Introduction
Lung cancer is the most prevalent human cancer and
remains the main cause of cancer-associated mortality, with a
5-year survival rate of 15% (1,2). In
addition, non-small cell lung cancer (NSCLC) subtype accounts for
~80% of lung cancer cases and comprises adenocarcinoma and squamous
cell carcinoma. Patients with NSCLC have a 5-year survival rate of
<15%, which is due to numerous factors, including difficulties
in early diagnosis, frequent relapse and absence of effective
treatments for advanced cases (3).
It is therefore crucial to determine the underlying mechanisms of
NSCLC onset and progression and develop novel therapeutic
strategies.
Interleukin-9 (IL-9) is a T helper (Th) 2 cytokine
that contributes to allergic diseases, including asthma and
rhinitis (4). A previous study
demonstrated that IL-9 is involved in tumor immunity mediated by
regulatory T cells (Tregs) and mast cells (5). Increasing evidence indicates that IL-9
participates in the pathogenesis of different types of cancer, such
as lung cancer, breast cancer and gastric cancer, predominantly
acting as a cancer promoting factor, particularly in nonsolid
tumors (6–8). Signal transducer and activator of
transcription 3 (STAT3) is a latent cytoplasmic transcription
factor, originally discovered as a transducer of signals from cell
surface receptors to the nucleus (9). Numerous evidences suggest that STAT3 is
constitutively activated in various types of cancer and serve a
crucial role in tumor growth and metastasis (9–12). In
addition, STAT3 regulates cellular proliferation, invasion,
migration and angiogenesis, which are essential for cancer
metastasis (13).
Epithelial-to-mesenchymal-transition (EMT) is a fundamental
biological process in which epithelial cells undergo biochemical
shifts to acquire mesenchymal properties (14). It is known that during EMT,
epithelial cells gain a mesenchymal phenotype, resulting in
increased invasion and metastasis in cancer (15). Consequently, epithelial markers, such
as E-cadherin, are downregulated, while mesenchymal markers, such
as vimentin and N-cadherin are upregulated (16). Accumulating evidence has elucidated
the essential role of EMT in the progression of NSCLC (11–13).
MicroRNAs (miRs) are evolutionary conserved
non-coding RNA molecules composed of 20–24 ribonucleotides, which
bind to mRNA 3′ untranslated regions (3′-UTRs), activating their
degradation or impairing the translation process (17–19).
Numerous studies reported that dysregulated expression of certain
miRNAs is associated with the development of esophageal (20,21),
hepatocellular (22) and breast
(23) carcinomas in humans. The
miR-208 family includes miR-208a, miR-208b and miR-449 (24–26).
Deep miRNA sequencing demonstrated that numerous members of the
miR-208 family are involved in the onset and progression of cardiac
diseases, such as cardiac ischemia reperfusion injury (24) and acute myocardial infarction
(25).
In particular, it was reported that the expression
level of −3p and −5p isoforms of miR-208a and miR-208b is highly
expressed in cardiac tissue and is dysregulated in various
cardiovascular diseases (26–30).
Inhibition of miR-208 improves cardiac function and patient
survival during heart failure (30,31).
However, the expression and function of miR-208 in different types
of cancer, particularly in NSCLC remain unknown.
The present study aimed to determine the function
and underlying molecular mechanisms of miR-208b-5p in the
progression of NSCLC.
Materials and methods
Tissue specimens
A total of 62 tumor samples were isolated from
patients with NSCLC who underwent surgical lung resection at the
Zhejiang Provincial Zhongshan Hospital between January 2011 and
December 2013. Patients had not been treated by radiotherapy or
chemotherapy prior to the surgery. Tumor tissues were collected
from the edge of NSCLC lesions. Adjacent normal tissues were also
collected 5 cm from the lesion. NSCLC and adjacent non-tumor
tissues were confirmed by at least two expert pathologists. All
patients with NSCLC received checkup every 2–3 months during the
first two years and every 3–6 months afterwards until the end of
the follow-up period in January 2015 by two physicians who were
blinded to the study. Samples were snap-frozen after resection and
maintained at −80°C prior to use. This study was approved by the
Ethics Committee of The First Hospital of Longyan city, Fujian
Medical University. All patients provided written informed consent
prior to the study. The clinicopathological characteristics of the
patients included in the present study are presented in Table I.
 | Table I.Association between miR-208b-5p
expression level and the clinicopathological characteristic of
patients with non-small cell lung cancer. |
Table I.
Association between miR-208b-5p
expression level and the clinicopathological characteristic of
patients with non-small cell lung cancer.
|
|
| miR-208b-5p
expression |
|
|---|
|
|
|
|
|
|---|
| Characteristic | Patient, n | High n=30 | Low n=32 | P-value |
|---|
| Sex |
|
|
| 0.075 |
|
Male | 51 | 22 | 29 |
|
|
Female | 11 | 8 | 3 |
|
| Age, years |
|
|
| 0.553 |
|
<60 | 39 | 20 | 19 |
|
|
≥60 | 23 | 10 | 13 |
|
| Smoking status |
|
|
| 0.851 |
|
Previous/Current | 40 | 19 | 21 |
|
|
Never | 22 | 11 | 11 |
|
| Pathological
type |
|
|
| 0.779 |
|
Adenocarcinoma | 28 | 13 | 15 |
|
|
Squamous carcinoma | 34 | 17 | 17 |
|
| Tumor
differentiation |
|
|
| 0.286 |
| Well
and moderate | 49 | 22 | 27 |
|
|
Poor | 13 | 8 | 5 |
|
| Tumor size, cm |
|
|
| 0.830 |
|
<3 | 51 | 25 | 26 |
|
| ≥3 | 11 | 5 | 6 |
|
| Lymphatic
metastasis |
|
|
| 0.014 |
|
Yes | 22 | 6 | 16 |
|
| No | 40 | 24 | 16 |
|
| TNM
classification |
|
|
| 0.021 |
|
I–II | 22 | 15 | 7 |
|
|
III–IV | 40 | 15 | 25 |
|
Cell lines and transfection
The eight human NSCLC cell lines H460, PC9, H1299,
A549, H1703, H1437, CALU-1 and H520, the 293T cell line and the
normal HBEC Beas-2b cell line were purchased from The Cell Bank of
Type Culture Collection of the Chinese Academy of Sciences. Cells
were cultured in RPMI-1640 (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.) and 1% penicillin/streptomycin (Gibco; Thermo
Fisher Scientific, Inc.) and placed at 37°C in a humidified
incubator containing 5% CO2.
