Vascular development begins with the differentiation
of mesodermal cells into endothelial cell precursors (angioblasts),
which form primary vessels de novo by a process known as
vasculogenesis (1). The primary
vessels are subsequently remodeled by the sprouting and branching
of new blood vessels, a process known as angiogenesis.
Physiologically, angiogenesis establishes the first vascular tree
and adequate vasculature for the growth and development of organs
in the embryo (2), whilst in adults,
angiogenesis occurs during the ovarian cycle and wound healing
(3). The process is controlled by
balancing inducers and inhibitors of angiogenesis (4). Cancer cells use angiogenesis to fulfil
the increased need for nutrients and oxygen to the growing tumor.
Angiogenesis also promotes tumor invasion and metastasis, and has
been described as one of the six hallmarks of cancer (5).
Angiogenesis begins with the activation of quiescent
endothelial cells (ECs) in response to angiogenic stimuli. A number
of proteins are important for angiogenesis. The VEGF family of
secreted proteins and their receptors, Fibroblast growth factors
and Notch signaling are some of the most well studied regulators of
angiogenesis. Platelet derived growth factor, the angiopoietins and
tie receptors are associated with vessel maturation (6). The extracellular matrix (ECM) serves a
pivotal role in the regulation of both physiological and
pathological angiogenesis (7).
Endothelial cell-cell adhesion and adhesion with the ECM are
essential to establish the appropriate cellular configuration for
growth, survival and differentiation. Cell adhesion molecules,
including integrins, cadherin, immunoglobulin families and selectin
are critically involved in angiogenesis (8). In absence of appropriate cell contact,
the ECs may undergo programmed cell death or unable to form new
capillary blood vessels and extension and maturation of new vessels
(8,9). Activated ECs secrete proteinases to
breakdown the surrounding basement matrix and invade the ECM
(1,10). Once free from the ECM, ECs
proliferate and migrate towards chemotactic and angiogenic stimuli.
The newly formed vessels are stabilized by basement membrane
synthesis and the recruitment of pericytes, and fresh sprouts fuse
to establish blood flow (11–13).
The C-type lectin family XIV members are expressed
on angiogenic blood vessels and are vital for cell-cell adhesion
and cell-ECM interactions during angiogenesis. The ECM also
releases proteiolytic enzymes that results in degradation of matrix
molecules and soluble factors that promote angiogenesis (7). The C-type lectin family XIV members are
associated with increased expression of ECM degrading enzymes like
MMPs and plasminogen activators. They have been associated with
increased rate of angiogenesis in a variety of cancers. These
proteins are also implicated in other diseases involving
endothelial dysfunction and have been used as a biomarker in these
diseases. High plasma levels of thrombomodulin has been observed in
preeclampsia, diffuse intravascular coagulation, Shiga
toxin-producing E. coli (STEC)-induced and atypical
hemolytic uremic syndrome, thrombotic thrombocytopenic purpura,
scleroderma-associated pulmonary hypertension, and arterial
hypertension (14–19). The role of endosialin in rheumatoid
arthritis and Salmonella infection is well established
(20,21). CD93 expression is found to be altered
in systemic lupus erythematosus, rheumatoid arthritis and coronary
artery disease (22–24). In this review study, however, we have
focused primarily on the role of these proteins in regulating
tumour angiogenesis.
CLEC14A is a transmembrane glycoprotein containing
an extracellular signal peptide, a CTLD, a sushi-like domain, a
single EGF-like domain, a mucin-like domain, a single transmembrane
domain and an intracellular cytoplasmic domain. Human
CLEC14a is an intronless gene located on chromosome 14q21.1,
and CLEC14A is expressed specifically in the embryonic vasculature
of mice and zebrafish, and by human ECs (31). Reverse transcription-quantitative PCR
analysis revealed that CLEC14A expression begins from 12 h
post-fertilization in zebrafish embryos, which coincides with the
generation of hemangioblasts. Also, CLEC14A expression level was
shown to increase in the later stages of angiogenic development
(32). CRISPR-Cas9-mediated knockout
of clec14a in zebrafish resulted in the malformation of
inter-segmental vessels (ISVs), and the knockout of both
cd93 and clec14a resulted in the inhibition of
cadherin 5 expression in ISVs (33).
Furthermore, the CLEC14A expression level was significantly higher
in the tumor vasculature compared with the normal vasculature
(32). CLEC14A is one of the primary
genes of the tumor angiogenesis signature, highly expressed in head
and neck squamous cell carcinoma, breast cancers and clear cell
renal cell carcinoma (34), and is
therefore considered to be a tumor endothelial marker (TEM). Its
expression was also shown to increase in response to hypoxia in
HUVECs (35).
CLEC14A has also been reported to regulate
pro-angiogenic phenotypes such as filopodia formation, cell
migration and tube formation in HUVECs (31,32); it
localizes to the inter-cellular boundary and regulates cell
adhesion through its CTLD (31).
Targeted neutralization of CLEC14A using an anti-CTLD antibody was
shown to inhibit endothelial cell migration, cell-cell contact and
tube formation by blocking CTLD-CTLD interactions, and
downregulating CLEC14A expression at the endothelial cell surface
(36,37). A Stable Isotope Labelling with Amino
Acids in Cell Culture-based proteomics study showed the
upregulation of CLEC14A expression during tubule morphogenesis, and
the analysis of post-translational modifications of CLEC14A
identified a phosphorylation site at Ser483, near the PDZ-binding
domain in the cytoplasmic tail (38). The PDZ domain serves a vital role in
protein-protein interactions (39),
although no functional studies have been reported for the
phosphorylation site and the PDZ domain of CLEC14A.
Secretory CLEC14A co-localizes with fibronectin
(FN1), laminin alpha 4 (LAMA4) and multimerin 2 (MMRN2) in the ECM,
and its expression is upregulated during tumor angiogenesis in
spontaneous mouse tumors (38). FN1
and LAMA4 were also reportedly deregulated during tumor
angiogenesis (40,41), and MMRN2 was shown to suppress
vascular endothelial growth factor A (VEGFA)/Vascular endothelial
growth factor receptor 2 (VEGFR2) signaling by sequestering VEGFA
(42). However, the pro-angiogenic
properties of MMRN2 have also been reported (38,43,44).
