Introduction
Breast cancer is a highly heterogeneous malignant
tumor, and its morbidity and mortality are second only to lung
cancer, which poses a great threat to human health (1). In recent years, the mortality rate of
breast cancer has been increasing year by year (2). Although the prognosis of breast cancer
patients is greatly improved under comprehensive treatment, 20–25%
of patients will eventually have local recurrence or even distant
metastasis, eventually leading to death. Studies have shown that
the development, early diagnosis, comprehensive treatment and
prognosis of breast cancer are closely related to the abnormal
level of non-coding ribonucleic acid (RNA) (3), so it is necessary to explore the
pathogenesis of breast cancer from the perspective of non-coding
RNA.
Long-chain non-coding RNA (lncRNA) is a RNA molecule
that is more than 200 nt in length and has been proved to be
involved in various biological processes in cells (4,5). lncRNA
can actively participate in the regulation of various biological
functions and plays an important role in a variety of tumors mainly
through epigenetic changes, cell cycle regulation, regulation of
miRNAs and participation in signaling pathways (6,7). For
instance, lncRNA plays an important role in the development of
breast cancer (8). By gene chip
high-throughput analysis of differential level of lncRNA in breast
cancer and normal breast tissue, studies have found that 220
lncRNAs are abnormally expressed in breast cancer tissues, of which
129 lncRNAs are specifically expressed in breast cancer and can be
used as breast cancer biomarkers (9). Another study found that lncRNA H19 is
overexpressed in estrogen receptor positive breast cancer and
promotes breast cancer growth and proliferation (10). lncRNA GACAT1 is highly expressed in
gastric cancer and closely related to the prognosis of gastric
cancer, which can be used as a tumor marker for gastric cancer
(11,12). Through previous studies, we found
that GACAT1 is also highly expressed in breast cancer, but the
study on GACAT1 in breast cancer has not been reported.
MicroRNAs (miRNAs) are a class of small-molecule
non-coding RNAs found in eukaryotes. They are composed of ~22
nucleotides and bind to the corresponding target mRNA, resulting in
degradation of target mRNA or inhibited post-transcriptional
translation. At present, scientists have identified 474 miRNAs in
the human genome, which could regulate ~30% of the encoded protein
genes, and each miRNA regulates ~100 target mRNAs (13). The relationship between miRNAs and
human life activities and diseases is receiving increasing
attention. In particular, recent studies have shown that they are
closely related to the occurrence and development of human
diseases. For example, in patients with lung cancer, the level of
miRNA1et-7 is conspicuously reduced. Meanwhile, let-7a transfection
into A549 cell line could inhibit lung cancer cell proliferation
(14). In the study on chronic
lymphocytic leukemia, it was found that the level of miR-15 and
miR-16 in leukemia cells of patients with chronic lymphocytic
leukemia is downregulated, while the level of B-cell lymphoma-2
(Bcl-2) is increased. There is a significant negative correlation
between miR-15 and miR-16 expression and Bcl-2 protein levels, and
miR-15 and miR-16 functions by targeting degradation of Bcl-2
(15).
In the present, through bioinformatics analysis, it
was found that microRNA-875-3p might be a potential target gene for
long-chain non-coding RNA GACAT1, moreover, its expression was low
in breast cancer tissues and cells. This study investigated the
role of GACAT1 in breast cancer to verify whether it could function
by degrading microRNA-875-3p.
Patients and methods
General information
Fresh breast cancer tissue and adjacent normal
tissues were collected from 24 patients with breast cancer who
underwent surgery. The enrollment conditions included: i) exclusion
of other malignant tumor history; ii) first surgery, no
radiotherapy before surgery; iii) the pathology diagnosis was
performed by the pathologist's reading film; iv) the clinical
medical record data and the postoperative follow-up data were
complete. This study was approved by the Ethics Committee of Linyi
Cancer Hospital (Linyi, China). Signed informed consents were
obtained from all participants before the study. The specimens were
taken from the tumors removed during the operation, and stored in a
liquid nitrogen tank within 15 min.
Cell culture
Normal Hs578Bst cells as well as breast cancer cell
lines MDA-MB-231, BCap-37 and MCF-7 were purchased from American
Type Culture Collection (ATCC). The cells were cultured in
Dulbecco's modified Eagles medium (DMEM) (Gibco; Thermo Fisher
Scientific, Inc.) (containing 10% fetal bovine serum (FBS), 100
U/ml penicillin and 0.1 mg/ml streptomycin) at 37°C, in a 5%
CO2 incubator.
Cell transfection
The cells were inoculated one day before the
transfection. The next day, plasmid, miRNA mimics and their
respective negative control sequences (50–100 nM) were transfected
by Lipofectamine 3000 (Invitrogen; Thermo Fisher Scientific, Inc.).
