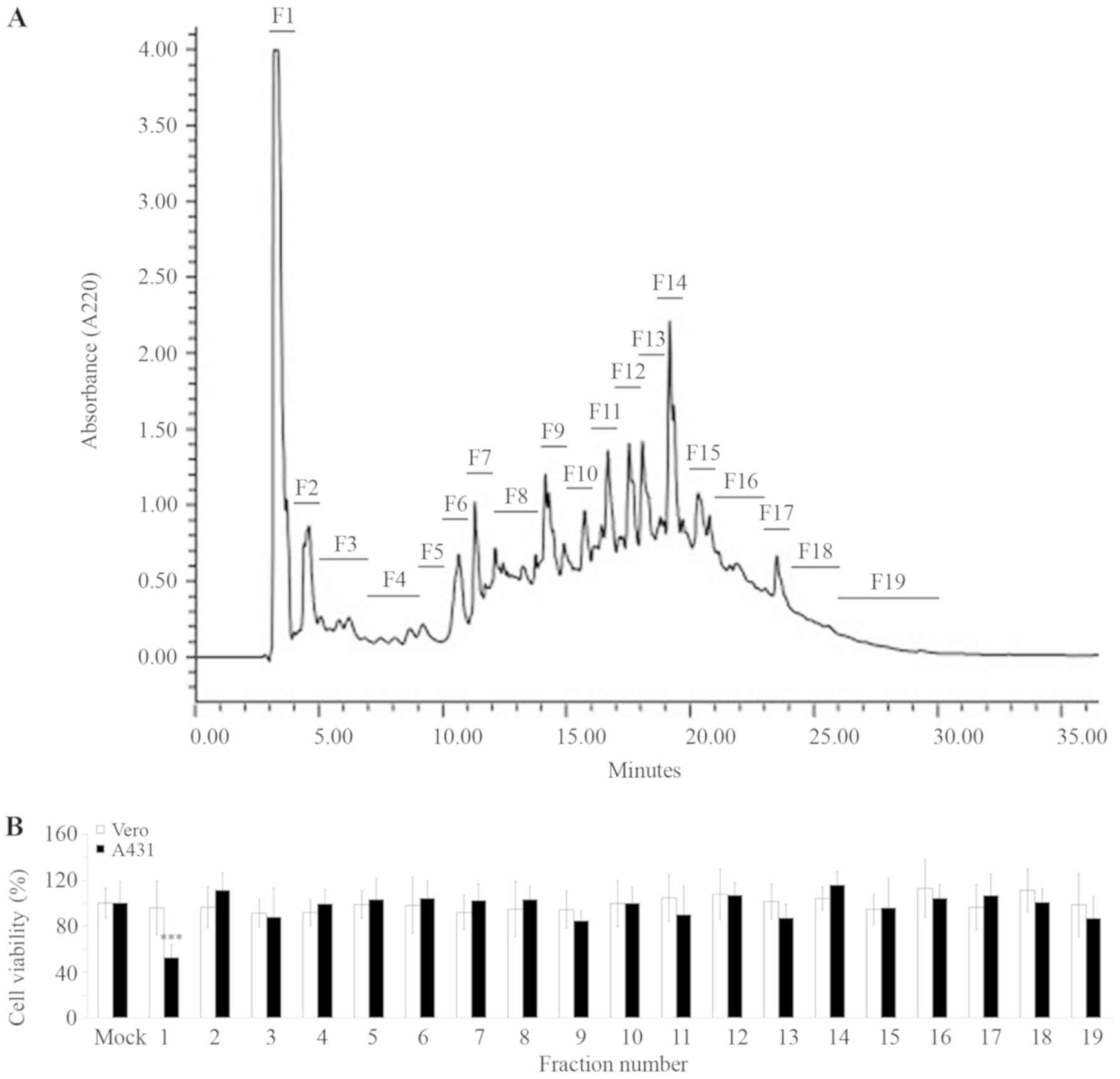
phase (RP) HPLC with a UV detector was performed. The chromatogram
indicated several major and minor peaks, which were sorted into 19
fractions (Fig. 2A). According to
the IC50 value of A. ebracteatus protein
hydrolysate against A431 cells (Fig.
1B), the concentration at 425.9 ng protein/ml of each fraction
was then administered to the A431 cells. After 5 days of
incubation, the cells were subjected to an MTT assay. The results
demonstrated that the majority of the fractions had no inhibitory
effects on the cells (Fig. 2B).