Furthermore, antibody-mediated disruption of the interaction
between CLEC14A and MMRN2 inhibited sprouting angiogenesis and
tumor growth (45). As an ECM
protein, MMRN2 binds to the CTLD of CLEC14A; CLEC14A-CTLD is also
reported to interact with Hsp70-1A and to be crucial for
Hsp70-1A-induced angiogenesis via
extracellular-signal-regulated kinase (ERK) activation (46). Moreover, CLEC14A is cleaved by
rhomboid-like protein 2 (a membrane-embedded proteolytic enzyme),
and the cleaved extracellular domain has sprout-inhibiting and
anti-migratory properties. This inhibition is thought to have been
due to competition between the cleaved extracellular domain and
wild type forms of CLEC14A for the binding of MMRN2 (47).
In contrast to the above observations, increased
angiogenesis and lymphangiogenesis were observed in
CLEC14A-knockout mice, and CLEC14A-deficient mice exhibited
abnormal tumor vasculature and reduced survival of tumor-bearing
mice. Additionally, the loss of CLEC14A was attributed to the loss
of VEGFR3 expression and suppressed Notch/Dll4 and Notch
target gene expression (48).
Furthermore, the deletion of EC-specific VEGFR3 was shown to
induce hypervascularity, and VEGFR-3 knockdown using siRNA,
followed by VEGF treatment, increased the level of VEGFR-2
phosphorylation in HUVECs (49). The
role of CLEC14A may be context-dependent, and future studies to
identify CLEC14A-interacting partners (of both to the extracellular
domain and the cytoplasmic domain) and their signaling pathways may
enhance understanding of their precise roles in angiogenesis.
Thrombomodulin (also known as CD141) is a
membrane-bound glycoprotein with an N-terminal signal peptide, a
CTLD, six tandem EGF-like domain repeats, an O-glycosylation
site-rich domain, a transmembrane domain and a short cytoplasmic
C-terminal loop (50,51). Thrombomodulin is an intronless gene
(location, chromosome 20p11.21) initially identified as a gene
expressed in the vascular endothelium (52). It was later discovered to also be
expressed in smooth muscle cell lines (53), and by both circulating and tissue
mononuclear phagocytes (54).
Additionally, treatment with VEGF also increased the expression
level of thrombomodulin in human aortic ECs (55). The CTLD of thrombomodulin was
reported to mediate Ca2+-dependent cell-cell adhesion,
and antibody-targeted inhibition of the CTLD prevented cell-cell
contact, whereas an antibody towards the EGF domain of
thrombomodulin did not (56). In
addition, thrombomodulin was shown to co-localize with actin
filaments, and cell-cell adhesion was abolished by mannose,
chondroitin sulfate A and chondroitin sulfate C administration
(56).
It was postulated that the lectin-like domain of
thrombomodulin (expressed at the tumor surface) interacted with
cell membrane and ECM proteins, and also facilitated cell-cell
adhesion (57,58). The CTLD of thrombomodulin also
interacts with the ECM protein fibronectin in the tumor
vasculature, enhancing cell adhesion and migration. It has also
been reported to promote the phosphorylation of focal adhesion
kinase 1 (FAK) and the expression of matrix metalloproteinase 9
(MMP9) (59). The role of FAK in
angiogenesis has also been reported (60,61).
Knockdown of thrombomodulin in HaCaT cells inhibited
E-cadherin trafficking to the cell membrane, bestowing a more
fibroblast-like phenotype (62). In
another study, the recombinant lectin-like domain of TM-TM domain 1
(rTMD1) was found to inhibit HUVEC tube formation by Matrigel
assays, as well as disrupting the interaction between rTMD1 and
Lewis Y Ag (LeY)-modified EGFR, resulting in the inhibition of
EGF-mediated EGFR signaling and angiogenesis. This was believed to
be due to rTMD1-associated interference of LeY (a cell surface
tetra-saccharide) in endothelial cell connection and capillary
formation (63).
A soluble form of thrombomodulin containing the CTLD
has been shown to retard tumor cell invasiveness, whereas soluble
thrombomodulin lacking this domain was unable to inhibit cell
invasion, suggesting an anti-metastatic role for the CTLD of
thrombomodulin (64). The CTLD of
thrombomodulin was also shown to suppress
lipopolysaccharide-induced ERK1/2 phosphorylation (65). Additionally, thrombomodulin
expression was inversely correlated with tumor cell proliferation
in lung squamous cell carcinoma (66), esophageal squamous cell carcinoma
(67), hepatocellular carcinoma
(57), colorectal cancer (68), and malignant melanoma (69); its expression level was also
increased in a number of other cancer types, including colorectal
cancer, pancreatic cancer, mammary carcinoma, leukemia (70) and glioblastoma (71).
The CTLD and the cytoplasmic domain of
thrombomodulin were also discovered to be necessary for reduced
cell proliferation (69); it was
revealed that the cytoplasmic domain of thrombomodulin interacted
with the N-terminal membrane-cytoskeleton linker
ezrin/radixin/moesin (ERM) family protein ezrin. Thrombomodulin,
ezrin and F-actin were shown to co-localize at intercellular
filopodia, and the interaction between ezrin and CD44 has been
reported to facilitate cancer cell migration. It was also
hypothesized that the thrombomodulin-ezrin interaction may prevent
the binding of CD44 to ezrin, resulting in the reduced migration of
thrombomodulin-expressing cells compared with thrombomodulin
knockdown cells (72).
In other epithelial and tumor cell lines, the
expression of thrombomodulin and Snail was inversely correlated;
Snail is a transcription factor involved in epithelial mesenchymal
transition (EMT) that has been shown to bind the thrombomodulin
promoter and suppress its expression (62). Thrombomodulin was also able to
reverse EMT by upregulating E-cadherin and downregulating
N-cadherin expression levels in lung cancer cells (73). By contrast, the recombinant
thrombomodulin fragment TMD23 (with a 6-tandem EGF-like domain and
O-glycosylation site-rich domain) was reported to stimulate
angiogenesis (63,74). The C loop of the C-terminal
sub-domain of the fifth EGF-like domain of TMD23 has pro-angiogenic
and cytoprotective effects, in a G protein-coupled receptor
15-dependent manner (75).
Angiogenesis was also mediated by the phosphorylation of ERK1/2,
p38, protein kinase B (Akt) and Endothelial nitric oxide synthase
(eNOS) (74), and by Fibroblast
growth factor receptor 1-A (76).
rTMD23 was reported to stimulate the endothelial cell expression of
MMPs and plasminogen activators that mediate ECM degradation, and
subsequently, angiogenesis (74).
This suggests that rTMD23 is responsible for cellular proliferation
and migration, and that the CTLD may possess anti-angiogenic
properties.