RNase-free diethyl pyrocarbonate (DEPC) water (both from Beyotime)
and Eppendorf (EP) tubes were used for transfection.
Tissue and cell total RNA extraction
and quantitative real-time polymerase chain reaction (qRT-PCR)
Extraction was carried out by the TRIzol method
(Invitrogen; Thermo Fisher Scientific, Inc.). Chloroform (200 µl)
was added to each l ml of TRIzol-dissolved EP tube, mixed by
inversion, placed in an ice box for 5 min, and centrifuged at 4°C,
12,000 × g for 15 min carefully pipeting. The extracted RNA was
applied to the NanoDrop 2000c assay to determine its concentration
and the 260/280 absorbance was used to evaluate the quality. The
measurement was repeated twice, and the 260/280 absorbance ratio
was between 1.8 and 2.0. The total RNA was then reverse-transcribed
into complementary deoxyribonucleic acid (cDNA) which was used for
further real-time PCR. Detection of the level of the target gene
was performed using an ABI StepOnePlus real-time quantitative
fluorescent PCR instrument by the SYBR real-time qPCR kit (Applied
Biosystems; Thermo Fisher Scientific, Inc.). Each well contained 10
µl SYBR-Green Master Mix, 1 µl upstream primer, 1 µl downstream
primer, 0.4 µl 50X Rox Dye, 2 µl cDNA and 5.6 µl enzyme-free water.
The reaction conditions were 95°C for 10 min, 95°C for 15 sec and
60°C for 60 sec, for 40 cycles. The data were statistically
analyzed using the StepOne Software version v2.1 (Applied
Biosystems; Thermo Fisher Scientific, Inc.) software and the primer
sequences were derived as follows: microRNA-875-3p, forward,
5′-ACACTCCAGCTGGGUAUACCUCAGUUUUAU-3′ and reverse,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGCACCUGA-3′; U6, forward,
5′-CTCGCTTCGGCAGCAGCACATATA-3′ and reverse,
5′-AAATATGGAACGCTTCACGA-3′; GACAT1, forward,
5′-ACCGGAGGAAAATCCCTAGC-3′ and reverse, 5′-CCATAAAAGGGGCGGCTG-3′;
GAPDH, forward, 5′-GAAGAGAGAGACCCTCACGCTG-3′ and reverse,
5′-ACTGTGAGGAGGGGAGATTCAGT-3′; Stonin2 (STON2), forward,
5′-ACCATGTGATTGCCACCCAC and reverse, 5′-AGCTCTCGGACTGGTCTGG-3′.
Luciferase reporting assay
Reporter plasmid was constructed and termed
pmirGL0-STON2-7 7 UTR-wild type (Wt) or pmirGLO-STON2-37 UTR-mutant
(Mut), which were inserted into the dual-fluorescence of wild-type
and mutant STON2-3′UTR, respectively. Breast cancer cells were
seeded into 48-well plates, and the plasmid was co-transfected with
miRNA-875-5p mimics, then the luciferase activity was detected 48 h
later.
Detection of cell cycle
Twenty-four hours after transfection, cells were
collected, washed twice with phosphate-buffered saline (PBS) and
then 1 ml of DNA Staining solution (Takara) was added, vortexed and
mixed, and incubated at room temperature in the dark. After 30 min,
the samples were detected by MACS flow cytometry (Beckman
Coulter).
Cell proliferation
Transfected cells were seeded into 96-well plates
(Corning, Inc.) at a density of 3–5×103 cells per well,
and 10 replicate wells were set for each group. 100 µl of a mixture
of cell counting kit-8 (CCK-8) reagent (Dojindo) premixed in a
ratio of 1:10 was added and then the cells were incubated in a 37°C
incubator for 1 h. Then the enzyme was added. The optical density
(OD) value of each replicate well was measured by microplate reader
at 450 nm wavelength.
Statistical analysis
The measurement data were expressed as mean ±
standard deviation, and t-test was used to examine the mean
difference between two groups. Single factor variance analysis was
used when multiple sets of data meet normality and the homogeneity
of the variance. Pairwise comparison was used if there was a
difference between the groups. Nonparametric test was used when the
data did not meet normality and the homogeneity of the variance.
P<0.05 was considered statistically significant.