Nevertheless, the fraction number 1 (FR1) demonstrated
approximately 53±11% inhibition against A431 cell viability
(P<0.0001) without affecting the Vero normal cells (Fig. 2B). Therefore, the FR1 was selected
for further analysis to identify their potential peptide sequences
using LC-MS/MS and de novo analyses.
Evaluation of anti-epidermoid cancer
activity of obtained peptides from aerial part of A. ebracteatus
using A431 cells
The LC-MS/MS and bioinformatic analyses on the FR1
identified four peptide sequences; these were named AE1-AE4, as
presented in Table I. Excluding
peptide AE1, peptides AE2-AE3 had a molecular weight of ~1,300
g/mol and pI values <7. On the other hand, AE1 was the smallest
size about 929 g/mol and had a pI of 9.9.
These peptides were synthesized and their
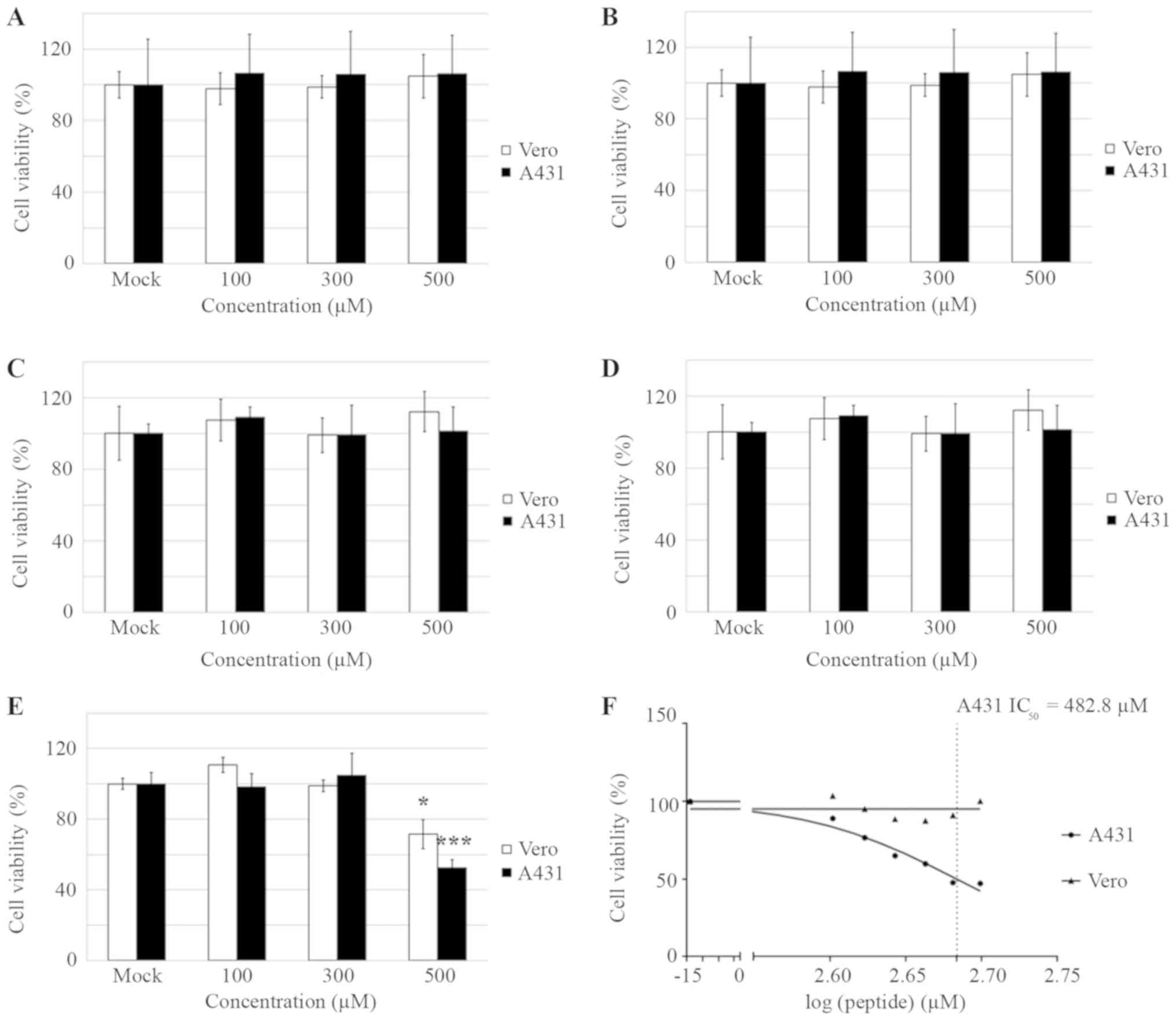
anti-epidermoid cancer activities were evaluated using an MTT assay
with A431 cells. Subsequently, A431 cells were treated with single
or combined peptides ranging from 100–500 µM. After 5 days of
incubation, the cells were subjected to the MTT assay. The results
revealed that each single peptide had no cytotoxicity effects on
either A431 or Vero cells (Fig.
