Introduction
Prostate cancer is a common malignant tumor of the
male reproductive system. Its incidence has been rising with the
changes of social environment in recent years, and its mortality
rate ranks high among tumors of the urinary system (1,2). The
disease is difficult to be diagnosed in its early stage due to the
lack of effective diagnostic methods, so it has usually progressed
to the advanced stage when confirmed. Accordingly, many patients
with the disease cannot be operated for radical cure, which
seriously endangers their life and health (3). With the development of molecular
biology, the role of microRNA (miRNA) in tumors has been
increasingly valued, which also provides a new direction for the
diagnosis and treatment of prostate cancer.
As a non-coding single-stranded RNA, miRNA affects
the biological functions of cells through its complete or
incomplete complementary binding to the 3′-end of target genes
(4,5). miR-129 is a miRNA located in the
genomic region near the fragile site of chromosome 7q (6), and fragile site loss is closely related
to the malignancy of prostate cancer (7). miR-129-3p is a miRNA closely correlated
with the development and progression of tumors and the expression
is low in gastric cancer (8) and
breast cancer (9), functioning as a
tumor suppressor gene. Smad3 is a transporter that plays a pivotal
role in transforming growth factor-β (TGF-β) signaling pathway, and
it can promote the invasion and metastasis of tumor cells (10). In this study, a bioinformatics
website (TargetScan) predicted that Smad3 may be a target gene of
miR-129-3p.
In this study, the effects of miR-129-3p on the
biological functions of prostate cancer cells as well as its
potential targeted and regulatory mechanism were explored, so as to
provide more experimental data for the mechanism research of
prostate cancer.
Materials and methods
Experimental reagents and
materials
A total of 74 patients who were pathologically
diagnosed with prostate cancer and then underwent radical
prostatectomy in Gansu Provincial Hospital of TCM (Lanzhou, China)
from 2015 to 2018 were enrolled. All of them had stages I–III of
prostate cancer. Detailed information is shown in Table I. After receiving consent, their
prostate cancer and adjacent tissues (n=74 each) were obtained
during the operation and stored in a liquid nitrogen container.
Prostate cancer cells (PC-3, DU-145, and LNCaP cells) and human
prostate epithelial cell RWPE-1 (Shanghai Institute of Cell
Biology); fetal bovine serum (FBS) and trypsin (Gibco; Thermo
Fisher Scientific, Inc.); phosphate buffer solution (PBS) (Hyclone;
GE Healthcare Life Sciences); dimethyl sulfoxide (DMSO)
(Sigma-Aldrich; Merck KGaA); TRIzol reagent (Invitrogen; Thermo
Fisher Scientific, Inc.); dual luciferase reporter gene assay
detection kit (Solarbio); reverse transcription kit and PCR master
mix (Fermentas; Thermo Fisher Scientific, Inc.); RIPA and BCA
protein kit (Thermo Fisher Scientific, Inc.); Annexin V-FITC/PI
apoptosis kit (Jiangsu KeyGEN Bio TECH Corp., Ltd.); Transwell
chamber (Corning, Inc.); Matrigel (Beijing BioDee Biotechnology
Co., Ltd.); Smad3, Bax, Bcl-2 and β-actin antibodies (Cell
Signaling Technology); goat anti-rabbit IgG secondary antibody
(Wuhan Boster Biological Technology Co., Ltd.); ECL developer
(Thermo Fisher Scientific, Inc.). Primers for miR-129-3p and miR-NC
were designed and synthesized by Sangon Biotech Shanghai Co.,
Ltd.
 | Table I.General information. |
Table I.
General information.
| Information | Patients with
prostate cancer (n=74) |
|---|
| Age (years) | 58.34±8.46 |
| BMI
(kg/m2) | 22.89±1.22 |
| Pathological
types |
|
|
Adenocarcinoma | 25 (33.78) |
| Squamous
cell carcinoma | 27 (36.49) |
|
Adenosquamous carcinoma | 22 (29.73) |
| Pathological
stages |
|
| Stage
I | 21 (30.43) |
| Stage
II | 26 (37.68) |
| Stage
III | 22 (31.88) |
| Degree of
differentiation |
|
| High | 20 (28.99) |
|
Moderate | 23 (33.33) |
| Low | 26 (37.68) |
The study was approved by the Ethics Committee of
Gansu Provincial Hospital of TCM (Lanzhou, China).
RT-PCR detection of miR-129-3p and
Smad3 expression levels
The prostate cancer tissue and the adjacent tissue
were taken from the liquid nitrogen container for grinding. PC-3,
DU-145, LNCaP and RWPE-1 cells were prepared into a cell
suspension. The TRIzol reagent was used to extract total RNA from
the tissues and cells, and an ultraviolet spectrophotometer was
used to detect its purity and concentration. Each 5 µg of the total
RNA was reverse transcribed into cDNA according to the instructions
of the kits. The parameters were as follows: at 37°C for 15 min, at
42°C for 42 min, and at 70°C for 5 min. The transcribed cDNA was
used for PCR amplification, during which β-actin was considered as
an internal reference for Smad3 mRNA and U6 was considered as an
internal reference for miR-129-3p. The reaction system was as
follows: 1 µl of cDNA, each 0.2 µl of upstream and downstream
primers, 10 µl of 2X Real-time PCR Master mix, 0.4 µl of Passive
Reference Dye (50X), and ddH2O up to 20 µl. The
conditions for Smad3 mRNA were as follows: pre-denaturation at 95°C
for 5 min, and then cycling at 95°C for 30 sec and at 60°C for 45
sec for 40 times. The conditions for miR-129-3p were:
pre-denaturation at 95°C for 15 min, cycling at 94°C for 10 sec and
at 55°C for 40 sec, 40 times, and finally extension at 72°C for 30
sec. The upstream and downstream primers for miR-129-3p were
5′-CTTGTTGCGGTCTGG-3′ and 5′-TGCAGGGTCCGAGGT-3′. The primers for U6
were 5′-CTCGTCTCGGCAGCACA-3′ and 5′-AACGCTTCACGAATTTGCGT-3′. The
primers for Smad3 mRNA were 5′-GGAACTTACAAGGCGACAC-3′ and
5′-TGGGAGACTGGACGAAA-3′. The primers for β-actin were
5′-CCCATCTACGAGGGCTAT-3′ and 5′-TGTCACGCACGATTTCC-3′. The relative
expression of genes was expressed by 2−∆∆CT. A PCR
instrument was used for real-time fluorescence quantitative PCR.