Results
GACAT1 is clearly expressed in breast
cancer
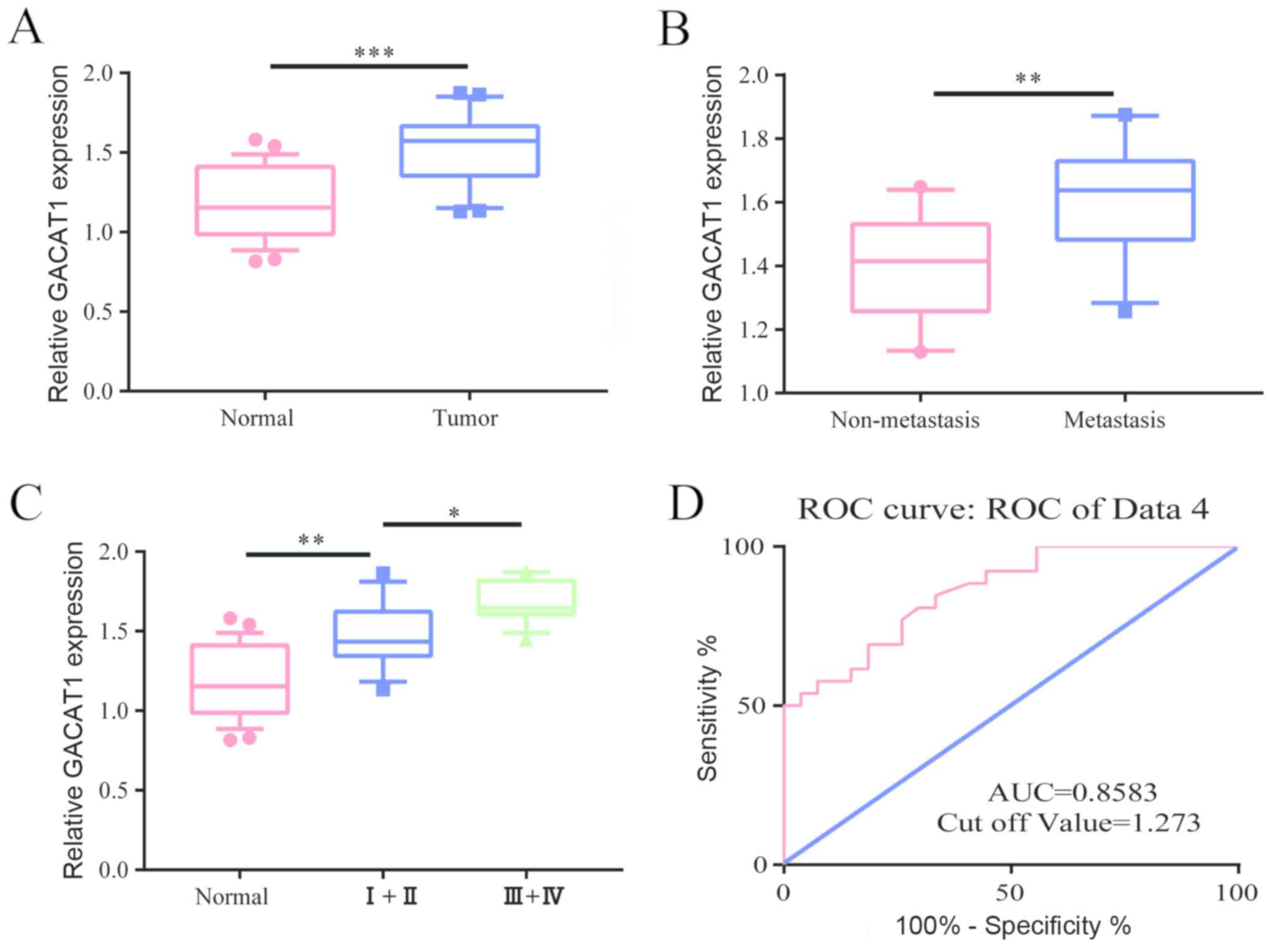
First, we examined the level of GACAT1 in 24 breast
cancer patients. The results showed that microRNA-875-3p was
conspicuously expressed in breast cancer tissues compared to that
of adjacent tissues (Fig. 1A).
Furthermore, we found that in patients with cancer metastasis, the
level of GACAT1 was conspicuously higher than that of patients
without metastasis (Fig. 1B). We
then examined the level of GACAT1 in breast cancer at different
stages, and the results showed that GACAT1 level was conspicuously
higher in patients with grade III to IV breast cancer than in
patients with stage I to II (Fig.
1C). By analyzing the clinical information of breast cancer
patients, the survival curve was plotted and the results showed
that the area under the curve was 0.8583 and the cutoff value was
1.273 (Fig. 1D), which indicated
that GACAT1 can be used as a biomarker for breast cancer.