3A-D). Nevertheless, 500 µM of the combined treatment with the
4 peptides (in the same ratio, 125 µM of each peptide) revealed ~35
and 50% inhibition against Vero and A431 cells, respectively.
(Fig. 3E). These results implied
that the mixture of the 4 peptides provided synergistic cytotoxic
effects against the cultured cells. Further MTT assays were then
performed and the IC50 of the combined peptides against
A431 cells was equal to 482.8 µM (Fig.
3F). Despite this IC50 demonstrating no cytotoxic
effect against the normal cells (Fig.
3F), the value was considerably high. Subsequently, the
partially purified FR1 (Fig. 2B) was
considered and investigated to identify its effect on the apoptosis
pathway.
Partially purified protein hydrolysate
extracted from A. ebracteatus significantly induces the apoptosis
pathway
To investigate the effects of partially purified
peptides on the apoptosis pathway, A431 cells were treated with
425.9 ng protein/ml of the FR1 and incubated for 3 days rather than
5 days to ensure that enough cell population were presented for
testing the apoptotic assay. After 3 days of incubation, cells
defined as positive control were treated with 50 µg/ml
dihydrochloride hydrate, and apoptosis inducer. All cells were then
stained using the Muse® Annexin V and Dead Cell kit that
uses a single reagent and two stains to reliably stain and
differentiate live, dead and apoptotic cells. The assay relies on
the binding of fluorescently labeled Annexin V to
phosphatidylserine (PS) molecules, which translocate to the outer
surface of the cell membrane upon the onset of apoptosis (20). 7-Amino-Actinomycin D (7-AAD) is a
fluorescence indicator that is used as a dead cell marker. It also
binds selectively to GC regions of DNA (21). Following staining, apoptotic cells
were analyzed using a flow cytometer.
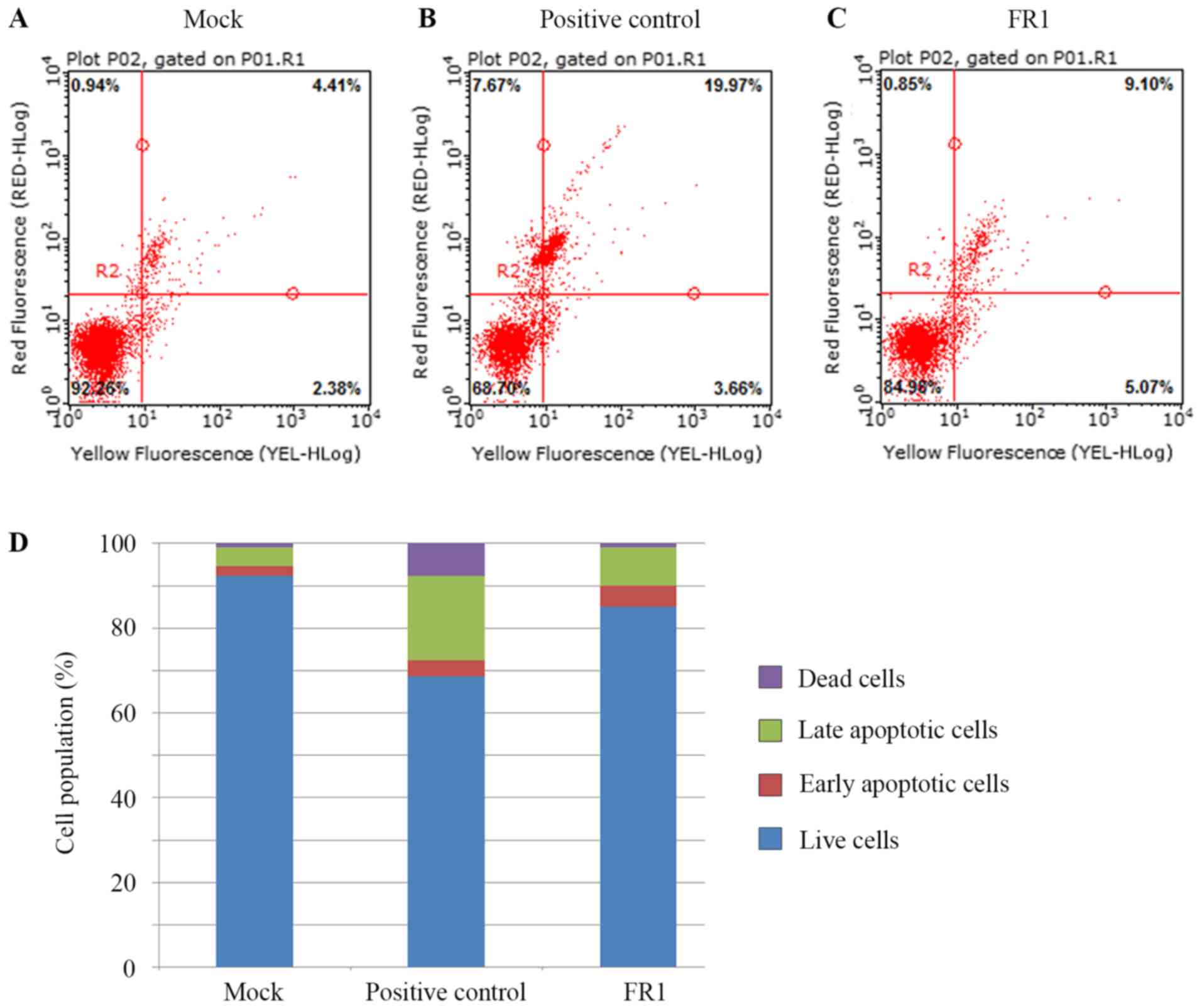
The result from the mock analysis revealed that the
