Introduction
Hepatocellular carcinoma (HCC) is a malignant tumor
with high incidence and the number of novel cases is expected to
increase by 1 million every year in the next decade (1). The main causative factors include
alcoholic liver disease, hepatitis B and hepatitis C viral
infections, and non-alcoholic fatty liver disease (2). However, the pathogenesis and exact
molecular mechanism of HCC are not fully understood. Although
international guidelines suggest HCC screening for patients with
cirrhosis, regular monitoring presents various limitations in
clinical practice (3). The large
number of undiagnosed patients leads to a low HCC monitoring rate
and high late diagnosis rate, causing the patients with HCC to be
diagnosed only when the tumor exhibits a large size, leading to
poor prognosis (4).
During tumor development and progression, multiple
cell types interact with tumor cells, including astrocytes, B
cells, lymphocytes, macrophages, monocytes, natural killer cells
and T cells, constituting the tumor immune microenvironment
(5,6). These cells, together with the
fibroblasts, vascular endothelial cells and other factors, which
are collectively called the tumor stroma, as well as the
extracellular matrix, oxygen levels and pH values, constitute the
tumor microenvironment (7,8). Notably, the interactions between immune
cells and tumor cells affect the growth and remodeling of the tumor
microenvironment (9). Immune cells
can stimulate tumor cells to secrete cytokines, which mediate the
tumor growth by promoting the growth of new blood and lymphatic
vessels (6). Different cell types
may serve pro- and anti-tumorigenic roles in the tumor
microenvironment. For example, targeting T cell activation is
considered as an important novel strategy to repress tumor growth
(10,11). S100A4 has been shown to be an
oncogene able to promote inflammation (12) and affect angiogenesis (13). Accumulating evidence showed that
expression of S100A4 in tumor cells is related to the
tumor-associated T cell deficiency (14). The rich blood supply and unique
sinusoid structure of the liver provide a plastic environment for
the formation and function of the tumor immune microenvironment
(15). Therefore, it is of great
significance to study the molecular characteristics and
intercellular interactions in the HCC immune microenvironment.
Massively parallel sequencing data have provided
novel insights in the field of cancer research. In particular,
RNA-sequencing (RNA-seq) has been used to detect genomic mutations
and rearrangement signatures in the human genome and transcriptome.
However, conventional bulk RNA-seq can only provide the average
expression signal of transcripts in the whole tumor tissue, without
considering the tumor heterogeneity. By contrast, single-cell
RNA-seq may facilitate the identification of complex and rare cells
populations, thus allowing investigation of the tumor immune
microenvironment (16), especially
in HCC. For example, using single-cell RNA-seq, a previous study
identified 11 HCC-related T cell subpopulations, which provided
valuable insights for the understanding of the cancer immune
microenvironment (17). In addition,
a previous study has described the molecular characteristics of
immune cells that infiltrate HCC to determine whether certain types
of drugs may be effective against liver cancer (18).
Moreover, chromatin immunoprecipitation (ChIP)-seq
results and protein-protein interaction (PPI) network may
facilitate the detection of gene regulatory networks and
interaction events, such as the bindings between transcription
factors (TFs) and promoters. The present study compared the
single-cell RNA-seq data of normal peripheral blood mononuclear
cells (PBMCs) with that of in vivo tumor cells and in
vitro cell lines using single cell classification and
identification. By integrating differential expression analysis,
ChIP-seq data and PPI networks, the present results suggested that
the JunB proto-oncogene (JUNB) may serve an important role
in the development and progression of HCC and the immune response.
In addition, apolipoprotein A2 (APOA2), which encodes a
genetically susceptible protein in HCC (19), was found to exhibit the same
expression pattern as JUNB. The present results may
contribute to the identification of novel therapeutic targets for
the treatment of HCC patients.
Materials and methods
Data collection
In vivo tumor cells were isolated from a
patient who had undergone resection at the National Institutes of
Health (NIH) Clinical Center. The tissue acquisition procedures
were approved by the Institutional Review Board of The NIH
(20). In total, two in vitro
cell lines (HuH1 and HuH7) from The Health Science Research
Resources Bank (cat. nos. JCRB0199 and JCRB0403) were pooled and
used for 10× Genomics single-cell RNA-seq. These data were
collected and downloaded from the GEO database (database no.
GSE103867) (20). Single-cell data
of PBMCs were downloaded from the GEO database (database no.
GSE111360) (21). Gene expression
profile and clinical information of a cohort of 360 patients with
HCC were collected from The Cancer Genome Atlas (TCGA; http://cancergenome.nih.gov/). The PPI network was
obtained using STRING (v11.0) with only highly strong interactions
(score, 0.4) being used (22).
Preprocessing for 10× Genomics
single-cell RNA-seq data
Seurat v2.1 (http://satijalab.org/seurat/) was used to analyze the
10× Genomics data (23). Genes whose
expression was detected in ≥3 cells and cells with ≥10 genes were
used in this study. Variable genes were identified using cutoffs
(x.low.cutoff = 0.05; y.cutoff = 0.1). The top 20 principal
components were used in the clustering analysis (resolution = 0.6).
Gene expression levels were quantified using the unique molecular
identifier counts. Dimensionality reduction was based on the t-SNE
algorithm. Subsequently, cell populations were clustered by
principal component analysis.
Differential expression gene and
pathway enrichment analysis
Log2Fold-Change represented the ratio of
gene expression between one cluster of cells and all the other
cells. P-values were calculated using the negative binomial test
and adjusted by the Benjamini-Hochberg method. Gene Ontology (GO)
terms and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway
enrichment analyses were performed using DAVID (version 6.8)
(24).