GACAT1 promotes proliferation and cycle
of breast cancer cells
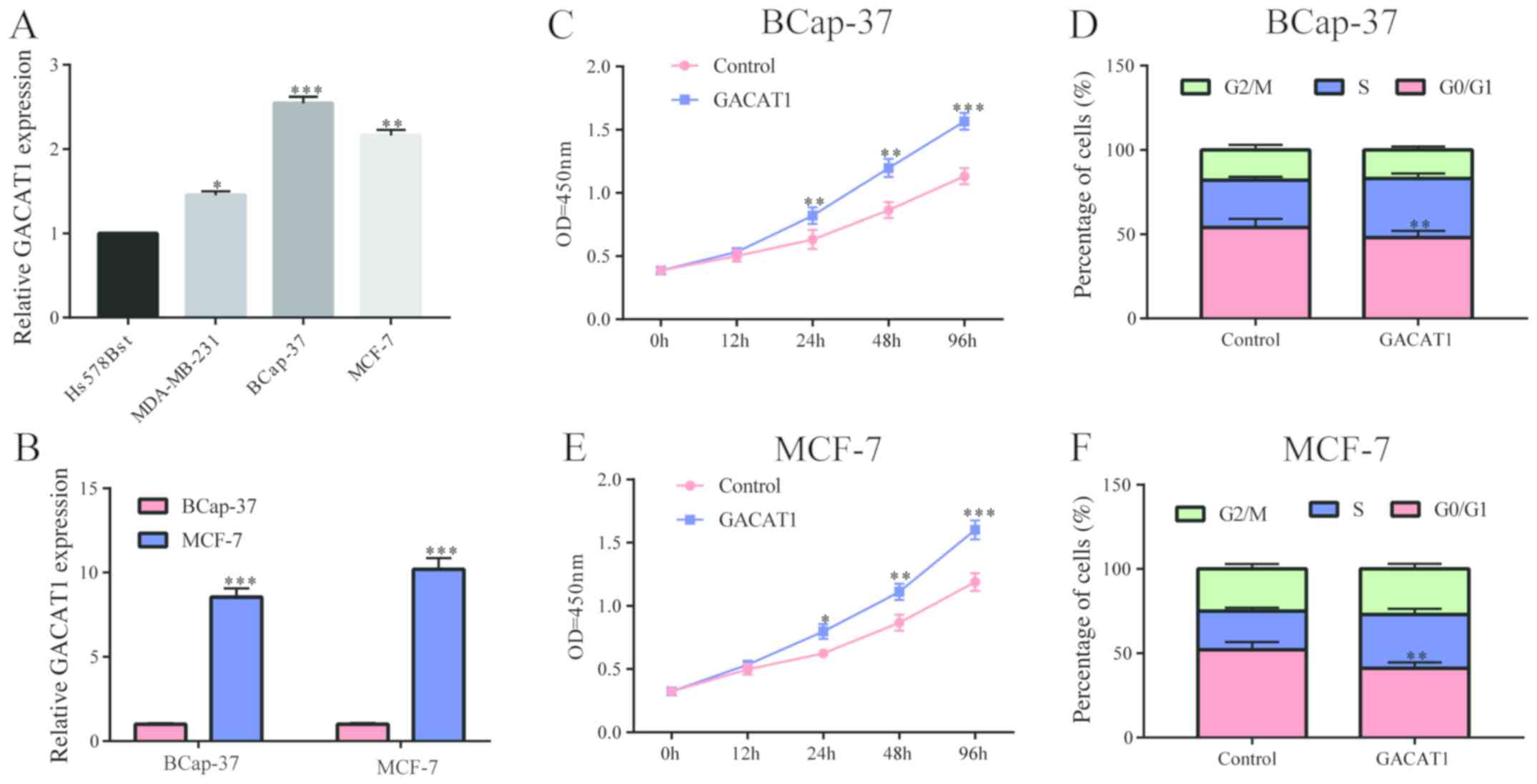
In breast cancer cell lines, we also
examined the GACAT1 level
The results showed that it was generally highly
expressed in breast cancer cell lines compared to normal breast
cells (Fig. 2A). After transfection
of the GACAT1 overexpression plasmid in MCF-7 and BCap-37 cells, we
examined the efficiency of transfection, and the results showed
that GACAT1 was high and stably expressed (Fig. 2B). Subsequently, we applied CCK-8
assay and flow cytometry to examine the effect of GACAT1 on
proliferation and cell cycle of breast cancer cells. The results
showed that the upregulation of GACAT1 conspicuously increased the
proliferation of BCap-37, and also clearly promoted the cell cycle
of BCap-37 (Fig. 2C and D), which
was consistent with MCF-7 cells (Fig. 2E
and F). These results demonstrated that GACAT1 promoted
proliferation and cell cycle of breast cancer cells.