majority of the A431 cells were alive (~90%) with 5% of naturally
occurring late apoptotic cells and 1% of naturally occurring early
apoptotic cells and 1% of cell dead (Fig. 4A and D). The addition of the
apoptotic inducer increased the number of late apoptotic cells by 3
fold. However, the apoptotic inducer only significantly increased
numbers of late apoptotic cells (Fig. 4B
and D) whereas the effects of FR1 increased numbers of early
apoptotic cells by 2 fold, and moderately increased late apoptotic
cells by ~1.5 fold when compared with mock (Fig. 4C and D). Therefore, these results
suggested that the FR1 contains bioactive compounds that may
inhibit the viability of cancer cells by promoting the apoptosis
pathway in A431 cells. Subsequently, the effects FR1 on the protein
expression of p53, Ser15P, NF-κB (RelA/p65) and cyclin D1 were
further investigated.
Effects of partially purified protein
hydrolysate extracted from A. ebracteatus on protein expression of
p53, NF-κB and cyclin D1
A431 cells are reported to carry a p53 mutation at
codon 273 (p53-R273H) (22,23). This R273H mutation affected the
DNA-binding ability of p53, thus, p53-R273 could not act as a
transcription factor (24).
Furthermore, p53-R273H has been reported to induce drug resistance
(25). Therefore, whether the FR1
could restore p53 transcriptional activity in A431 cells or not was
further investigated via assessing the protein expression levels of
p53 and Ser15P. Furthermore, the effects of partially purified
peptide FR1 were also determined via the protein expression levels
of another important transcription factor, RelA (p65), which is
involved in the regulation of cell apoptosis, the cell cycle and
carcinogen transformation (26).
Finally, the effects of FR on the protein expression of cyclin D1,
an important regulator of cell cycle progression (27), was also investigated.