CD93 (also known as complement component C1q
receptor and AA4.1) is a type I transmembrane glycoprotein with one
C-type lectin-like domain, five tandem EGF-like domain repeats, a
serine threonine-rich mucin-like domain, a transmembrane domain and
a cytoplasmic domain (77). CD93 is
located on chromosome 20p11.21 and has two exons separated by a
single intron (78). CD93 was
identified as one of the top 20 genes in the core human primary
tumor angiogenesis signature (34).
Its expression is prominent in ECs and certain hematopoietic
subsets, including myeloid cells, platelets and hematopoietic stem
cells (79–81). It is also highly expressed in tumor
ECs, but exhibits low expression levels in non-proliferating ECs
(34,82–85). A
soluble form of the CD93 ectodomain (containing the CTLD and
EGF-like domain repeats) was detected in normal human plasma
(80) and during inflammatory
stimulation in vivo (86).
In situ hybridization of mouse CD93 revealed its expression
in the vascular endothelium of 9 day old embryos; this correlated
with the remodeling of blood vessels in the intersomitic branches
of the dorsal aorta and developing perineural plexus, suggesting an
angiogenic role for CD93 (87).
The CTLD of CD93 is essential for intercellular
adhesion not sensitive to calcium chelators (79). Also, CD93 knockdown inhibited tube
formation, migration and adhesion of ECs (82), and the growth of
orthotopically-implanted syngeneic GL261 gliomas was retarded in
CD93−/− mice, which was associated with abnormal tumor
vessel growth (82).
The cytoplasmic domain of CD93 interacts with the
ERM protein moesin, establishing a link between CD93 and actin that
contributes to cytoskeletal reorganization, an essential process
for cellular adhesion (88). CD93
also interacts with dystroglycan, an ECM receptor and
laminin-binding protein. Through dystroglycan, the tyrosine
residues in the cytoplasmic domain of CD93 are phosphorylated
following adhesion to laminin, and this phosphorylation is
necessary for endothelial cell migration (89). In addition to the CTLD, the DX domain
(a 79 residue-long stretch situated between the CTLD and 5 EGF-like
domain repeats) is necessary for the interaction between CD93 and
MMRN2; blocking this interaction by targeting the DX domain
retarded the angiogenesis (90). In
endothelial filopodia, interaction with MMRN2 stabilizes CD93 to
prevent the shedding of its extracellular domain. This stable
complex is required for the activation of β1 integrin, which
initiates the phosphorylation of FAK and the organization of
fibronectin into fibrillar structures (43).
The recombinant CD93 protein rCD93-D23 (containing
the EGF domain and a serine-threonine rich-mucin-like domain)
induced HUVEC proliferation and migration via the ERK1/2,
PI3K/Akt/eNOS pathways and EGFR signaling; the CTLD of this protein
did not influence angiogenesis in vivo (or did so to a
moderate degree only), as indicted by the removal of the entire
ectodomain (including the CTLD), and suggesting that the CTLD of
CD93 may possess an anti-angiogenic function (91). Monoclonal antibody-targeting of the
extracellular domain of CD93 inhibited the proliferation, migration
and sprouting of ECs without influencing endothelial cell survival
and the inhibition of angiogenesis was suggested to result from the
prohibition of cell adhesion. The epitope recognized by this
antibody is in the region overlapping the CTLD and DX domain, and
lies outside of the EGF domain; it does not impair CD93-dependent
EGFR activation (which is dependent on the EGF domain) suggesting
that CD93 may possess a different angiogenic function as a
membrane-intercalated protein than insoluble form (92).
The extracellular domain of endosialin [also known
as tumor endothelial marker 1 (TEM1) and CD248] comprises a CTLD, a
Sushi domain (also known as a short consensus repeat or complement
control protein domain) and three EGF-like domains, followed by a
transmembrane and a cytoplasmic domain (93). Endosialin was initially identified as
a cell surface glycoprotein and TEM (94), though further studies have suggested
endosialin as a marker of cancer-associated fibroblasts (CAFs) and
tumor vessel associated mural cells, rather than a mesenchymal stem
cell (MSC) marker. The expression of endosialin by both CAFs and
MSCs indicates the involvement of the latter in tumor stroma
formation via differentiation into tumor stromal fibroblasts
(95). Furthermore,
endosialin-downregulated fibroblasts showed a platelet-derived
growth factor-BB-mediated reduction in migration and proliferation
(96).
The reported binding partners of endosialin are
metastasis-related protein Mac2-BP/90K (97) and MMRN2 (44). Its CTLD interacts with ECM proteins
such as collagen type I/IV and fibronectin, both of which mediate
cell adhesion and migration. CHO cells overexpressing endosialin
proliferated in clusters to form web-like structures, that formed
larger clusters over time compared with normal CHO cells, which
proliferate as singular cells (98).
Endosialin-null mice showed normal physiological angiogenesis;
however, there was a reduction in tumor growth, an increased number
of small and immature tumor vessels and a decreased number of
larger and mature vessels, highlighting the importance of
endosialin in tumor microvasculature maturation (99). Moreover, mice lacking the cytoplasmic
domain of endosialin exhibited reduced expression levels of VEGF,
hypoxia-inducible factor-1α (HIF1α), placental growth factor
(PlGF), MMP9 (20), and increased
expression levels of the tumor suppressor transgelin (SM22α) and
the downstream effector of Notch (100). VEGF, HIF1α, P1GF and MMP9 are
pro-angiogenic, whereas SM22α is known to have tumor suppressive
properties (101). Depending on the
tissue and cellular context, Notch can be either oncogenic or tumor
suppressive (102). It was also
revealed that endosialin expression was induced in hypoxic
conditions and regulated by HIF2α; endosialin transcription was
also enhanced by the interaction of HIF2α with the
proto-oncoprotein protein C-ets-1 (103).
As a number of pathways are able to compensate for
the VEGF-targeted inhibition of angiogenesis, anti-angiogenic
therapies that target VEGF alone are not sufficient (104). Acquired resistance to the
inhibition of VEGF signaling, and its toxicity towards normal
physiology demand a broader range of therapeutic approaches,
targeting multiple aspects of angiogenesis.
