Introduction
Colorectal cancer (CRC) is one of the most common
types of cancer in men and women, and the third leading cause of
cancer-associated mortality in the United States (1). Age is the most significant risk factor
(2), and as more individuals shift
to a western diet and lifestyle, the incidence of CRC increases
(3). Up to 5% of all CRC cases are
caused by hereditary syndromes, which include polyposis and
nonpolyposis CRCs (4,5). Endoscopic polypectomy is an effective
method to reduce the incidence and mortality of early-stage CRC
(6), but the majority of patients
are diagnosed at an advanced stage. Out of all patients with CRC,
35–55% develop hematogenous liver metastasis (7). The 5-year survival rate is ~90% for
patients with early-stage CRC, but <10% in patients with distant
metastases (8). Therefore, it is
critical to seek effective anticancer drugs for the treatment of
CRC; however, the exact mechanisms of anticancer drugs in CRC are
largely unknown.
Ginseng (the root of Panax ginseng C.A.
Meyer) is a valued traditional medicinal herb found in China and
Korea, which is widely used for its health benefits, particularly
in these countries (9). Ginsenoside
Rh1 (Rh1), found in red ginseng, is a metabolite of the major
ginsenosides Re and Rg1, formed by intestinal microbiota following
the oral ingestion of ginseng (10,11). Rh1
has been reported to possess anti-allergic, anti-inflammatory,
anti-aging, antioxidant and antitumor activity (12–14). Rh1
may also increase learning and memory capacity, as well as
hippocampal excitability (15,16).
Additionally, other pharmacological effects have been noted,
including myocardial injury (17)
and obesity prevention (18), as
well as antiplatelet effects (19).
Additionally, Rh1 induces anticancer effects in several tumor
cells, including human hepatocellular carcinoma (20), astroglioma (21) and acute monocytic leukemia cells
(22).
Although several studies have investigated the
anticancer effects of Rh1, to the best of our knowledge, there have
been no reports regarding the effects of Rh1 in CRC to date. The
underlying mechanisms of Rh1 on CRC migration and invasion remain
unknown. Therefore, in the present study, the regulation of CRC
invasion and migration by Rh1 was investigated in vitro and
in vivo.
Materials and methods
Cell culture
The human CRC cell line SW620 was purchased from the
Type Culture Collection of the Chinese Academy of Sciences. Cells
were cultured in RPMI-1640 medium (Gibco; Thermo Fisher Scientific,
Inc.) supplemented with 10% heat-inactivated fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc.), 1% streptomycin and
1% penicillin (all from Invitrogen; Thermo Fisher Scientific,
Inc.). The cultured cells were incubated at 37°C with 5%
CO2.
Analysis of cell viability and cell
proliferation
Ginsenoside Rh1 (purity >97%) was purchased from
Chengdu Must Bio-Technology Co., Ltd. (cat. no. A0240). The effects
of Rh1 on SW620 cell viability and proliferation were determined by
Cell Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.)
assays. Briefly, SW620 cells in the logarithmic growth phase were
seeded into 96-well plates at a density of 2×103
cells/well. Next, for examining cell viability, cells were treated
with Rh1 at concentrations of 0, 50 or 100 µM at 37°C for 24 or 48
h, and cells were treated with 0, 50 or 100 µM Rh1 at 37°C for 0,
12, 24 and 48 h to examine cell proliferation. Untreated cell
served as the control group. Then, 10 µl CCK-8 reagent was added to
each well and cells were incubated at 37°C with 5% CO2
for another 2 h. The absorbance of each well was measured at 450 nm
using a microplate reader (Thermo Fisher Scientific, Inc.).
Cell migration assay
Cell migration was determined by a wound healing
test. Cells (5×105 cells/well) were seeded into 6-well
plates and incubated at 37°C until >90% confluence was reached.
Cell monolayers were carefully scratched using a 10-µl sterile
plastic pipette tip to make an artificial wound. The floating cell
debris was rinsed twice with PBS. Subsequently, fresh serum-free
RPMI-1640 medium containing different concentrations of Rh1 (0 or
100 µM) was added. Images of the wound area were captured at 0 and
24 h under an inverted light microscope (magnification, ×200;
Olympus Corporation).
Matrigel invasion assay
Transwell chambers (pore size, 8-µm; Corning Inc.)
were used to examine the cell invasion capacity. The chamber filter
was pre-coated with 100 µl diluted Matrigel (BD Biosciences). Cell
suspensions (200 µl; 2×105 cells in serum-free RPMI-1640
medium containing 0 or 100 µM Rh1) were added to the upper chamber.
The bottom chamber was filled with complete RPMI-1640 medium
containing 10% FBS. Following incubation at 37°C with 5%
CO2 for 24 h, the remaining cells on the top surface of
the filter were removed. The invasive cells on the lower surface of
the filter were fixed in 4% paraformaldehyde for 10 min and stained
in 0.1% crystal violet for 15 min both at room temperature. The
invasive cells were imaged and counted using a light microscope
(Olympus Corporation) at ×200 magnification in five random
fields.