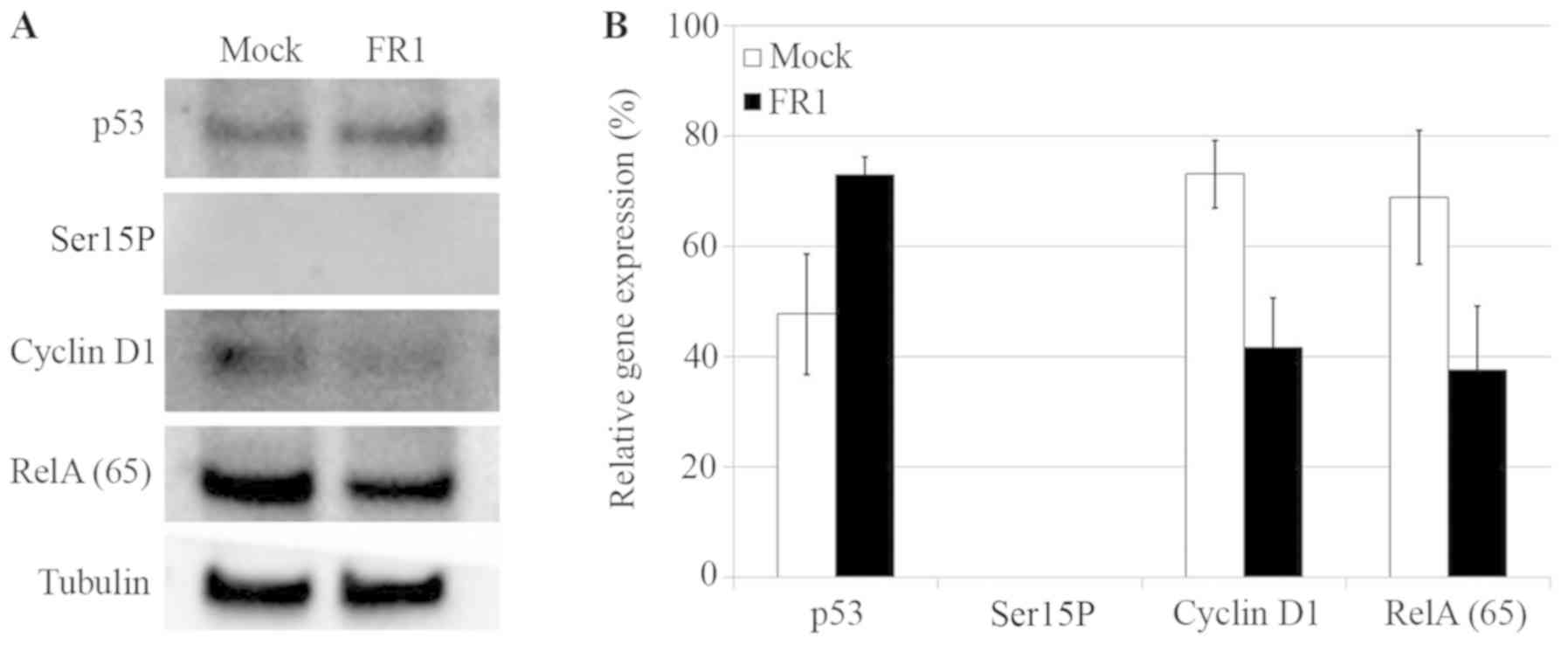
The results from the western blot analysis indicated
that partially purified peptide FR1 from A. ebracteatus may
increase the expression of p53. On the other hand, FR1 decreased
the protein expression of RelA (p65) and cyclin D1 (Fig. 5A and B). Ser15P was barely detected
in mock as its expression is often very low in carrying p53 mutants
such as cancer cells (28–30); however, FR1 had no effect on the
expression levels of Ser15P (Fig. 5A and
B).
Discussion
Previously, the stem of A. ebracteatus was
indicated to contain bioactive polysaccharides such as galactose,
3-O-methylgalactose and arabinose, which could strongly affect the
complement system (31). In
addition, the results from the present study suggested that the
protein hydrolysate of A. ebracteatus (<3 kDa) (Fig. 1) and partially purified protein
hydrolysate, FR1, possessed anti-skin cancer activity (Fig. 2). New peptides derived from aerial
part of A. ebracteatus were also identified (Table I). Although a single peptide could
not provide potent cytotoxic effects on the epidermoid cancer cell
line, it was demonstrated that the combined peptides resulted in
the synergistic effects against cell viability of A431 cells
(Fig. 3E and F). Similarly, previous
studies have reported that the combination of bioactive peptides
displayed a synergism of biological activities. For example, the
effects of the combined two antimicrobial peptides, magainin-2 and
PGLa were synergistic against E. coli and human melanoma
cells (32). Co-administration
therapy between pro-apoptosis peptide (kla-TAT) and the cationic
anticancer peptide (HPRP-Al) also drastically increased anticancer
activities against cancer cell lines (33). In the present study it was
hypothesized that the synergistic effects from mixtures of several
polypeptides, oligopeptides and free amino acids are crucial for
the anticancer activity of A. ebracteatus. Notably,
temperatures ranging from 60–70°C were used for inactivating pepsin
activity. This high temperature is likely to denature the majority
of temperature sensitive polypeptides or oligopeptides. Therefore,
it was proposed that FR1 contains oligopeptides that are relatively
stable at temperatures around 60–70°C.
In addition, p53 is a protein that serves an
important role in several cellular mechanisms. Elevation of the p53
protein can promote apoptosis in cancer cells. In the p53 signaling
pathway, p53 can regulate downstream targets resulting in
inhibition of cyclin D1, which leads to inhibition of cell cycle
progression (34). In particular,
A431 cells carry mutant p53 with a missense mutation (R273H) at a
hotspot for DNA binding (23).