Correlation analysis
Gene expression of 360 patients with HCC, obtained
from the TCGA database, was used as an independent external test
set to validate the putative genes of interest. Co-expression
analysis based on Spearman's Correlation was performed using
cBioPortal (http://www.cbioportal.org/) (25).
Survival analysis
Data from patients with HCC derived from TCGA were
divided into two groups according to the expression of APOA2
(high or low, with an mRNA Z-score >0 or <0, respectively)
and the expression of JUNB (high or low, with an mRNA
Z-score >0 or <0, respectively). Survival curves were
estimated by the Kaplan-Meier method and compared with the log-rank
test.
Transcriptional regulation
analysis
For each gene, interactions between proteins and
their promoters and enhancers were obtained from r GeneHancer
(26), a database of ChIP-seq data
classify by inferred target genes. The interactions between TFs and
their binding sites in the promoter and enhancer regions were
supported by ChIP-sequencing. Subsequently, the TFs predicted to
regulate both JUNB and APOA2 were used.
Results
Identification of significant
differences in gene expression between in vivo and in vitro
cells
Differential expression analysis was performed
between the transcriptomes of in vivo tumor cells and two
in vitro cell lines (Fig.
S1). Top 2 cell population-specific marker gene expression is
presented in Fig. S2A. Top 9 cell
type-specific expressed genes are presented in Fig. S2B (HuH7 cells, Fig. S2Ba; P1T and P1C cells, Fig. S2Bb; P1B cells, Fig. S2Bc; and HuH1 cells; Fig. S2Bd). The specific expression was
potentially due to the invasion of immune cells in the tumor and
the immune response elicited by tumor cells (27). These significant differences
suggested that the validation of the RNA-seq data should be a
crucially important component for the in vitro analysis of
the immune response in tumors. Cell population identifications of
PBMCs are presented in Fig. S3A-D.
Subsequently, an integrated comparison among tumor cells, cell
lines and PBMCs was performed (Fig.
1). As expected, HuH1 and HuH7 cells clustered together in the
analysis (Fig. 1A). Interestingly,
the clustering analysis showed that a small number of epithelial
cells were observed among the PBMCs (Fig. 1A-C). According to previous studies
(28–30), these epithelial cells may be vascular
endothelial cells, which have the potential to regulate the tumor
cells. In addition to B cells and T cells, numerous types of
mononuclear cells were detected among the PBMCs (Fig. 1C). A heat map was used to observe the
