Cancer is a major global public health burden, with
~21.6 million new cancer cases predicted for 2030 (1). In general, tumorigenesis is the process
that promotes the transformation of normal cells into invasive
cells, overcoming the constraints that usually limit proliferation
and survival. These alterations can give rise to a number of
potentially deleterious circumstances or vulnerabilities that can
be lethal to patients if left unchecked (2). Despite extensive research investigating
tumorigenesis, the precise underlying molecular mechanisms remain
unclear.
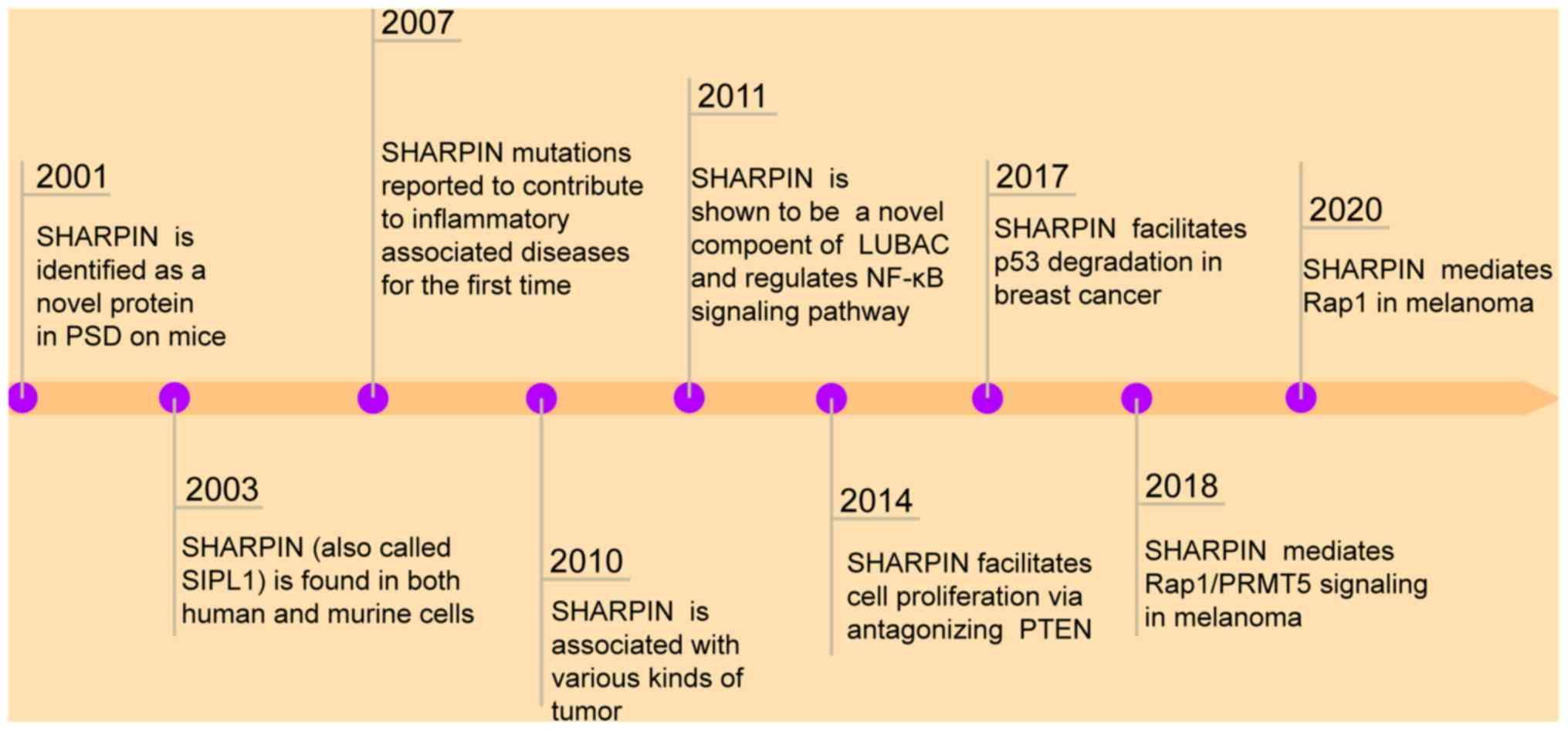
Shank-associated RH domain interactor (SHARPIN) is
an ~40-kDa multifunctional adaptor protein that is amplified and
overexpressed in a number of human cancer types. Studies have shown
that the SHARPIN promotes cancer cell proliferation, tumor
formation and metastasis (3–6). SHARPIN was first identified in
C57BL/KaLawRij mice, functioning as a shank binding protein at the
postsynaptic density of excitatory synapses in the central nervous
system (7). In addition, SHARPIN has
important physiological functions in several organisms (7) and is ubiquitously expressed in various
types of cells and tissues (8).
SHARPIN is an autosomal gene that is conserved among a
number of mammalian species, including humans, chimpanzees, dogs,
rats and mice (8). SHARPIN is
located on cell membranes and in the nuclei, and primarily
functions in immune and inflammatory responses (8). A previous study has reported that
mutations in this gene contribute to chronic proliferative
dermatitis, which is accompanied by immune system malfunction and
multi-organ inflammation (7).