The experiment was carried out 3 times. Calculation methods were as
follows: for all test samples and calibration samples, the CT value
of the internal reference gene was used to normalize that of the
target gene: ∆CT (test) = CT (target, test) - CT (ref, test) ∆CT
(calibrator) = CT (target, calibrator) - CT (ref, calibrator).
Next, the ∆CT value of the calibration samples was used to
normalize that of the test samples: ∆∆CT = ∆CT (test) - ∆CT
(calibrator). Finally, the ratio of expression level was
calculated: 2-∆∆CT = the ratio of expression.
Cell culture, passage, and
transfection
The detection of miR-129-3p and Smad3 RNA expression
levels showed that the expression level of miR-129-3p in PC-3 and
LNCaP cells was lower than that in DU-145 cells, so PC-3 and LNCaP
cells were selected for transfection and subsequent experiments.
The two kinds of cells were cultured in a DMEM containing 10% PBS
at 37°C and with 5% CO2. When the adherent growth and
fusion reached 85%, the cells were digested with 25% trypsin and
then continuously cultured in the medium for passage. Cas9 backbone
plasmid was digested with two restriction endonucleases
(KpnI and XhoI), and then the digested product was
recovered through agarose gel electrophoresis. The digested
fragment was about 5 kb. After that, miR-129-3p-mimics,
miR-129-3p-inhibitor, miR-NC, Si-Smad3, and Si-NC were,
respectively, transfected into cells in the logarithmic phase.
According to the instruction of the Lipofectamine 2000
manufacturer's kit, Lipofectamine 2000 was mixed with
miR-129-3p-mimics and then incubated at room temperature for 5 min.
Finally, the mixture was evenly mixed with the cells and then
transfected at 37°C with 5% CO2.
MTT assay for cell proliferation
The cells transfected for 48 h were inoculated into
a 96-well plate, ~100 µl/well and 2×103 cells/ml, and
then incubated at 37°C. At the first, third, and fifth days of the
incubation, the cells in each well were added with 20 µl of MTT
solution respectively, and then continuously cultured in an
incubator for 4 h. Then, they were added with 150 µl of DMSO and
shaken for 10 min. Finally, a microplate reader was used to measure
the optical density (OD) values at 490 nm, so as to detect cell
proliferation. The experiment was carried out 3 times.
Transwell chamber assay for cell
invasion in vitro
Matrigel was placed at 4°C overnight, and then the
liquefied Matrigel was diluted at 1:8. The transfected PC-3 cells
were starved for 24 h and then resuspended with FBS-free DMEM
culture solution. After the cell density was adjusted to
2×105 cells/ml, the cells were inoculated in the
Transwell chambers of a 24-well plate. After each chamber was added
with about 200 µl of the cell suspension, the lower chamber in the
plate was added with 600 µl of DMEM culture solution containing 10%
FBS, and then the cells were cultured for 24 h in an incubator at
37°C. The supernatant was removed with cotton swabs and the chamber
was washed with PBS. The cells in the lower chamber were fixed with
95% ethanol solution for 30 min, and then the chamber was washed
again with PBS. Each well was added with about 600 µl of 0.1%
crystal violet for staining. The number of migrated cells in 5
random wells was calculated with a microscope, to calculate the
average value. The experiment was carried out 3 times.
Apoptosis
Annexin V-FITC/PI double staining combined with flow
cytometry was used to detect apoptosis. The transfected cells were
inoculated in a 6-well plate at 3×105 cells/well,
digested with 0.25% trypsin, and then washed twice with PBS. The
cells were added with 100 µl of binding buffer to prepare into a
cell suspension at 1×106 cells/ml. The cells were
sequentially added with Annexin V-FITC and PI, and then incubated
at room temperature in the dark for 5 min. Finally, a flow
cytometer was used to detect apoptosis and calculate the average
value. The experiment was carried out 3 times.
Western blotting
RIPA lysis method was used to extract the total
protein from the collected cells, and BCA protein assay was used to
detect its concentration. The protein was separated by
electrophoresis with 12% SDS-PAGE, transferred to PVDF membrane,
and then sealed with 5% skimmed milk powder at room temperature for
2 h. After that, the membrane was added with mouse monoclonal
antibodies [Smad3 (1:500), Bax (1:500), Bcl-2 (1:500), and β-actin
(1:1,000)], and sealed overnight at 4°C. After washing to remove
the primary antibodies, the membrane was added with goat
anti-rabbit (1:3,000), incubated at 37°C for 1 h, and rinsed with
PBS 3 times. Finally, the excess liquid was absorbed dry, and the
protein was luminesced and developed with ECL.
Dual luciferase reporter gene
assay
Dual luciferase reporter gene assay was performed to
determine whether Smad3 was the direct target gene of miR-129-3p.