expression of the differentially expressed genes (marker genes) in
the comparisons (Fig. 1D). The
expression of the marker genes in each group of single cells is
presented in Fig. 1E.
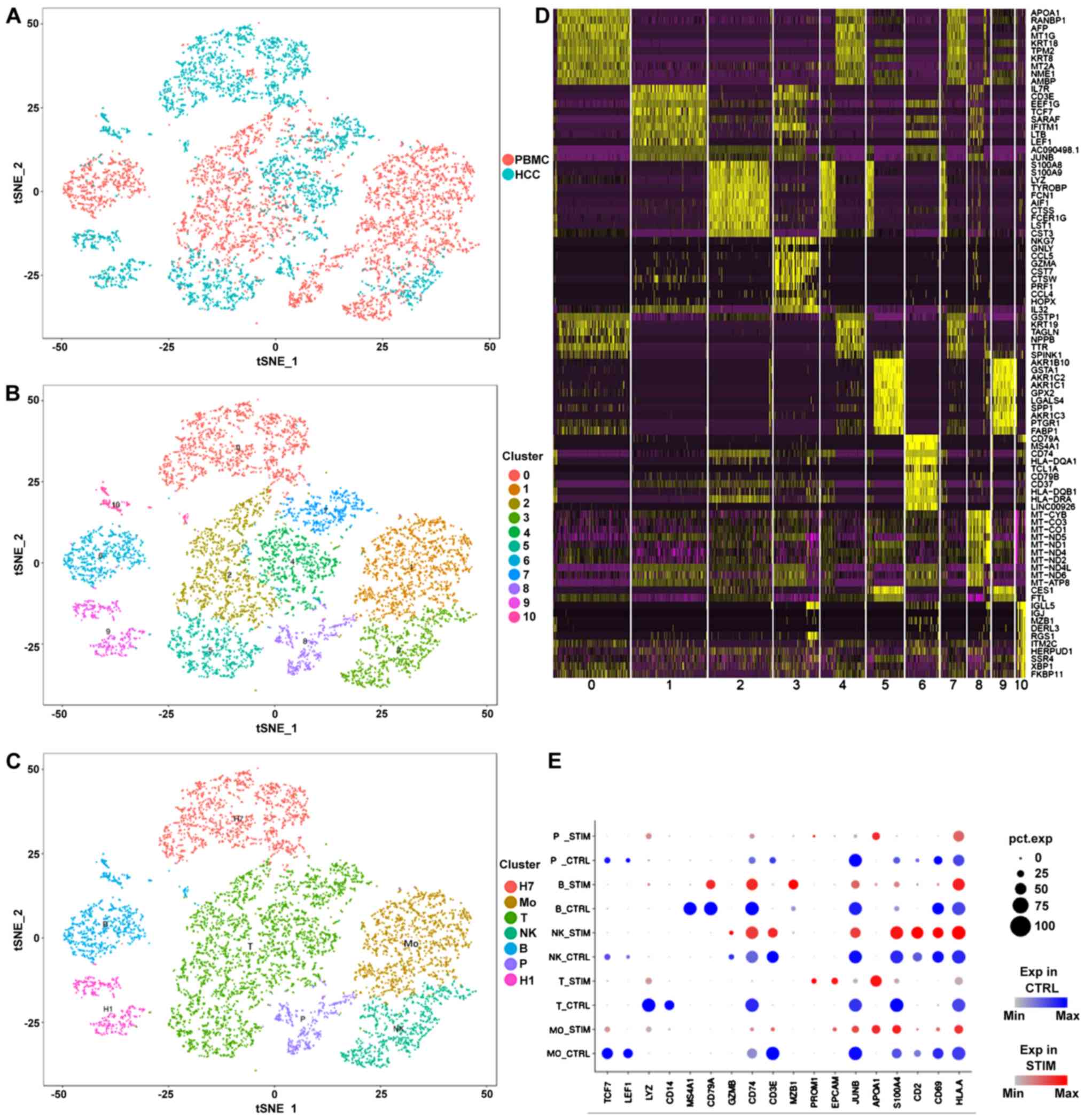 | Figure 1.Single-cell transcriptomic data of
PBMCs, HCC tumor cells and liver cancer cell lines integration. (A)
Integration of PMBC and tumor samples. (B) Identification of cell
populations by PCA. (C) Definition of cell types according to the
marker genes. (D) Heatmap of the markers identified by PCA. (E)
Comparison between expression of marker genes in PBMC and tumor
samples. The color blue indicates the expression levels in PBMC
samples, the color red in tumor samples. The size of each dot
represents the percentage of the cells that expressed the
corresponding gene. H1, HuH1; H7, HuH7; Mo, monocytes; NK, natural
killer cells; P, epithelial cells; T, T cells; B, B cells; PBMC,
peripheral blood mononuclear cells; PCA, principal component
analysis; STIM, tumor samples; CTRL, PBMC samples; HCC,
hepatocellular carcinoma. |
Tumor-infiltrating immune cells
exhibit a higher transcription level
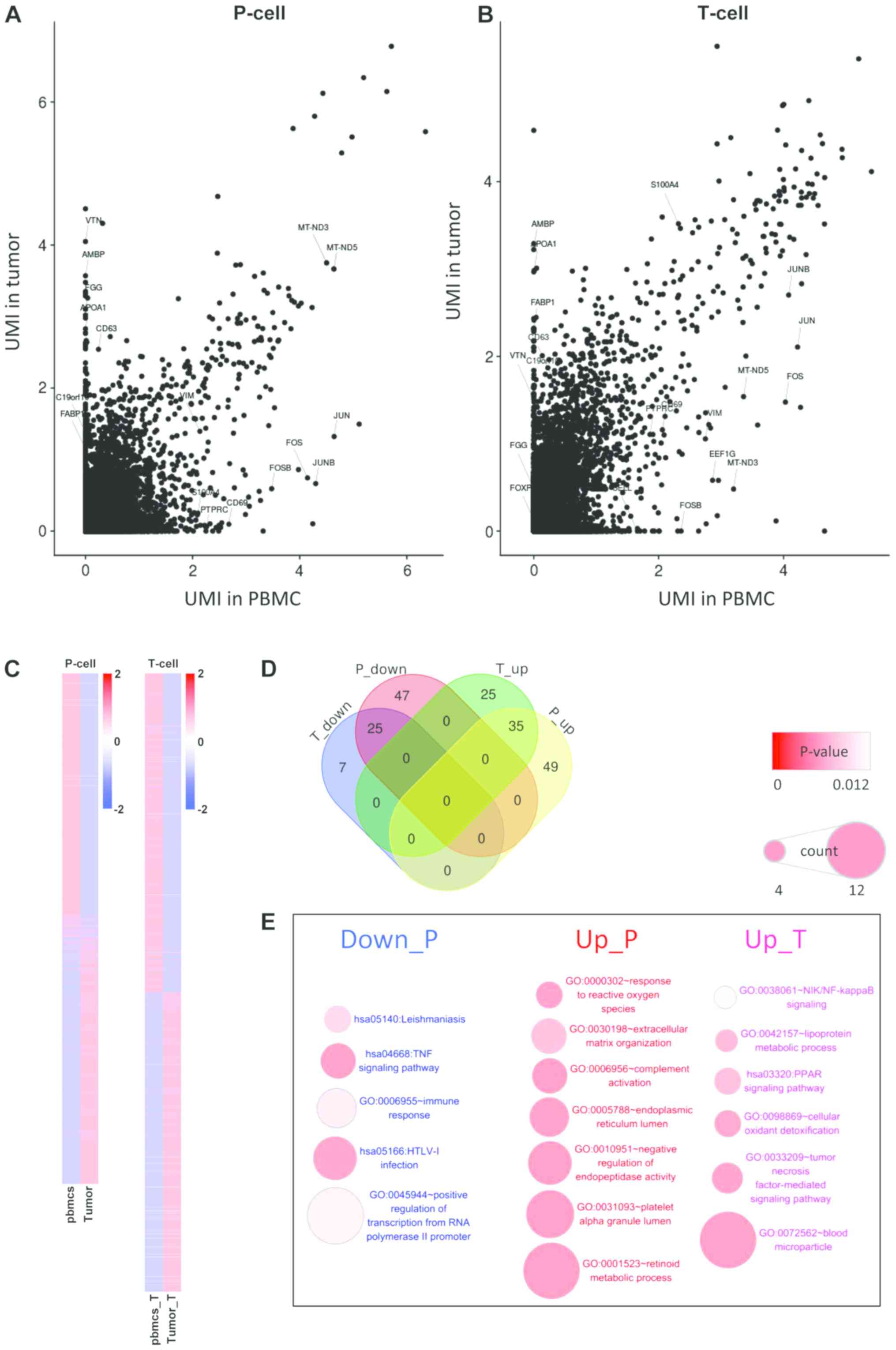
In the analysis of differentially expressed genes,
the gene expression levels between tumor cells and vascular
epithelial cells in peripheral blood (Fig. 2A) were compared. In addition, the
differentially expressed genes between T cells infiltrated in
tumors and T cells in peripheral blood were examined (Fig. 2B). In the two comparisons, the number
of the differentially expressed genes in T cells was significantly
decreased compared with those in epithelial cells, while most of
the differentially expressed genes in T cells were due to
differences among cell types (Fig. 2C
and D). Enrichment analysis of these differentially expressed
genes showed that the differentially expressed genes in epithelial
cells were significantly enriched in ‘inflammation’ and ‘immune
response’ (Fig. 2E). The inhibition
of some positive regulators of T cells, such as tumor necrosis
factor (TNF), NF-κB inhibitor α (NFKBIA), Fos
proto-oncogene (FOS), JUN and DEAD-box helicase 3
X-linked (DDX3X) may lead to the downregulation of the T
cell receptor signaling pathway, and the inhibition of the
hepatitis B and cellular oxidant detoxification pathways, which may
be caused by the abnormal growth of tumor cells and the
accumulation of stress-associated factors. In this condition, tumor
cells may require more energy to sustain their proliferation and to
adapt to the hypoxic micro-environment.