Reverse transcription-quantitative PCR
(RT-qPCR)
SW620 cells (2×105) in the logarithmic
growth phase were seeded in 6-well plates until 90% confluence was
reached. Subsequently, they were treated with 0 or 100 µM Rh1 at
37°C for 24 h. Total RNA from cells was extracted using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). cDNA synthesis was performed using a PrimeScript™ RT reagent
kit with gDNA Eraser (Takara Biotechnology Co., Ltd.). RT reaction
conditions were as follows: 37°C for 15 min followed by 85°C for 5
sec. qPCR was performed using SYBR Premix Ex Taq II (Tli RNaseH
Plus; Takara Biotechnology Co., Ltd.), according to manufacturer's
protocol, on an ABI PRISM 7500 Real-Time PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The PCR conditions
were as follows: Initial denaturation at 95°C for 30 sec, followed
by 40 cycles of denaturation at 95°C for 5 sec and
annealing/extension at 60°C for 34 sec. The melting curve was
analyzed at the end of amplification. The fold change of each gene
was calculated using the 2−ΔΔCq method (23) and normalization to GAPDH. The primers
were synthesized by GenScript, and the sequences used were: Matrix
metallopeptidase (MMP)1 forward, 5′-AGGACTCCAAGGTAGACACAC-3′ and
reverse, 5′-TTGCCGTTCTTGTAGGTGAACGC-3′; MMP3 forward,
5′-CCTGCTTTGTCCTTTGATGC-3′, and reverse,
5′-TGAGTCAATCCCTGGAAAGTC-3′; tissue inhibitor of metalloproteinases
3 (TIMP3) forward, 5′-AGTTACCCAGCCCTATGA-3′ and reverse,
5′-GCAAAGGCTTAAACATCT-3′; and GAPDH forward,
5′-GAAGGTGAAGGTCGGAGTC-3′ and reverse,
5′-GAAGATGGTGATGGGATTTC-3′.
Western blot analysis
SW620 cells (2×105) treated with 0 or 100
µM Rh1 for 24 h as aforementioned were harvested and lysed in RIPA
buffer (Beyotime Institute of Biotechnology). Protein concentration
was determined by the bicinchoninic acid (BCA) method using a BCA
Protein Assay kit (Beijing Solarbio Science & Technology Co.,
Ltd.) according to the manufacturer's protocol. A total of 30 µg of
protein was separated via 10% SDS-PAGE. Following electrophoresis,
proteins were transferred onto PVDF membranes (Thermo Fisher
Scientific, Inc.). Membranes were blocked in TBS with 0.05% Tween
20 (Beijing Solarbio Science & Technology Co., Ltd.) containing
5% skimmed milk at room temperature for 1 h, and subsequently
incubated with primary antibodies at 4°C overnight. The primary
antibodies (all from Abcam) included: Anti-MMP1 (cat. no. ab38929;
1:5,000), anti-MMP3 (cat. no. ab53015; 1:1,000), anti-ERK1+ERK2
(cat. no. ab17942; 1:1,000), anti-phosphorylated (p-)ERK1 + ERK2
(p-T202 + T204; cat. no. ab214362; 1:1,000), anti-P38 (cat. no.
ab197348; 1:1,000), anti-p-P38 (p-T180; cat. no. ab178867; 1:1,000
dilution), anti-JNK1+JNK2 (cat. no. ab112501; 1:1,000), anti-p-JNK1
+ JNK2 (p-T183 + T185; cat. no. ab4821; 1:1,000), anti-TIMP3 (cat.
no. ab39184; 1:1,000) and anti-GAPDH (cat. no. ab181602; 1:10,000).
Next, the membranes were incubated with secondary antibody
conjugated to horseradish peroxidase (goat anti-rabbit
immunoglobulin G; 1:2,000 dilution; cat. no. ab205718; Abcam) for 1
h at room temperature. The immunopositive bands were visualized
using a Pierce ECL Western Blotting kit (Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocols. Band intensity was
determined with the ImageQuant™ LAS 4010 biomolecular
imager (GE Healthcare Life Sciences) and normalized to GAPDH.
Nude mouse xenograft model and Rh1
treatment
A total of 6 adult male BALB/c nude mice (4–6 weeks;
20–22 g) were selected for the present study. Nude mice were
purchased from Shanghai Experimental Animal Center of the Chinese
Academy of Sciences (Shanghai, China). The mice were housed
individually in a specific pathogen-free environment with free
access to water and food under 26–28°C, 40–60% humidity and 10 h
light/14 h dark conditions. The experimental procedures were
approved by the Institutional Animal Care and Use Committee of
Lanzhou University Second Hospital (Lanzhou, China). Logarithmic
growth phase SW620 cells (3×106) were suspended in 250
µl PBS and Matrigel (1:1 ratio; BD Biosciences) and injected
subcutaneously into the dorsal right flank of mice. Mice were
randomly divided into control and Rh1 groups (3 mice/group). Mice
in the control group were orally administered with 2 ml distilled
water/day, while mice in Rh1 group were orally administrated with
20 mg/kg/day Rh1, dissolved in 2 ml distilled water, for 35
consecutive days. Tumor volumes were measured once per week. At day
35, mice were sacrificed by decapitation and the tumors were
dissected at 12 h after the last drug delivery. The tumor length
and width were measured with a caliper, and the volume was
calculated using the formula: Volume=0.5 × (length ×
width)2. Images of the tumors in each group were
captured and the tumors were weighed.