Smad3 3′UTR dual luciferase reporter plasmids (WT and MUT) that
were chemically synthetized by XbaI/XbaI digestion
were, respectively, co-transfected into PC-3 cells with
miR-129-3p-mimics or miR-NC using Lipofectamine 2000. After 48-h
incubation, the luciferase activity was detected by the dual
luciferase reporter system.
Statistical methods
SPSS 19.0 was used for statistical analysis.
GraphPad Prism 6 was used to plot figures. Measurement data were
expressed as mean ± standard deviation (means ± SD) and analyzed by
t-test. Comparison between groups was analyzed by independent
samples t-test and represented by t-test. Comparison between
multiple groups was analyzed by one-way analysis of variance, and
post hoc pairwise comparison was analyzed by LSD t-test. Pearson
was used for correlation analysis. P<0.05 was considered to
indicate a statistically significant difference.
Results
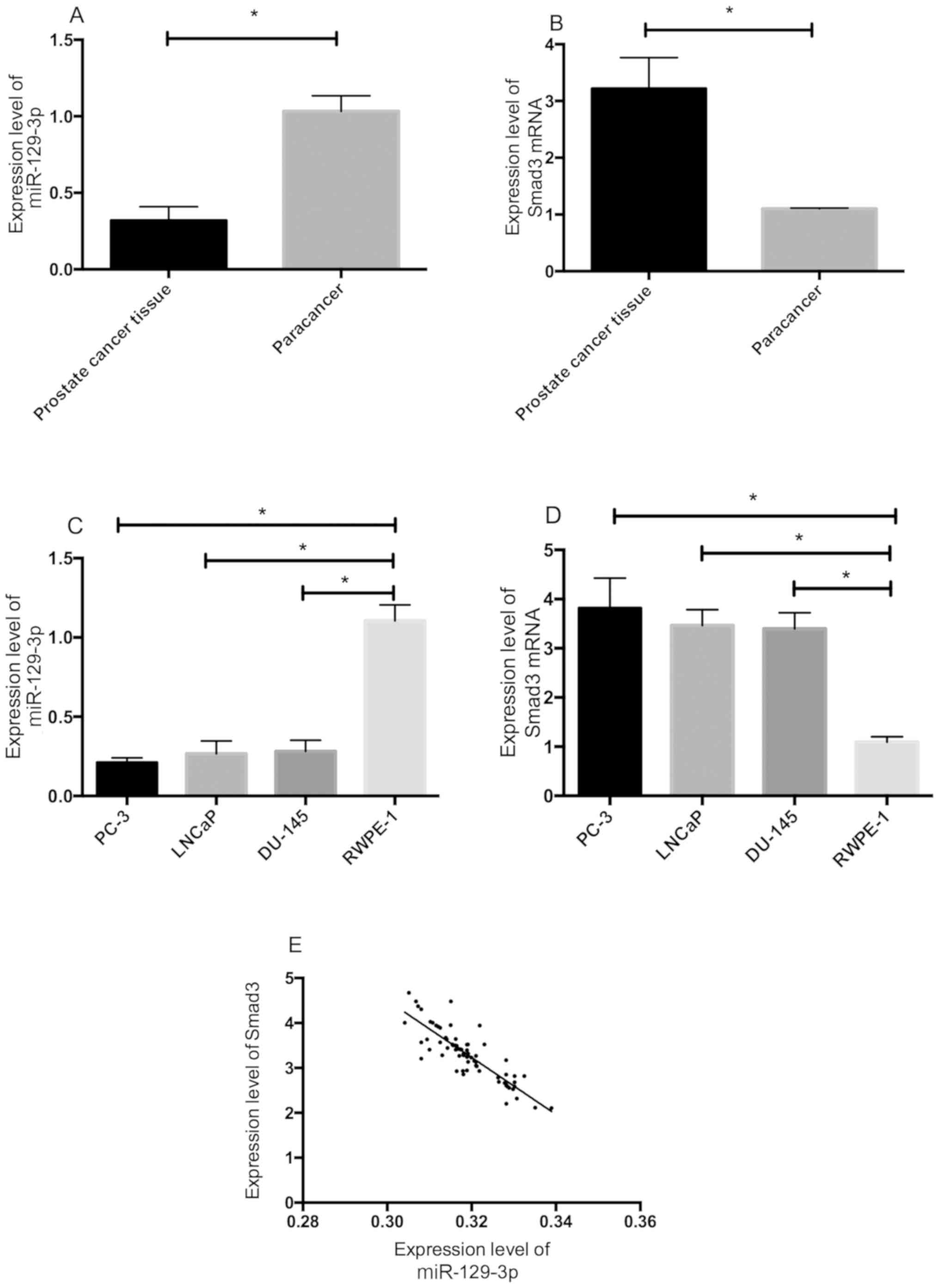
Expression levels of miR-129-3P and
Smad3 and their correlation
The expression levels of miR-129-3p and Smad3 mRNA
in prostate cancer and adjacent tissues were detected by qRT-PCR.
Compared with those in the adjacent tissue, the expression level of
miR-129-3p in the prostate cancer tissue significantly decreased
(P<0.05), while the expression level of Smad3 significantly
increased (P<0.05). The expression levels of miR-129-3p and
Smad3 protein in PC-3, DU-145, LNCaP, and RWPE-1 cells were
detected. The expression level of miR-129-3p in PC-3, DU-145, and
LNCaP cells was lower than that in RWPE-1 cells (P<0.05), while
the expression level of Smad3 was higher than that in RWPE-1 cells
(P<0.05). The expression levels of miR-129-3p and Smad3 protein
were negatively correlated (r=−0.855, P<0.001). According to the
analysis of the correlations of miR-129-3p and Smad3 with the
clinicopathological features, the expression levels of miR-129-3p
and Smad3 were closely related to the degree of tumor
differentiation, TNM staging, and lymph node metastasis (P<0.05)
(Table II and Fig. 1).
| Table II.Correlation of miR-129-3p and Smad3
with clinicopathological features. |
Table II.