GACAT1 works by adsorbing
microRNA-875-3p
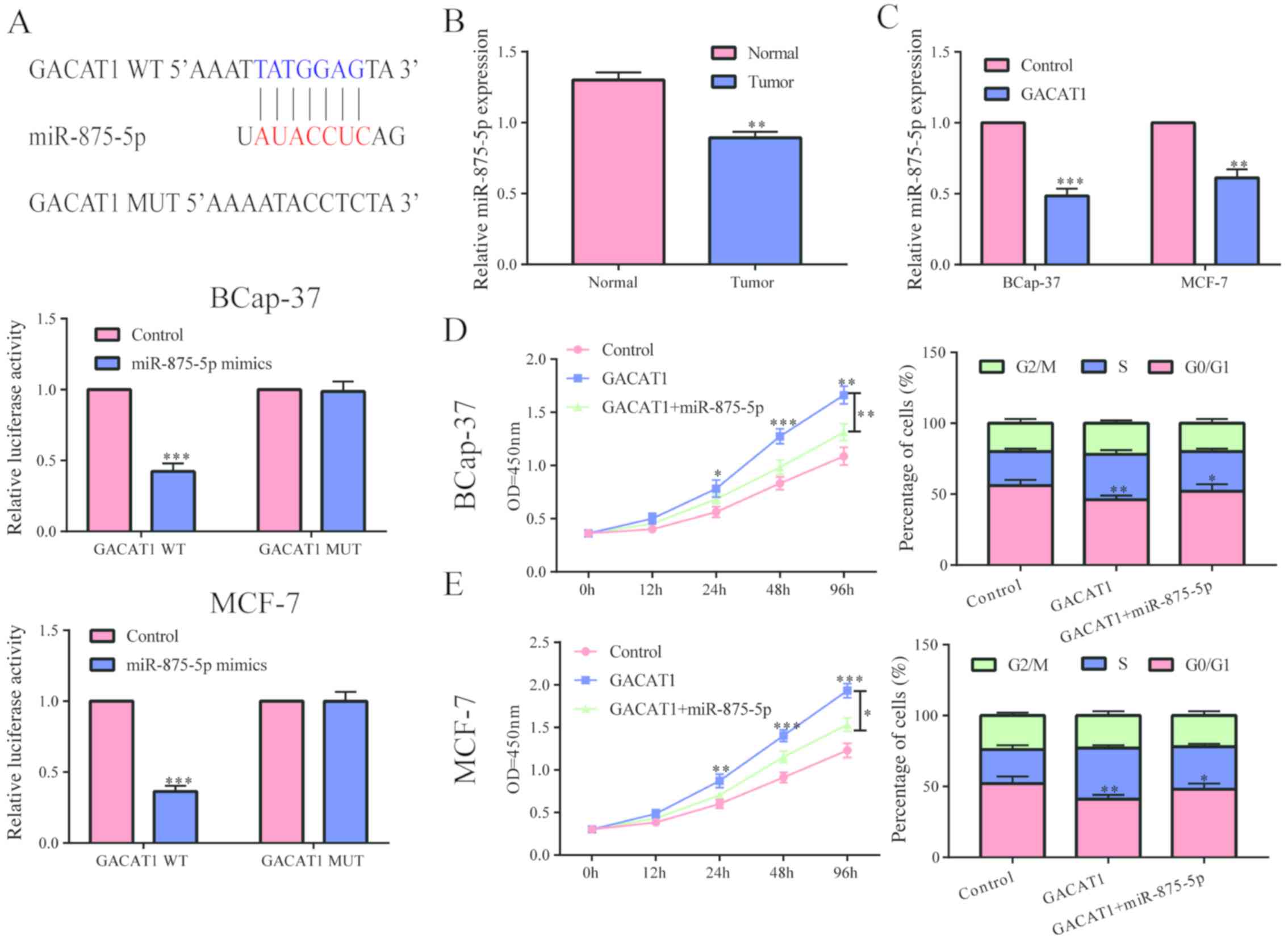
Through bioinformatics analysis, we found that
GACAT1 might bind to microRNA-875-3p, and confirmed their binding
by dual luciferase reporter gene experiments (Fig. 3A). Further studies found that
microRNA-875-3p was conspicuously under expressed in breast cancer
tissues (Fig. 3B). After
upregulating the level of GACAT1 in BCap-37 and MCF-7 cells, the
level of microRNA-875-3p was clearly decreased (Fig. 3C). The above results indicated that
GACAT1 can inhibit the level of microRNA-875-3p. To verify whether
GACAT1 functioned by targeting microRNA-875-3p, we first
upregulated GACAT1 and then overexpressed microRNA-875-3p as well,
and found that the effect of GACAT1 on cell proliferation in
BCap-379 and MCF-7 cells was partially inhibited by microRNA-875-3p
overexpression (Fig. 3D). Moreover,
we found that upregulation of microRNA-875-3p in BCap-379 and MCF-7
cells partially inhibited the promoting effect of GACAT1 on the
cell cycle (Fig. 3E). Based on the
above results, we concluded that GACAT1 may play a role by
adsorbing microRNA-875-3p and then regulate its downstream target
genes.
