Introduction
Hepatocellular carcinoma (HCC) is one of the most
malignant cancer types worldwide, especially in east Asia (1). Although significant improvements in
diagnosis and treatment have been accomplished, the mortality rate
for patients with HCC remains high, with ~782,000 deaths worldwide
annually (1–3). The rapid and unnoticeable development
and metastases of HCC are crucial to its progression, resulting in
unresectable vascular thrombi, intrahepatic satellite nodules or
cancer recurrence (4,5). Hence, it is important to discover
effective diagnostic biomarkers to identify HCC in the early stage
to improve the prognosis of patients with HCC.
NF-κB inhibitor α (IκBα) is an isoform in the IκB
family, members of which have multiple ankyrin repeat domains at
the C-terminus (6). IκBα combines
with the NF-κB dimer to retain its existence in the cytoplasm and
prevent its translocation into the nucleus (6). The IκB kinase (IKK) complex is induced
by diverse extracellular signals, such as lipopolysaccharide, tumor
necrosis factor (TNF) and growth factors, and the activated IKK
complex phosphorylates IκBα. Then, phosphorylated IκBα is
specifically recognized by an E3 ubiquitin ligase, leading to its
ubiquitination and degradation (7).
While previous studies have reported its inhibition of NF-κB, the
remaining functions of IκBα in HCC are yet to be fully
elucidated.
p65 is a well-known member of the NF-κB family of
transcriptional factors participating in numerous pathways in
biological responses, such as inflammation, adaptive immunity and
stress reactions (8). Previous
studies have revealed that NF-κB is involved in cancer oncogenesis
and that it specifically promotes proliferation, metastasis and
cell survival, particularly in inflammation-related cancer types
(9,10). In healthy cells, activation of NF-κB
after interaction with cellular stimuli occurs for a short duration
and then returns to the inactive form. However, NF-κB is activated
aberrantly in cancer cells, and thus continues to upregulate the
transcription of its target gene (11). To achieve its transcriptional
function, NF-κB/Rel proto-oncogene, NF-κB subunit (Rel) complex
should shuttle to nucleus; but, IκBα combines with the dimer to
anchor it in cytoplasm and thus inhibit its activity (8). After its phosphorylation is initiated
by proinflammation signals or immune stimulation, IκBα is degraded
by the ubiquitin-protease system, which enables the NF-κB/Rel dimer
to translocate in nucleus (12).
Therefore, reduced expression of IκBα is an important cause of the
abnormal elevated expression of NF-κB in cancer cells.
Erbb2 interacting protein (Erbin) is a target gene
of NF-κB, encoding Erbin. Containing 16 leucine-rich repeats and
one PSD-95/Dlg/ZO-1 (PDZ) domain, Erbin is a member of the
leucine-rich repeat and PDZ domain family (13). Furthermore, Erbin is an adaptor of
the Erb-b2 receptor tyrosine kinase 2 (ERBB2) protein and, by
binding with ERBB2, regulates its function and localization
(13). Erbin has also been reported
to be involved in several pathways (14). For instance, Erbin suppresses
tumorigenesis via inhibiting Akt activity or Ras/Raf signaling
(15,16). In addition, Erbin exerts an
ontogenetic function by activating estrogen receptor (ER) α or
inhibiting STAT3 signaling (17,18).
Thus, there is conflicting data regarding the role of Erbin in
cancer. A previous study showed that Erbin is upregulated following
exposure of macrophages to muramyl dipeptide, TNF α and
lipopolysaccharide (19). Although
it has been observed that Erbin is upregulated in HCC tissues, the
underlying mechanism via which Erbin becomes dysregulated remains
unknown.
The present study investigated the clinical
importance of IκBα, Erbin and their molecular interactions, which
suggested that reduced IκBα expression promoted HCC cell
proliferation and migration via the NF-κB/Erbin pathway.