Statistical analysis
Statistical analysis was performed using GraphPad
Prism version 7 software (GraphPad Software, Inc.). Data are
expressed as the means ± SEM of at least three independent
experiments. The differences between two groups were analyzed using
a Student's t-test, and the differences among multiple groups were
assessed using one-way ANOVA followed by a Newman Keuls post hoc
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Effects of Rh1 on the viability and
proliferation of CRC cells
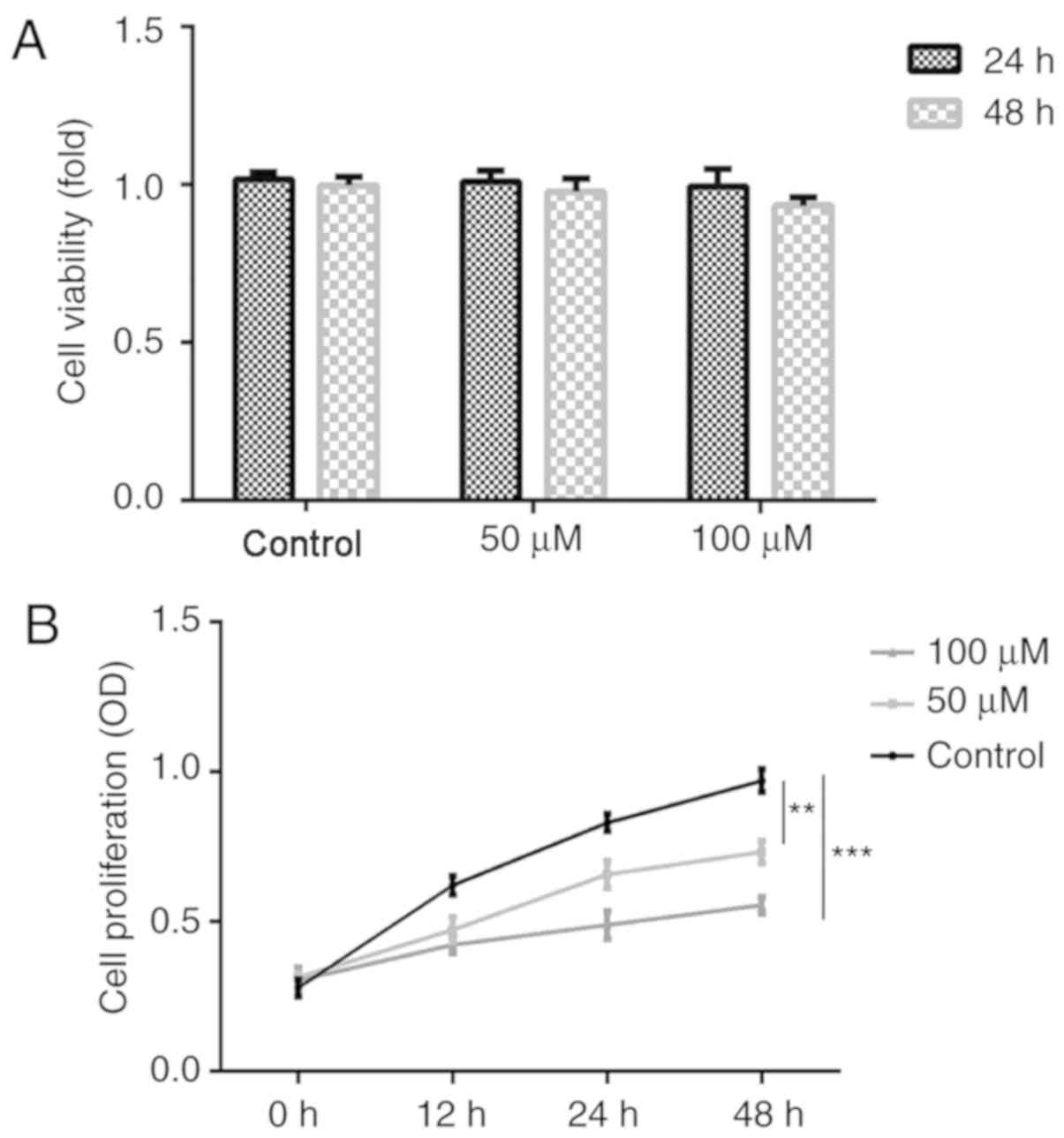
To investigate the effects of Rh1 on cell viability
and proliferation, SW620 cells were treated with various
concentrations (0, 50 or 100 µM) of Rh1, and CCK-8 assays were
performed. As shown in Fig. 1A, 50
and 100 µM Rh1 did not influence cell viability in the treatment
groups, compared with in the control group. Furthermore, Rh1
treatment for 24 or 48 h did not significantly influence cell
viability for the same Rh1 concentrations. These results indicated
that Rh1 did not exert significant cytotoxicity on SW620 cells at
0–100 µM for 24 or 48 h. However, 50 and 100 µM Rh1 significantly
inhibited cell proliferation (50 µM, P<0.01; 100 µM, P<0.001)
in the treatment group compared with in the control group (Fig. 1B). The nontoxic concentration of 100
µM was selected for subsequent experimentation.