Moreover, SHARPIN deficiency results in an autoinflammatory
phenotype in an inflammatory mouse model (7). There is evidence demonstrating that
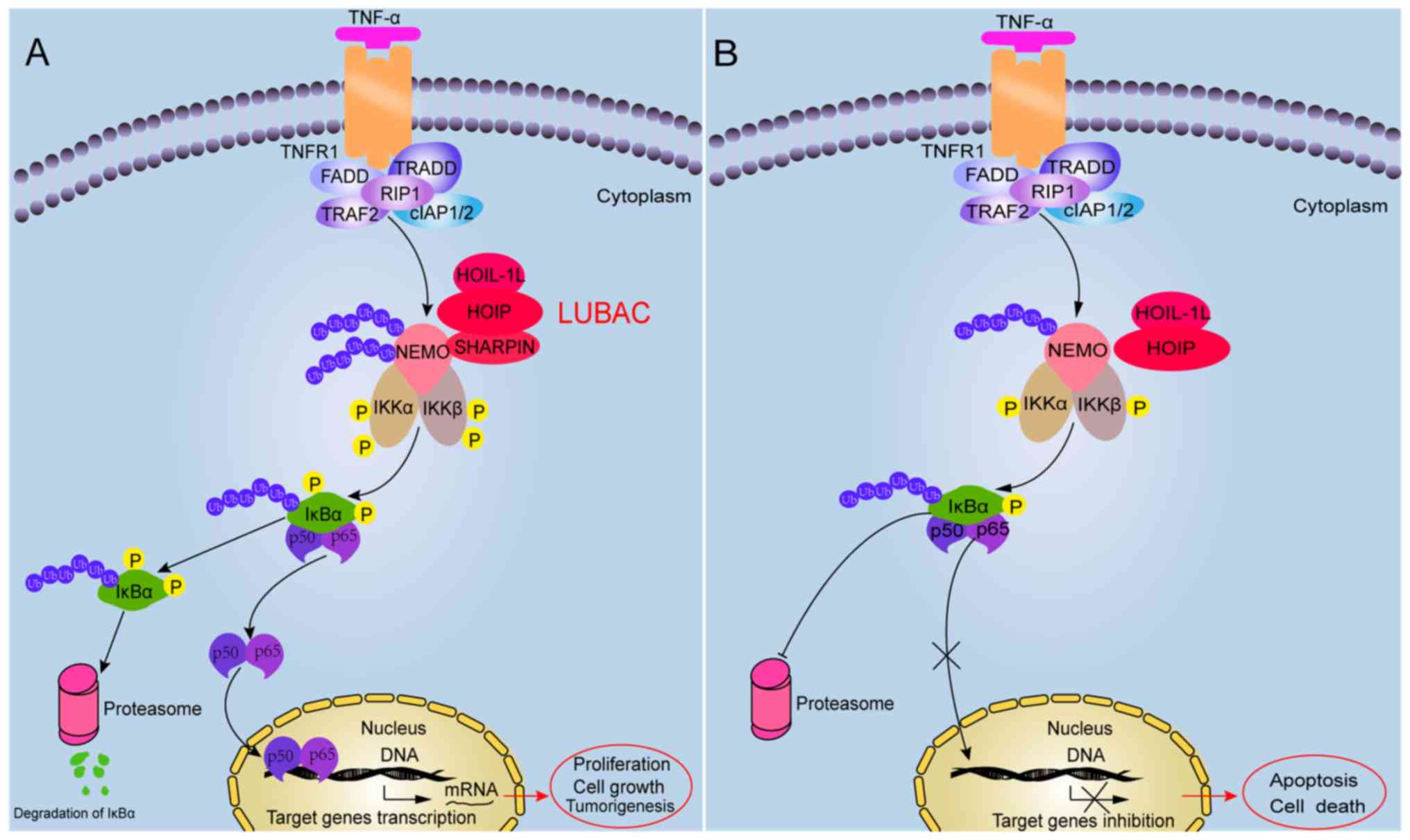
SHARPIN is a component of the linear ubiquitin chain assembly
complex (LUBAC), which participants in a range of complex
biological functions (9,10) (Fig.
1). Moreover, the LUBAC is also involved in various molecular
and cellular processes, such as embryogenesis (11) and apoptosis (12). SHARPIN has been well characterized as
a crucial regulator of canonical NF-κB signaling during the
inflammatory response (10,13) and T-cell differentiation (14,15). A
recent study reported that SHARPIN serves an important role in
promoting breast cancer progression (16). A previous study also showed that
SHARPIN regulates p53 protein levels in tumor cell lines through
the mouse double minute 2 homolog (MDM2)-dependent pathway
(17). Additionally, it has been
reported that SHARPIN interaction with protein arginine
methyltransferase 5 (PRMT5) mediates tumor cell growth (3). Notably, Zhou et al (18) demonstrated that SHARPIN upregulates
ras-associated protein-1 (Rap1), which promotes melanoma
development through p38 and c-Jun N-terminal kinases (JNK)/c-Jun
signaling pathways. A previous study also revealed that SHARPIN
mediates the phosphatase and tensin homologue deleted on chromosome
10 (PTEN) signaling pathway, which promotes tumorigenesis by
phosphoinositide 3-kinase (PI3K)/AKT signaling (19). Therefore, research investigating
SHARPIN has improved our understanding of its functions in humans;
however, a comprehensive summary of mechanisms by which SHARPIN
regulates tumorigenesis has not been available until now. Hence,
the following sections summarize the current data on the possible
functions of the aforementioned proteins and SHARPIN in
tumorigenesis.
Numerous studies have demonstrated that LUBAC
consists of three structurally related proteins: Heme-oxidized IRP2
ligase 1L (HOIL-1L), HOIL-1 interacting protein (HOIP) and SHARPIN,
with molecular weights of 120, 58 and 40 kD, respectively (20). LUBAC is involved in
post-translational modifications that regulate a multitude of
cellular processes, including cell death, development,
carcinogenesis and autoimmune diseases (21,22). The
HOIL-1L subunit is an accessory molecule that is involved in the
stabilization of LUBAC. For example, cells lacking HOIL-1L have
significantly decreased linear ubiquitylation (23), and the primary function of SHARPIN is
to maintain the linear ubiquitylation activity in LUBAC. HOIP is a
catalytic subunit that is associated with regulatory proteins, such
as HOIL-1L and SHARPIN (24). A
recent study demonstrated that the aforementioned subunits interact
with each other in the trimeric core of LUBAC, contributing to the
overall stabilization of the complex (25).