Mechanistically, IκBα was downregulated in HCC tissues compared
with healthy liver tissues, and decreased expression of IκBα
weakened the inhibition of NF-κB. Abnormally activated NF-κB
promoted Erbin expression at both the mRNA and protein levels,
which served an oncogenic function in HCC. Moreover, IκBα was
negatively associated with Erbin, and decreased IκBα and increased
Erbin expression levels were associated with poor outcomes in
patients with HCC. Therefore, the present results may facilitate
the development of a potential strategy to target Erbin for HCC
treatment.
Materials and methods
Patients with HCC and sample
collection
In all, 107 human HCC tissue specimens were acquired
from patients (aged between 19 and 73 years old, and 93 male, 14
female) between January 2011 and December 2013 at the Department of
Hepatobiliary Surgery of Southwest Hospital, Army Medical
University (Chongqing, China). All patients provided written
informed consent and this study obtained approval from the Ethics
Committee of Southwest Hospital. All patients used in this study
were followed up for 5 years since their cancer diagnosis. Upon
acquisition, HCC and paracancerous liver tissues (free of tumor to
the eye) were conventionally 10% formalin-fixed for 48 h on a
rotator at room temperature, paraffin-embedded and cut into 10-µm
sections.
Immunohistochemistry (IHC)
staining
To perform IHC staining, sections were first
deparaffinized, rehydrated and then boiled for 2.5 min in 100°C
0.01 mol/l sodium citrate solution for antigen retrieval. Then the
IHC detection kit (SP-9001; ZSGB-Bio, Beijing, China) was employed
from blockage of peroxidase to incubation of secondary antibodies
according to the manufacturer's instructions. Briefly, 3%
H2O2 was used to block the endogenous
peroxidase at room temperature for 15 min. After blocking with 5%
goat serum at room temperature for 30 min, tissues were incubated
with specific antibodies: Anti-IκBα (sc-945; 1:50 dilution; Santa
Cruz Biotechnology, Inc.) and anti-Erbin (22438–1-AP; 1:50
dilution; ProteinTech Group, Inc.) overnight at 4°C. The following
day the sections were incubated with horseradish
peroxidase-conjugated secondary antibody, before positive signals
were detected using 3,3′-diaminobenzidine and nuclei were stained
with hematoxylin at room temperature for 20 sec. After dehydration,
representative images were captured with an Olympus BX41 microscope
under bright field (magnification, ×100). Histological grading was
referred to clinical standards (20).
Cell culture and transfection
Huh7 and HCCLM3 cells were purchased from Fudan Cell
Bank (Shanghai, China). Cells were maintained in DMEM (Gibco;
Thermo Fisher Scientific, Inc.) supplemented with 10% FBS (Gibco;
Thermo Fisher Scientific, Inc.) and 1% penicillin-streptomycin
solution (Gibco; Thermo Fisher Scientific, Inc.), and cultured at
37°C with 5% CO2.
Cell transfection with plasmids (HA-tagged IκBα,
Flag-tagged NF-κB p65 and corresponding empty vector as control;
all Shanghai GeneChem Co., Ltd.) or small interfering RNAs (siRNAs;
Shanghai Genepharma Co., Ltd.) was performed using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's instructions. In
total, 400,000 cells/well were seeded into 6-well plates 1 day
before transfection. Plasmids or siRNAs were diluted in Opti-MEM
(Gibco; Thermo Fisher Scientific, Inc.) before mixing with
Lipofectamine® 2000. Overexpression vector plasmids were
pcDNA3.1 (Shanghai GeneChem Co., Ltd.). Each well of the 6-well
plate was added with 2 µg plasmid for transfection. The sequences
of siRNA oligos were: siIκBα, 5′-CUCCGAGACUUUCGAGGAATT-3′; siNF-κB
p65, 5′-GAUUGAGGAGAAACGUAAAdTdT-3′; and siErbin,
5′-CACACUGUUGUAUGAUCAACCAU-3′. Each well of a 6-well plate was
added with 8 µl siRNA (20 µmol/l) for transfection. After 48 h,
cells were harvested for successive experiments.
Cell Counting Kit (CCK)-8
Cell proliferation was assessed using CCK-8
according to the instructions of the manufacturer (Bimake). Then,
24 h after transfection, cells were trypsinized and counted
manually. Cells (2,000-3,000 cells/well) were seeded into a 96-well
plate and after another 24 h, 10 µl CCK-8 solution was added into
each well. The cells were incubated at 37°C for another 2–4 h. A
spectrophotometer was then used to measure absorbance at 450
nm.