miR-208b-5p mimics (miR-208b-5p) and control mimics
(control) were purchased from Sigma-Aldrich; Merck KGaA. A549 and
H1299 cells (1×105) were co-transfected with IL-9 cDNA
plasmid (without the 3′UTR; Shanghai GenePharma Co., Ltd.), an
empty vector (pcDNA-NC; Shanghai GenePharma Co., Ltd.) was used as
the control for IL-9 overexpression experiments and miR-208b-5p
mimics for 72 h. The primer sequences were as follows: miR-208b-5p
mimic forward, 5′-GUGCAUUAUGGUUGCAUCCCA-3′ and reverse,
5′-CAAUGCAACUACAAUGCACUU-3′; and NC mimic forward,
5′-UUCUCCGAACGUGUCACGUTT-3′ and reverse,
5′-ACGUGACACGUUCGGAGAATT-3′. miR-208b-5p mimic transfections were
performed using Lipofectamine™ 3000 reagent (Invitrogen; Thermo
Fisher Scientific, Inc.), according to the manufacturer's
protocol.
C188-9 is a small-molecule inhibitor of STAT3 that
targets the phosphotyrosyl peptide binding site within the STAT3
Src homology 2 (SH2) domain, with Ki=136 nm (9). It does not inhibit upstream Jak or Src
kinases (9). In order to define the
role of miR-208b-5p inhibition NSCLC cells EMT through IL-9/STAT3
signaling pathway, A549 and H1299 cells were pre-treated with
C188-9 (0.9 µM; Selleck Chemicals) for 1 h (9) and subsequently transfected with
miR-208b-5p mimics or control mimics for 48 h. Cells without C188-9
treatment were used as the control.
Reverse transcription-quantitative
(RT-qPCR)
Total RNA was extracted from NSCLC cell lines and 62
NSCLC tissues using TRIzol™ (Invitrogen; Thermo Fisher Scientific,
Inc.). cDNA was generated with the Mir-X™ miRNA First-Strand
Synthesis kit (Takara Bio, Inc.) for miRNA expression level
analysis or using PrimeScript RT Reagent Kit (Takara Bio, Inc.) for
mRNA expression level analysis according to the manufacturers'
instructions. RT-qPCR was performed using SYBR Premix Ex Taq II kit
(Takara Bio, Inc.) on a ViiA 7 RT-PCR instrument (Applied
Biosystems; Thermo Fisher Scientific, Inc.) as described by the
manufacturers. RT-qPCR reactions were performed as follows: 30
cycles of denaturation at 94°C for 30 sec, annealing at 56°C for 30
sec and extension at 72°C for 30 sec. U6 or β-actin were used as an
internal controls. The primers were designed by Shanghai Sangong
Pharmaceutical Co., Ltd., and are as follows: miR-208b-5p forward,
5′-GTCGTATCCAGTGCGTGTCGTC-3′ and reverse,
5′-CACACTCTTGATGTTCCAGGA-3′; U6 forward, 5′-CTCGCTTCGGCAGCACA-3′
and reverse, 5′-AACGCTTCACGAATTTGCGT-3′; IL-9 forward,
5′-CTCTGTTTGGGCATTCCCTCT-3′ and reverse,
5′-GGGTATCTTGTTTGCATGGTGG-3′; β-actin forward,
5′-TGACGTGGACATCCGCAAAG-3′ and reverse, 5′-CTGGAAGGTGGACAGCGAGG-3′;
E-cadherin forward, 5′-TACACTGCCCAGGAGCCAGA-3′ and reverse,
5′-TGGCACCAGTGTCCGGATTA-3′; N-cadherin forward,
5′-TCAGGCGTCTGTAGAGGCTT-3′ and reverse,
5′-ATGCACATCCTTCGATAAGACTG-3′; and vimentin forward,
5′-GACGCCATCAACACCGAGTT-3′ and reverse,
5′-CTTTGTCGTTGGTTAGCTGGT-3′. The relative expressions levels were
normalized to endogenous controls and were expressed as
2−ΔΔCq (32).
In vitro cell migration and
invasion
NSCLC cell migration and invasion were investigated
via Transwell assays. For the migration assay, 2000 cells in
FBS-free medium were seeded in the upper compartment of the
Transwell (pore size, 8 µm; Corning Inc.). For the invasion assay,
the chamber was precoated with Matrigel (Corning Inc.) for 48 h at
37°C prior to cell seeding (2,000 cells in FBS-free medium). The
lower chamber was filled with medium supplemented with 20% FBS.
After 24 h incubation, non-migratory cells were swabbed and
removed. Cells that have migrated were fixed with 4%
paraformaldehyde for 30 min at 25°C followed by crystal violet
staining for 30 min at 25°C. The values for invasion or migration
were obtained by counting 10 randomly selected fields per membrane
using a light microscope (magnification, ×40; Olympus Corporation)
and represented the average of 3 independent experiments.
Western blotting
Proteins were extracted from A549 and H1299 NSCLC
cells on ice using RIPA buffer (Beyotime Institute of
Biotechnology) supplemented with protease inhibitors (Cell
Signaling Technology, Inc.) for 20 min. Protein concentration was
measured using Bradford Protein Assay kit (Beyotime Institute of
Biotechnology). Proteins (15 µg) were separated via SDS-PAGE on a
10% gel and transferred onto polyvinylidene fluoride membranes.
Membranes were blocked with 5% non-fat milk dissolved in TBST for 1
h at room temperature and incubated with primary antibodies against
E-cadherin (cat. no. ab194982; 1:1,000), N-cadherin (cat. no.
ab202030; 1:1,000), vimentin (cat. no. ab193555; 1:1,000), IL-9
(cat. no. ab203386; 1:1,000), STAT3 (cat. no. ab68153; 1:1,000),
phospho-STAT3 (cat. no. ab76315; 1:1,000) and β-actin (cat. no.
ab179467; 1:5,000), which were all purchased from Abcam, overnight
at 4°C. Subsequently, membranes were incubated with goat
anti-rabbit horseradish peroxidase-conjugated secondary antibody
(cat. no. ab150077; 1:10,000) for 1 h at room temperature. Enhanced
chemiluminescence reagent (Beyotime Institute of Biotechnology) was
used to detect the signal on the membrane. The data were analyzed
via densitometry using Bio-Rad Image Lab software (version 4.1;
Bio-Rad Laboratories, Inc.) and normalized to the expression of the
internal control, β-actin.