A recent study demonstrated that deficiency in
SHARPIN or HOIP in mice causes severe inflammation in adulthood or
embryonic lethality, respectively. By contrast, HOIL-1 deficiency
contributes no overt phenotype (11). Previously, the LUBAC was shown to
serve a pivotal role in complete activation of the canonical NF-κB
signaling pathway, whereas the absence of one subunit resulted in
attenuated activation of this pathway, particularly when SHARPIN
was absent (26). These data suggest
that HOIP, HOIL-1L and SHARPIN are all necessary for efficient
activation of the NF-κB signaling pathway. Notably, Rodgers et
al (27) suggested that linear
ubiquitination is required for activation of the NACHT, LRR and PYD
domains-containing protein 3 inflammasome, and this finding further
expands the role of LUBAC as an innate immune regulator.
In brief, the canonical NF-κB signaling pathway can
be triggered by different stimulators, such as tumor necrosis
factor (TNF)-α, interleukin (IL)-1 and pathogen-associated
molecular patterns (28,29). When stimulators combine with the TNF
receptor (TNFR), TNFR type 1-associated death domain protein,
FAS-associated death domain protein and TNFR-associated factor 2
proteins and protein kinases are recruited in the cytoplasm as a
result of phosphorylation and activation of the inhibition of the
κB kinase (IKK) complex (30). The
classical IKK complex consists of two catalytic subunits and a
regulatory subunit, IKKα, IKKβ and the NF-κB essential modulator
(NEMO), respectively. Furthermore, the activated IKK complex
facilitates the phosphorylation of IκB, which releases NF-κB dimers
that freely translocate into the nucleus and bind with DNA,
promoting the transcription of relevant target genes (30,31).
Recent studies have demonstrated that the LUBAC mediates linear
ubiquitylation, which is involved in the canonical NF-κB signaling
pathway (Fig. 2A).
Numerous studies have suggested that NEMO possesses
a specific ubiquitin-binding region that interacts with the LUBAC
(23,32–34). The
NF-κB activated state is influenced by the process of NEMO
conjugating with a linear poly-ubiquitin chain (35). Furthermore, NEMO deficiency leads to
decreased interaction with LUBAC, preventing SHARPIN-mediated
linear ubiquitination and NF-κB activation (14,36).
Consistent with this, SHAPRIN deficiency leads to inhibition of
LUBAC-mediated linear poly-ubiquitination of endogenous NEMO and
attenuates the activation of the NF-κB signaling pathway (37). Therefore, p65/p50 cannot translocate
into the nucleus to induce target gene expression due to decreased
phosphorylation and degradation of NF-κB inhibitor α (IKBα)
(28) (Fig. 2B). These findings suggest that the
NEMO-SHARPIN interaction is essential to mediate canonical NF-κB
signaling. Hence, these results suggest that SHARPIN is important
in regulating canonical NF-κB signaling and that this disruption
may impact downstream physiological functions.
It is well known that p53 is a vital tumor
suppressor and its presence was initially reported in response to
various types of stress >40 years ago (38). The p53 protein acts as a
transcription factor in cells and is functionally inactivated in
the majority of cancer types (39).
Several studies have demonstrated that activated p53 is associated
with numerous downstream responses and is involved in important
physiological and pathological processes, including cell cycle
arrest, DNA repair, apoptosis, metabolism, invasion, metastasis and
tumorigenesis (40–42). p53 is maintained at very low cellular
levels in normal cells due to binding with E3 ubiquitin ligase
mouse double minute 2 homolog (MDM2), which contributes to p53
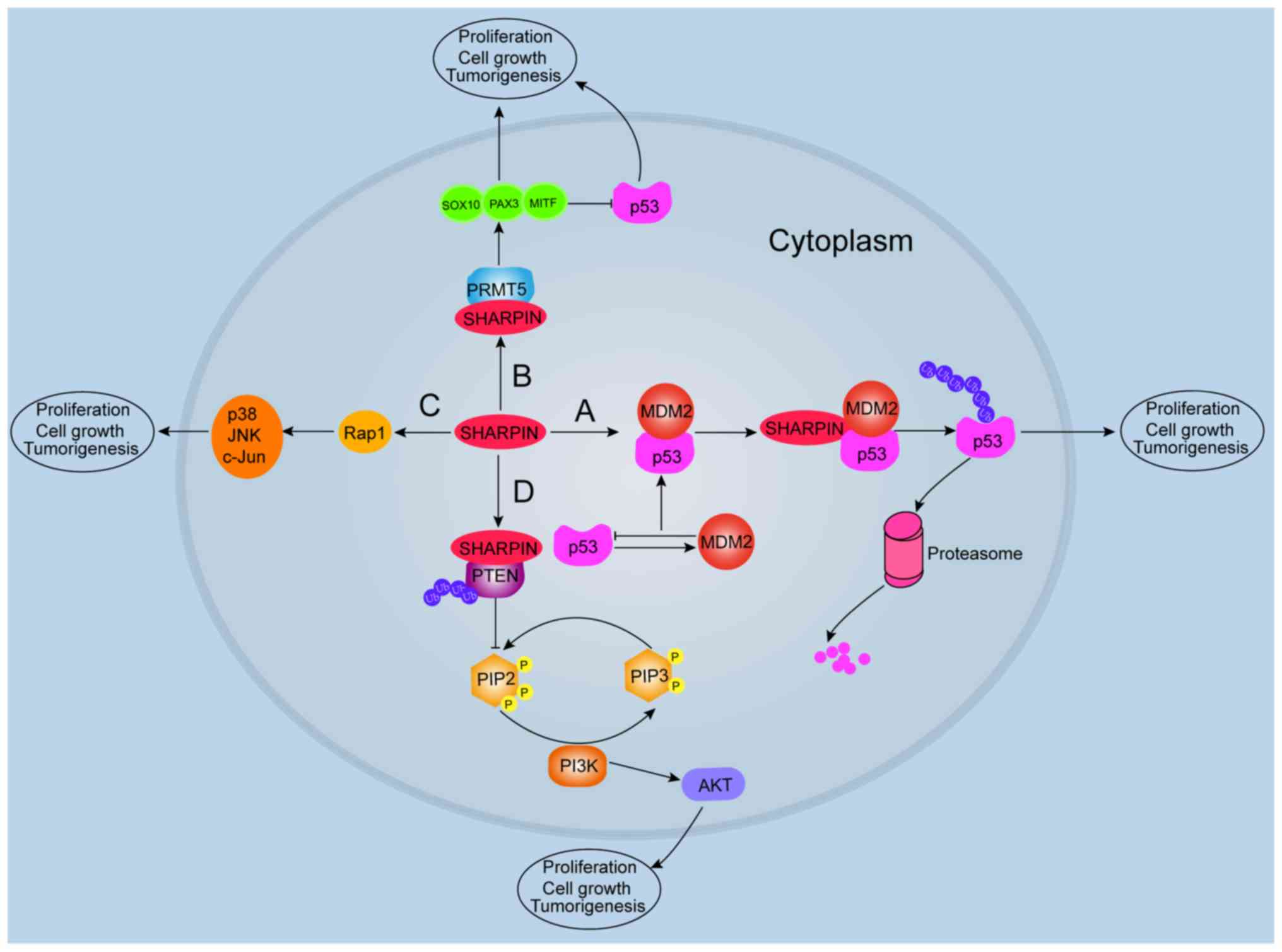
ubiquitination and rapid degradation by the proteasome (43). SHARPIN may be involved in promoting
p53 ubiquitination and maintaining cellular levels of p53 (44). Mechanistically, MDM2 recognizes the
N-terminal trans-activation domain of p53 and mediates p53
transcription as an inhibitor of p53 transcriptional activation