Clonogenic survival assay
Transfected cells in DMEM with 10% FBS were seeded
into 6-well plate, ~500 cells per well. After 10–14 days, cells
that formed visualized colonies were stained with 0.1% crystal
violet staining solution (Beijing Solarbio Science & Technology
Co., Ltd.) at room temperature for 30 min and images were captured.
Colonies with >50 cells were counted manually.
Transwell migration assay
To compare the difference of cell migration after
different transfection, 5×104−1.5×105
transfected cells were plated into an 8 µm-pore Transwell chamber,
fit for a 24-well plate, with serum free-DMEM. The lower section of
the chamber, a 24-well plate, contained normal medium with 10% FBS.
FBS acted as attractant to stimulate cell migrate through the
membrane of chambers. After incubating for 24 h, cells were fixed
by 4% paraformaldehyde for 15 min at room temperature and stained
with 0.1% crystal violet staining solution (Beijing Solarbio
Science & Technology Co., Ltd.) at room temperature for 30 min.
Three different fields were captured under bright field and cells
were counted manually (magnification, ×40).
RNA extraction and reverse
transcription-quantitative (RT-q)PCR
RNA extraction, reverse transcription (RT) and
real-time PCR were performed using RNAiso Plus, PrimeScript™ RT
reagent kit with gDNA Eraser (Perfect Real Time) and TB Green
Premix Ex Taq (Tli RNaseH Plus) (all Takara Bio, Inc.), following
the manufacturer's instructions. Temperature and duration for
reverse transcription were 37°C for 15 min and 85°C for 5 min. The
thermocycling conditions were as follows: Activation of enzyme at
95°C for 30 sec; 30 cycles of denaturation and annealing at 95°C
for 5 sec and 60°C for 30 sec. GAPDH was used as the reference
gene. The quantitative method to analyze relative gene expression
was 2−ΔΔCq (21). The
5′-3′sequences of primers used in RT-qPCR were as follows: GAPDH
forward, TGGCACCGTCAAGGCTGAGAA and reverse, TGGTGAAGACGCCAGTGGACTC;
NF-κB p65 forward, CGCTGCATCCACAGTTTCCA and reverse,
AGGGGTTGTTGTTGGTCTGG; Erbin forward, TGTGGGTGTGAAGACCTCAG and
reverse, GTCGCATCTCCGCCATTTTC; c-Rel forward GGTTGGTCCTGCCTCCTTAC,
and reverse, GCTGGAGTCCCAATGACGAA; and enhancer of zeste 2 polycomb
repressive complex 2 subunit (Ezh2) forward, GGACCACAGTGTTACCAGCAT
and reverse, GTGGGGTCTTTATCCGCTCAG.
Western blot analysis
Proteins were extracted using RIPA lysis buffer
(Beyotime Institute of Biotechnology) with complete protease
inhibitor cocktail tablet (Bimake). Protein concentration was
calculated by a BCA protein concentration kit (Beyotime Institute
of Biotechnology) and 5X SDS loading buffer (Shanghai Shenggong
Biology Engineering Technology Service, Ltd.) was added for
extraction. The samples were gently vortexed and boiled at 100°C
for 10 min before 30 µg was loaded onto 10% polyacrylamide gel
(EpiZyme, Inc.). Following electrophoresis, proteins were
transferred to nitrocellulose membranes. The membranes were blocked
in 5% non-fat milk solution at room temperature for at least 1 h,
incubated with primary antibodies at 4°C overnight and then
HRP-conjunct secondary antibodies at room temperature for 1 h.
Protein blots were detected using the super ECL kit (Nanjing KeyGen
Biotech Co., Ltd.). Antibodies used in this study were: Anti-IκBα
(sc-945, 1:600 dilution, Santa Cruz Biotechnology, Inc.);
anti-NF-κB p65 (#8242, 1:1,000 dilution, Cell Signaling Technology,
Inc.); anti-GAPDH (10494-1-AP, 1:5,000 dilution, ProteinTech group,
Inc.) and anti-Erbin (22438-1-AP, 1:500 dilution, ProteinTech
group, Inc.).