Correlation of miR-129-3p and Smad3
with clinicopathological features.
| Factors | Relative expression
of miR-129-3p | t/F-value | P-value | Relative expression
of Smad3 | t/F-value | P-value |
|---|
| Age |
| 0.953 | 0.344 |
| 0.391 | 0.697 |
| <58
years (n=36) | 0.51±0.05 |
|
| 2.12±0.34 |
|
|
| ≥58 years (n=38) | 0.52±0.04 |
|
| 2.15±0.32 |
|
|
| TNM staging |
| 17.43 | <0.001 |
| 15.93 | <0.001 |
| I and II
(n=47) | 0.69±0.07 |
|
| 1.69±0.21 |
|
|
| III
(n=22) | 0.41±0.04 |
|
| 2.95±0.45 |
|
|
| Pathological
types |
| 0.838 | 0.437 |
| 0.292 | 0.747 |
|
Adenocarcinoma (n=25) | 0.46±0.05 |
|
| 2.09±0.32 |
|
|
| Squamous
cell carcinoma (n=27) | 0.47±0.06 |
|
| 2.16±0.34 |
|
|
|
Adenosquamous carcinoma
(n=22) | 0.45±0.05 |
|
| 2.13±0.33 |
|
|
| Lymph node
metastasis |
| 18.63 | <0.001 |
| 13.46 | <0.001 |
| Yes
(n=29) | 0.40±0.05 |
|
| 3.02±0.48 |
|
|
| No
(n=45) | 0.71±0.08 |
|
| 1.72±0.35 |
|
|
| Degree of
differentiation |
| 14.95 | <0.001 |
| 10.26 | <0.001 |
| Low
(n=20) | 0.40±0.04 |
|
| 2.86±0.62 |
|
|
|
Moderate and high (n=49) | 0.62±0.06 |
|
| 1.68±0.31 |
|
|
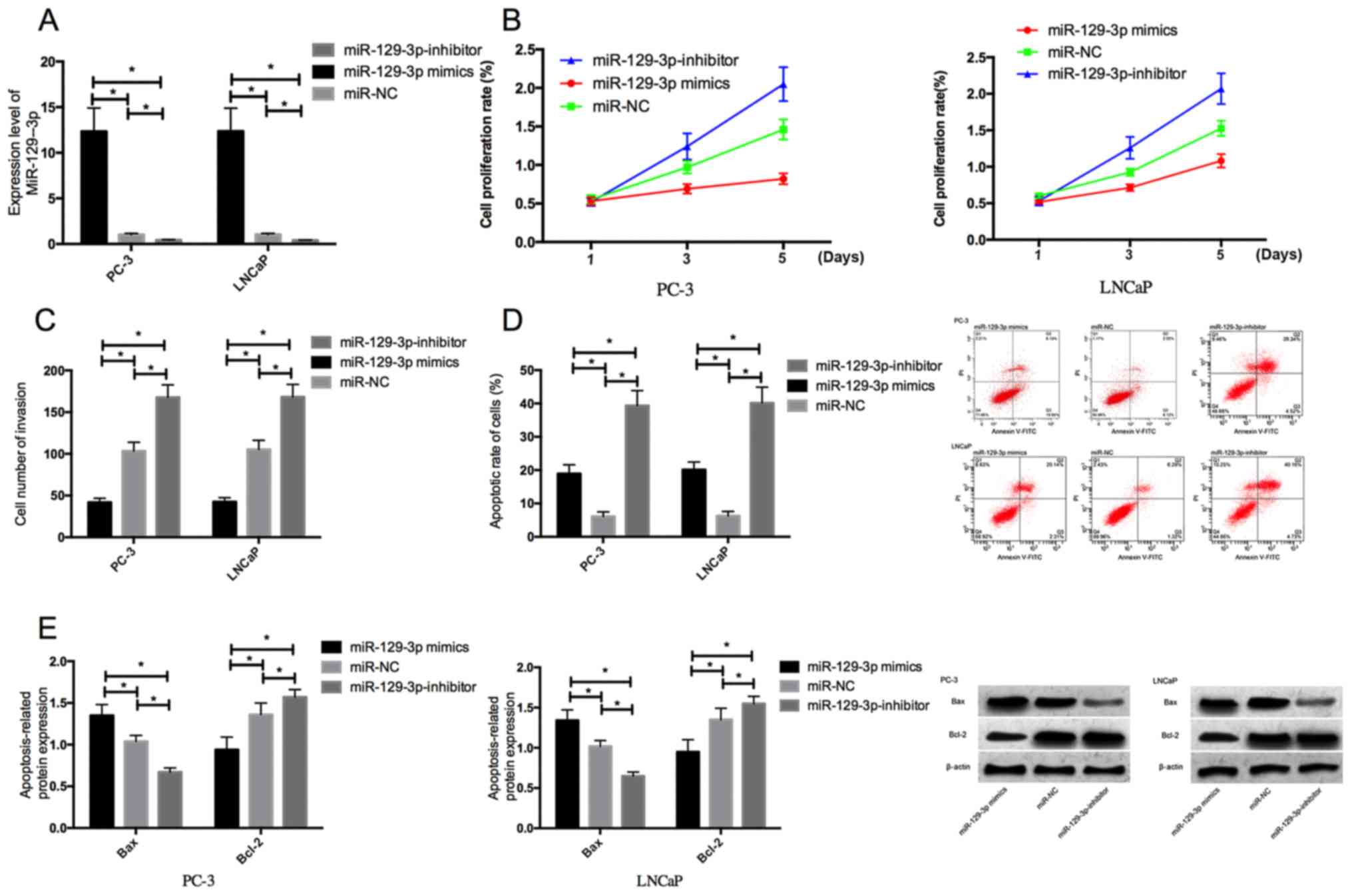
Effects of miR-129-3p on biological
functions of prostate cancer cells
PC-3 and LNCaP cells were transfected with
miR-129-3p mimics, miR-129-3p inhibitor, or miR-NC for 48 h, to
explore the effects of miR-129-3p overexpression on the
proliferation, invasion, and apoptosis of prostate cancer cells.