The members of the C-type lectin family XIV are
transmembrane proteins expressed at the cell surface, and are
therefore relatively easy to target. With the exception of
thrombomodulin, C-type lectin family XIV members are predominantly
expressed by tumor ECs. Endosialin is expressed by the tumor
vasculature, tumor stromal cells and MSCs, making it an attractive
target for anti-angiogenic therapy in various types of tumor
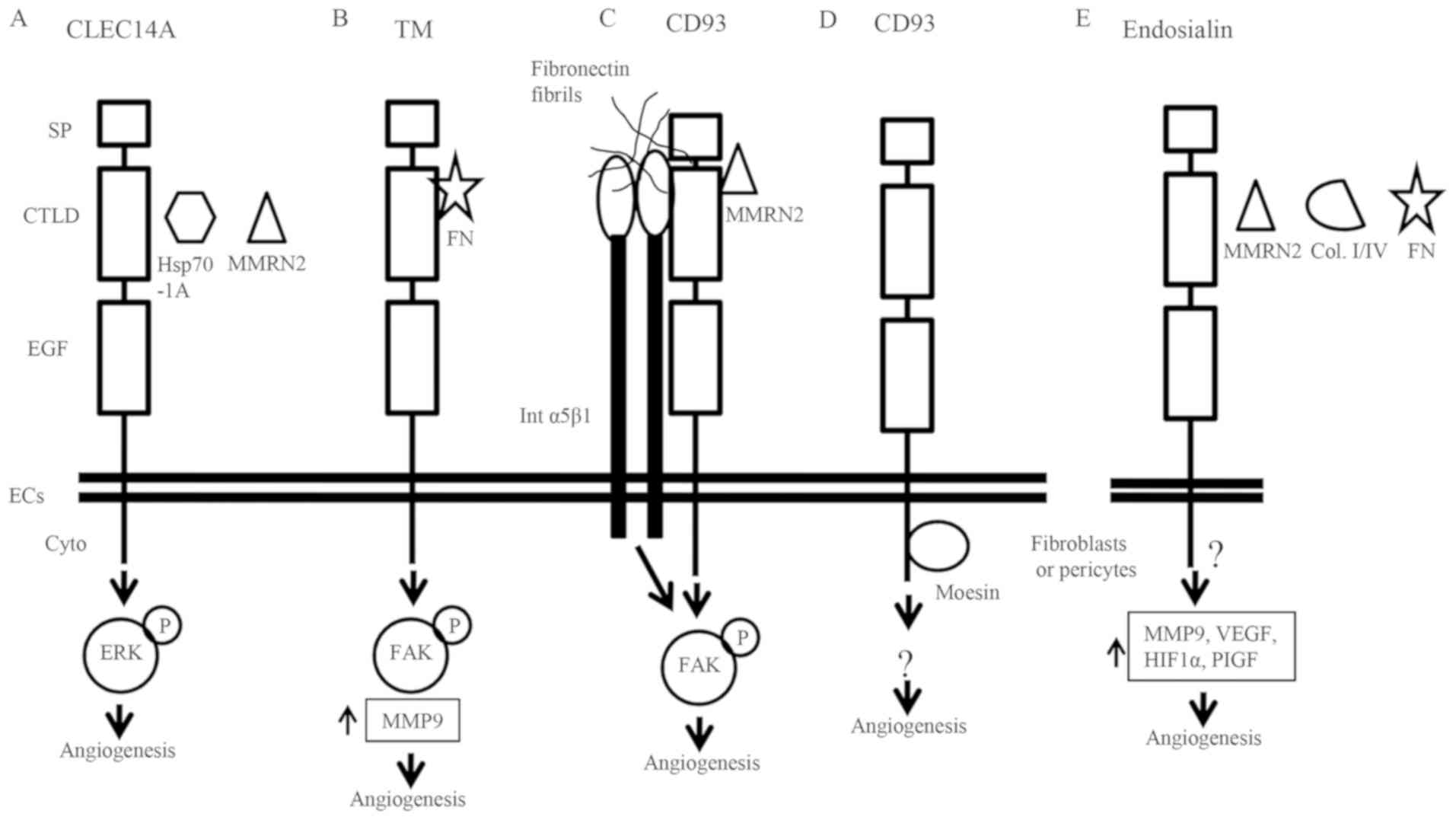
(95). CLEC14A induces filopodia and
tube formation (32), and its
interaction with MMRN2 promotes tumor growth and angiogenesis
(45). Thrombomodulin interacts with
fibronectin during tumor angiogenesis and maintains the endothelial
tube structure (59). CD93 interacts
with MMRN2, fibronectin fibrils and α5β1 integrin, promoting
angiogenesis via FAK phosphorylation (43). The interaction between the
cytoplasmic tail of CD93 and moesin induces cytoskeletal
reorganization (88), and the
interaction between endosialin, collagen and fibronectin mediates
cell adhesion and migration (98).
As discussed, all members of the C-type lectin
family XIV interact with ECM proteins and support endothelial cell
migration. Inhibiting the function of these proteins may lead to
reduced endothelial cell migration and angiogenesis. Particular
family members also promote downstream signaling mechanisms, such
as the phosphorylation of FAK and an increase in the expression
levels of MMP9 and plasminogen activators (Fig. 2). The C-type lectin family XIV
members have been reported to enhance angiogenesis in different
cancer types (Table I); however,
these findings do not clearly illustrate the mechanisms by which
they regulate angiogenesis. The CTLD has been shown to exert both
pro-and anti-angiogenic activity in different members of C-type
lectin family XIV. Additionally, the EGF-like domain is necessary
for their angiogenic capacity, and may be a potential target for
anti-angiogenic therapy.
Improved characterization of the structural motifs
and domains of members of C-type lectin family XIV will aid in the
understanding of their mechanisms of signal transduction and
angiogenesis. Specific binding partners of the family are known
(Table II), yet detailed mechanisms
of the roles of these proteins in angiogenesis require further
elucidation. It is evident that members of C-type lectin family XIV
are important regulators of physiological and pathological
angiogenesis, and therefore present as attractive therapeutic
targets.
The authors would like to thank Mr S. Rajivgandhi
(Institute of Life Sciences, Bhubaneswar) for comments on the
manuscript.
RKS is supported by DBT grant
(6242-P64/RGCB/PMD/DBT/RJKS/2015), SERB-EMR (EMR/2016/003780) and
intramural funds from ILS, which is an autonomous institute of DBT,
Government of India. SB is a recipient of DBT-SRF.
Not applicable.
RKS and DV conceptualized the manuscript. SB wrote
the manuscript with input from RKS and DV. RKS and DV critically
reviewed and edited the manuscript. All authors have read and
approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Risau W: Mechanisms of angiogenesis.