Luciferase reporter assay
To perform luciferase reporter assay, a pMIR-REPORT
vector (Promega Corporation) was constructed. The 3′-UTR fragments
of IL-9 mRNA, containing miR-208b-5p candidate binding sites
synthesized by Shanghai Shandong Biology Engineering Technology
Service, Ltd., was PCR-amplified and inserted into the vectors,
referred to as the pMIR-IL-9-WT vectors (Promega Corporation).
Furthermore, pMIR-IL-9-Mut vectors (Promega Corporation) were built
with IL-9 subjected to site-directed mutation of the miR-208b-5p
binding sequence via a Quik-Change Site-directed Mutagenesis Kit
(Agilent Technologies, Inc.). Assays were performed in 96-well
plates with 1.5×104 293T cells per well. Cells were
incubated at 37°C for 24 h, followed by miR-208b-5p or negative
control co-transfection using Effectene reagent (Qiagen China Co.,
Ltd.), with pMIR-target-WT or pMIR-target-Mut vectors for 48 h.
Subsequently, luciferase activity was analyzed using the
Dual-Luciferase Reporter Assay System (Promega Corporation)
IL-9 ELISA
Following A549 and H1299 NSCLC cell transfection
with miR-208b-5p mimic or mimic NC for 48 h, supernatant was
collected and IL-9 level was measured by using the Human IL-9 ELISA
Kit (cat. no. ab46027; Abcam) according to the manufacturers'
instructions (detection limit, 0.5 pg/ml).
Statistical analysis
The data were presented as the means ± standard
deviation. Significant differences were determined using GraphPad
software version 5.0 (GraphPad Software, Inc.). Comparison between
NSCLC and adjacent normal tissues was performed using paired
Student's t-test. Student's t-test was used to determine
differences between two groups. One-way analysis of variance,
followed by Tukey's post-hoc test was used to compare differences
between three or more groups. χ2 test was used to
determine the association between miR-208b-5p expression level and
the clinicopathological characteristics of patients with NSCLC.
Survival curves were plotted using Kaplan-Meier method and were
compared via log-rank test. P<0.05 was considered to indicate a
statistically significant difference. Each experiment was repeated
three times.
Results
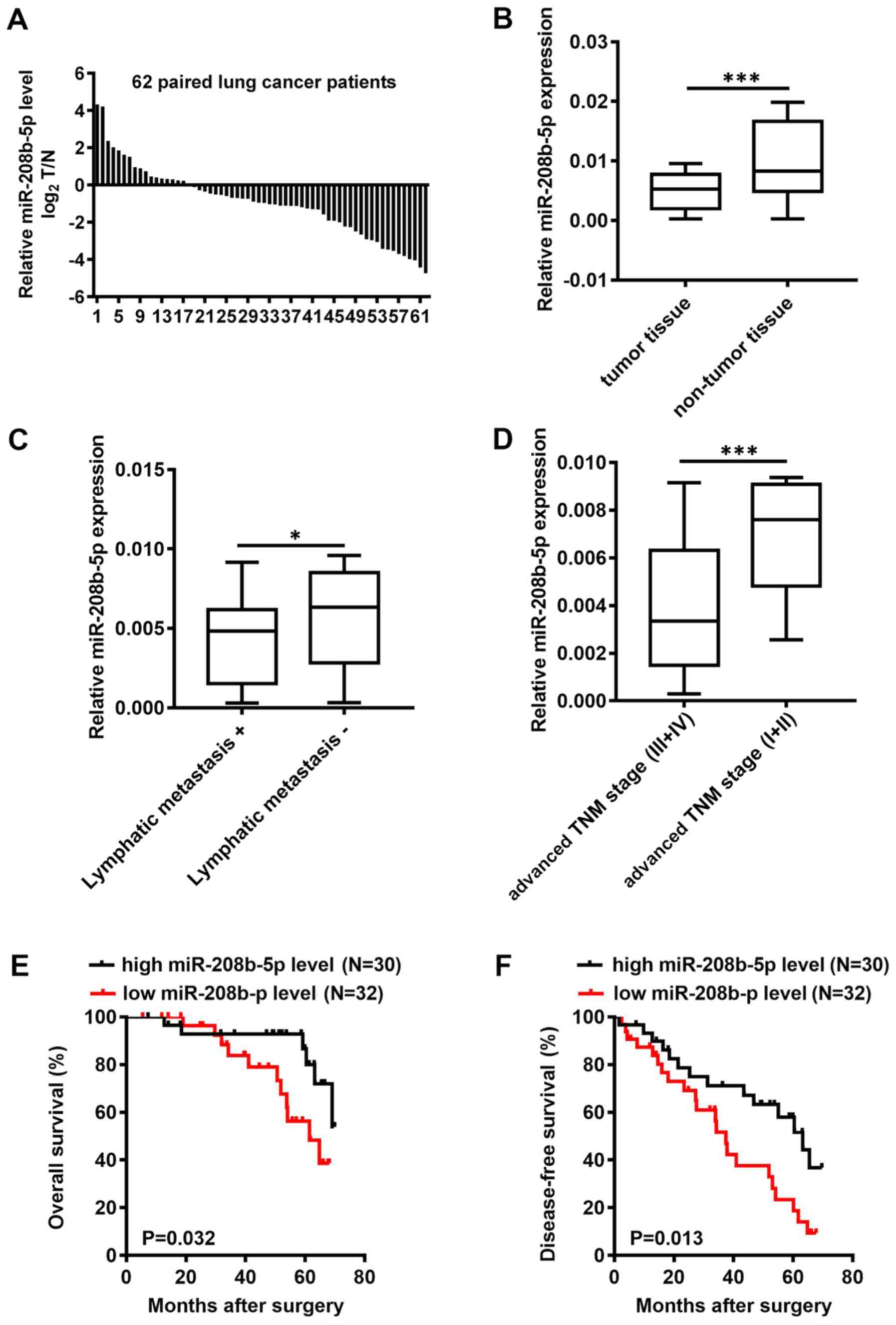
miR-208b-5p is downregulated in NSCLC
tissues and inversely associated with prognosis
miR-208b-5p expression level was assessed by RT-qPCR
in 62 pairs of NSCLC and adjacent normal tissues. The results
demonstrated that miR-208b-5p expression level in tumor tissues was
decreased in tumor tissues of 45 patients with NSCLC compared with
adjacent normal tissues with 45 out of 62 patients have a lower
miR-208b-5p level in NSCLC tumors compared to adjacent normal
tissues (P<0.001; Fig. 1A and B).