Compared with those transfected with miR-NC, the expression level
of miR-129-3p in the PC-3 and LNCaP cells transfected with
miR-129-3p-mimics significantly increased, while the expression
level in those transfected with miR-129-3p-inhibitor significantly
decreased (Fig. 2A and B,
P<0.05). After transfection, MTT assay, cell invasion detection,
and flow cytometry were conducted. The overexpression of miR-129-3p
significantly inhibited the proliferation and invasion of the cells
and promoted apoptosis (Fig. 2C-E,
P<0.05). According to the results of western blotting, the
overexpression increased the expression level of pro-apoptotic
protein Bax while decreased the expression level of anti-apoptotic
protein Bcl-2. However, the inhibition of miR-129-3p expression
significantly promoted the proliferation and invasion of the cells,
inhibited apoptosis, and decreased the expression level of
pro-apoptotic protein Bax, as well as increased the expression
level of anti-apoptotic protein Bcl-2 (Fig. 2).
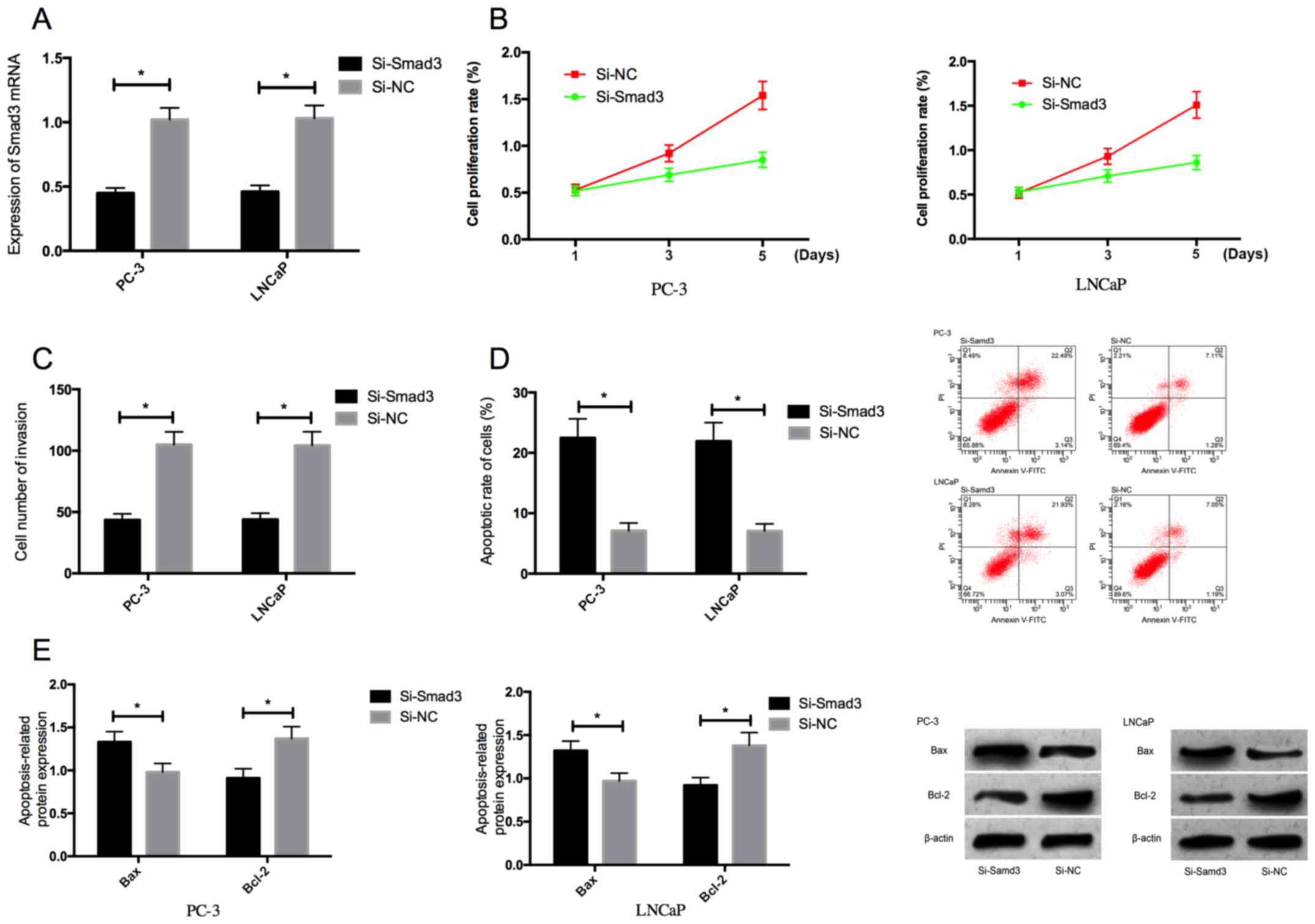
Effects of Smad3 on biological
functions of prostate cancer cells
PC-3 and LNCaP cells were transfected with Si-Smad3
or Si-NC for 48 h, to explore the effects of Smad3 on the
proliferation, invasion and apoptosis of prostate cancer cells.