Nature. 386:671–674. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Folkman J: Angiogenesis in cancer,
vascular, rheumatoid and other disease. Nat Med. 1:27–31. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Klagsbrun M and D'Amore PA: Regulators of
angiogenesis. Ann Review Physiol. 53:217–239. 1991. View Article : Google Scholar
|
|
4
|
Folkman J: Fundamental concepts of the
angiogenic process. Curr Mol Med. 3:643–651. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Patel-Hett S and D'Amore PA: Signal
transduction in vasculogenesis and developmental angiogenesis. Int
J Dev Biol. 55:353–363. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Neve A, Cantatore FP, Maruotti N, Corrado
A and Ribatti D: Extracellular matrix modulates angiogenesis in
physiological and pathological conditions. Biomed Res Int.
2014:7560782014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bischoff J: Cell adhesion and
angiogenesis. J Clin Inves. 99:373–376. 1997. View Article : Google Scholar
|
|
9
|
Ramjaun AR and Hodivala-Dilke K: The role
of cell adhesion pathways in angiogenesis. Int J Biochem Cell Biol.
41:521–530. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mignatti P and Rifkin DB: Plasminogen
activators and matrix metalloproteinases in angiogenesis. Enzyme
Protein. 49:117–137. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lamalice L, Le Boeuf F and Huot J:
Endothelial cell migration during angiogenesis. Circ Res.
100:782–794. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Conway EM, Collen D and Carmeliet P:
Molecular mechanisms of blood vessel growth. Cardiovasc Res.
49:507–521. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Papetti M and Herman IM: Mechanisms of
normal and tumor-derived angiogenesis. American journal of
physiology. Cell Physiol. 282:C947–C970. 2002. View Article : Google Scholar
|
|
14
|
Dusse LM, Carvalho MG, Getliffe K, Voegeli
D, Cooper AJ and Lwaleed BA: Increased circulating thrombomodulin
levels in pre-eclampsia. Clin Chim Acta. 387:168–171. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wada H, Minamikawa K, Wakita Y, Nakase T,
Kaneko T, Ohiwa M, Tamaki S, Deguchi K, Shirakawa S, Hayashi T, et
al: Increased vascular endothelial cell markers in patients with
disseminated intravascular coagulation. Am J Hematol. 44:85–88.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mori Y, Wada H, Okugawa Y, Tamaki S,
Nakasaki T, Watanabe R, Gabazza EC, Nishikawa M, Minami N and Shiku
H: Increased plasma thrombomodulin as a vascular endothelial cell
marker in patients with thrombotic thrombocytopenic purpura and
hemolytic uremic syndrome. Clin Appl Thromb Hemost. 7:5–9. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Stratton RJ, Pompon L, Coghlan JG, Pearson
JD and Black CM: Soluble thrombomodulin concentration is raised in
scleroderma associated pulmonary hypertension. Ann Rheum Dis.
59:132–134. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dohi Y, Ohashi M, Sugiyama M, Takase H,
Sato K and Ueda R: Circulating thrombomodulin levels are related to
latent progression of atherosclerosis in hypertensive patients.
Hypertens Res. 26:479–483. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Papadopoulos DP, Thomopoulos C, Mourouzis
I, Kotrotsou A, Sanidas E, Papazachou U, Daskalaki M and Makris TK:
Masked hypertension unfavourably affects haemostasis parameters.
Blood Press. 20:218–221. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Maia M, de Vriese A, Janssens T, Moons M,
van Landuyt K, Tavernier J, Lories RJ and Conway EM: CD248 and its
cytoplasmic domain: A therapeutic target for arthritis. Arthritis
Rheum. 62:3595–3606. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lax S, Hou TZ, Jenkinson E, Salmon M,
MacFadyen JR, Isacke CM, Anderson G, Cunningham AF and Buckley CD:
CD248/Endosialin is dynamically expressed on a subset of stromal
cells during lymphoid tissue development, splenic remodeling and
repair. FEBS Lett. 581:3550–3556. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jeon JW, Jung JG, Shin EC, Choi HI, Kim
HY, Cho ML, Kim SW, Jang YS, Sohn MH, Moon JH, et al: Soluble CD93
induces differentiation of monocytes and enhances TLR responses. J
Immunol. 185:4921–4927. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Moosig F, Fahndrich E, Knorr-Spahr A,
Böttcher S, Ritgen M, Zeuner R, Kneba M and Schröder JO: C1qRP
(CD93) expression on peripheral blood monocytes in patients with
systemic lupus erythematosus. Rheumatol Int. 26:1109–1112. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
van der Net JB, Oosterveer DM, Versmissen
J, Defesche JC, Yazdanpanah M, Aouizerat BE, Steyerberg EW, Malloy
MJ, Pullinger CR, Kastelein JJ and Kane JP: Replication study of 10
genetic polymorphisms associated with coronary heart disease in a
specific high-risk population with familial hypercholesterolemia.
Eur Heart J. 29:2195–2201. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Drickamer K: Demonstration of
carbohydrate-recognition activity in diverse proteins which share a
common primary structure motif. Biochem Soci Trans. 17:13–15. 1989.
View Article : Google Scholar
|
|
26
|
Drickamer K: C-type lectin-like domains.
Curr Opinion Struct Biol. 9:585–590. 1999. View Article : Google Scholar
|
|
27
|
Drickamer K and Fadden AJ: Genomic
analysis of C-type lectins. Biochem Soci Symp. 59–72. 2002.
View Article : Google Scholar
|
|
28
|
Zelensky AN and Gready JE: C-type
lectin-like domains in Fugu rubripes. BMC Genomics. 5:512004.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Drickamer K: Evolution of Ca(2+)-dependent
animal lectins. Prog Nucleic Acid Res Mol Biol. 45:207–232. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zelensky AN and Gready JE: The C-type
lectin-like domain superfamily. FEBS J. 272:6179–6217. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rho SS, Choi HJ, Min JK, Lee HW, Park H,
Park H, Kim YM and Kwon YG: Clec14a is specifically expressed in
endothelial cells and mediates cell to cell adhesion. Biochem
Biophys Res Commun. 404:103–108. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mura M, Swain RK, Zhuang X, Vorschmitt H,
Reynolds G, Durant S, Beesley JF, Herbert JM, Sheldon H, Andre M,
et al: Identification and angiogenic role of the novel tumor
endothelial marker CLEC14A. Oncogene. 31:293–305. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Du J, Yang Q, Luo L and Yang D: C1qr and
C1qrl redundantly regulate angiogenesis in zebrafish through
controlling endothelial Cdh5. Biochem Biophys Res Commun.