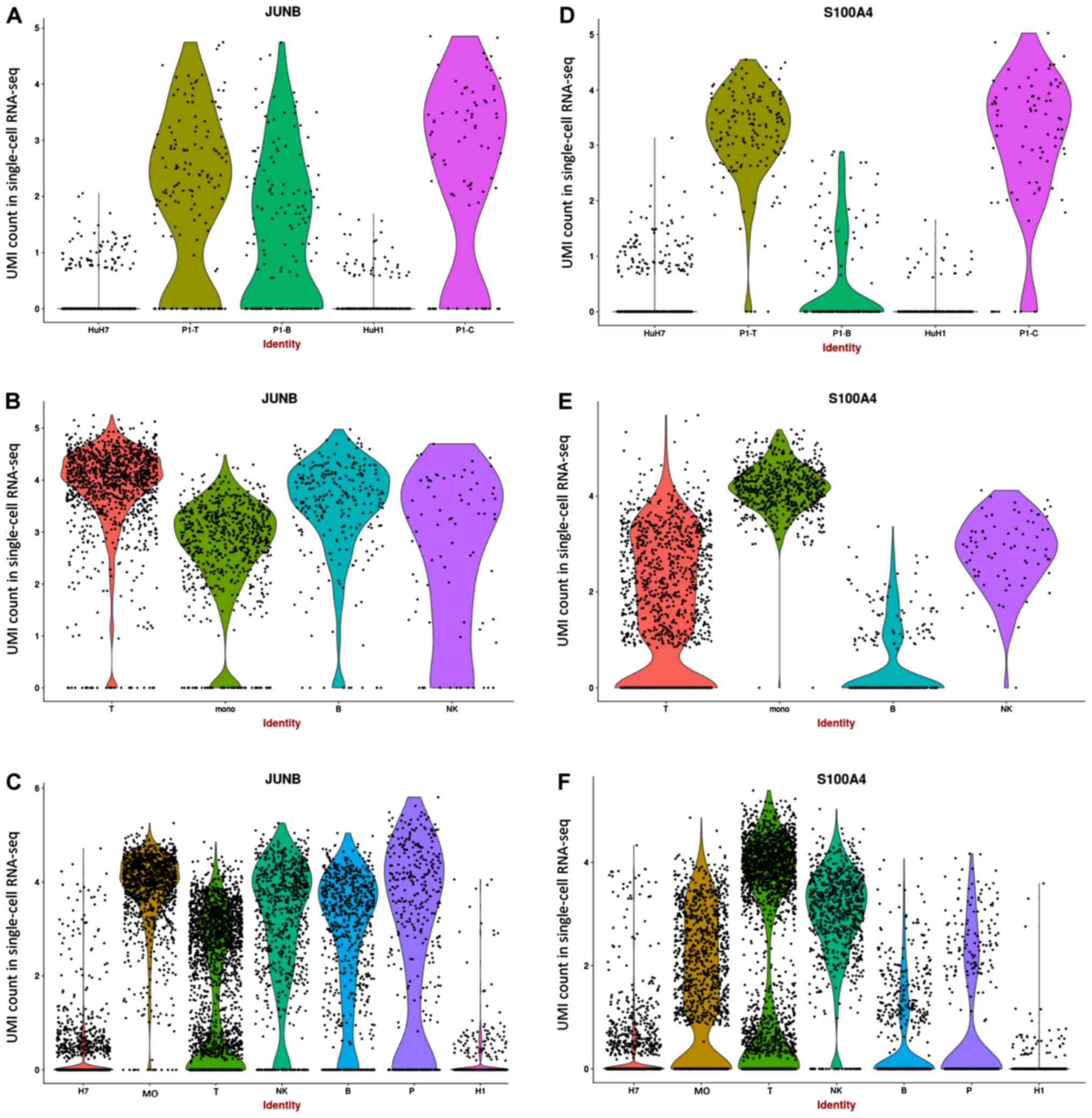
JUNB may serve a crucial role in HCC
tumor immune microenvironment
JUNB, which was reported as highly expressed
in cancer cells in previous studies (22,31–33), was
observed to be downregulated in numerous tumor cells, although it
was observed to be highly expressed in certain tumor cell lines
(Fig. 3A). Additionally, all PBMCs
presented high expression levels of JUNB (Fig. 3B). Since JUNB is associated
with lipid metabolism, high expression of JUNB was expected
in activated cells. S100A4, which is associated with the
development and progression of the tumor and immune infiltration
(34,35), exhibited high expression in in
vivo epithelial cells and T cells, but low expression in in
vitro cell lines (Fig. 3D and
E). Although the indirect interaction between JUNB and
S100A4 was found to function through annexin A2
(ANXA2; Fig. S3E), a
positive association between ANXA2, JUNB and S100A4
was detected in patients with HCC from the TCGA dataset (Fig. S3F). In addition, the functional
roles of ANXA2, JUNB and S100A4 in tumor cells were
found to be associated with T cells (14,36,37),
suggesting that the function of JUNB in HCC tumor cells may
be associated with the interaction between these genes in the tumor
immune microenvironment. By combining the data from PBMCs and tumor
samples, JUNB and S1004A were found to be decreased
in epithelial cells (Fig. 3C and F).