Based on the mean expression of miR-208b-5p in NSCLC tissues
(cut-off value, 0.0057), patients were divided into a high
miR-208b-5p expression group and a low miR-208b-5p expression
group. In addition, miR-208b-5p expression level in NSCLC tissues
was inversely associated with the presence of lymphatic metastasis
and clinical TNM stage (Table I;
P=0.014 and P=0.021, respectively). Patients with lymphatic
metastasis or advanced TNM stage presented with decreased
miR-208b-5p expression level (Fig. 1C
and D; P<0.05 and P<0.0001, respectively). Furthermore,
patients with low miR-208b-5p expression levels had significantly
shorter overall survival (OS) and disease-free survival (DFS)
compared with patients with high miR-208b-5p expression levels
(Fig. 1E and F; P=0.032 and P=0.013,
respectively). These findings suggested that miR-208b-5p
downregulation in NSCLC tissues was inversely associated with
lymphatic metastasis and clinical TNM stage. In addition, the
results from OS and DFS analyses suggested that miR-208b-5p
expression level may be considered as an independent prognostic
factor in patients with NSCLC.
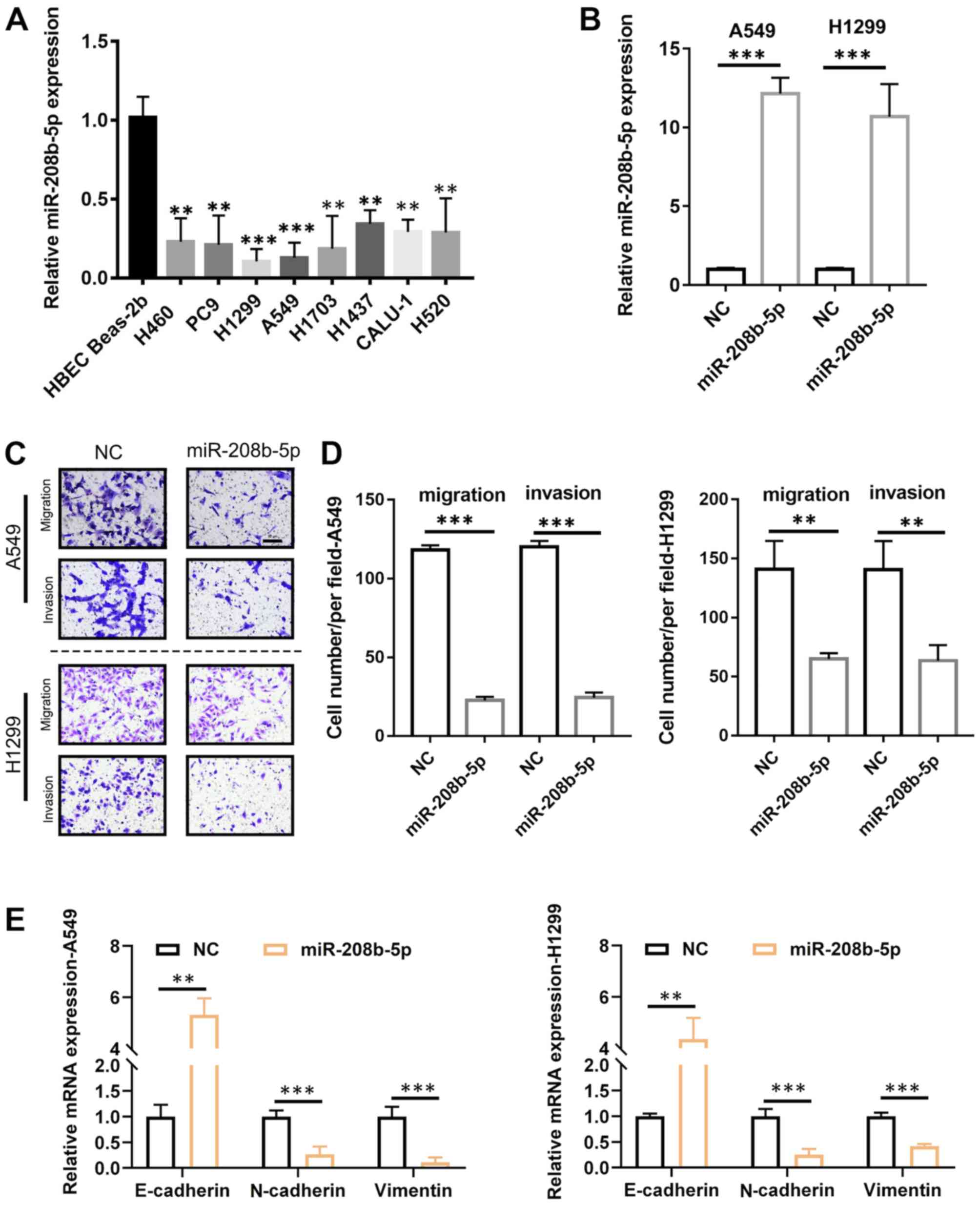
miR-208b-5p inhibits the migratory and
invasive abilities and the EMT in NSCLC cells
Following the determination of the association
between miR-208b-5p expression level and metastasis in patients
with NSCLC, the underlying mechanisms were evaluated. To do so,
miR-208b-5p expression level in numerous NSCLC-derived cell lines
was measured. The results demonstrated that miR-208b-5p expression
level was significantly decreased in these cell lines compared with
the normal Beas-2b bronchial cell line (Fig. 2A; P<0.05 and P<0.01). The cell
lines A549 and H1299 were selected for subsequent analyses, since
they were characterized by the lowest miR-208b-5p levels. To
overexpress miR-208b-5p in these two cell lines, infection with a
lentiviral vector was performed, and successful transfection was
confirmed by RT-qPCR (Fig. 2B). The
results from Transwell assays demonstrated that A549 and H1299
cells overexpressing miR-208b-5p had significantly decreased
migratory and invasive abilities compared with control (Fig. 2C and D; P<0.001 for A549 cells and
P<0.01 for H1299 cells). Furthermore, miR-208b-5p-overexpressing
cells exhibited significantly increased mRNA expression of the
epithelial marker E-cadherin and decreased mRNA expression of the
mesenchymal markers N-cadherin and vimentin (P<0.01 and
P<0.001; Fig. 2E). These findings
suggested that miR-208b-5p may inhibit EMT and decrease NSCLC cell
migratory and invasive abilities.