Rh1 inhibits the migration and
invasion of CRC cells in vitro
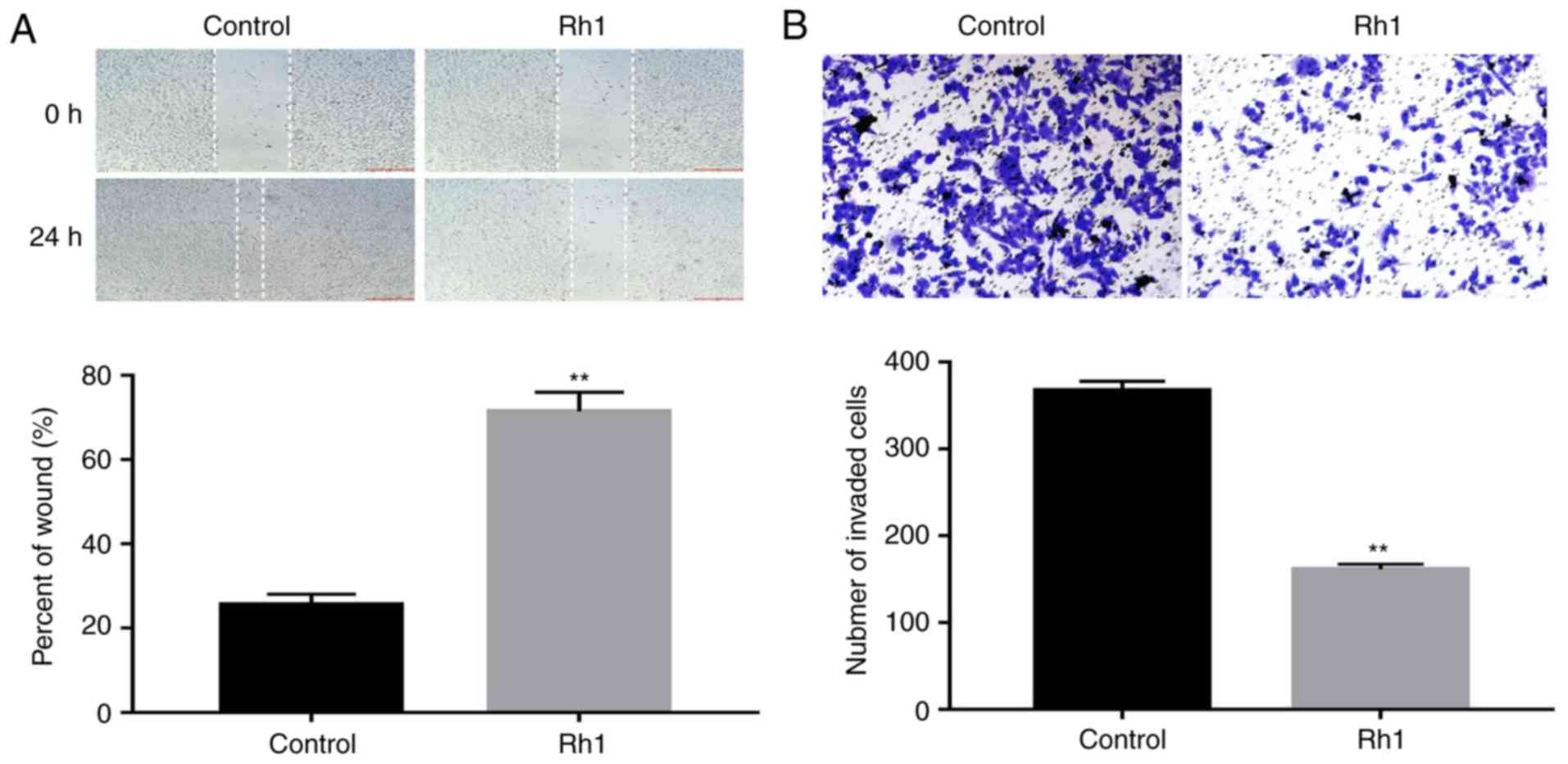
To evaluate the effects of Rh1 on cell migration and
invasion, SW620 cells were treated with 0 or 100 µM Rh1 for 24 h.
The wound healing assay demonstrated that Rh1 significantly
inhibited wound closure. The percentage of the wound area in the
Rh1 group was significantly greater than that of the control group
(P<0.01; Fig. 2A). Similarly, the
Transwell invasion assay demonstrated that Rh1 treatment
significantly decreased the number of invasive cells, compared with
the control group (P<0.01; Fig.
2B). These data suggested that Rh1 effectively inhibited the
migration and invasion of CRC cells in vitro.
Rh1 suppresses MMP1 and MMP3
expression, increases the expression levels of TIMP3 and inhibits
mitogen-activated protein kinase (MAPK) signaling pathway
activation
The expression levels of MMP1, MMP3 and TIMP3 were
measured in CRC cells via RT-qPCR and western blot analyses,
respectively. The results revealed that Rh1 treatment markedly
decreased the mRNA expression levels of MMP1 and MMP3, and
increased TIMP3 mRNA expression, compared with the control group
(P<0.01; Fig. 3A-C).
Consistently, the protein expression levels of MMP1 and MMP3 were
significantly decreased, and TIMP3 expression was increased in the
Rh1 group (P<0.01; Fig. 3D-G).
Additionally, as presented in Fig. 3H
and I, it was demonstrated that Rh1 significantly decreased the
ratios of p-P38/P38, p-ERK1/2/ERK1-2 and p-JNK/JNK compared with
the control group (P<0.01). These results suggested that Rh1
inhibited MAPK signaling.
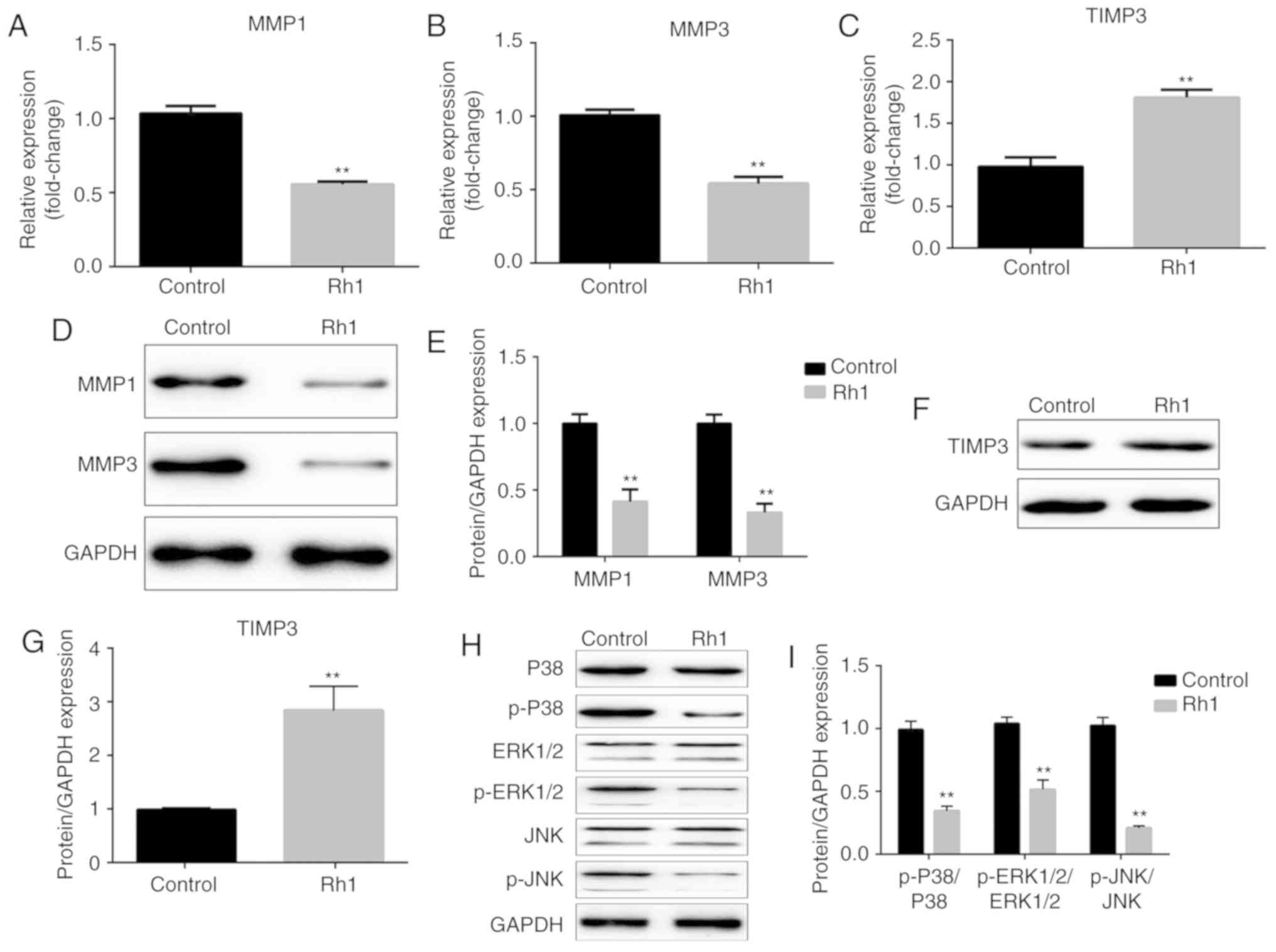 | Figure 3.Rh1 decreases the mRNA and protein
expression levels of MMP1 and MMP3, enhances TIMP3 expression and
inhibits the phosphorylation of ERK1/2, P38 and JNK. mRNA
expression levels of (A) MMP1, (B) MMP3 and (C) TIMP3 were detected
using reverse transcription-quantitative PCR following exposure of
SW620 cells to 0 or 100 µM Rh1 for 24 h. (D) Protein expression
levels of MMP1 and MMP3 were measured using western blotting. GAPDH
was used as an internal control. (E) Fold change of MMP1 and MMP3
protein levels was normalized using GAPDH. (F) Protein expression
levels of TIMP3 were detected by western blot analysis. GAPDH was
used for normalization. (G) Semi-quantified TIMP3 protein
expression. (H) Protein expression levels of P38, p-P38, ERK1/2,