However, the expression level of S100A4 was found to be
increased in tumor-associated T cells, while the expression of
JUNB was decreased. JUNB was identified to be
downregulated in various tumors in the previous studies (31).
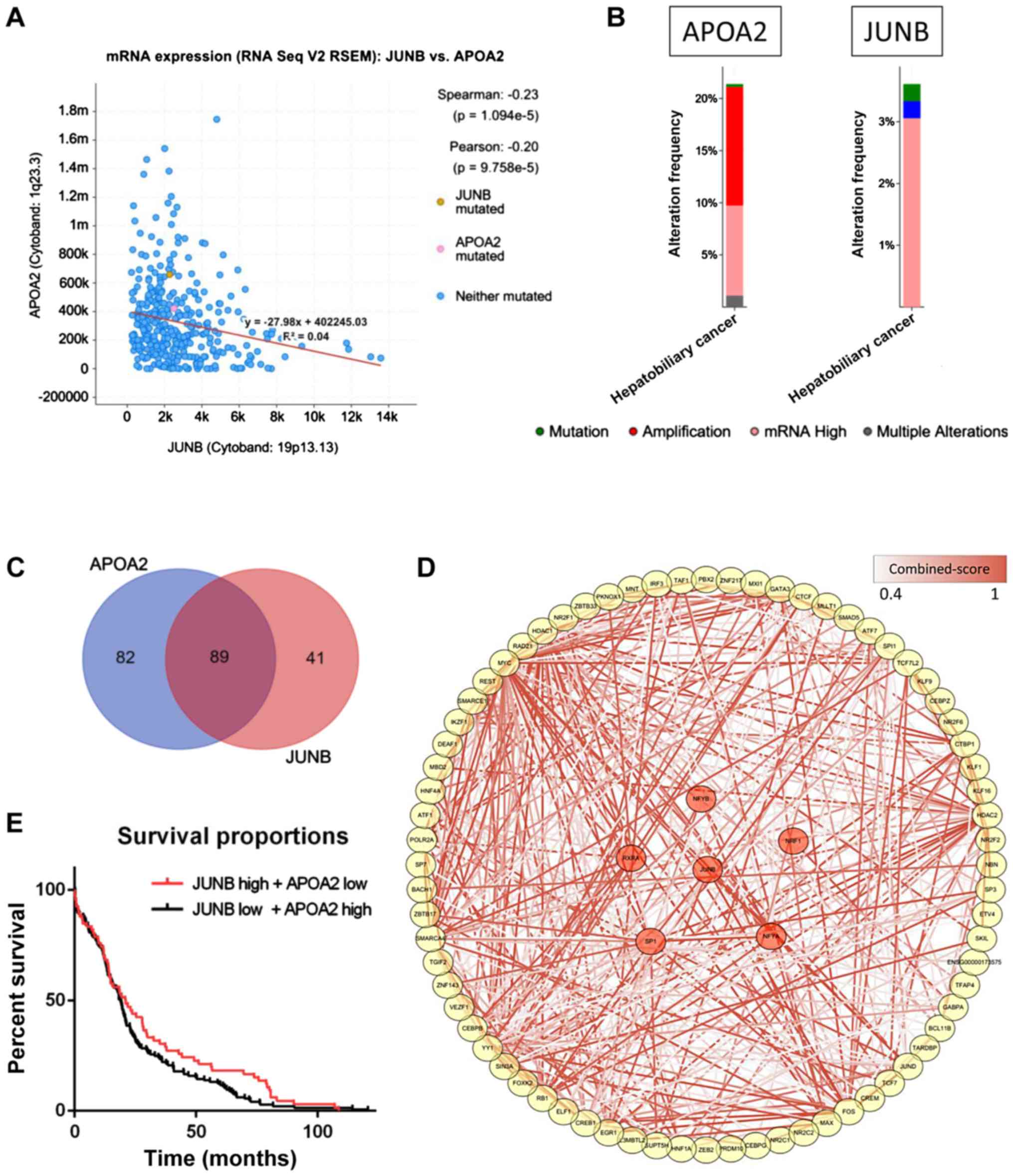
Validation of the key role of JUNB in
an independent dataset
The potential role of JUNB was further
investigated by integrating a cohort of 360 HCC patients from the
TCGA database. APOA2 was found to be significantly
negatively associated with JUNB in the dataset
(P=1.094×10−5; Fig. 4A).