(45). Nevertheless, in response to
cellular stress signals, p53 is rapidly activated by
phosphorylation, which releases MDM2 and promotes p53 stabilization
(46). MDM2 regulates p53 through
direct binding, mono-ubiquitylation, polyubiquitylation (44) or negative feedback regulation
(47), which maintains appropriate
cellular levels of p53.
A recent study demonstrated that SHARPIN may be
upstream of p53 signaling in breast cancer cells, as depleted
SHARPIN resulted in decreased cell proliferation and increased
expression of p53 (17).
Furthermore, the study reported that SHARPIN modulates p53 protein
levels through poly-ubiquitination and degradation in a
MDM2-dependent manner, which determines the fate of p53 in tumor
cells (Fig. 3). Therefore, p53
protein levels in cells are indirectly regulated by SHARPIN.
Nevertheless, whether other accessory molecules or signaling
pathways are involved in SHARPIN-mediated regulation of the
p53/MDM2 complex needs further clarification.
PRMT5 is a member of the PRMT family of proteins and
can catalyze symmetric methylation of histone and non-histone
proteins. PRMT5 catalyzes the transfer of a methyl group from
S-adenosylmethionine to the guanidino nitrogen atoms of arginine
(48). PRMT5 is commonly activated
in cancer, which is mediated in part by the PRMT5 co-factor
methylosome protein 50 (also known as p44) (49). Researchers have demonstrated that
PRMT5 serves an essential role in lung cancer, leukemia (49), tumorigenesis (50), cell survival and human embryonic stem
cell proliferation (51). Stopa
et al (52) provides an
overall literature review of PRMT5 overexpression, which appears to
be an important factor in the tumorigenicity of a large number of
cancer types, such as gastric (53)
and breast cancer (54). In
addition, PRMT5 mediates the methylation of Arg1175 of epidermal
growth factor receptor, which controls extracellular
signal-regulated kinase activation (55). Similarly, E2F-1 may be directly
methylated by PRMT5, which influences the stability of E2F-1.
Depleting PRMT5 using small interfering RNA resulted in decreased
arginine methylation of E2F-1 but increased E2F-1 levels in one
study (56).
Recent reports have shown that the interaction of
SHARPIN with PRMT5 contributes to regulating the transcription of
cancer-associated genes (3,56). For example, Fu et al (57) reported that SHARPIN activates PRMT5
to specifically target histone H3R2 for mono-methylation, which is
responsible for the subsequent activation of cancer-associated
genes and mediates metastasis in invasive lung cancer cells. Tamiya
et al (3) also demonstrated
that PRMT5 activity is increased after SHARPIN binding. This study
observed that SHARPIN activates PRMT5 by regulating SRY-box
transcription factor 10, paired box gene 3 (PAX3) and
microphthalmia-associated (MITF) transcription factors in melanoma
development (Fig. 3). These results
demonstrate that both SHARPIN and PRMT5 are involved in
tumorigenesis.
SHARPIN regulates PRMT5 activity, which induces the
transcriptional activity of PAX3 and MITF in melanoma growth; in
turn, PAX3 and MITF can inhibit p53 activation (3). Consistent with these findings, a recent
study demonstrated that PAX3 specifically binds to the promoter of
p53 leading to repressed p53 expression in glioblastoma (65). In addition, MITF binds p53 to
regulate cyclin-dependent kinase inhibitor 1A in melanoma cells
(66). Overall, these data suggest
that PRMT5 is capable of regulating p53 indirectly, and SHARPIN
mediates p53 through PRMT5-dependent signaling with PAX3 and
MITF.
Studies have shown that PTEN is a pivotal tumor
suppressor gene, and that it is frequently deleted in late-stage
human cancer types and has important functions in crosstalk with
the PI3K/ATK signaling pathway, mediating several fundamental
cellular processes under different circumstances via multiple
downstream targets (68–70). For example, a study has suggested
that PI3K/AKT signaling is attenuated in the brains of patients
with Alzheimer's disease (71).