microRNA-875-3p functions by targeting
degradation of STON2
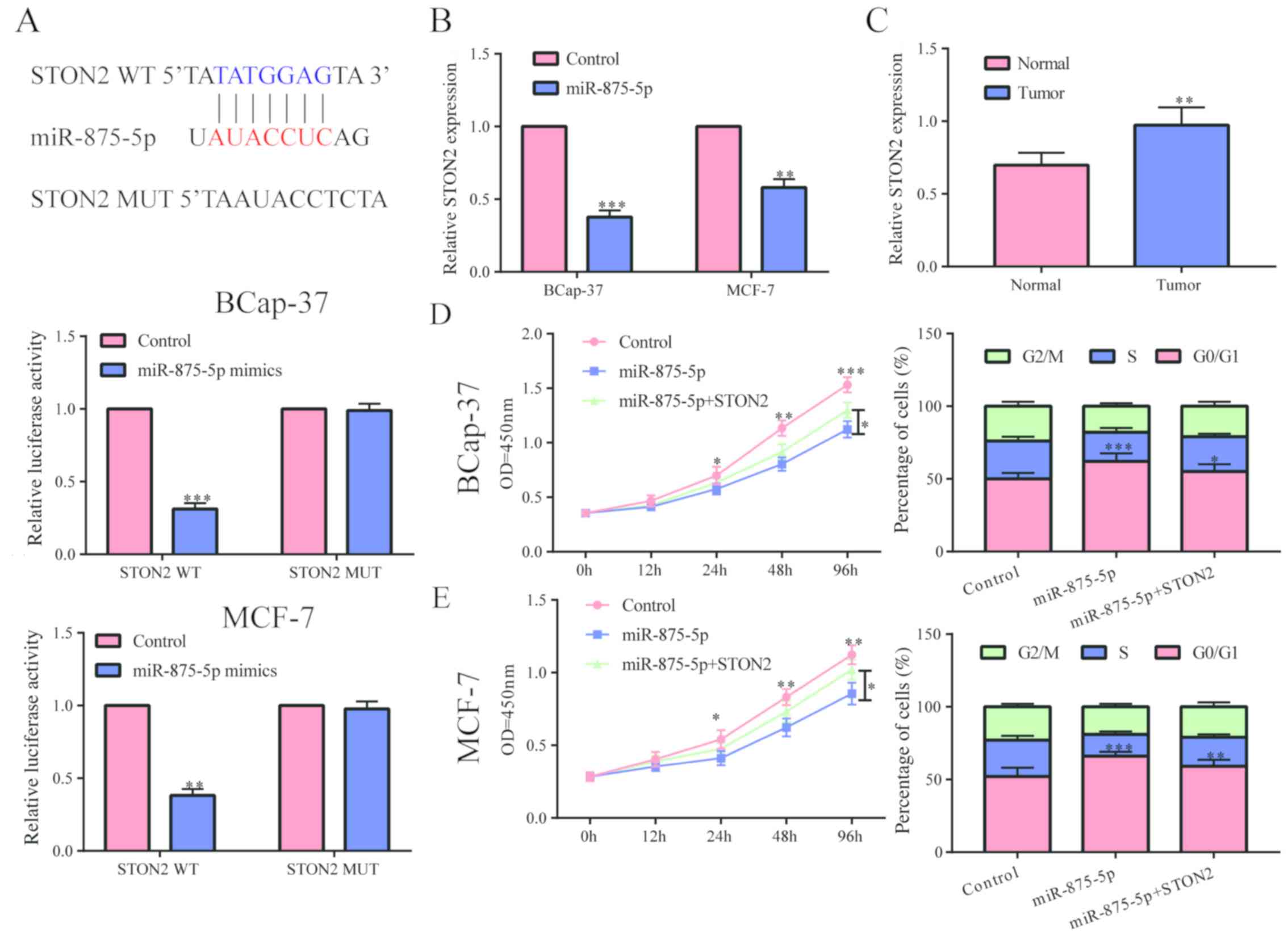
It was predicted by website that STON2 may be a
potential target gene of microRNA-875-3p, and we confirmed their
binding relationship by luciferase reporter gene experiment
(Fig. 4A). In breast cancer cells,
we found that the level of STON2 was conspicuously reduced after
overexpressing microRNA-875-3p (Fig.
4B), indicating that STON2 can be degraded by microRNA-875-3p.
We examined the level of STON2 in breast cancer tissues and found
that it existed at a significant high level in breast cancer
(Fig. 4C). To test whether
microRNA-875-3p worked by inhibiting the level of STON2, a series
of recovery experiments were performed. The results showed that
upregulation of STON2 in MCF-7 and BCap-37 cells partially
abolished the inhibition of cell proliferation and cycle by
microRNA-875-3p (Fig. 4D and E).
Based on the above results, we hypothesized that GACAT1 may
function by inhibiting the degradation of downstream STON2 by
adsorbing microRNA-875-3p.