Among the 360 HCC samples, >20% of the samples showed
APOA2 mRNA upregulation, which may be explained by the low
expression of JUNB in HCC samples (Fig. 4B). The high expression of JUNB
was previously predicted as a poor survival indicator in patients
with tumors (32,33), while APOA2 was found to serve
a role in the development and progression of the tumor through the
peroxisome proliferator activated receptor α (PPARα) pathway
(38). Moreover, >50% of the TFs
that interact with the promoters and enhancers of these two genes
were found to regulate both APOA2 and JUNB (Fig. 4C). Further data enrichment analysis
showed that the pathways associated with the TFs regulating
APOA2 and JUNB were involved in the development and
progression of HCC and immune response, which may represent a
potential novel mechanism underlying HCC (Fig. 4D), although further experimental
evidence is required to test this hypothesis. Although survival
analysis showed a slightly different result when analyzing the
overall survival time in two groups of patients (P=0.11), a
significant difference was observed when analyzing only longer
survival time (>15 months; Fig.
4E), indicating that JUNB and APOA2 may play a
key role in improving the survival time in patients with HCC.
Discussion
Increased understanding of tumor-host interactions
has accelerated the development of novel cancer immunotherapies. In
addition, drug resistance in biomarker therapy and immunotherapy
have recently been investigated (21). Despite their success,
immune-checkpoint inhibitors present certain limitations.
Therefore, in addition to improving the treatment of patients
presenting with tumors at an advanced stage, the identification of
driver biomarkers may contribute to immunotherapy in an early
clinical stage (39). Since
immune-checkpoint inhibitors could be used in HCC treatment,
combining molecular targeted therapy with immunotherapy has become
a therapeutic method to stimulate the immune response. The
previously described mechanisms underlying tumor development based
on cell lines studies have been found to be unreliable,
particularly for immune response and immunotherapy-related studies.
In order to investigate the role of the immune response in tumor
progression, the tumor microenvironment and the balance between
tumor cells and immune cells must be considered.
The present results suggested that the inhibition of
JUNB may be a key indicator of the regulation of the
APOA2-associated PPARα pathway in HCC (31). APOA2 is a well-known member of
the apolipoprotein family (40),
which is functionally involved in triglyceride, fatty acid and
glucose metabolism. This gene family has been previously reported
to be overexpressed in HCC and regulated by the PPARα pathway
(41). In addition, together with
the co-expression of APOA2 and JUNB observed in the
TCGA dataset and the transcriptional regulation analysis, a
potential regulation of APOA2 and JUNB by the PPARα
pathway was identified. Finally, the present survival analysis in
HCC patients suggested that the investigation of JUNB may
facilitate the identification of novel therapeutic targets for HCC
patients. The inhibition of some positive regulators of T cells,
such as TNF, NFKBIA, FOS, JUN and DDX3X may lead to
the downregulation of the T cell receptor signaling pathway
(42–44), affecting the hepatitis B and cellular
oxidant detoxification response, which may be caused by the
abnormal growth of tumor cells and the accumulation of
stress-associated factors. In this condition, tumor cells may
require more energy to sustain their proliferation and to adapt to
the hypoxic micro-environment.
Although the present results could potentially
contribute to the development of novel therapeutics to treat
patients with HCC, the lack of experimental validations on mRNA
levels are still the main limitations for the study. In addition,
investigating the expression levels of the products of candidate
mRNAs, the proteins, is equally critical to validate the results,
as they play a central role in biological processes. Furthermore,
validations using both in vivo and in vitro models
should be further investigated, such as ChIP to detect the
potential regulatory regions on JUNB, Junb defective mice to
study the important functions of Junb.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81501561); the
Natural Science Foundation of Guangdong Province (grant nos.
2014A030310043 and 2017A030313873) and the Science and Technology
Planning Project of Zhuhai (grant no. 20171009E030008).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
PY, BZ, PP and JM conceived and designed the
experiments. PY, YM, AW, XH, YL, YY and YW analyzed the data and
wrote the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable. No patients were enrolled in the
study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
HCC
|
hepatocellular carcinoma
|
|
PBMC
|
peripheral blood mononuclear cells
|
|
TCGA
|
The Cancer Genome Atlas
|
|
TF
|
transcription factors
|
|
UMI
|
unique molecular identifier
|
References
|
1
|
Harris PS, Hansen RM, Gray ME, Massoud OI,
McGuire BM and Shoreibah MG: Hepatocellular carcinoma surveillance:
An evidence-based approach. World J Gastroenterol. 25:1550–1559.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bou-Nader M, Caruso S, Donne R,
Celton-Morizur S, Calderaro J, Gentric G, Cadoux M, L'Hermitte A,
Klein C, Guilbert T, et al: Polyploidy spectrum: A new marker in
HCC classification. Gut. (pii): gutjnl-2018-318021. 2019.(Epub
ahead of print). PubMed/NCBI
|
|
3
|
Kim HL, An J, Park JA, Park SH, Lim YS and
Lee EK: Magnetic resonance imaging is cost-effective for
hepatocellular carcinoma surveillance in high-risk patients with
cirrhosis. Hepatology. 69:1599–1613. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yang Z, Zou R, Zheng Y, Qiu J, Shen J,
Liao Y, Zhang Y, Wang C, Wang Y, Yuan Y, et al: Lipiodol deposition
in portal vein tumour thrombus predicts treatment outcome in HCC
patients after transarterial chemoembolisation. Eur Radiol.