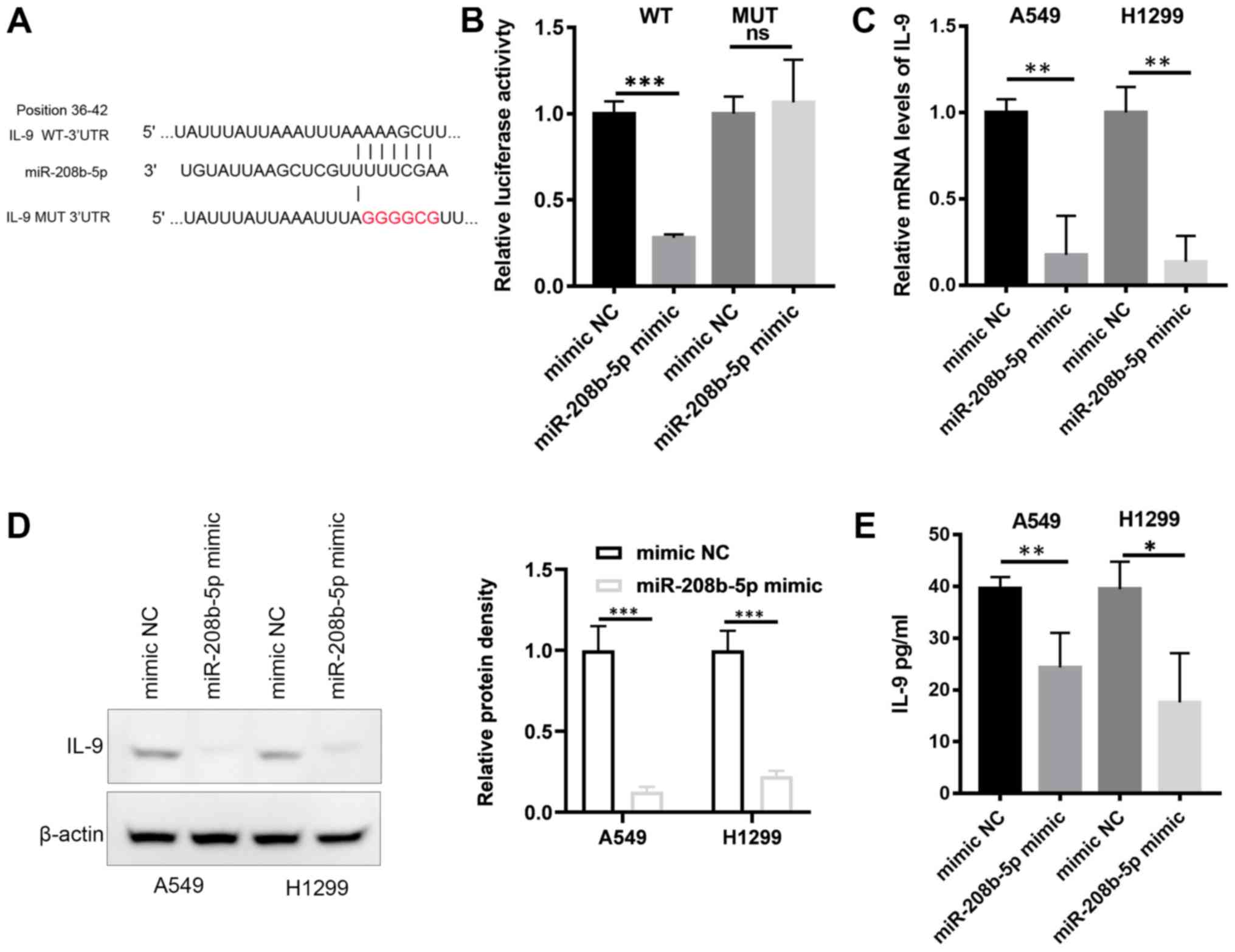
miR-208b-5p targets IL-9 in NSCLC
cells
Analysis of the miRNA target database reported IL-9
as a potential target of miR-208b-5p (Fig. 3A). To analyze the interaction between
miR-208b-5p and IL-9 mRNA, a wild type (WT) luciferase reporter was
constructed including the 3′-UTR of IL-9. A mutant type (Mut)
reporter with a mutated miR-208b-5p binding sequence was also
generated (Fig. 3A). The constructs
and miR-208b-5p were co-transfected into the 293T cell line.
Compared with control cells (mimic NC), cells transfected with
miR-208b-5p and the WT constructs displayed a significantly
decreased luciferase activity (Fig.
3B; P<0.001). This effect was absent in cells transfected
with the Mut reporter. These findings suggested a direct binding of
miR-208b-5p to the 3′-UTR of IL-9. This result was confirmed by
RT-qPCR, western blotting and ELISA assays, which demonstrated
decrease in mRNA and protein levels of IL-9 in A549 and H1299 cells
overexpressing miR-208b-5p (Fig.
3C-E, P<0.05). IL-9 mRNA may therefore be directly targeted
by miR-208b-5p.
IL-9/STAT3 signaling pathway mediates
the inhibition of EMT in NSCLC cells by miR-208b-5p
To further elucidate the interrelationship among
miR-208b-5p expression level, IL-9 signaling and EMT, RT-qPCR
analyses were performed. A549 and H1299 NSCLC cells were
transfected with pcDNA.IL9, and the results demonstrated increased
IL9 mRNA expression levels in these cells compared with the
negative control (pcDNA.NC; P<0.05; Fig. 4A and B). Furthermore, miR-208b-5p
overexpression increased E-cadherin mRNA expression, while
decreasing N-Cadherin and vimentin mRNA expression levels in A549
and H1299 NSCLC cells (P<0.05; Fig.
4C and D). Conversely, increasing IL-9 levels decreased
E-cadherin mRNA expression, while increasing N-Cadherin and
vimentin mRNA expression levels in A549 and H1299 NSCLC cells
(P<0.05; Fig. 4C and D).
Furthermore, co-transfection with miR-208b-5p mimic + IL9 reversed
the effects of miR-208b-5p overexpression on increasing E-Cadherin
mRNA expression level, and decreasing N-Cadherin and vimentin mRNA
expression levels in A549 and H1299 NSCLC cells (P<0.05;
Fig. 4C and D). The STAT3 inhibitor,
C188-9 attenuated the expression level of EMT-promoting proteins.
In addition, miR-208b-5p overexpression had no further effect on
the expression level of these proteins when combined with C188-9
treatment (Fig. 4E and F,
P<0.05). These findings suggested that miR-208b-5p may inhibit
EMT in NSCLC cells via STAT3-mediated decreased IL-9
expression.