p-ERK1/2, JNK and p-JNK were assessed using western blotting. GAPDH
was used as an internal control. (I) Relative protein expression
levels of p-P38, p-ERK1/2 and p-JNK were compared with their total
amount. Data are presented as the means ± SEM. **P<0.01 vs.
control group. MMP, matrix metallopeptidase; p-, phosphorylated-;
Rh1, ginsenoside Rh1; TIMP3, tissue inhibitor of metalloproteinases
3. |
Inhibitory effects of Rh1 on xenograft
tumor growth and MAPK signaling in vivo
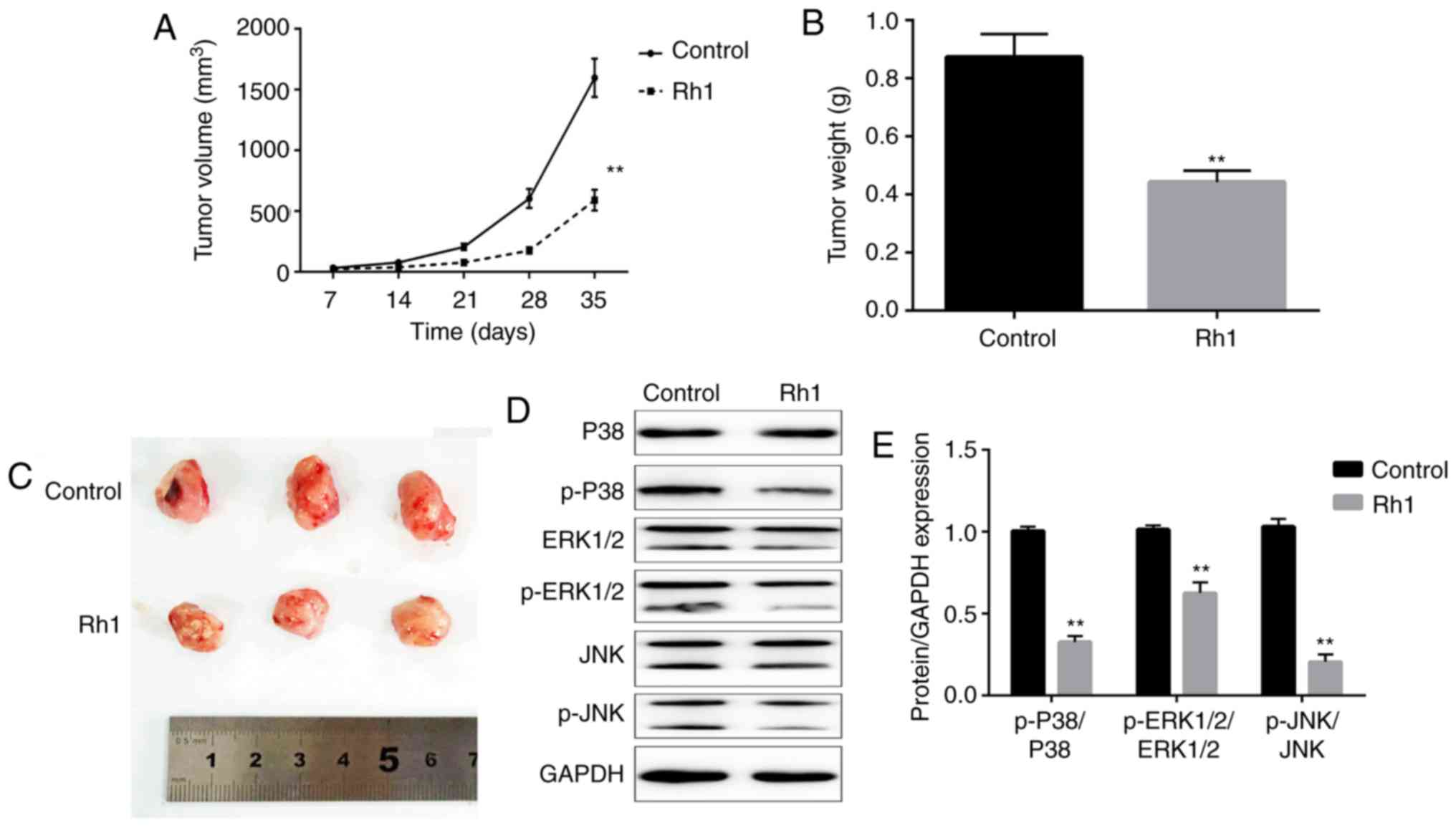
To further investigate the antitumor effects of Rh1,
a xenograft model of nude mice was established with SW620 cells.
Rh1 (0 or 20 mg/kg) was orally administered to treat the tumors in
mice, and tumor growth was determined. The tumor volume was
significantly decreased in the Rh1 group compared with in the
control group after 35 days (P<0.01; Fig. 4A). When the tumors were harvested,
the tumor weights were measured. Rh1 treatment significantly
decreased the tumor weight, compared with the control group
(P<0.01; Fig. 4B). Additionally,
tumor size was markedly decreased in the Rh1 group compared with
that in the control group (Fig. 4C).
These results suggested that Rh1 inhibited tumor growth in
vivo. Additionally, whether Rh1 regulates the MAPK signaling
pathway in vivo remains unclear. The ratios of p-P38/P38,
p-ERK1/2/ERK1-2 and p-JNK/JNK were significantly downregulated in
Rh1 treated mice tumor tissues compared with in tissues from the
control group (P<0.01; Fig. 4D and
E).
Discussion
The present study revealed that different
concentrations of Rh1 exhibited no toxicity in CRC cells. Rh1 (100
µM) significantly inhibited the proliferation, migration and
invasion of CRC cells in vitro, and 20 mg/kg Rh1 inhibited
tumor growth in vivo.
Rh1 is a hydrolysis product that reaches the
systemic circulation following ginseng ingestion (24), and has been reported to exhibit
certain biological activities, including anti-allergic,
anti-inflammatory and antitumor activities (12–14).
Previous studies have revealed that Rh1 is involved in cell
viability and proliferation. For example, Rh1 inhibits the
proliferation of non-alcoholic fatty liver cells (25). Conversely, Wang et al
(26) revealed that Rh1 has no
effect on human peripheral blood mononuclear cell proliferation up
to 100 mg/l. In addition, the cytotoxicity in neuronal cells is not