Discussion
The competitive endogenous RNA (ceRNAs) hypothesis
reveals a novel mode of regulation of RNA-interacting interactions,
including messenger RNA (mRNA), pseudogene, long-chain non-coding
RNA (lncRNA) and circular RNA (circRNA). By stimulating the same
miRNA through microRNA response elements (MREs), the level of miRNA
is reduced, thereby reducing or reducing the effect of miRNA on
target genes. ceRNA is widely involved in a series of physiological
and pathological processes including cell differentiation,
proliferation, apoptosis, and growth and invasion of tumor cells
(16,17). There have been many studies on the
role of ceRNA. Studies have found that lncRNA HOTAIR is
conspicuously upregulated in renal cell tumors and acts as an
endogenous RNA, which could promote tumor progression by
downregulating the level of miR-217. Other studies have shown that
lncRNA RMRP also plays a role as a miRNA sponge in lung cancer. By
adsorbing miR-206, it upregulates the level of miRNA downstream
target genes and plays a role in the development of lung
cancer.
In this study, we found that GACAT1 was highly
expressed in breast cancer by qPCR. After staging breast cancer
patients and dividing them into metastasis group and non-metastasis
group, we further analyzed that GACAT1 level was closely related to
breast cancer staging and metastasis, suggesting that GACAT1 could
indicate poor prognosis. In vitro, we overexpressed GACAT1
in breast cancer cell lines and found that GACAT1 conspicuously
promoted the proliferation and cell cycle of breast cancer cells,
suggesting that it may play a role in the development of breast
cancer.
To explore whether GACAT1 also functioned as a miRNA
sponge, we performed prediction analysis and found that
microRNA-875-3p may be a potential target for GACAT1. Previous
studies have shown that microRNA-875-3p plays a role as a tumor
suppressor gene in a variety of tumors, and its upregulation can
often inhibit the proliferation and invasion of tumor cells
(18,19). Therefore, we also examined the level
of microRNA-875-3p in breast cancer patients, and the results
showed that microRNA-875-3p was conspicuously under-expressed in
breast cancer. We confirmed the binding relationship between GACAT1
and microRNA-875-3p by luciferase reporter gene assay. Besides, the
level of microRNA-875-3p was conspicuously decreased after
upregulating GACAT1, indicating that GACAT1 can play the role of
ceRNA to downregulate microRNA-875-3p level. To further confirm
that GACAT1 functioned through microRNA-875-3p, we simultaneously
upregulated GACAT1 and microRNA-875-3p in breast cancer cell lines,
and found that microRNA-875-3p partially inhibited the inhibition
effect of GACAT1 on cell proliferation and cycle.
In addition, since miRNAs play a role in the
targeted degradation of their target genes, we further explored
their potential target genes through site prediction and analysis.
We found that STON2 may be a potential target gene for
microRNA-875-3p. The STON2 gene is a clathrin-related protein that
is closely related to the regulation of intracellular complexes
(20,21). In ovarian cancer, studies have shown
that high level of STON2 can promote tumor invasion and predict a
poor prognosis (22). In this study,
we verified that microRNA-875-3p could bind to STON2 by dual
luciferase reporter gene assay and that STON2 was highly expressed
in breast cancer. Through a series of recovery experiments, we
found that overexpression of STON2 partially abolished the
inhibitory effect of microRNA-875-3p on proliferation and cell
cycle of breast cancer cells, indicating that microRNA-875-3p may
exert its role through regulating STON2.
Based on the above results, we hypothesized that
lncRNA GACAT1 can adsorb microRNA-875-3p to upregulate the level of
STON2, thereby promoting the progression of breast cancer.
In conclusion, in this study, we conducted an
in-depth study of the role and mechanism of GACAT1 in breast
cancer, and demonstrated that GACAT1 could play a vital role in the
progression of breast cancer, which also provides theoretical basis
for new prevention and treatment for breast cancer.
Acknowledgements
Not applicable.
Funding
Not funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
QW and XQ designed the study and performed the
experiments, QW and JX collected the data, QR and XL analyzed the
data, QW and XQ prepared the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
This study was approved by the Ethics Committee of
Linyi Cancer Hospital (Linyi, China). Signed informed consents were
obtained from the patients and/or guardians.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhuang X and Wang J: Correlations of MRP1
gene with serum TGF-β1 and IL-8 in breast cancer patients during
chemotherapy. J BUON. 23:1302–1308. 2018.PubMed/NCBI
|
|
3
|
Huang NS, Chi YY, Xue JY, Liu MY, Huang S,
Mo M, Zhou SL and Wu J: Long non-coding RNA metastasis associated
in lung adenocarcinoma transcript 1 (MALAT1) interacts with
estrogen receptor and predicted poor survival in breast cancer.
Oncotarget. 7:37957–37965. 2016.PubMed/NCBI
|
|
4
|
Ponting CP, Oliver PL and Reik W:
Evolution and functions of long noncoding RNAs. Cell. 136:629–641.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhou CX, Wang X, Yang N, Xue SK, Li WC and
Xie PP: lncRNA LET function as a tumor suppressor in breast cancer
development. Eur Rev Med Pharmacol Sci. 22:6002–6007.