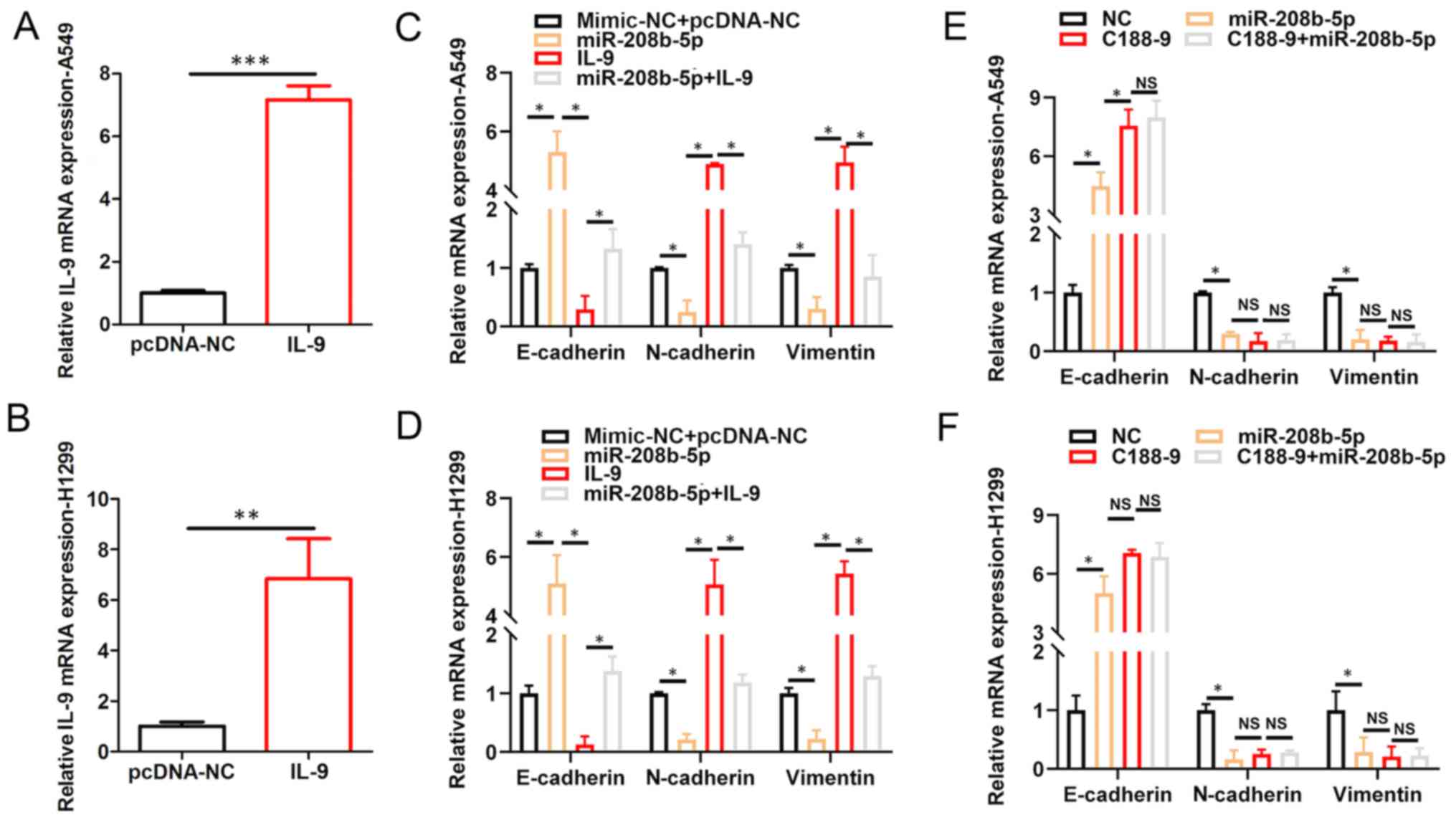 | Figure 4.IL-9/STAT3 signaling pathway mediates
the inhibition of EMT in NSCLC cells by miR-208b-5p. IL9 mRNA
expression in (A) A549 and (B) H1299 cells transfected with IL9.
Relative mRNA expression of E-cadherin, N-cadherin and vimentin
assessed via RT-qPCR analysis in (C) A459 and (D) H1299 cells
transfected with NC, miR-208b-5p, IL-9 or miR-208b-5p+IL-9.
Relative mRNA expression of N-cadherin, E-cadherin and vimentin
assessed by RT-qPCR in (E) A459 and (F) H1299 cells transfected or
treated with NC, miR-208b-5p, C188-9 or C188-9+miR-208b-5p.
*P<0.05, **P<0.01 and ***P<0.001. miR, microRNA, IL-9,
interleukin 9; NC, negative control; NS, not significant; RT-qPCR,
reverse transcription-quantitative PCR. |
Discussion
Increasing evidence indicates that several types of
miRNAs, such as miR-675-5p, miR-361-3p, miR-4286, miR-150-5p
miR-625 and miR-4319 play crucial roles in NSCLC regulation
(33–38). miR-208 is highly expressed in cardiac
tissue and is dysregulated in several types of cardiovascular
diseases (26–30), such as cardiac ischemia reperfusion
injury (24) and acute myocardial
infarction (25). Inhibition of
miR-208 has been demonstrated to improve cardiac function and
patient survival during heart failure (30,31).
However, the expression and function of miR-208 in different types
of cancer, particularly in NSCLC remain unknown. Thus, the present
study aimed to investigate the potential role of miR-208b-5p in
inhibiting NSCLC cell migration and invasion.
The present study demonstrated that miR-208b-5p
expression level in NSCLC tissues was significantly decreased
compared with adjacent normal tissues. Furthermore, miR-208b-5p
expression level was associated with the presence of metastases,
clinical TNM stage, OS and DFS. miR-208b-5p overexpression
attenuated the migratory and invasive abilities of NSCLC cells and
inhibited EMT. In addition, this study demonstrated that IL-9 mRNA
may be a novel target for miR-208b-5p. Furthermore, miR-208b-5p
ability to attenuate the STAT3 pathway was demonstrated.
miR-208b-5p may therefore be considered as a cancer suppressor gene
and may be used as a therapeutic target and prognostic marker in
NSCLC.