affected by different concentrations of Rh1 (27). Rh1 at the concentrations of 0–100 µM
was not toxic to CRC cells in the present study. However, cell
proliferation was significantly suppressed by Rh1 in a
dose-dependent manner. These findings suggested that Rh1 suppressed
tumor progression partially due to the inhibition of cell
proliferation rather than due to toxic effects. Therefore, a
nontoxic concentration was selected for a more accurate assessment
of CRC cell migration and invasion.
Migration and invasion are major obstacles in the
treatment of cancer, which induce poor prognosis and affect patient
survival (28). Previous studies
have revealed that Rh1 inhibits human cancer migration and
invasion: For example, Rh1 exerts inhibitory effects on migration
and invasion by suppressing MMP1, MMP3 and MMP9 expression in human
astroglioma (21). Additionally, Rh1
inhibits THP-1 acute monocytic leukemia cell migration and invasion
(22). Similarly, the results of the
present study demonstrated that Rh1 inhibited CRC cell migration
and invasion. However, the molecular mechanisms underlying the
inhibition of migration and invasion remain unclear.
MMPs, including MMP1 and MMP3, serve an important
role in tumor progression, and are involved in tumor metastasis
processes, including angiogenesis, migration and invasion (29). High expression levels of MMP1 are
associated with lymph node metastasis, and promote tumor growth and
metastasis in esophageal squamous cell carcinoma (30). In CRC, MMP1 is a prognostic indicator
for hematogenous metastasis (31).
MMP3 is a tumor promoter involved in tumorigenesis and metastasis,
which promotes tumor cell migration and invasion (32). TIMPs inhibit activated MMPs, keeping
the balance of TIMPs and MMPs (33).
TIMP3 has been reported to be associated with anticancer
capability, and regulates cell migration and invasion in
osteosarcoma (34), cervical cancer
(35) and esophageal squamous cell
carcinoma (36). In CRC, TIMP3 is
downregulated in tumor tissues and inhibits cell migration and
invasion (37). The present study
measured the expression levels of MMP1 and MMP3, which were
decreased following Rh1 treatment. Additionally, Rh1 treatment led
to an increase in the expression levels of TIMP3. Thus, it was
inferred that Rh1 suppressed CRC cell migration and invasion, at
least partially, via suppressing MMP1 and MMP3 expression and
promoting TIMP3 expression.