Mutant versions of p53, such as p53-R273H, have been revealed to
lose transcriptional activity, but maintain the ability to enhance
tumor progression, metastasis and drug resistance (35,36).
Therefore, restoration of p53 transcriptional activity and
depletion of mutant p53 may be potential strategies to combat
different types of cancer. For example, styrylquinazoline compound
(CP-31398) could induce mitochondrial translocation of mutant
p53-R273H in A431 cells, resulting in cytochrome c release
and the induction of apoptosis (37). On the other hand, geldanamycin or
Hsp90 inhibitors may promote the degradation of different varieties
of p53 mutants, such as p53-R175H, p53-L194F and p53-R273H
(38). In the present study,
partially purified peptide FR1 demonstrated the ability to induce
apoptosis (Fig. 4). Although FR1
could increase expression levels of p53, it could not upregulate
the expression of Ser15P (Fig. 5).
Therefore, it was hypothesized that FR1 could induce apoptosis
through other proteins, such as NF-κB and associated family
members.
In cancer cells, NF-κB is also known as a
transcriptional factor, which plays an important role in cell
proliferation, cell survival, the cell cycle and cell death
(39). Thus, downregulation of NF-κB
may inhibit proliferation and promote apoptosis of cancer cells
(40,41). A number of studies have demonstrated
that natural products from plants could alter the protein
expression levels of NF-κB. For example, crude extracts of C.
formosum demonstrated the activation of protein expression of
p53 and downregulation of NF-κB and cyclin D1 proteins (42). Partially purified peptides from
Gloriosa superba rhizome could significantly inhibit the
cell viability of colon cancer cell line SW620, by inducing
apoptosis signaling via upregulation of p53 and downregulation of
(43). In the present study, FR1
decreased expression levels of RelA (p65) (Fig. 5), which is important for
NF-κB-mediated gene transactivation (26). Beg et al (44) reported that RelA (p65)-deficient mice
die during embryonic development due to apoptosis of hepatocytes.
It was therefore proposed that FR1 is likely to induce apoptosis
via the downregulation of NF-κB and also leads to the depletion of
cyclin D1 expression (Fig. 5) as
NF-κB promotes the cell-cycle transition from the G1 to
S phase, and cyclin D1 is a downstream signaling molecule of NF-κB
(26). Nevertheless, investigation
on protein expression of nuclear or chromatin-bound NF-κB should be
performed in the future studies because it is a better indicator of
NF-κB-mediated transcription.
In conclusion, the present study demonstrated that
the partially purified peptide FR1 from aerial parts of A.
ebracteatus possessed anti-epidermoid cancer activity. FR1
induced apoptosis via the downregulation of RelA (p65) protein
expression resulting in decreased cyclin D1 expression and
upregulation of p53, but not Ser15P. Therefore, FR1 has the
potential for further development as a therapeutic mechanism
against anti-non-melanoma skin cancer.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank the Department of
Biochemistry, Faculty of Science, Kasetsart University (Bangkok,
Thailand) for providing laboratory equipment.
Funding
The present study was supported by Kasetsart
University Research and Development Institute (grant no.
Vor-Tor-Dor 169.58) and the Faculty of Science, Kasetsart
University (grant no. RFG 1–6).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
AK carried out all experiments and drafted the
manuscript. DJ performed western blot analysis and revised the
manuscript. OR performed LC-MS/MS, analyzed and identified peptide
sequences and also drafted the manuscript. NPTT conceived the idea,
designed and coordinated the study and prepared the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Subudhi HN, Choudhury BP and Acharya BC:
Potential medicinal plants from Mahanadi delta in the state of
Orissa. J Econ Taxon Bot. 16:479–487. 1992.
|
|
2
|
Perry LM and Metzger J: Medicinal plants
of East and Southeast Asia: Attributed properties and uses.
Bibliography. 2:447–493. 1980.
|
|
3
|
Laupattarakasem P, Houghton PJ, Hoult JR
and Itharat A: An evaluation of the activity related to
inflammation of four plants used in Thailand to treat arthritis. J
Ethnopharmacol. 85:207–215. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pongboonrod S: Medicinal Plants of East
and Southeast Asia Attributed Properties and Use. Kasem Bunnakit
Publishing; Bangkok: 1971
|
|
5
|
Somchaichana J, Bunaprasert T and Patumraj
S: Acanthus ebracteatus Vahl. ethanol extract enhancement of
the efficacy of the collagen scaffold in wound closure: A study in
a full-thickness- wound mouse model. J Biomed Biotechnol.