29:5752–5762. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Al-Zoughbi W, Huang J, Paramasivan GS,
Till H, Pichler M, Guertl-Lackner B and Hoefler G: Tumor
macroenvironment and metabolism. Semin Oncol. 41:281–195. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mantovani A, Allavena P, Sica A and
Balkwill F: Cancer-related inflammation. Nature. 454:436–444. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wu T and Dai Y: Tumor microenvironment and
therapeutic response. Cancer Lett. 387:61–68. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hirata E and Sahai E: Tumor
microenvironment and differential responses to therapy. Cold Spring
Harb Perspect Med. 7:a0267812017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li Q, Ma L, Shen S, Guo Y, Cao Q, Cai X,
Feng J, Yan Y, Hu T, Luo S, et al: Intestinal
dysbacteriosis-induced IL-25 promotes development of HCC via
alternative activation of macrophages in tumor microenvironment. J
Exp Clin Cancer Res. 38:3032019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Francis JM, Kiezun A, Ramos AH, Serra S,
Pedamallu CS, Qian ZR, Banck MS, Kanwar R, Kulkarni AA, Karpathakis
A, et al: Somatic mutation of CDKN1B in small intestine
neuroendocrine tumors. Nat Genet. 45:1483–1486. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kang HJ, Oh JH, Chun SM, Kim D, Ryu YM,
Hwang HS, Kim SY, An J, Cho EJ, Lee H, et al: Immunogenomic
landscape of hepatocellular carcinoma with immune cell stroma and
EBV-positive tumor-infiltrating lymphocytes. J Hepatol. 71:91–103.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yuan Q, Hou S, Zhai J, Tian T, Wu Y, Wu Z,
He J, Chen Z and Zhang J: S100A4 promotes inflammation but
suppresses lipid accumulation via the STAT3 pathway in chronic
ethanol-induced fatty liver. J Mol Med (Berl). 97:1399–1412. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Schmidt-Hansen B, Ornås D, Grigorian M,
Klingelhöfer J, Tulchinsky E, Lukanidin E and Ambartsumian N:
Extracellular S100A4(mts1) stimulates invasive growth of mouse
endothelial cells and modulates MMP-13 matrix metalloproteinase
activity. Oncogene. 23:5487–5495. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Grum-Schwensen B, Klingelhöfer J,
Grigorian M, Almholt K, Nielsen BS, Lukanidin E and Ambartsumian N:
Lung metastasis fails in MMTV-PyMT oncomice lacking S100A4 due to a
T-cell deficiency in primary tumors. Cancer Res. 70:936–947. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ringelhan M, Pfister D, O'Connor T,
Pikarsky E and Heikenwalder M: The immunology of hepatocellular
carcinoma. Nat Immunol. 19:222–232. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hwang B, Lee JH and Bang D: Single-cell
RNA sequencing technologies and bioinformatics pipelines. Exp Mol
Med. 50:962018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zheng C, Zheng L, Yoo JK, Guo H, Zhang Y,
Guo X, Kang B, Hu R, Huang JY, Zhang Q, et al: Landscape of
infiltrating t cells in liver cancer revealed by single-cell
sequencing. Cell. 169:1342–1356 e16. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sia D, Jiao Y, Martinez-Quetglas I, Kuchuk
O, Villacorta-Martin C, Castro de Moura M, Putra J, Camprecios G,
Bassaganyas L, Akers N, et al: Identification of an immune-specific
class of hepatocellular carcinoma, based on molecular features.
Gastroenterology. 153:812–826. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhong DN, Ning QY, Wu JZ, Zang N, Wu JL,
Hu DF, Luo SY, Huang AC, Li LL and Li GJ: Comparative proteomic
profiles indicating genetic factors may involve in hepatocellular
carcinoma familial aggregation. Cancer Sci. 103:1833–1838. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zheng H, Pomyen Y, Hernandez MO, Li C,
Livak F, Tang W, Dang H, Greten TF, Davis JL, Zhao Y, et al:
Single-cell analysis reveals cancer stem cell heterogeneity in
hepatocellular carcinoma. Hepatology. 68:127–140. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Neal JT, Li X, Zhu J, Giangarra V,
Grzeskowiak CL, Ju J, Liu IH, Chiou SH, Salahudeen AA, Smith AR, et
al: Organoid modeling of the tumor immune microenvironment. Cell.
175:1972–1988 e16. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee KH and Kim JR: Regulation of
HGF-mediated cell proliferation and invasion through NF-κB, JunB,
and MMP-9 cascades in stomach cancer cells. Clin Exp Metastasis.
29:263–272. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang L, Fan J, Francis JM, Georghiou G,
Hergert S, Li S, Gambe R, Zhou CW, Yang C, Xiao S, et al:
Integrated single-cell genetic and transcriptional analysis
suggests novel drivers of chronic lymphocytic leukemia. Genome Res.
27:1300–1311. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gao J, Aksoy BA, Dogrusoz U, Dresdner G,
Gross B, Sumer SO, Sun Y, Jacobsen A, Sinha R, Larsson E, et al:
Integrative analysis of complex cancer genomics and clinical
profiles using the cBioPortal. Sci Signal. 6:ppl12013. View Article : Google Scholar
|
|
26
|
Fishilevich S, Nudel R, Rappaport N, Hadar
R, Plaschkes I, Iny Stein T, Rosen N, Kohn A, Twik M, Safran M, et
al: GeneHancer: Genome-wide integration of enhancers and target
genes in GeneCards. Database (Oxford). 2017. View Article : Google Scholar
|
|
27
|
Fennemann FL, de Vries IJM, Figdor CG and
Verdoes M: Attacking tumors from all sides: Personalized multiplex
vaccines to tackle intratumor heterogeneity. Front Immunol.