483:482–487. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Masiero M, Simoes FC, Han HD, Snell C,
Peterkin T, Bridges E, Mangala LS, Wu SY, Pradeep S, Li D, et al: A
core human primary tumor angiogenesis signature identifies the
endothelial orphan receptor ELTD1 as a key regulator of
angiogenesis. Cancer Cell. 24:229–241. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Delcourt N, Quevedo C, Nonne C, Fons P,
O'Brien D, Loyaux D, Diez M, Autelitano F, Guillemot JC, Ferrara P,
et al: Targeted identification of sialoglycoproteins in hypoxic
endothelial cells and validation in zebrafish reveal roles for
proteins in angiogenesis. J Biol Chem. 290:3405–3417. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ki MK, Jeoung MH, Choi JR, Rho SS, Kwon
YG, Shim H, Chung J, Hong HJ, Song BD and Lee S: Human antibodies
targeting the C-type lectin-like domain of the tumor endothelial
cell marker clec14a regulate angiogenic properties in vitro.
Oncogene. 32:5449–5457. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kim TK, Park CS, Jang J, Kim MR, Na HJ,
Lee K, Kim HJ, Heo K, Yoo BC, Kim YM, et al: Inhibition of
VEGF-dependent angiogenesis and tumor angiogenesis by an optimized
antibody targeting CLEC14a. Mol Oncol. 12:356–372. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zanivan S, Maione F, Hein MY,
Hernández-Fernaud JR, Ostasiewicz P, Giraudo E and Mann M:
SILAC-based proteomics of human primary endothelial cell
morphogenesis unveils tumor angiogenic markers. Mol Cell
Proteomics. 12:3599–3611. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Dev KK: Making protein interactions
druggable: Targeting PDZ domains. Nat Rev Drug Discov. 3:1047–1056.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kim S, Bell K, Mousa SA and Varner JA:
Regulation of angiogenesis in vivo by ligation of integrin
alpha5beta1 with the central cell-binding domain of fibronectin. Am
J Pathol. 156:1345–1362. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Huang X, Ji G, Wu Y, Wan B and Yu L:
LAMA4, highly expressed in human hepatocellular carcinoma from
Chinese patients, is a novel marker of tumor invasion and
metastasis. J Cancer Res Clin Oncol. 134:705–714. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lorenzon E, Colladel R, Andreuzzi E,
Marastoni S, Todaro F, Schiappacassi M, Ligresti G, Colombatti A
and Mongiat M: MULTIMERIN2 impairs tumor angiogenesis and growth by
interfering with VEGF-A/VEGFR2 pathway. Oncogene. 31:3136–3147.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lugano R, Vemuri K, Yu D, Bergqvist M,
Smits A, Essand M, Johansson S, Dejana E and Dimberg A: CD93
promotes beta1 integrin activation and fibronectin fibrillogenesis
during tumor angiogenesis. J Clin Invest. 128:3280–3297. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Khan KA, Naylor AJ, Khan A, Noy PJ,
Mambretti M, Lodhia P, Athwal J, Korzystka A, Buckley CD, Willcox
BE, et al: Multimerin-2 is a ligand for group 14 family C-type
lectins CLEC14A, CD93 and CD248 spanning the endothelial pericyte
interface. Oncogene. 36:6097–6108. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Noy PJ, Lodhia P, Khan K, Zhuang X, Ward
DG, Verissimo AR, Bacon A and Bicknell R: Blocking CLEC14A-MMRN2
binding inhibits sprouting angiogenesis and tumour growth.
Oncogene. 34:5821–5831. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Jang J, Kim MR, Kim TK, Lee WR, Kim JH,
Heo K and Lee S: CLEC14a-HSP70-1A interaction regulates
HSP70-1A-induced angiogenesis. Sci Re. 7:106662017.
|
|
47
|
Noy PJ, Swain RK, Khan K, Lodhia P and
Bicknell R: Sprouting angiogenesis is regulated by shedding of the
C-type lectin family 14, member A (CLEC14A) ectodomain, catalyzed
by rhomboid-like 2 protein (RHBDL2). FASEB J. 30:2311–2323. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lee S, Rho SS, Park H, Park JA, Kim J, Lee
IK, Koh GY, Mochizuki N, Kim YM and Kwon YG: Carbohydrate-binding
protein CLEC14A regulates VEGFR-2- and VEGFR-3-dependent signals
during angiogenesis and lymphangiogenesis. J Clin Invest.
127:457–471. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Tammela T, Zarkada G, Nurmi H, Jakobsson
L, Heinolainen K, Tvorogov D, Zheng W, Franco CA, Murtomäki A,
Aranda E, et al: VEGFR-3 controls tip to stalk conversion at vessel
fusion sites by reinforcing Notch signalling. Nat Cell Biol.
13:1202–1213. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Suzuki K, Kusumoto H, Deyashiki Y,
Nishioka J, Maruyama I, Zushi M, Kawahara S, Honda G, Yamamoto S
and Horiguchi S: Structure and expression of human thrombomodulin,
a thrombin receptor on endothelium acting as a cofactor for protein
C activation. EMBO J. 6:1891–1897. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Conway EM: Thrombomodulin and its role in
inflammation. Semin Immunopathol. 34:107–125. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Maruyama I, Bell CE and Majerus PW:
Thrombomodulin is found on endothelium of arteries, veins,
capillaries, and lymphatics, and on syncytiotrophoblast of human
placenta. J Cell Biol. 101:363–371. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Soff GA, Jackman RW and Rosenberg RD:
Expression of thrombomodulin by smooth muscle cells in culture:
Different effects of tumor necrosis factor and cyclic adenosine
monophosphate on thrombomodulin expression by endothelial cells and
smooth muscle cells in culture. Blood. 77:515–518. 1991.PubMed/NCBI
|
|
54
|
McCachren SS, Diggs J, Weinberg JB and
Dittman WA: Thrombomodulin expression by human blood monocytes and
by human synovial tissue lining macrophages. Blood. 78:3128–3132.
1991.PubMed/NCBI
|
|
55
|
Calnek DS and Grinnell BW:
Thrombomodulin-dependent anticoagulant activity is regulated by
vascular endothelial growth factor. Exp Cell Res. 238:294–298.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Huang HC, Shi GY, Jiang SJ, Shi CS, Wu CM,
Yang HY and Wu HL: Thrombomodulin-mediated cell adhesion:
Involvement of its lectin-like domain. J Biol Chem.
278:46750–46759. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Suehiro T, Shimada M, Matsumata T,
Taketomi A, Yamamoto K and Sugimachi K: Thrombomodulin inhibits
intrahepatic spread in human hepatocellular carcinoma. Hepatology.