The MAPK signaling pathway consists of four
independent cascades. The terminal kinases of these cascades
include ERK, Big MAPK-1, JNK and P38 (38). The MAPK signaling pathways link
extracellular signals to the machinery that controls basic cellular
processes, including proliferation, differentiation, invasion,
migration and apoptosis (39,40). A
previous study revealed that Rh1 suppresses MMP1 expression via
inhibition of MAPK signaling (20).
In addition, Rh1 inhibited the migration and invasion of acute
monocytic leukemia cells by inactivating the MAPK signaling pathway
(22). The present study revealed
that Rh1 inhibited the phosphorylation of ERK1/2, p38 and JNK in
SW620 cells and tumor-bearing mice tissues, which suggested that
Rh1 inhibited the activation of the MAPK signaling pathway not only
in vitro but also in vivo. From this, it was
concluded that Rh1 may have suppressed CRC cell migration and
invasion via MAPK signaling pathway inactivation.
Furthermore, a xenograft model was established to
investigate the effects of Rh1 on tumor growth in mice. It was
identified that Rh1 reduced tumor volume and weight, indicating
that Rh1 inhibited tumor growth in vivo. To the best of our
knowledge, this was the first study demonstrating that Rh1 exerted
anticancer activity in CRC in vivo.
In conclusion, the present study demonstrated that
Rh1 significantly reduced CRC cell proliferation, migration and
invasion. Rh1 exerted anticancer activity, potentially through
inhibiting MMP1 and MMP3 expression, increasing TIMP3 expression,
and MAPK signaling pathway inactivation. Furthermore, Rh1 inhibited
tumor growth and the MAPK signaling pathway in vivo. These
findings indicated that Rh1 has potential for development into a
novel drug for adjuvant therapy of CRC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
XL and YCZ designed the study. XL, XX, AS, JG and
YWZ performed the experiments and analyzed data. XL mainly wrote
the manuscript and YCZ revised it. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
The experimental procedures were approved by the
Institutional Animal Care and Use Committee of Lanzhou University
Second Hospital (Lanzhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Binefa G, Rodríguez-Moranta F, Teule A and
Medina-Hayas M: Colorectal cancer: From prevention to personalized
medicine. World J Gastroenterol. 20:6786–6808. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Brody H: Colorectal cancer. Nature. 521
(Suppl):S12015. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Balaguer F: Hereditary and familial
colorectal cancer. Gastroenterol Hepatol Suppl. 3:77–84. 2014.
View Article : Google Scholar
|
|
5
|
Vasen HF, Tomlinson I and Castells A:
Clinical management of hereditary colorectal cancer syndromes. Nat
Rev Gastroenterol Hepatol. 12:88–97. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pellisé M: Colonoscopy in the screening,
follow-up and treatment of colorectal cancer and precursor lesions.
Gastroenterol Hepatol. 38 (Suppl 1):S71–S77. 2015.(In Spanish).
View Article : Google Scholar
|
|
7
|
Akgül Ö, Çetinkaya E, Ersöz Ş and Tez M:
Role of surgery in colorectal cancer liver metastases. World J
Gastroenterol. 20:6113–6122. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ting WC, Chen LM, Pao JB, Yang YP, You BJ,
Chang TY, Lan YH, Lee HZ and Bao BY: Common genetic variants in Wnt
signaling pathway genes as potential prognostic biomarkers for
colorectal cancer. PLoS One. 8:e561962013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xiang YZ, Shang HC, Gao XM and Zhang BL: A
comparison of the ancient use of ginseng in traditional Chinese
medicine with modernpharmacological experiments and clinical
trials. Phytother Res. 22:851–858. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee SM, Bae BS, Park HW, Ahn NG, Cho BG,
Cho YL and Kwak YS: Characterization of Korean red ginseng (Panax
ginseng Meyer): History, preparation method and chemical
composition. J Ginseng Res. 39:384–391. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ko SR, Suzuki Y, Choi KJ and Kim YH:
Enzymatic preparation of genuine prosapogenin, 20(S)-ginsenoside
Rh1, from ginsenosides Re and Rg1. Biosci Biotechnol Biochem.
64:2739–2743. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Park EK, Choo MK, Han MJ and Kim DH:
Ginsenoside Rh1 possesses antiallergic and anti-inflammatory
activities. Int Arch Allergy Immunol. 133:113–120. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jiang Y, Liu W, Wang XM, Zhong GG, Zhang
WJ, Chen L, Zhan S, Qi H, Zhao CY, Ma XY, et al: Calcium channel
blockade and anti-free-radical actions of panaxatriol saponins in
cultured myocardiocytes. Zhongguo Yao Li Xue Bao. 17:138–141.
1996.PubMed/NCBI
|
|
14
|
Cheng Y, Shen LH and Zhang JT:
Anti-amnestic and anti-aging effects of ginsenoside Rg1 and Rb1 and
its mechanism of action. Acta Pharmacol Sin. 26:143–149. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang YZ, Chen J, Chu SF, Wang YS, Wang XY,
Chen NH and Zhang JT: Improvement of memory in mice and increase of
hippocampal excitability in rats by ginsenoside Rg1's metabolites
ginsenoside Rh1 and protopanaxatriol. J Pharmacol Sci. 109:504–510.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hou J, Xue J, Lee M, Yu J and Sung C:
Long-term administration of ginsenoside Rh1 enhances learing and
memory by promoting cell survival in the mouse hippocampus. Int J
Mol Med. 33:234–240. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gai Y, Ma Z, Yu X, Qu S and Sui D: Effect
of ginsenoside Rh1 on myocardial injury and heart function in
isoproterenol-induced cardiotoxicity in rats. Toxicol Mech Methods.