The present study demonstrated that the mRNA coding
IL-9 protein may be a novel direct target of miR-208b-5p. IL-9
belongs to the Th2-type of cytokines that have a pleiotropic effect
on inflammatory responses in allergic diseases (4). A previous study reported the
involvement of IL-9 in Treg and mast cells-mediated tumor immunity
(5). Increasing evidence indicates
that IL-9 participates in the pathogenesis of several types of
cancer, predominantly acting as a cancer promoting factor,
particularly in nonsolid tumors (6–8). IL-9
contributes to the signaling pathways of IL-2, IL-4, IL-7, IL-15
and IL-21, which involve heterodimeric receptors that interact with
the common γ-chain members of the STATs family (39). STAT3 transduces signals from the cell
membrane receptors to the nucleus, where it acts as a transcription
factor. STAT3 is known to be constitutively activated in various
types of cancer (9), such as
cutaneous melanoma (10), squamous
cell carcinoma (11) and oral
squamous cell carcinoma (12). In
addition to promoting angiogenesis, STAT3 regulates cell
proliferation, migration and invasion of tumor cells, indicating
its crucial role in the control of tumor growth and metastasis
(10,40–42). The
present study demonstrated that STAT3 inhibitor C188-9 attenuated
the expression of EMT-promoting mRNA levels. Furthermore,
miR-208b-5p overexpression did not further alter the mRNA levels of
related-EMT genes in NSCLC cells treated with C188-9. Taken
together, these findings suggested that miR-208b-5p may exert a
tumor suppressor role in NSCLC cells by regulating IL-9/STAT3
pathway. Further investigation is required to confirm this
hypothesis and determine the underlying mechanisms of miR-208-5p in
NSCLC.
In conclusion, the present study demonstrated that
miR-208b-5p was downregulated in NSCLC, and that its expression
level was associated with metastasis and patient survival. These
results suggested that miR-497-5p may act as a potential tumor
suppressor in NSCLC. In addition, miR-208b-5p may prevent the
activation of EMT, and decrease the migratory and invasive
abilities of NSCLC cells by targeting IL-9/ STAT3 pathway.
Acknowledgements
Not applicable.
Funding
This study was supported by the Special Fund for
Scientific Research in Non-affiliated Hospitals of Fujian Medical
University (grant. no. FZS13013Y) and the Youth Research Fund of
Fujian Provincial Department of Health (grant. no. 2013-2-150).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
JM and HT wrote the manuscript and contributed to
the conception of the study. JL and FC performed the data analysis.
CW, CC and JW contributed to data acquisition and analysis and
revised the manuscript. SH performed the clinic patients' data
analysis.. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
This study was approved by the Institutional Ethics
Committee of the First Hospital of Longyan city, Fujian Medical
University (Longyan, China; approval no. 2018-LSKY-22. Signed
informed consents were obtained from the patients or their
guardians.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
3′-UTRs
|
3′ untranslated regions
|
|
EMT
|
epithelial-mesenchymal transition
|
|
IL-9
|
interleukin-9
|
|
microRNA-208b-5p
|
miR-208b-5p
|
|
NSCLC
|
non-small cell lung cancer
|
|
TNM
|
Tumor-Node-Metastasis
|
References
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Heist RS and Engelman JA: SnapShot:
Non-small cell lung cancer. Cancer Cell. 21:448.e22012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Soroosh P and Doherty TA: Th9 and allergic
disease. Immunology. 127:450–458. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tete S, Saggini A, Maccauro G, Rosati M,
Conti F, Cianchetti E, Tripodi D, Toniato E, Fulcheri M, Salini V,
et al: Interleukin-9 and mast cells. J Biol Regul Homeost Agents.
26:319–326. 2012.PubMed/NCBI
|
|
6
|
Knoops L and Renauld JC: IL-9 and its
receptor: From signal transduction to tumorigenesis. Growth
Factors. 22:207–215. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen N, Lu K, Li P, Lv X and Wang X:
Overexpression of IL-9 induced by STAT6 activation promotes the
pathogenesis of chronic lymphocytic leukemia. Int J Clin Exp
Pathol. 7:2319–2323. 2014.PubMed/NCBI
|
|
8
|
Zhang J, Wang WD, Geng QR, Wang L, Chen
XQ, Liu CC and Lv Y: Serum levels of interleukin-9 correlate with
negative prognostic factors in extranodal NK/T-cell lymphoma. PLoS
One. 9:e946372014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Redell MS, Ruiz MJ, Alonzo TA, Gerbing RB
and Tweardy DJ: Stat3 signaling in acute myeloid leukemia:
Ligand-dependent and -independent activation and induction of
apoptosis by a novel small-molecule Stat3 inhibitor. Blood.
117:5701–5709. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
He Y, Yang Y, Xu J, Liao Y, Liu L, Deng L
and Xiong X: IL22 drives cutaneous melanoma cell proliferation,
migration and invasion through activation of miR-181/STAT3/AKT
axis. J Cancer. 11:2679–2687. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zheng S, Liu Q, Liu T, Yang L, Zhang Q,
Shen T, Zhang X, Han X and Lu X: NME4 modulates PD-L1 expression
via the STAT3 signaling pathway in squamous cell carcinoma. Biochem
Biophys Res Commun. S0006-291X(30535-0)2020.[Epub ahead of
print].
|
|
12
|
Qu Y, He Y, Yang Y, Li S, An W, Li Z, Wang
X, Han Z and Qin L: ALDH3A1 acts as a prognostic biomarker and
inhibits the epithelial mesenchymal transition of oral squamous
cell carcinoma through IL-6/STAT3 signaling pathway. J Cancer.
11:2621–2631. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fionda C, Malgarini G, Soriani A, Zingoni
A, Cecere F, Iannitto ML, Ricciardi MR, Federico V, Petrucci MT,
Santoni A and Cippitelli M: Inhibition of glycogen synthase
kinase-3 increases NKG2D ligand MICA expression and sensitivity to
NK cell-mediated cytotoxicity in multiple myeloma cells: Role of
STAT3. J Immunol. 190:6662–6672. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li W, Jia G, Qu Y, Du Q and Liu B and Liu
B: Long non-coding RNA (LncRNA) HOXA11-AS promotes breast cancer
invasion and metastasis by regulating epithelial-mesenchymal
transition. Med Sci Monit. 23:3393–3403. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhao M, Ang L, Huang J and Wang J:
MicroRNAs regulate the epithelial-mesenchymal transition and
influence breast cancer invasion and metastasis. Tumour Biol.