21:1285–1290. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Tabata M, Sugihara K, Yonezawa S,
Yamashita S and Maruyama I: An immunohistochemical study of
thrombomodulin in oral squamous cell carcinoma and its association
with invasive and metastatic potential. J Oral Pathol Med.
26:258–264. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Hsu YY, Shi GY, Wang KC, Ma CY, Cheng TL
and Wu HL: Thrombomodulin promotes focal adhesion kinase activation
and contributes to angiogenesis by binding to fibronectin.
Oncotarget. 7:68122–68139. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Shen TL, Park AY, Alcaraz A, Peng X, Jang
I, Koni P, Flavell RA, Gu H and Guan JL: Conditional knockout of
focal adhesion kinase in endothelial cells reveals its role in
angiogenesis and vascular development in late embryogenesis. J Cell
Biol. 169:941–952. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Peng X, Ueda H, Zhou H, Stokol T, Shen TL,
Alcaraz A, Nagy T, Vassalli JD and Guan JL: Overexpression of focal
adhesion kinase in vascular endothelial cells promotes angiogenesis
in transgenic mice. Cardiovasc Rese. 64:421–430. 2004. View Article : Google Scholar
|
|
62
|
Kao YC, Wu LW, Shi CS, Chu CH, Huang CW,
Kuo CP, Sheu HM, Shi GY and Wu HL: Downregulation of
thrombomodulin, a novel target of Snail, induces tumorigenesis
through epithelial-mesenchymal transition. Mol Cell Biol.
30:4767–4785. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Kuo CH, Chen PK, Chang BI, Sung MC, Shi
CS, Lee JS, Chang CF, Shi GY and Wu HL: The recombinant lectin-like
domain of thrombomodulin inhibits angiogenesis through interaction
with Lewis Y antigen. Blood. 119:1302–1313. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Hosaka Y, Higuchi T, Tsumagari M and Ishii
H: Inhibition of invasion and experimental metastasis of murine
melanoma cells by human soluble thrombomodulin. Cancer Lett.
161:231–240. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Conway EM, Van de Wouwer M, Pollefeyt S,
Jurk K, Van Aken H, De Vriese A, Weitz JI, Weiler H, Hellings PW,
Schaeffer P, et al: The lectin-like domain of thrombomodulin
confers protection from neutrophil-mediated tissue damage by
suppressing adhesion molecule expression via nuclear factor kappaB
and mitogen-activated protein kinase pathways. J Exp Med.
196:565–577. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Hamatake M, Ishida T, Mitsudomi T, Akazawa
K and Sugimachi K: Prognostic value and clinicopathological
correlation of thrombomodulin in squamous cell carcinoma of the
human lung. Clin Cancer Res. 2:763–766. 1996.PubMed/NCBI
|
|
67
|
Tezuka Y, Yonezawa S, Maruyama I,
Matsushita Y, Shimizu T, Obama H, Sagara M, Shirao K, Kusano C,
Natsugoe S, et al: Expression of thrombomodulin in esophageal
squamous cell carcinoma and its relationship to lymph node
metastasis. Cancer Res. 55:4196–4200. 1995.PubMed/NCBI
|
|
68
|
Hanly AM, Redmond M, Winter DC, Brophy S,
Deasy JM, Bouchier-Hayes DJ and Kay EW: Thrombomodulin expression
in colorectal carcinoma is protective and correlates with survival.
Br J Cancer. 94:1320–1325. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Zhang Y, Weiler-Guettler H, Chen J,
Wilhelm O, Deng Y, Qiu F, Nakagawa K, Klevesath M, Wilhelm S,
Böhrer H, et al: Thrombomodulin modulates growth of tumor cells
independent of its anticoagulant activity. J Clin Invest.
101:1301–1309. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
70
|
Lindahl AK, Boffa MC and Abildgaard U:
Increased plasma thrombomodulin in cancer patients. Thromb Haemost.
69:112–114. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Salmaggi A, Eoli M, Frigerio S, Ciusani E,
Silvani A and Boiardi A: Circulating intercellular adhesion
molecule-1 (ICAM-1), vascular cell adhesion molecule-1 (VCAM-1) and
plasma thrombomodulin levels in glioblastoma patients. Cancer Lett.
146:169–172. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Hsu YY, Shi GY, Kuo CH, Liu SL, Wu CM, Ma
CY, Lin FY, Yang HY and Wu HL: Thrombomodulin is an
ezrin-interacting protein that controls epithelial morphology and
promotes collective cell migration. FASEB J. 26:3440–3452. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Zheng N, Huo Z, Zhang B, Meng M, Cao Z,
Wang Z and Zhou Q: Thrombomodulin reduces tumorigenic and
metastatic potential of lung cancer cells by up-regulation of
E-cadherin and down-regulation of N-cadherin expression. Biochem
Biophys Res Commun. 476:252–259. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Shi CS, Shi GY, Chang YS, Han HS, Kuo CH,
Liu C, Huang HC, Chang YJ, Chen PS and Wu HL: Evidence of human
thrombomodulin domain as a novel angiogenic factor. Circulation.
111:1627–1636. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Wang X, Pan B, Honda G, Wang X, Hashimoto
Y, Ohkawara H, Xu K, Zeng L and Ikezoe T: Cytoprotective and
pro-angiogenic functions of thrombomodulin are preserved in the C
loop of the fifth epidermal growth factor-like domain.
Haematologica. 103:1730–1740. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Kuo CH, Sung MC, Chen PK, Chang BI, Lee
FT, Cho CF, Hsieh TT, Huang YC, Li YH, Shi GY, et al: FGFR1
mediates recombinant thrombomodulin domain-induced angiogenesis.
Cardiovasc Res. 105:107–117. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Nepomuceno RR, Henschen-Edman AH, Burgess
WH and Tenner AJ: cDNA cloning and primary structure analysis of
C1qR(P), the human C1q/MBL/SPA receptor that mediates enhanced
phagocytosis in vitro. Immunity. 6:119–129. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Malarstig A, Silveira A, Wagsater D,
Öhrvik J, Bäcklund A, Samnegård A, Khademi M, Hellenius ML, Leander
K, Olsson T, et al: Plasma CD93 concentration is a potential novel
biomarker for coronary artery disease. J Intern Med. 270:229–236.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
McGreal EP, Ikewaki N, Akatsu H, Morgan BP
and Gasque P: Human C1qRp is identical with CD93 and the mNI-11
antigen but does not bind C1q. J Immunol. 168:5222–5232. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Bohlson SS, Silva R, Fonseca MI and Tenner
AJ: CD93 is rapidly shed from the surface of human myeloid cells
and the soluble form is detected in human plasma. J Immunol.