Statistics analysis
Data are presented as the mean ± SEM, which
represented ≥3 independent trials. Comparison between two different
groups was performed using unpaired Student's t-test. Comparison
among multiple groups was performed using one-way ANOVA and least
significance difference was used as the post hoc test.
χ2 test was used to detect associations between protein
expression and clinical characteristics. Survival analysis was
performed using Kaplan-Meier method and comparisons with the
log-rank test. Statistical analyses were performed using GraphPad
Prism 5 (GraphPad Software, Inc.) and SPSS software (22.0 version;
IBM Corp.). P<0.05 was considered to indicate a statistically
significant difference.
Results
IκBα expression is downregulated in
HCC and positively associated with poor prognosis of patients with
HCC
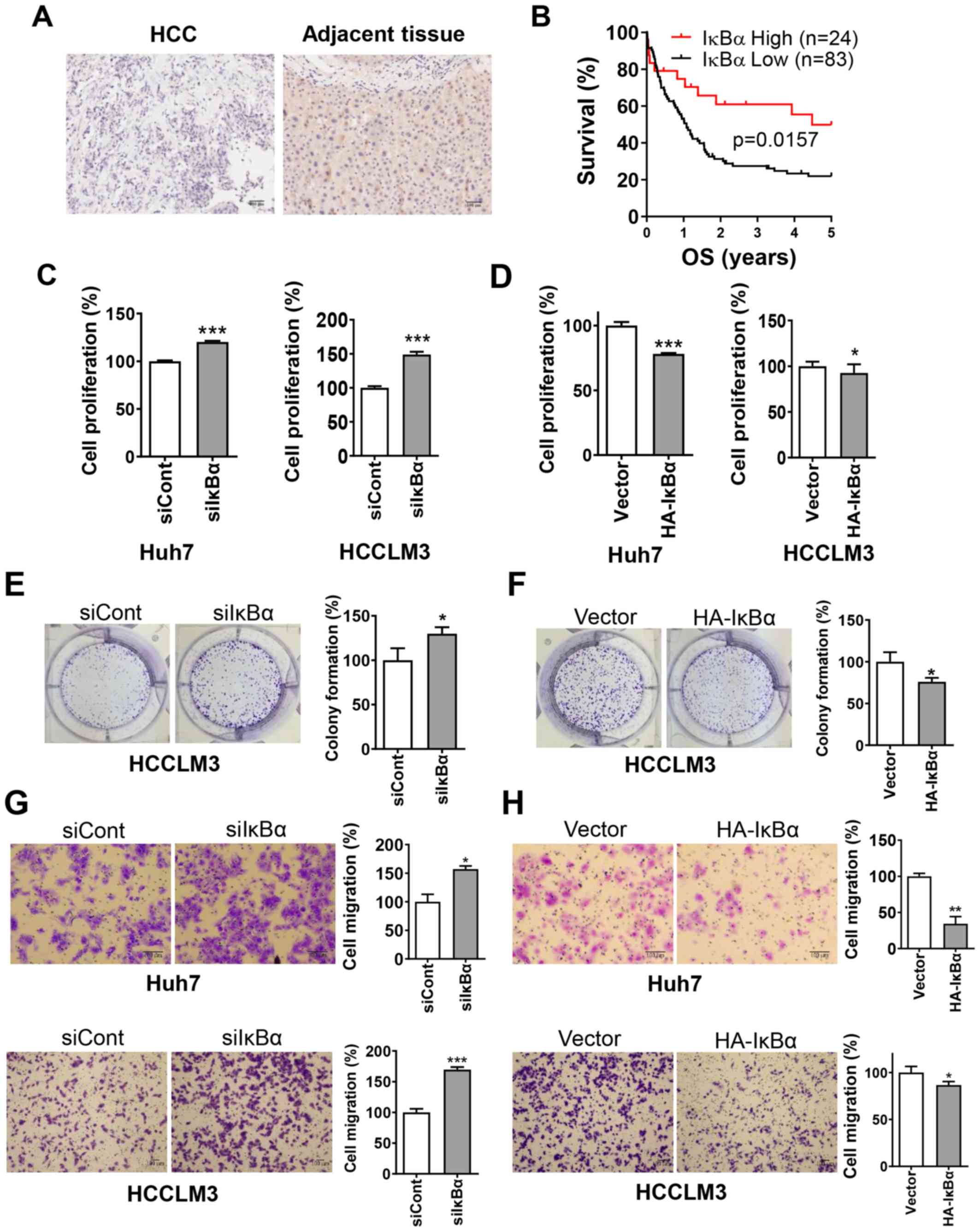
The results indicated that IκBα protein expression
was markedly decreased in HCC tissues compared with adjacent
tissues (Fig. 1A). In total, 77.6%
(83/107) of HCC cases exhibited low expression compared with the
matched peri-tumor tissues. Kaplan-Meier survival analysis
demonstrated that patients with HCC with low expression of IκBα had
a significantly shorter survival time compared with patients with
high expression (Fig. 1B). In
addition, the association between the expression of IκBα and cancer
recurrence was statistically significant (Table I).
 | Table I.Association between IκBα expression
and clinicopathological variables of 107 patients with HCC. |
Table I.
Association between IκBα expression
and clinicopathological variables of 107 patients with HCC.
|
| IκBα
expression |
|
|
|---|
|
|
|
|
|
|---|
| Parameter | High (n=24) | Low (n=83) | χ2 | P-value |
|---|
| Sex |
|
| 0.873 | 0.35 |
|
Male | 19 | 74 |
|
|
|
Female | 5 | 9 |
|
|
| Age, years |
|
| 0.97 | 0.325 |
|
≤50 | 17 | 49 |
|
|
|
>50 | 7 | 33 |
|
|