Moreover, PI3K/AKT/PTEN is mediated by intracellular ROS production
(72). The main function of PTEN is
to catalyze the conversion of phosphatidylinositol (3,4,5)-trisphosphate (PIP3) to
phosphatidylinositol (4,5)-bisphosphate (73). PTEN is an antagonist of PI3K and,
when dephosphorylated, serves as a negative regulator of PI3K/AKT
signaling in normal cells, while loss or inactivation of PTEN
contributes to hyperactivation of PI3K/AKT in primary T-cell
leukemia (74) and adult B-cell
acute lymphoblastic leukemia (75).
Once activated, the PI3K/AKT pathway promotes cell survival by
blocking the function of pro-apoptotic proteins and promoting
protein synthesis and cell proliferation. Consequently, these
results indicate that inactivation of PTEN is a critical step
during tumorigenesis.
There has been much interest in the transcriptional
and post-translational modulation of PTEN expression, protein
stability and activity mediated by miRNA (76), phosphorylation (77), acetylation (78) and ubiquitination (79). A recent study reports that PTEN
knockdown could significantly promote mouse neuronal cell
proliferation and differentiation in vitro (80). Moreover, a previous study
demonstrated that SHARPIN is a negative regulator of PTEN in human
tumor cell lines and human primary cervical cancer cells both in
vitro and in vivo (81).
Furthermore, it has been reported that SHARPIN interacts with PTEN
through its ubiquitin-like domain (81). It is well known that the major
function of PTEN is inhibition of the PI3K signaling pathway, while
loss of PTEN activates the PI3K/AKT pathway (82). Additionally, attenuation of PTEN
function activates PI3K/AKT signaling and elicits tumorigenesis
(82). Hence, reducing PTEN-induced
PIP3 phosphatase activity and enhancing the activity of the
PI3K/AKT signaling pathway promotes tumorigenesis (83). In accordance with this, De Melo et
al (84) suggested that SHARPIN
facilitated PTEN poly-ubiquitination via lysine63 and formation of
the SHARPIN/PTEN complex, which did not lead to PTEN degradation.
These biochemical processes alter the affinity of the SHARPIN/PTEN
complex and reduce the phosphatase activity of PTEN (Fig. 3). The study proposed that SHARPIN
promotes poly-ubiquitination of PTEN in a manner that does not
alter PTEN stability, and that SHARPIN/PTEN binding is enhanced by
poly-ubiquitination of PTEN.
Overall, there has been a marked increase in the
understanding of the function of SHARPIN in recent years. The
present review summarizes our understanding of how SHARPIN mediates
canonical NF-κB signaling and crosstalk with various mechanisms,
regulating tumorigenesis. Furthermore, the function of SHARPIN in
the NF-κB, p53, PRMT5, Rap1 and PTEN signaling pathways was
explored. The aforementioned results shed light on possible
functions of these proteins in tumor cells, and may provide novel
targets for cancer treatment in the future. However, the detailed
molecular function of associated co-activators or co-repressors
contributing to these pathways needs further investigation.
The authors would like to thank Professor Shaogang
Qu (Nanfang Hospital, Southern Medical University, Guangzhou,
China) for assistance with the manuscript revision.
The present study was supported by grants from the
Science and Technology Program of Foshan (grant no. 1920001000694),
the Scientific Research Startup of Shunde Hospital (grant no.
SRSP2019012) and the Medical Scientific Research Foundation of
Guangdong Province (grant no. A2020175).
Not applicable.
CZ, DX, KZ and JY jointly conceived and designed the
review, researched the literature and wrote the manuscript. All
authors read and approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Bray F, Jemal A, Grey N, Ferlay J and
Forman D: Global cancer transitions according to the Human
development index (2008–2030): A population-based study. Lancet
Oncol. 13:790–801. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Luo J, Solimini NL and Elledge SJ:
Principles of cancer therapy: Oncogene and non-oncogene addiction.
Cell. 136:823–837. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tamiya H, Kim H, Klymenko O, Kim H, Feng
Y, Zhang T, Han JY, Murao A, Snipas SJ, Jilaveanu L, et al:
SHARPIN-mediated regulation of protein arginine methyltransferase 5
controls melanoma growth. J Clin Invest. 128:517–530. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jung J, Kim JM, Park B, Cheon Y, Lee B,
Choo SH, Koh SS and Lee S: Newly identified tumor-associated role
of human Sharpin. Mol Cell Biochem. 340:161–167. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ojo D, Wu Y, Bane A and Tang D: A role of
SIPL1/SHARPIN in promoting resistance to hormone therapy in breast
cancer. Biochim Biophys Acta Mol Basis Dis. 1864:735–745. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bii VM, Rae DT and Trobridge GD: A novel
gammaretroviral shuttle vector insertional mutagenesis screen
identifies SHARPIN as a breast cancer metastasis gene and
prognostic biomarker. Oncotarget. 6:39507–39520. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Seymour RE, Hasham MG, Cox GA, Shultz LD,
Hogenesch H, Roopenian DC and Sundberg JP: Spontaneous mutations in
the mouse Sharpin gene result in multiorgan inflammation, immune
system dysregulation and dermatitis. Genes Immun. 8:416–421. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang Z, Potter CS, Sundberg JP and
Hogenesch H: SHARPIN is a key regulator of immune and inflammatory
responses. J Cell Mol Med. 16:2271–2279. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rittinger K and Ikeda F: Linear ubiquitin
chains: Enzymes, mechanisms and biology. Open Biol. 7:1700262017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tokunaga F, Nakagawa T, Nakahara M, Saeki
Y, Taniguchi M, Sakata S, Tanaka K, Nakano H and Iwai K: SHARPIN is
a component of the NF-κB-activating linear ubiquitin chain assembly
complex. Nature. 471:633–636. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Peltzer N, Darding M, Montinaro A, Draber
P, Draberova H, Kupka S, Rieser E, Fisher A, Hutchinson C,
Taraborrelli L, et al: LUBAC is essential for embryogenesis by
preventing cell death and enabling haematopoiesis. Nature.