2012:7545272012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mahasiripanth T, Hokputsa S, Niruthisard
S, Bhattarakosol P and Patumraj S: Effects of Acanthus
ebracteatus Vahl on tumor angiogenesis and on tumor growth in
nude mice implanted with cervical cancer. Cancer Manag Res.
4:269–279. 2012.PubMed/NCBI
|
|
7
|
WHO: World Health Statistics, . WHO Press;
Geneva: 2018
|
|
8
|
Chinembiri TN, du Plessis LH, Gerber M,
Hamman JH and du Plessis J: Review of natural compounds for
potential skin cancer treatment. Molecules. 19:11679–11721. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sarkar FH, Li Y, Wang Z and Kong D:
Cellular signaling perturbation by natural products. Cell Signal.
21:1541–1547. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Banjerdpongchai R, Wudtiwai B and Sringarm
K: Cytotoxic and apoptotic-inducing effects of purple rice extracts
and chemotherapeutic drugs on human cancer cell lines. Asian Pac J
Cancer Prev. 14:6541–6548. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cho JJ, Cho CL, Kao CL, Chen CM, Tseng CN,
Lee YZ, Liao LJ and Hong YR: Crude aqueous extracts of Pluchea
indica (L.) Less. Inhibit proliferation and migration of cancer
cells through induction of p53-dependent cell death. BMC Complement
Altern Med. 12:2652012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sreelatha S, Jeyachitra A and Padma PR:
Antiproliferation and induction of apoptosis by Moringa oleifera
leaf extract on human cancer cells. Food Chem Toxicol.
49:1270–1275. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nourazarian SM, Nourazarian A, Majidinia M
and Roshaniasl E: Effect of root extracts of medicinal herb
glycyrrhiza glabra on HSP90 gene expression and apoptosis in the
HT-29 colon cancer cell line. Asian Pac J Cancer Prev.
16:8563–8566. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shahidi F and Zhong Y: Bioactive peptides.
J AOAC Int. 91:914–931. 2008.PubMed/NCBI
|
|
15
|
Hou Y, Wu Z, Dai Z, Wang G and Wu G:
Protein hydrolysates in animal nutrition: Industrial production,
bioactive peptides, and functional significance. J Anim Sci
Biotechnol. 8:242017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Marqus S, Pirogova E and Piva TJ:
Evaluation of the use of therapeutic peptides for cancer treatment.
J Biomed Sci. 24:212017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bonifacino JS, Dasso M, Harford JB,
Lippincott- Schwartz J and Yamada KM: Current Protocols in Cell
Biology. John Wiley; New York: 1998
|
|
18
|
Waiyaput W, Payungporn S, Issara-Amphorn J
and Panjaworayan NT: Inhibitory effects of crude extracts from some
edible Thai plants against replication of hepatitis B virus and
human liver cancer cells. BMC Complement Altern Med. 12:2462012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lear S and Cobb SL: Pep-Calc.com: A set of
web utilities for the calculation of peptide and peptoid properties
and automatic mass spectral peak assignment. J Comput Aided Mol
Des. 30:271–277. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Schutte B, Nuydens R, Geerts H and
Ramaekers F: Annexin V binding assay as a tool to measure apoptosis
in differentiated neuronal cells. J Neurosci Methods. 86:63–69.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Schmid I, Krall WJ, Uittenbogaart CH,
Braun J and Giorgi JV: Dead cell discrimination with
7-amino-actinomycin D in combination with dual color
immunofluorescence in single laser flow cytometry. Cytometry.
13:204–208. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Park DJ, Nakamura H, Chumakov AM, Said JW,
Miller CW, Chen DL and Koeffler HP: Transactivational and DNA
binding abilities of endogenous p53 in p53 mutant cell lines.