Compared with those transfected with Si-NC, the expression level of
Smad3 in the PC-3 and LNCaP cells transfected with Si-Smad3
significantly decreased (P<0.05). After transfection, MTT assay,
cell invasion detection, and flow cytometry were conducted. The
proliferation and invasion of the cells transfected with Si-Smad3
were significantly inhibited while the apoptotic rate significantly
increased. According to the results of western blotting, the
expression level of pro-apoptotic protein Bax in the cells
transfected with Si-Smad3 increased, while the expression level of
anti-apoptotic protein Bcl-2 decreased (Fig. 3).
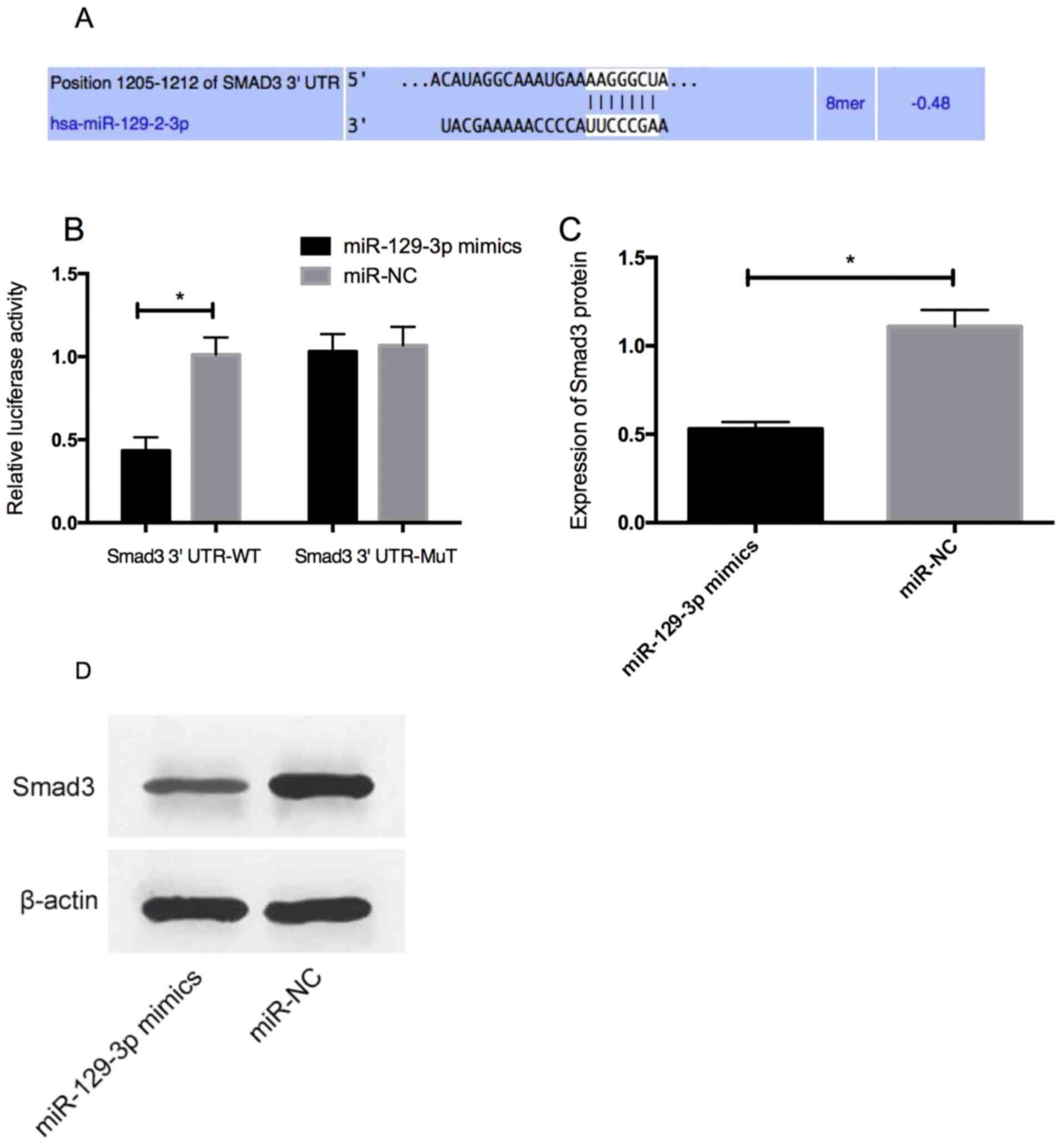
Smad3 as the direct target gene of
miR-129-3p
Bioinformatics analysis was conducted to predict the
target gene of miR-129-3p, to explore the potential mechanism of
miR-129-3p in prostate cancer. Smad3 was identified as the target
gene of miR-129-3p. Dual luciferase reporter gene assay was carried
out to determine whether Smad3 3′UTR could be directly targeted by
miR-129-3p. The overexpression of miR-129-3p decreased the
luciferase activity of position 1,205-1,212 of Smad3 3′UTR Wt
(P<0.05), without effect on position 1,205-1,212 of Smad3 3′UTR
Mut. According to the results of western blotting, the expression
level of Smad3 protein in PC-3 cells transfected with
miR-129-3p-mimics decreased (P<0.05) (Fig. 4).