557:112–117. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tang Y, Joo D, Liu G, Tu H, You J, Jin J,
Zhao X, Hung MC and Lin X: Linear ubiquitination of cFLIP induced
by LUBAC contributes to TNFα-induced apoptosis. J Biol Chem.
293:20062–20072. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ikeda F, Deribe YL, Skanland SS, Stieglitz
B, Grabbe C, Franz-Wachtel M, van Wijk SJ, Goswami P, Nagy V,
Terzic J, et al: SHARPIN forms a linear ubiquitin ligase complex
regulating NF-κB activity and apoptosis. Nature. 471:637–641. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Teh CE, Lalaoui N, Jain R, Policheni AN,
Heinlein M, Alvarez-Diaz S, Sheridan JM, Rieser E, Deuser S,
Darding M, et al: Linear ubiquitin chain assembly complex
coordinates late thymic T-cell differentiation and regulatory
T-cell homeostasis. Nat Commun. 7:133532016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Redecke V, Chaturvedi V, Kuriakose J and
Hacker H: SHARPIN controls the development of regulatory T cells.
Immunology. 148:216–226. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tian Z, Tang J, Yang Q, Li X, Zhu J and Wu
G: Atypical ubiquitin-binding protein SHARPIN promotes breast
cancer progression. Biomed Pharmacother. 119:1094142019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang H, Yu S, Wang W, Li X, Hou Y, Liu Z,
Shi Y, Mu K, Niu G, Xu J, et al: SHARPIN facilitates p53
degradation in breast cancer cells. Neoplasia. 19:84–92. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhou S, Liang Y, Zhang X, Liao L, Yang Y,
Ouyang W and Xu H: SHARPIN promotes melanoma progression via Rap1
signaling pathway. J Invest Dermatol. 140:395–403.e6. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
De Melo J, Wu V, He L, Yan J and Tang D:
SIPL1 enhances the proliferation, attachment, and migration of CHO
cells by inhibiting PTEN function. Int J Mol Med. 34:835–841. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sasaki K and Iwai K: Roles of linear
ubiquitinylation, a crucial regulator of NF-κB and cell death, in
the immune system. Immunol Rev. 266:175–189. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ikeda F: Linear ubiquitination signals in
adaptive immune responses. Immunol Rev. 266:222–236. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Iwai K, Fujita H and Sasaki Y: Linear
ubiquitin chains: NF-κB signalling, cell death and beyond. Nat Rev
Mol Cell Biol. 15:503–508. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tokunaga F, Sakata S, Saeki Y, Satomi Y,
Kirisako T, Kamei K, Nakagawa T, Kato M, Murata S, Yamaoka S, et
al: Involvement of linear polyubiquitylation of NEMO in NF-kappaB
activation. Nat Cell Biol. 11:123–132. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rivkin E, Almeida SM, Ceccarelli DF, Juang
YC, MacLean TA, Srikumar T, Huang H, Dunham WH, Fukumura R, Xie G,
et al: The linear ubiquitin-specific deubiquitinase gumby regulates
angiogenesis. Nature. 498:318–324. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fujita H, Tokunaga A, Shimizu S, Whiting
AL, Aguilar-Alonso F, Takagi K, Walinda E, Sasaki Y, Shimokawa T,
Mizushima T, et al: Cooperative domain formation by homologous
motifs in HOIL-1L and SHARPIN plays A crucial role in LUBAC
stabilization. Cell Rep. 23:1192–1204. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Matsunaga Y, Nakatsu Y, Fukushima T, Okubo
H, Iwashita M, Sakoda H, Fujishiro M, Yamamotoya T, Kushiyama A,
Takahashi S, et al: LUBAC formation is impaired in the livers of
mice with MCD-dependent nonalcoholic steatohepatitis. Mediators
Inflamm. 2015:1253802015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rodgers MA, Bowman JW, Fujita H, Orazio N,
Shi M, Liang Q, Amatya R, Kelly TJ, Iwai K, Ting J, et al: The
linear ubiquitin assembly complex (LUBAC) is essential for NLRP3
inflammasome activation. J Exp Med. 211:1333–1347. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tokunaga F: Linear ubiquitination-mediated
NF-κB regulation and its related disorders. J Biochem. 154:313–323.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Oeckinghaus A, Hayden MS and Ghosh S:
Crosstalk in NF-κB signaling pathways. Nat Immunol. 12:695–708.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
D'Ignazio L, Batie M and Rocha S: Hypoxia
and inflammation in cancer, focus on HIF and NF-κB. Biomedicines.
5:212017. View Article : Google Scholar
|
|
31
|
Israel A: The IKK complex, a central
regulator of NF-kappaB activation. Cold Spring Harb Perspect Biol.
2:a0001582010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Smit JJ, van Dijk WJ, El Atmioui D, Merkx
R, Ovaa H and Sixma TK: Target specificity of the E3 ligase LUBAC
for ubiquitin and NEMO relies on different minimal requirements. J
Biol Chem. 288:31728–31737. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tokunaga F and Iwai K: Involvement of
LUBAC-mediated linear polyubiquitination of NEMO in NF-kappaB
activation. Tanpakushitsu Kakusan Koso. 54:635–642. 2009.(In
Japanese). PubMed/NCBI
|
|
34
|
Iwai K and Tokunaga F: Linear
polyubiquitination: A new regulator of NF-kappaB activation. EMBO
Rep. 10:706–713. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Rahighi S, Ikeda F, Kawasaki M, Akutsu M,
Suzuki N, Kato R, Kensche T, Uejima T, Bloor S, Komander D, et al:
Specific recognition of linear ubiquitin chains by NEMO is
important for NF-kappaB activation. Cell. 136:1098–1109. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bal E, Laplantine E, Hamel Y, Dubosclard
V, Boisson B, Pescatore A, Picard C, Hadj-Rabia S, Royer G,
Steffann J, et al: Lack of interaction between NEMO and SHARPIN
impairs linear ubiquitination and NF-κB activation and leads to
incontinentia pigmenti. J Allergy Clin Immunol. 140:1671–1682.e2.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Niu J, Shi Y, Iwai K and Wu ZH: LUBAC
regulates NF-κB activation upon genotoxic stress by promoting
linear ubiquitination of NEMO. EMBO J. 30:3741–3753. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lane D and Levine A: p53 Research: The
past thirty years and the next thirty years. Cold Spring Harb
Perspect Biol. 2:a0008932010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ozaki T and Nakagawara A: Role of p53 in
cell death and human cancers. Cancers (Basel). 3:994–1013. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Bieging KT, Mello SS and Attardi LD:
Unravelling mechanisms of p53-mediated tumour suppression. Nat Rev
Cancer. 14:359–370. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Muller PA, Vousden KH and Norman JC: p53
and its mutants in tumor cell migration and invasion. J Cell Biol.