175:1239–1247. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Nepomuceno RR and Tenner AJ: C1qRP, the
C1q receptor that enhances phagocytosis, is detected specifically
in human cells of myeloid lineage, endothelial cells, and
platelets. J Immunol. 160:1929–1935. 1998.PubMed/NCBI
|
|
82
|
Langenkamp E, Zhang L, Lugano R, Huang H,
Elhassan TE, Georganaki M, Bazzar W, Lööf J, Trendelenburg G,
Essand M, et al: Elevated expression of the C-type lectin CD93 in
the glioblastoma vasculature regulates cytoskeletal rearrangements
that enhance vessel function and reduce host survival. Cancer Res.
75:4504–4516. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Bao L, Tang M, Zhang Q, You B, Shan Y, Shi
S, Li L, Hu S and You Y: Elevated expression of CD93 promotes
angiogenesis and tumor growth in nasopharyngeal carcinoma. Biochem
Biophys Res Commun. 476:467–474. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Tosi GM, Caldi E, Parolini B, Toti P, Neri
G, Nardi F, Traversi C, Cevenini G, Marigliani D, Nuti E, et al:
CD93 as a potential target in neovascular age-related macular
degeneration. J Cell Physiol. 232:1767–1773. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Dieterich LC, Mellberg S, Langenkamp E,
Zhang L, Zieba A, Salomäki H, Teichert M, Huang H, Edqvist PH,
Kraus T, et al: Transcriptional profiling of human glioblastoma
vessels indicates a key role of VEGF-A and TGFbeta2 in vascular
abnormalization. J Pathol. 228:378–390. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Greenlee MC, Sullivan SA and Bohlson SS:
Detection and characterization of soluble CD93 released during
inflammation. Inflamm Res. 58:909–919. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Petrenko O, Beavis A, Klaine M, Kittappa
R, Godin I and Lemischka IR: The molecular characterization of the
fetal stem cell marker AA4. Immunity. 10:691–700. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Zhang M, Bohlson SS, Dy M and Tenner AJ:
Modulated interaction of the ERM protein, moesin, with CD93.
Immunology. 115:63–73. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Galvagni F, Nardi F, Maida M, Bernardini
G, Vannuccini S, Petraglia F, Santucci A and Orlandini M: CD93 and
dystroglycan cooperation in human endothelial cell adhesion and
migration adhesion and migration. Oncotarget. 7:10090–10103. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Galvagni F, Nardi F, Spiga O, Trezza A,
Tarticchio G, Pellicani R, Andreuzzi E, Caldi E, Toti P, Tosi GM,
et al: Dissecting the CD93-Multimerin 2 interaction involved in
cell adhesion and migration of the activated endothelium. Matrix
Biol. 64:112–127. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Kao YC, Jiang SJ, Pan WA, Wang KC, Chen
PK, Wei HJ, Chen WS, Chang BI, Shi GY and Wu HL: The epidermal
growth factor-like domain of CD93 is a potent angiogenic factor.
PLoS One. 7:e516472012. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Orlandini M, Galvagni F, Bardelli M,
Rocchigiani M, Lentucci C, Anselmi F, Zippo A, Bini L and Oliviero
S: The characterization of a novel monoclonal antibody against CD93
unveils a new antiangiogenic target. Oncotarget. 5:2750–2760. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Christian S, Ahorn H, Koehler A,
Eisenhaber F, Rodi HP, Garin-Chesa P, Park JE, Rettig WJ and Lenter
MC: Molecular cloning and characterization of endosialin, a C-type
lectin-like cell surface receptor of tumor endothelium. J Biol
Chem. 276:7408–7414. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Rettig WJ, Garin-Chesa P, Healey JH, Su
SL, Jaffe EA and Old LJ: Identification of endosialin, a cell
surface glycoprotein of vascular endothelial cells in human cancer.
Proc Natl Acad Sci USA. 89:10832–10836. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Bagley RG, Weber W, Rouleau C, Yao M,
Honma N, Kataoka S, Ishida I, Roberts BL and Teicher BA: Human
mesenchymal stem cells from bone marrow express tumor endothelial
and stromal markers. Int J Oncol. 34:619–627. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Christian S, Winkler R, Helfrich I, Boos
AM, Besemfelder E, Schadendorf D and Augustin HG: Endosialin (Tem1)
is a marker of tumor-associated myofibroblasts and tumor
vessel-associated mural cells. Am J Pathol. 172:486–494. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Becker R, Lenter MC, Vollkommer T, Boos
AM, Pfaff D, Augustin HG and Christian S: Tumor stroma marker
endosialin (Tem1) is a binding partner of metastasis-related
protein Mac-2 BP/90K. FASEB J. 22:3059–3067. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Tomkowicz B, Rybinski K, Foley B, Ebel W,
Kline B, Routhier E, Sass P, Nicolaides NC, Grasso L and Zhou Y:
Interaction of endosialin/TEM1 with extracellular matrix proteins
mediates cell adhesion and migration. Proc Natl Acad Sci USA.
104:17965–17970. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Nanda A, Karim B, Peng Z, Liu G, Qiu W,
Gan C, Vogelstein B, St Croix B, Kinzler KW and Huso DL: Tumor
endothelial marker 1 (Tem1) functions in the growth and progression
of abdominal tumors. Proc Natl Acad Sci USA. 103:3351–3356. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Maia M, DeVriese A, Janssens T, Moons M,
Lories RJ, Tavernier J and Conway EM: CD248 facilitates tumor
growth via its cytoplasmic domain. BMC Cancer. 11:1622011.
View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Yeo M, Park HJ, Kim DK, Kim YB, Cheong JY,
Lee KJ and Cho SW: Loss of SM22 is a characteristic signature of
colon carcinogenesis and its restoration suppresses colon
tumorigenicity in vivo and in vitro. Cancer. 116:2581–2589.
2010.PubMed/NCBI
|
|
102
|
Nowell CS and Radtke F: Notch as a tumour
suppressor. Nat Rev Cancer. 17:145–159. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Ohradanova A, Gradin K, Barathova M,
Zatovicova M, Holotnakova T, Kopacek J, Parkkila S, Poellinger L,
Pastorekova S and Pastorek J: Hypoxia upregulates expression of
human endosialin gene via hypoxia-inducible factor 2. Br J Cancer.
99:1348–1356. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Zhao Y and Adjei AA: Targeting
angiogenesis in cancer therapy: Moving beyond vascular endothelial
growth factor. Oncologist. 20:660–673. 2015. View Article : Google Scholar : PubMed/NCBI
|