22:584–591. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gu W, Kim KA and Kim DH: Ginsenoside Rh1
ameliorates high fat diet-induced obesity in mice by inhibiting
adipocyte differentiation. Biol Pharm Bull. 36:102–107. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Teng CM, Kuo SC, Ko FN, Lee JC, Lee LG,
Chen SC and Huang TF: Antiplatelet actions of panaxynol and
ginsenosides isolated from ginseng. Biochim Biophys Acta.
990:315–320. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yoon JH, Choi YJ and Lee SG: Ginsenoside
Rh1 suppresses matrix metalloproteinase-1 expression through
inhibition of activator protein-1 and mitogen-activated protein
kinase signaling pathway in human hepatocellular carcinoma cells.
Eur J Pharmacol. 679:24–33. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jung JS, Ahn JH, Le TK, Kim DH and Kim HS:
Protopanaxatriol ginsenoside Rh1 inhibits the expression of matrix
metalloproteinases and the in vitro invasion/migration of human
astroglioma cells. Neurochem Int. 63:80–86. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Choi YJ, Yoon JH, Cha SW and Lee SG:
Ginsenoside Rh1 inhibits the invasion and migration of THP-1 acute
monocytic leukemia cells via inactivation of the MAPK signaling
pathway. Fitoterapia. 82:911–919. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lai L, Hao H, Liu Y, Zheng C, Wang Q, Wang
G and Chen X: Characterization of pharmacokinetic profiles and
metabolic pathways of 20(S)-ginsenoside Rh1 in vivo and in vitro.
Planta Med. 75:797–802. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen XJ, Liu WJ, Wen ML, Liang H, Wu SM,
Zhu YZ, Zhao JY, Dong XQ, Li MG, Bian L, et al: Ameliorative
effects of Compound K and ginsenoside Rh1 on non-alcoholic fatty
liver disease in rats. Sci Rep. 7:411442017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang Y, Wang BX, Liu TH, Minami M, Nagata
T and Ikejima T: Metabolism of ginsenoside Rg1 by intestinal
bacteria. II. Immunological activity of ginsenoside Rg1 and Rh1.
Acta Pharmacol Sin. 21:792–796. 2000.PubMed/NCBI
|
|
27
|
Li XF, Lui CN, Jiang ZH and Ken YK:
Neuroprotective effects of ginsenosides Rh1 and Rg2 on neuronal
cells. Chin Med. 6:192011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kramer N, Walzl A, Unger C, Rosner M,
Krupitza G, Hengstschläger M and Dolznig H: In vitro cell migration
and invasion assays. Mutat Res. 752:10–24. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Deryugina EI and Quigley JP: Matrix
metalloproteinases and tumor metastasis. Cancer Metastasis Rev.
25:9–34. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu M, Hu Y, Zhang MF, Luo KJ, Xie XY, Wen
J, Fu JH and Yang H: MMP1 promotes tumor growth and metastasis in
esophageal squamous cell carcinoma. Cancer Lett. 377:97–104. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sunami E, Tsuno N, Osada T, Saito S,
Kitayama J, Tomozawa S, Tsuruo T, Shibata Y, Muto T and Nagawa H:
MMP-1 is a prognostic marker for hematogenous metastasis of
colorectal cancer. Oncologist. 5:108–114. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chu C, Liu X, Bai X, Zhao T, Wang M, Xu R,
Li M, Hu Y, Li W, Yang L, et al: MiR-519d suppresses breast cancer
tumorigenesis and metastasis via targeting MMP3. Int J Biol Sci.
14:228–236. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jackson HW, Defamie V, Waterhouse P and
Khokha R: TIMPs: Versatile extracellular regulators in cancer. Nat
Rev Cancer. 17:38–53. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Han XG, Li Y, Mo HM, Li K, Lin D, Zhao CQ,
Zhao J and Tang TT: TIMP3 regulates osteosarcoma cell migration,
invasion and chemotherapeutic resistances. Tumour Biol.
37:8857–8867. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang Z, Wang J, Wang X, Song W, Shi Y and
Zhang L: MicroRNA-21 promotes proliferation, migration and invasion
of cervical cancer through targeting TIMP3. Arch Gynecol Obstet.
297:433–442. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu W, Li M, Chen X, Zhang D, Wei L, Zhang
Z, Wang S, Meng L, Zhu S and Li B: MicroRNA-373 promotes migration
and invasion in human esophageal squamous cell carcinoma by
inhibiting TIMP3 expression. Am J Cancer Res. 6:1–14.
2015.PubMed/NCBI
|
|
37
|
Lin H, Zhang Y, Wang H, Xu D, Meng X, Shao
Y, Lin C, Ye Y, Qian H and Wang S: Tissue inhibitor of
metalloproteinases-3 transfer suppresses malignant behaviors of
colorectal cancer cells. Cancer Gene Ther. 19:845–851. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cossa G, Gatti L, Cassinelli G, Lanzi C,
Zaffaroni N and Perego P: Modulation of sensitivity to antitumor
agents by targeting the MAPK survival pathway. Curr Pharm Des.
19:883–894. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Burotto M, Chiou VL, Lee JM and Kohn EC:
The MAPK pathway across different malignancies: A new perspective.
Cancer. 120:3446–3456. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yoon JH, Choi YJ, Cha SW and Lee SG:
Anti-metastatic effects of ginsenoside Rd via inactivation of MAPK
signaling and induction of focal adhesion formation. Phytomedicine.
19:284–292. 2012. View Article : Google Scholar : PubMed/NCBI
|