192:209–218. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Scoumanne A, Zhang J and Chen X: PRMT5 is
required for cell-cycle progression and p53 tumor suppressor
function. Nucleic Acids Res. 37:4965–4976. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Honda R, Tanaka H and Yasuda H:
Oncoprotein MDM2 is a ubiquitin ligase E3 for tumor suppressor p53.
FEBS Lett. 420:25–27. 2007. View Article : Google Scholar
|
|
44
|
Lee JT and Gu W: The multiple levels of
regulation by p53 ubiquitination. Cell Death Differ. 17:86–92.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lessel D, Wu D, Trujillo C, Ramezani T,
Lessel I, Alwasiyah MK, Saha B, Hisama FM, Rading K, Goebel I, et
al: Dysfunction of the MDM2/p53 axis is linked to premature aging.
J Clin Invest. 127:3598–3608. 2007. View Article : Google Scholar
|
|
46
|
Chen J, Lin J and Levine AJ: Regulation of
transcription functions of the p53 tumor suppressor by the mdm-2
oncogene. Mol Med. 1:142–152. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wu X, Bayle JH, Olson D and Levine AJ: The
p53-mdm-2 autoregulatory feedback loop. Genes Dev. 7:1126–1132.
1999. View Article : Google Scholar
|
|
48
|
Jin Y, Zhou J, Xu F, Jin B, Cui L, Wang Y,
Du X, Li J, Li P, Ren R and Pan J: Targeting methyltransferase
PRMT5 eliminates leukemia stem cells in chronic myelogenous
leukemia. J Clin Invest. 126:3961–3980. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Rehman I, Basu SM, Das SK, Bhattacharjee
S, Ghosh A, Pommier Y and Das BB: PRMT5-mediated arginine
methylation of TDP1 for the repair of topoisomerase I covalent
complexes. Nucleic Acids Res. 46:5601–5617. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Durant ST, Cho EC and La Thangue NB: p53
methylation-the Arg-ument is clear. Cell Cycle. 8:801–802. 2008.
View Article : Google Scholar
|
|
51
|
Gkountela S, Li Z, Chin CJ, Lee SA and
Clark AT: PRMT5 is required for human embryonic stem cell
proliferation but not pluripotency. Stem Cell Rev. 10:230–239.
2014. View Article : Google Scholar
|
|
52
|
Stopa N, Krebs JE and Shechter D: The
PRMT5 arginine methyltransferase: Many roles in development, cancer
and beyond. Cell Mol Life Sci. 72:2041–2059. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Liu M, Yao B, Gui T, Guo C, Wu X, Li J, Ma
L, Deng Y, Xu P, Wang Y, et al: PRMT5-dependent transcriptional
repression of c-Myc target genes promotes gastric cancer
progression. Theranostics. 10:4437–4452. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Vinet M, Suresh S, Maire V, Monchecourt C,
Nemati F, Lesage L, Pierre F, Ye M, Lescure A, Brisson A, et al:
Protein arginine methyltransferase 5: A novel therapeutic target
for triple-negative breast cancers. Cancer Med. 8:2414–2428. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Hsu JM, Chen CT, Chou CK, Kuo HP, Li LY,
Lin CY, Lee HJ, Wang YN, Liu M, Liao HW, et al: Crosstalk between
Arg 1175 methylation and Tyr 1173 phosphorylation negatively
modulates EGFR-mediated ERK activation. Nat Cell Biol. 13:174–181.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Cho EC, Zheng S, Munro S, Liu G, Carr SM,
Moehlenbrink J, Lu YC, Stimson L, Khan O, Konietzny R, et al:
Arginine methylation controls growth regulation by E2F-1. EMBO J.
31:1785–1797. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Fu T, Lv X, Kong Q and Yuan C: A novel
SHARPIN-PRMT5-H3R2me1 axis is essential for lung cancer cell
invasion. Oncotarget. 8:54809–54820. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Jansson M, Durant ST, Cho EC, Sheahan S,
Edelmann M, Kessler B and La Thangue NB: Arginine methylation
regulates the p53 response. Nat Cell Biol. 10:1431–1439. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Yang M, Sun J, Sun X, Shen Q, Gao Z and
Yang C: Caenorhabditis elegans protein arginine methyltransferase
PRMT-5 negatively regulates DNA damage-induced apoptosis. PLoS
Genet. 5:e10005142009. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Bezzi M, Teo SX, Muller J, Mok WC, Sahu
SK, Vardy LA, Bonday ZQ and Guccione E: Regulation of constitutive
and alternative splicing by PRMT5 reveals a role for Mdm4 pre-mRNA
in sensing defects in the spliceosomal machinery. Genes Dev.
27:1903–1916. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Gerhart SV, Kellner WA, Thompson C,
Pappalardi MB, Zhang XP, Montes de Oca R, Penebre E, Duncan K,
Boriack-Sjodin A, Le B, et al: Activation of the p53-MDM4
regulatory axis defines the anti-tumour response to PRMT5
inhibition through its role in regulating cellular splicing. Sci
Rep. 8:97112018. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Li Y and Diehl JA: PRMT5-dependent p53
escape in tumorigenesis. Oncoscience. 2:700–702. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Scaglione A, Patzig J, Liang J, Frawley R,
Bok J, Mela A, Yattah C, Zhang J, Teo SX, Zhou T, et al:
PRMT5-mediated regulation of developmental myelination. Nat Commun.