39:10104283176916822017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Banyard J and Bielenberg DR: The role of
EMT and MET in cancer dissemination. Connect Tissue Res.
56:403–413. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Croce CM: Causes and consequences of
microRNA dysregulation in cancer. Nat Rev Genet. 10:704–714. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ma J, Chen X, Xin G and Li X: Chronic
exposure to the ionic liquid [C8mim]Br induces
inflammation in silver carp spleen: Involvement of oxidative
stress-mediated p38MAPK/NF-κB signalling and microRNAs. Fish
Shellfish Immunol. 84:627–638. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou YW, Zhang H, Duan CJ, Gao Y, Cheng
YD, He D, Li R and Zhang CF: miR-675-5p enhances tumorigenesis and
metastasis of esophageal squamous cell carcinoma by targeting
REPS2. Oncotarget. 7:30730–30747. 2016.PubMed/NCBI
|
|
21
|
Wen J, Hu Y, Liu Q, Ling Y, Zhang S, Luo
K, Xie X, Fu J and Yang H: miR-424 coordinates multilayered
regulation of cell cycle progression to promote esophageal squamous
cell carcinoma cell proliferation. EBioMedicine. 37:110–124. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chang RM, Xiao S, Lei X, Yang H, Fang F
and Yang LY: miRNA-487a promotes proliferation and metastasis in
hepatocellular carcinoma. Clin Cancer Res. 23:2593–2604. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Song H, Li D, Wu T, Xie D, Hua K, Hu J,
Deng X, Ji C, Deng Y and Fang L: MicroRNA-301b promotes cell
proliferation and apoptosis resistance in triple-negative breast
cancer by targeting CYLD. BMB Rep. 51:602–607. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu C, Zheng H, Xie L and Zhang J:
Decreased miR-208 induced ischemia myocardial and reperfusion
injury by targeting p21. Pharmazie. 71:719–723. 2016.PubMed/NCBI
|
|
25
|
Yan X, Liu J, Wu H, Liu Y, Zheng S, Zhang
C and Yang C: Impact of miR-208 and its target gene nemo-like
kinase on the protective effect of ginsenoside Rb1 in
hypoxia/ischemia injuried cardiomyocytes. Cell Physiol Biochem.
39:1187–1195. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kakimoto Y, Tanaka M, Kamiguchi H, Hayashi
H, Ochiai E and Osawa M: MicroRNA deep sequencing reveals
chamber-specific miR-208 family expression patterns in the human
heart. Int J Cardiol. 211:43–48. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhou C, Cui Q, Su G, Guo X, Liu X and
Zhang J: MicroRNA-208b alleviates post-infarction myocardial
fibrosis in a rat model by inhibiting GATA4. Med Sci Monit.
22:1808–1816. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cai B, Pan Z and Lu Y: The roles of
microRNAs in heart diseases: A novel important regulator. Curr Med
Chem. 17:407–411. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Han M, Toli J and Abdellatif M: MicroRNAs
in the cardiovascular system. Curr Opin Cardiol. 26:181–189. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Malizia AP and Wang DZ: MicroRNAs in
cardiomyocyte development. Wiley Interdiscip Rev Syst Biol Med.
3:183–190. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Montgomery RL, Hullinger TG, Semus HM,
Dickinson BA, Seto AG, Lynch JM, Stack C, Latimer PA, Olson EN and
van Rooij E: Therapeutic inhibition of miR-208a improves cardiac
function and survival during heart failure. Circulation.
124:1537–1547. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
He D, Wang J, Zhang C, Shan B, Deng X, Li
B, Zhou Y, Chen W, Hong J, Gao Y, et al: Down-regulation of
miR-675-5p contributes to tumor progression and development by
targeting pro-tumorigenic GPR55 in non-small cell lung cancer. Mol
Cancer. 14:732015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen W, Wang J, Liu S, Wang S, Cheng Y,
Zhou W, Duan C and Zhang C: MicroRNA-361-3p suppresses tumor cell
proliferation and metastasis by directly targeting SH2B1 in NSCLC.
J Exp Clin Cancer Res. 35:762016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
An X, Ge J, Guo H, Mi H, Zhou J, Liu Y,
Weiyue and Wu Z: Overexpression of miR-4286 is an unfavorable
prognostic marker in individuals with non-small cell lung cancer. J
Cell Biochem. 120:17573–17583. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dai FQ, Li CR, Fan XQ, Tan L, Wang RT and
Jin H: miR-150-5p inhibits non-small-cell lung cancer metastasis
and recurrence by targeting HMGA2 and β-catenin signaling. Mol Ther
Nucleic Acids. 16:675–685. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tan X, Jiang L, Wu X, Feng W and Lin Q:
MicroRNA-625 inhibits the progression of non-small cell lung cancer
by directly targeting HOXB5 and deactivating the Wnt/β-catenin
pathway. Int J Mol Med. 44:346–356. 2019.PubMed/NCBI
|
|
38
|
Yang Y, Li H, Liu Y, Chi C, Ni J and Lin
X: miR-4319 hinders YAP expression to restrain non-small cell lung
cancer growth through regulation of LIN28-mediated RFX5 stability.
Biomed Pharmacother. 115:1089562019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hornakova T, Staerk J, Royer Y, Flex E,
Tartaglia M, Constantinescu SN, Knoops L and Renauld JC: Acute
lymphoblastic leukemia-associated JAK1 mutants activate the Janus
kinase/STAT pathway via interleukin-9 receptor alpha homodimers. J
Biol Chem. 284:6773–6781. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chen N, Feng L, Qu H, Lu K, Li P, Lv X and
Wang X: Overexpression of IL-9 induced by STAT3 phosphorylation is
mediated by miR-155 and miR-21 in chronic lymphocytic leukemia.
Oncol Rep. 39:3064–3072. 2018.PubMed/NCBI
|
|
41
|
Pérez C, Mondéjar R, García-Díaz N,
Cereceda L, León A, Montes S, Durán Vian C, Pérez Paredes MG,
González-Morán A, Alegre de Miguel V, et al: Advanced-stage mycosis
fungoides. Role of the signal transducer and activator of
transcription 3, nuclear factor-κB and nuclear factor of activated
T cells pathways. Br J Dermatol. 182:147–155. 2020.PubMed/NCBI
|
|
42
|
Qin JJ, Yan L, Zhang J and Zhang WD: STAT3
as a potential therapeutic target in triple negative breast cancer:
A systematic review. J Exp Clin Cancer Res. 38:1952019. View Article : Google Scholar : PubMed/NCBI
|