10:8242019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ali M, Khan SY, Vasanth S, Ahmed MR, Chen
R, Na CH, Thomson JJ, Qiu C, Gottsch JD and Riazuddin SA:
Generation and proteome profiling of pbmc-originated, ipsc-derived
corneal endothelial cells. Invest Ophthalmol Vis Sci. 59:2437–2444.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
van der Wijst MGP, Brugge H, de Vries DH,
Deelen P, Swertz MA; LifeLines Cohort Study BIOS Consortium, ;
Franke L: Single-cell RNA sequencing identifies celltype-specific
cis-eQTLs and co-expression QTLs. Nat Genet. 50:493–497. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Song Q, Hawkins GA, Wudel L, Chou PC,
Forbes E, Pullikuth AK, Liu L, Jin G, Craddock L, Topaloglu U, et
al: Dissecting intratumoral myeloid cell plasticity by single cell
RNA-seq. Cancer Med. 8:3072–3085. 2019.PubMed/NCBI
|
|
31
|
Guo C, Liu QG, Zhang L, Song T and Yang X:
Expression and clinical significance of p53, JunB and KAI1/CD82 in
human hepatocellular carcinoma. Hepatobiliary Pancreat Dis Int.
8:389–396. 2009.PubMed/NCBI
|
|
32
|
Guo C, Liu Q, Zhang L, Yang X, Song T and
Yao Y: Double lethal effects of fusion gene of wild-type p53 and
JunB on hepatocellular carcinoma cells. J Huazhong Univ Sci
Technolog Med Sci. 32:663–668. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chang YS, Yeh KT, Yang MY, Liu TC, Lin SF,
Chan WL and Chang JG: Abnormal expression of JUNB gene in
hepatocellular carcinoma. Oncol Rep. 13:433–438. 2005.PubMed/NCBI
|
|
34
|
Zhai X, Zhu H, Wang W, Zhang S, Zhang Y
and Mao G: Abnormal expression of EMT-related proteins, S100A4,
vimentin and E-cadherin, is correlated with clinicopathological
features and prognosis in HCC. Med Oncol. 31:9702014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Dukhanina EA, Lukyanova TI, Romanova EA,
Guerriero V, Gnuchev NV, Georgiev GP, Yashin DV and Sashchenko LP:
A new role for PGRP-S (Tag7) in immune defense: Lymphocyte
migration is induced by a chemoattractant complex of Tag7 with
Mts1. Cell Cycle. 14:3635–3643. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wu J, Ma S, Hotz-Wagenblatt A, Angel P,
Mohr K, Schlimbach T, Schmitt M and Cui G: Regulatory T cells sense
effector T-cell activation through synchronized JunB expression.
FEBS Lett. 593:1020–1029. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kim VM, Blair AB, Lauer P, Foley K, Che X,
Soares K, Xia T, Muth ST, Kleponis J, Armstrong TD, et al:
Anti-pancreatic tumor efficacy of a listeria-based, annexin
A2-targeting immunotherapy in combination with anti-PD-1
antibodies. J Immunother Cancer. 7:1322019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Nagasawa M, Akasaka Y, Ide T, Hara T,
Kobayashi N, Utsumi M and Murakami K: Highly sensitive upregulation
of apolipoprotein A-IV by peroxisome proliferator-activated
receptor alpha (PPARalpha) agonist in human hepatoma cells. Biochem
Pharmacol. 74:1738–1746. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Harris WP, Wong KM, Saha S, Dika IE and
Abou-Alfa GK: Biomarker-driven and molecular targeted therapies for
hepatobiliary cancers. Semin Oncol. 45:116–123. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ballester M, Revilla M, Puig-Oliveras A,
Marchesi JA, Castelló A, Corominas J, Fernández AI and Folch JM:
Analysis of the porcine APOA2 gene expression in liver,
polymorphism identification and association with fatty acid
composition traits. Anim Genet. 47:552–559. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Thulin P, Glinghammar B, Skogsberg J,
Lundell K and Ehrenborg E: PPARdelta increases expression of the
human apolipoprotein A-II gene in human liver cells. Int J Mol Med.
21:819–824. 2008.PubMed/NCBI
|
|
42
|
Lu G, Zhang G, Zheng X, Zeng Y, Xu Z, Zeng
W and Wang K: c9, t11- conjugated linoleic acid induces HCC cell
apoptosis and correlation with PPAR-γ signaling pathway. Am J
Transl Res. 7:2752–2763. 2015.PubMed/NCBI
|
|
43
|
Kahraman DC, Kahraman T and Cetin-Atalay
R: Targeting PI3K/Akt/mTOR pathway identifies differential
expression and functional role of IL-8 in liver cancer stem cell
enrichment. Mol Cancer Ther. 18:2146–2157. 2019.PubMed/NCBI
|
|
44
|
Liu L, Cao Y, Chen C, Zhang X, McNabola A,
Wilkie D, Wilhelm S, Lynch M and Carter C: Sorafenib blocks the
RAF/MEK/ERK pathway, inhibits tumor angiogenesis, and induces tumor
cell apoptosis in hepatocellular carcinoma model PLC/PRF/5. Cancer
Res. 66:11851–11858. 2006. View Article : Google Scholar : PubMed/NCBI
|