Discussion
miRNA is a non-coding RNA closely related to the
development and progression of tumors, and its role in prostate
cancer has been widely discussed in recent years (11). There are various miRNA targets that
can be used for the diagnosis and treatment of prostate cancer,
with more tumor suppressor genes and fewer oncogenes (12,13). A
study has shown that the upregulation of miR-101 inhibits the
proliferation and invasion of prostate cancer cell line DU145 by
inhibiting its target gene EZH2 (14). miR-129 is a newly discovered miRNA
that is closely associated with the pathogenesis of prostate
cancer. According to previous studies, it regulates the growth
hormone receptor (GHR) of target genes in LNCaP cells, thereby
inhibiting the proliferation of prostate cancer cells (15). It also controls the centrosome number
of metastatic prostate cancer cells by inhibiting CP110, thus
affecting prostate cancer (16).
In this study, bioinformatics prediction showed that
Smad3 was the direct target gene of miR-129-3p. A previous study
reported that Smad3 promotes epithelial-mesenchymal transition of
prostate cancer cells and functions as an oncogene in the cells
(17). Therefore, we speculated that
miR-129-3p could play a role in prostate cancer cells by regulating
Smad3. According to the results of qRT-PCR, the expression level of
miR-129-3p in the prostate cancer tissue and in PC-3, DU-145, and
LNCaP cells significantly increased, while that of Smad3 mRNA
significantly decreased. In the present study, bioinformatics
prediction showed the target relationship between miR-129-3p and
Smad3. According to the results of the dual luciferase reporter
gene assay, the overexpression of miR-129-3p reduced the luciferase
activity of position 1,205-1,212 of Smad3 3′UTR Wt, without effect
on position 1,205-1,212 of Smad3 3′UTR Mut. This suggests that
Smad3 may be a direct target gene of miR-129-3p. As a
multifunctional cytokine, transforming growth factor-β (TGF-β)
plays a certain role in cells in autocrine and paracrine manner.
Smad3 is an important member of the Smad protein family and a major
component of TGF-β/Smad signaling pathway (18,19). We
regulated the expression level of Smad3 and found that the
downregulation of the expression level significantly inhibited the
proliferation and invasion of prostate cancer cells, but
significantly increased the apoptotic rate of the cells. A study
has shown that the high expression of Smad3 is closely correlated
with the recurrence and prognosis of prostate cancer (20). At diagnosis, prostate cancer has
mostly progressed into castration-resistant prostate cancer
(21), in which Smad3 promotes the
expression of prostate specific antigens and finally the growth of
prostate cancer cells through the auxiliary activation of androgen
receptors (22,23). These studies support our
conclusions.
In order to further confirm the effects of
miR-129-3p on the biological functions of prostate cancer cells,
miR-129-3p in PC-3 cells was overexpressed through plasmid
transfection. The expression level of Smad3 and the proliferation,
invasion, and apoptosis of PC-3 cells after the overexpression were
also detected. The results showed that the overexpression of
miR-129-3p significantly inhibited the proliferation and invasion
of PC-3 cells, promoted apoptosis, and significantly decreased the
expression level of Smad3 protein. These findings indicate that
miR-129-3p may function as a tumor suppressor gene by inhibiting
the expression level of Smad3 protein. Apoptosis detection showed
that the overexpression of miR-129-3p increased the expression
level of pro-apoptotic protein Bax but decreased the expression
level of anti-apoptotic protein Bcl-2, which shows that the
pro-apoptotic function of miR-129-3p may be related to the
regulation of apoptosis-related proteins. Based on previous
studies, miR-129-3p regulates the expression level of oncogene WWP1
and then inhibits the proliferation and metastasis of gastric
cancer cells (24). It can also
inhibit the growth of glioblastoma cells through the targeted
regulation of E2F5 gene (25).
Based on our study miR-129-3p may affect the
biological functions of prostate cancer cells through the targeted
regulation of Smad3. However, it remains unclear how miR-129-3p
regulates other related targets and whether these targets have a
synergistic effect with Smad3, which needs further
investigation.
In summary, miR-129-3p expression is low in prostate
cancer tissue, and inhibits the proliferation and migration of the
cells and induces apoptosis by negatively regulating the expression
level of Smad3, so it may be a potential target for the diagnosis
and treatment of prostate cancer.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YJ designed the study and wrote the manuscript. YG
was responsible for all the experiments. JD helped with statistical
analysis. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
Gansu Provincial Hospital of TCM (Lanzhou, China). Patients who
participated in this research had complete clinical data. Signed
informed consents were obtained from the patients or the
guardians.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Fizazi K, Tran N, Fein L, Matsubara N,
Rodriguez-Antolin A, Alekseev BY, Özgüroğlu M, Ye D, Feyerabend S,
Protheroe A, et al: Abiraterone plus Prednisone in metastatic,
castration-sensitive prostate cancer. Lancet Oncol. 20:686–700.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gillessen S, Attard G, Beer TM, Beltran H,
Bossi A, Bristow R, Carver B, Castellano D, Chung BH, Clarke N, et
al: Management of patients with advanced prostate cancer: The
report of the Advanced Prostate Cancer Consensus Conference APCCC
2017. Eur Urol. 73:178–211. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Madaan S, Abel PD, Chaudhary KS, Hewitt R,
Stott MA, Stamp GW and Lalani EN: Cytoplasmic induction and
over-expression of cyclooxygenase-2 in human prostate cancer:
Implications for prevention and treatment. BJU Int. 86:736–741.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hao NB, He YF, Li XQ, Wang K and Wang RL:
The role of miRNA and lncRNA in gastric cancer. Oncotarget.