9:28402018. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Liu F, Cheng G, Hamard PJ, Greenblatt S,
Wang L, Man N, Perna F, Xu H, Tadi M, Luciani L, et al: Arginine
methyltransferase PRMT5 is essential for sustaining normal adult
hematopoiesis. J Clin Invest. 125:3532–3544. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Zhu H, Wang H, Huang Q, Liu Q, Guo Y, Lu
J, Li X, Xue C and Han Q: Transcriptional repression of p53 by PAX3
contributes to gliomagenesis and differentiation of glioma stem
cells. Front Mol Neurosci. 11:1872018. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Wang C, Zhao L, Su Q, Fan X, Wang Y, Gao
S, Wang H, Chen H, Chan CB and Liu Z: Phosphorylation of MITF by
AKT affects its downstream targets and causes TP53-dependent cell
senescence. Int J Biochem Cell Biol. 80:132–142. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Lilja J, Zacharchenko T, Georgiadou M,
Jacquemet G, De Franceschi N, Peuhu E, Hamidi H, Pouwels J, Martens
V, Nia FH, et al: SHANK proteins limit integrin activation by
directly interacting with Rap1 and R-Ras. Nat Cell Biol.
19:292–305. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Lee JT, Shan J, Zhong J, Li M, Zhou B,
Zhou A, Parsons R and Gu W: RFP-mediated ubiquitination of PTEN
modulates its effect on AKT activation. Cell Res. 23:552–564. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Graupera M, Guillermet-Guibert J, Foukas
LC, Phng LK, Cain RJ, Salpekar A, Pearce W, Meek S, Millan J,
Cutillas PR, et al: Angiogenesis selectively requires the p110alpha
isoform of PI3K to control endothelial cell migration. Nature.
453:662–666. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Worby CA and Dixon JE: Pten. Annu Rev
Biochem. 83:641–669. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Wang Y, Wu C, Han B, Xu F, Mao M, Guo X
and Wang J: Dexmedetomidine attenuates repeated propofol
exposure-induced hippocampal apoptosis, PI3K/Akt/Gsk-3β signaling
disruption, and juvenile cognitive deficits in neonatal rats. Mol
Med Rep. 14:769–775. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Nakanishi A, Wada Y, Kitagishi Y and
Matsuda S: Link between PI3K/AKT/PTEN pathway and NOX proteinin
diseases. Aging Dis. 5:203–211. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Engelman JA, Luo J and Cantley LC: The
evolution of phosphatidylinositol 3-kinases as regulators of growth
and metabolism. Nat Rev Genet. 7:606–619. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Silva A, Yunes JA, Cardoso BA, Martins LR,
Jotta PY, Abecasis M, Nowill AE, Leslie NR, Cardoso AA and Barata
JT: PTEN posttranslational inactivation and hyperactivation of the
PI3K/Akt pathway sustain primary T cell leukemia viability. J Clin
Invest. 118:3762–3774. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Gomes AM, Soares MV, Ribeiro P, Caldas J,
Povoa V, Martins LR, Melao A, Serra-Caetano A, de Sousa AB, Lacerda
JF, et al: Adult B-cell acute lymphoblastic leukemia cells display
decreased PTEN activity and constitutive hyperactivation of
PI3K/Akt pathway despite high PTEN protein levels. Haematologica.
99:1062–1068. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Poliseno L, Salmena L, Zhang J, Carver B,
Haveman WJ and Pandolfi PP: A coding-independent function of gene
and pseudogene mRNAs regulates tumour biology. Nature.
465:1033–1038. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Al-Khouri AM, Ma Y, Togo SH, Williams S
and Mustelin T: Cooperative phosphorylation of the tumor suppressor
phosphatase and tensin homologue (PTEN) by casein kinases and
glycogen synthase kinase 3beta. J Biol Chem. 280:35195–35202. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Ikenoue T, Inoki K, Zhao B and Guan KL:
PTEN acetylation modulates its interaction with PDZ domain. Cancer
Res. 68:6908–6912. 2018. View Article : Google Scholar
|
|
79
|
Yang JM, Schiapparelli P, Nguyen HN,
Igarashi A, Zhang Q, Abbadi S, Amzel LM, Sesaki H,
Quinones-Hinojosa A and Iijima M: Characterization of PTEN
mutations in brain cancer reveals that pten mono-ubiquitination
promotes protein stability and nuclear localization. Oncogene.
36:3673–3685. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Song Z, Han X, Shen L, Zou H, Zhang B, Liu
J and Gong A: PTEN silencing enhances neuronal proliferation and
differentiation by activating PI3K/Akt/GSK3β pathway in vitro. Exp
Cell Res. 363:179–187. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Hopkins BD, Hodakoski C, Barrows D, Mense
SM and Parsons RE: PTEN function: The long and the short of it.
Trends Biochem Sci. 39:183–190. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Wu X, Senechal K, Neshat MS, Whang YE and
Sawyers CL: The PTEN/MMAC1 tumor suppressor phosphatase functions
as a negative regulator of the phosphoinositide 3-kinase/Akt
pathway. Proc Natl Acad Sci USA. 95:15587–15591. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
He L, Ingram A, Rybak AP and Tang D:
Shank-interacting protein-like 1 promotes tumorigenesis via PTEN
inhibition in human tumor cells. J Clin Invest. 120:2094–2108.
2012. View Article : Google Scholar
|
|
84
|
De Melo J, Lin X, He L, Wei F, Major P and
Tang D: SIPL1-facilitated PTEN ubiquitination contributes to its
association with PTEN. Cell Signal. 26:2749–2756. 2014. View Article : Google Scholar : PubMed/NCBI
|