2018.PubMed/NCBI
|
|
6
|
Khan FS, Ali I, Afridi UK, Ishtiaq M and
Mehmood R: Epigenetic mechanisms regulating the development of
hepatocellular carcinoma and their promise for therapeutics.
Hepatol Int. 11:45–53. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xia T, Liao Q, Jiang X, Shao Y, Xiao B, Xi
Y and Guo J: Long noncoding RNA associated-competing endogenous
RNAs in gastric cancer. Sci Rep. 4:60882014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dvinge H, Git A, Gräf S, Salmon-Divon M,
Curtis C, Sottoriva A, Zhao Y, Hirst M, Armisen J, Miska EA, et al:
The shaping and functional consequences of the microRNA landscape
in breast cancer. Nature. 497:378–382. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gibb EA, Vucic EA, Enfield KS, Stewart GL,
Lonergan KM, Kennett JY, Becker-Santos DD, MacAulay CE, Lam S,
Brown CJ, et al: Human cancer long non-coding RNA transcriptomes.
PLoS One. 6:e259152011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sun H, Wang G, Peng Y, Zeng Y, Zhu QN, Li
TL, Cai JQ, Zhou HH and Zhu YS: H19 lncRNA mediates
17β-estradiol-induced cell proliferation in MCF-7 breast cancer
cells. Oncol Rep. 33:3045–3052. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xing C, Cai Z, Gong J, Zhou J, Xu J and
Guo F: Identification of potential biomarkers involved in gastric
cancer through integrated analysis of non-coding RNA associated
competing endogenous RNAs network. Clin Lab. 64:1661–1669. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shi X, Wang X and Hua Y: lncRNA GACAT1
promotes gastric cancer cell growth, invasion and migration by
regulating miR-149-mediated of ZBTB2 and SP1. J Cancer.
9:3715–3722. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lajos R, Braicu C, Jurj A, Chira S,
Cojocneanu-Petric R, Pileczki V and Berindan-Neagoe I: A miRNAs
profile evolution of triple negative breast cancer cells in the
presence of a possible adjuvant therapy and senescence inducer. J
BUON. 23:692–705. 2018.PubMed/NCBI
|
|
14
|
Takamizawa J, Konishi H, Yanagisawa K,
Tomida S, Osada H, Endoh H, Harano T, Yatabe Y, Nagino M, Nimura Y,
et al: Reduced expression of the let-7 microRNAs in human lung
cancers in association with shortened postoperative survival.
Cancer Res. 64:3753–3756. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cimmino A, Calin GA, Fabbri M, Iorio MV,
Ferracin M, Shimizu M, Wojcik SE, Aqeilan RI, Zupo S, Dono M, et
al: miR-15 and miR-16 induce apoptosis by targeting BCL2. Proc Natl
Acad Sci USA. 102:13944–13949. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Salmena L, Poliseno L, Tay Y, Kats L and
Pandolfi PP: A ceRNA hypothesis: The Rosetta Stone of a hidden RNA
language? Cell. 146:353–358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rui X, Xu Y, Jiang X, Ye W, Huang Y and
Jiang J: Long non-coding RNA C5orf66-AS1 promotes cell
proliferation in cervical cancer by targeting miR-637/RING1 axis.
Cell Death Dis. 9:11752018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang T, Cai X, Li Q, Xue P, Chen Z, Dong
X and Xue Y: Hsa-miR-875-5p exerts tumor suppressor function
through downregulation of EGFR in colorectal carcinoma (CRC).
Oncotarget. 7:42225–42240. 2016.PubMed/NCBI
|
|
19
|
Wang J, Lu Y, Ding H, Gu T, Gong C, Sun J,
Zhang Z, Zhao Y and Ma C: The miR-875-5p inhibits SATB2 to promote
the invasion of lung cancer cells. Gene. 644:13–19. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Luan Z, Zhang Y, Lu T, Ruan Y, Zhang H,
Yan J, Li L, Sun W, Wang L, Yue W, et al: Positive association of
the human STON2 gene with schizophrenia. Neuroreport. 22:288–293.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kaempf N, Kochlamazashvili G, Puchkov D,
Maritzen T, Bajjalieh SM, Kononenko NL and Haucke V: Overlapping
functions of stonin 2 and SV2 in sorting of the calcium sensor
synaptotagmin 1 to synaptic vesicles. Proc Natl Acad Sci USA.
112:7297–7302. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sun X, Zhang W, Li H, Niu C, Ou Y, Song L
and Zhang Y: Stonin 2 overexpression is correlated with unfavorable
prognosis and tumor invasion in epithelial ovarian cancer. Int J
Mol Sci. 18:16532017. View Article : Google Scholar
|