8:81572–81582. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fang H, Xie J, Zhang M, Zhao Z, Wan Y and
Yao Y: miRNA-21 promotes proliferation and invasion of
triple-negative breast cancer cells through targeting PTEN. Am J
Transl Res. 9:953–961. 2017.PubMed/NCBI
|
|
6
|
Liu K, Huang J, Ni J, Song D, Ding M, Wang
J, Huang X and Li W: MALAT1 promotes osteosarcoma development by
regulation of HMGB1 via miR-142-3p and miR-129-5p. Cell Cycle.
16:578–587. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lin VC, Huang SP, Ting HJ, Ma WL, Yu CC,
Huang CY, Yin HL, Huang TY, Lee CH, Chang TY, et al: Vitamin D
receptor-binding site variants affect prostate cancer progression.
Oncotarget. 8:74119–74128. 2017.PubMed/NCBI
|
|
8
|
Lu C, Shan Z, Li C and Yang L: miR-129
regulates cisplatin- resistance in human gastric cancer cells by
targeting P-gp. Biomed Pharmacother. 86:450–456. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zuo Y, Li Y, Zhou Z, Ma M and Fu K: Long
non-coding RNA MALAT1 promotes proliferation and invasion via
targeting miR-129-5p in triple-negative breast cancer. Biomed
Pharmacother. 95:922–928. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tang PM, Zhou S, Meng XM, Wang QM, Li CJ,
Lian GY, Huang XR, Tang YJ, Guan XY, Yan BP, et al: Smad3 promotes
cancer progression by inhibiting E4BP4-mediated NK cell
development. Nat Commun. 8:146772017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pashaei E, Pashaei E, Ahmady M, Ozen M and
Aydin N: Meta-analysis of miRNA expression profiles for prostate
cancer recurrence following radical prostatectomy. PLoS One.
12:e01795432017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lu S, Wang MS, Chen PJ, Ren Q and Bai P:
miRNA-186 inhibits prostate cancer cell proliferation and tumor
growth by targeting YY1 and CDK6. Exp Ther Med. 13:3309–3314. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu C, Liu R, Zhang D, Deng Q, Liu B, Chao
HP, Rycaj K, Takata Y, Lin K, Lu Y, et al: MicroRNA-141 suppresses
prostate cancer stem cells and metastasis by targeting a cohort of
pro-metastasis genes. Nat Commun. 8:142702017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Varambally S, Cao Q, Mani RS, Shankar S,
Wang X, Ateeq B, Laxman B, Cao X, Jing X, Ramnarayanan K, et al:
Genomic loss of microRNA-101 leads to overexpression of histone
methyltransferase EZH2 in cancer. Science. 322:1695–1699. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Elzein S and Goodyer CG: Regulation of
human growth hormone receptor expression by microRNAs. Mol
Endocrinol. 28:1448–1459. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bijnsdorp IV, Hodzic J, Lagerweij T,
Westerman B, Krijgsman O, Broeke J, Verweij F, Nilsson RJ,
Rozendaal L, van Beusechem VW, et al: miR-129-3p controls
centrosome number in metastatic prostate cancer cells by repressing
CP110. Oncotarget. 7:16676–16687. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huang S, Liao Q, Li L and Xin D: PTTG1
inhibits SMAD3 in prostate cancer cells to promote their
proliferation. Tumour Biol. 35:6265–6270. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Majumder S, Bhowal A, Basu S, Mukherjee P,
Chatterji U and Sengupta S: Deregulated E2F5/p38/SMAD3 circuitry
reinforces the pro-tumorigenic switch of TGFβ signaling in prostate
cancer. J Cell Physiol. 231:2482–2492. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yao B, Zhao J, Li Y, Li H, Hu Z, Pan P,
Zhang Y, Du E, Liu R and Xu Y: Elf5 inhibits TGF-β-driven
epithelial-mesenchymal transition in prostate cancer by repressing
SMAD3 activation. Prostate. 75:872–882. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lu S, Lee J, Revelo M, Wang X, Lu S and
Dong Z: Smad3 is overexpressed in advanced human prostate cancer
and necessary for progressive growth of prostate cancer cells in
nude mice. Clin Cancer Res. 13:5692–5702. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Virgo KS, Rumble RB and Singer EA:
Second-line hormonal therapy for men with chemotherapy-naïve
castration-resistant prostate cancer: American Society of Clinical
Oncology Provisional Clinical Opinion Summary. J Oncol Pract.
13:459–461. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Teixeira AL, Gomes M, Nogueira A, Azevedo
AS, Assis J, Dias F, Santos JI, Lobo F, Morais A, Maurício J, et
al: Improvement of a predictive model of castration-resistant
prostate cancer: Functional genetic variants in TGFβ1 signaling
pathway modulation. PLoS One. 8:e72419. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Song K, Wang H, Krebs TL, Wang B, Kelley
TJ and Danielpour D: DHT selectively reverses
Smad3-mediated/TGF-beta-induced responses through transcriptional
down-regulation of Smad3 in prostate epithelial cells. Mol
Endocrinol. 24:2019–2029. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ma L, Chen X, Li C, Cheng R, Gao Z, Meng
X, Sun C, Liang C and Liu Y: miR-129-5p and −3p co-target WWP1 to
suppress gastric cancer proliferation and migration. J Cell
Biochem. Nov 11–2018.(Epub ahead of print). doi:
10.1002/jcb.28027.
|
|
25
|
Fang D-Z, Wang Y-P, Liu J, Hui XB, Wang
XD, Chen X and Liu D: MicroRNA-129-3p suppresses tumor growth by
targeting E2F5 in glioblastoma. Eur Rev Med Pharmacol Sci.
22:1044–1050. 2018.PubMed/NCBI
|