Oncogene. 9:1899–1906. 1994.PubMed/NCBI
|
|
23
|
Kwok TT, Mok CH and Menton-Brennan L:
Up-regulation of a mutant form of p53 by doxorubicin in human
squamous carcinoma cells. Cancer Res. 54:2834–2836. 1994.PubMed/NCBI
|
|
24
|
Rivlin N, Koifman G and Rotter V: p53
orchestrates between normal differentiation and cancer. Semin
Cancer Biol. 32:10–17. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wong RP, Tsang WP, Chau PY, Co NN, Tsang
TY and Kwok TT: p53-R273H gains new function in induction of drug
resistance through down-regulation of procaspase-3. Mol Cancer
Ther. 6:1054–1061. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen F, Castranova V and Shi X: New
insights into the role of nuclear factor-kappaB in cell growth
regulation. Am J Pathol. 159:387–397. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Alao JP: The regulation of cyclin D1
degradation: Roles in cancer development and the potential for
therapeutic invention. Mol Cancer. 6:242007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang W, Takimoto R, Rastinejad F and
El-Deiry WS: Stabilization of p53 by CP-31398 inhibits
ubiquitination without altering phosphorylation at serine 15 or 20
or MDM2 binding. Mol Cell Biol. 23:2171–2181. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pang LY, Saunders L and Argyle DJ:
Epidermal growth factor receptor activity is elevated in glioma
cancer stem cells and is required to maintain chemotherapy and
radiation resistance. Oncotarget. 8:72494–72512. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Loughery J, Cox M, Smith LM and Meek DW:
Critical role for p53-serine 15 phosphorylation in stimulating
transactivation at p53-responsive promoters. Nucleic Acids Res.
42:7666–7680. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hokputsa S, Harding SE, Inngjerdingen K,
Jumel K, Michaelsen TE, Heinze T, Koschella A and Paulsen BS:
Bioactive polysaccharides from the stems of the Thai medicinal
plant Acanthus ebracteatus: Their chemical and physical
features. Carbohydr Res. 339:753–762. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Westerhoff HV, Zasloff M, Rosner JL,
Hendler RW, De Waal A, Vaz Gomes A, Jongsma PM, Riethorst A and
Juretić D: Functional synergism of the magainins PGLa and
magainin-2 in Escherichia coli, tumor cells and liposomes. Eur J
Biochem. 228:257–264. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hu C, Chen X, Huang Y and Chen Y:
Synergistic effect of the pro-apoptosis peptide kla-TAT and the
cationic anticancer peptide HPRP-A1. Apoptosis. 23:132–142. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Casimiro MC, Crosariol M, Loro E, Li Z and
Pestell RG: Cyclins and cell cycle control in cancer and disease.
Genes Cancer. 3:649–657. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Prives C and Hall PA: The p53 pathway. J
Pathol. 187:112–126. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sigal A and Rotter V: Oncogenic mutations
of the p53 tumor suppressor: The demons of the guardian of the
genome. Cancer Res. 60:6788–6793. 2000.PubMed/NCBI
|
|
37
|
Tang X, Zhu Y, Han L, Kim AL, Kopelovich
L, Bickers DR and Athar M: CP-31398 restores mutant p53 tumor
suppressor function and inhibits UVB-induced skin carcinogenesis in
mice. J Clin Invest. 117:3753–3764. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li D, Marchenko ND, Schulz R, Fischer V,
Velasco-Hernandez T, Talos F and Moll UM: Functional inactivation
of endogenous MDM2 and CHIP by HSP90 causes aberrant stabilization
of mutant p53 in human cancer cells. Mol Cancer Res. 9:577–588.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Brantley DM, Chen CL, Muraoka RS, Bushdid
PB, Bradberry JL, Kittrell F, Medina D, Matrisian LM, Kerr LD and
Yull FE: Nuclear factor-kappaB (NF-kappaB) regulates proliferation
and branching in mouse mammary epithelium. Mol Biol Cell.
12:1445–1455. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hew CS, Khoo BY and Gam LH: The
anti-cancer property of proteins extracted from Gynura
procumbens (Lour.) Merr. PLoS One. 8:e685242013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Prateep A, Sumkhemthong S, Suksomtip M,
Chanvorachote P and Chaotham C: Peptides extracted from edible
mushroom: Lentinus squarrosulus induces apoptosis in human lung
cancer cells. Pharm Biol. 55:1792–1799. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Issara-Amphorn J and T-Thienprasert NP:
Preliminary in vitro pro-apoptotic effects of Cratoxylum formosum
crude leaf extracts. Int J Appl Res Nat Prod. 7:26–30. 2014.
|
|
43
|
Budchart P, Khamwut A, Sinthuvanich C,
Ratanapo S, Poovorawan Y and T-Thienprasert NP: Partially purified
gloriosa superba peptides inhibit colon cancer cell viability by
inducing apoptosis through p53 upregulation. Am J Med Sci.
354:423–429. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Beg AA, Sha WC, Bronson RT, Ghosh S and
Baltimore D: Embryonic lethality and liver degeneration in mice
lacking the RelA component of NF-kappa B. Nature. 376:167–170.
1995. View Article : Google Scholar : PubMed/NCBI
|