| TNM stage |
|
| 0.410 | 0.522 |
| I | 11 | 32 |
|
|
|
II–IV | 13 | 51 |
|
|
| Tumor thrombi |
|
| 1.706 | 0.192 |
|
Yes | 4 | 25 |
|
|
| No | 20 | 58 |
|
|
| Histological
grade |
|
| 0.001 | 0.98 |
| I | 3 | 8 |
|
|
|
II–III | 21 | 75 |
|
|
| Intrahepatic
metastasis |
|
| 0.363 | 0.547 |
|
Yes | 6 | 14 |
|
|
| No | 18 | 69 |
|
|
| Tumor
recurrence |
|
| 7.035 | 0.008a |
|
Yes | 10 | 59 |
|
|
| No | 14 | 24 |
|
|
To determine the tumor suppressive function of IκBα,
CCK-8, clonogenic survival and Transwell migration assays were used
to detect proliferation and migration of HCC cells. Knockdown of
IκBα significantly promoted cell proliferation and migration,
whereas overexpression of IκBα significantly inhibited cell
proliferation and migration (Fig.
1C-H). These findings suggested that IκBα was downregulated in
HCC tissues and that knockdown of IκBα was positively associated
with poor prognosis of patients with HCC.
IκBα negatively regulates Erbin
expression via NF-κB
Although it has been reported that elevated Erbin
promotes HCC tumorigenesis (18),
the underlying mechanism via which Erbin is upregulated in HCC
remains unknown. The results indicated that Erbin expression was
upregulated in the HCC tissues from 107 patients in southwest of
China (Fig. 2A). Kaplan-Meier
survival analysis demonstrated that high expression of Erbin was
positively associated with short survival time of patients with HCC
(Fig. 2B). Moreover, expression of
Erbin was associated with malignant characteristics of HCC, such as
TNM stage, existence of vascular tumor thrombi and cancer
recurrence (Table II). As both IκBα
and Erbin are associated with poor survival of patients with HCC,
it was determined whether there was an association between IκBα and
Erbin. It was found that knockdown of IκBα promoted Erbin protein
expression, while overexpression of IκBα reduced this expression
(Fig. 2C and D).
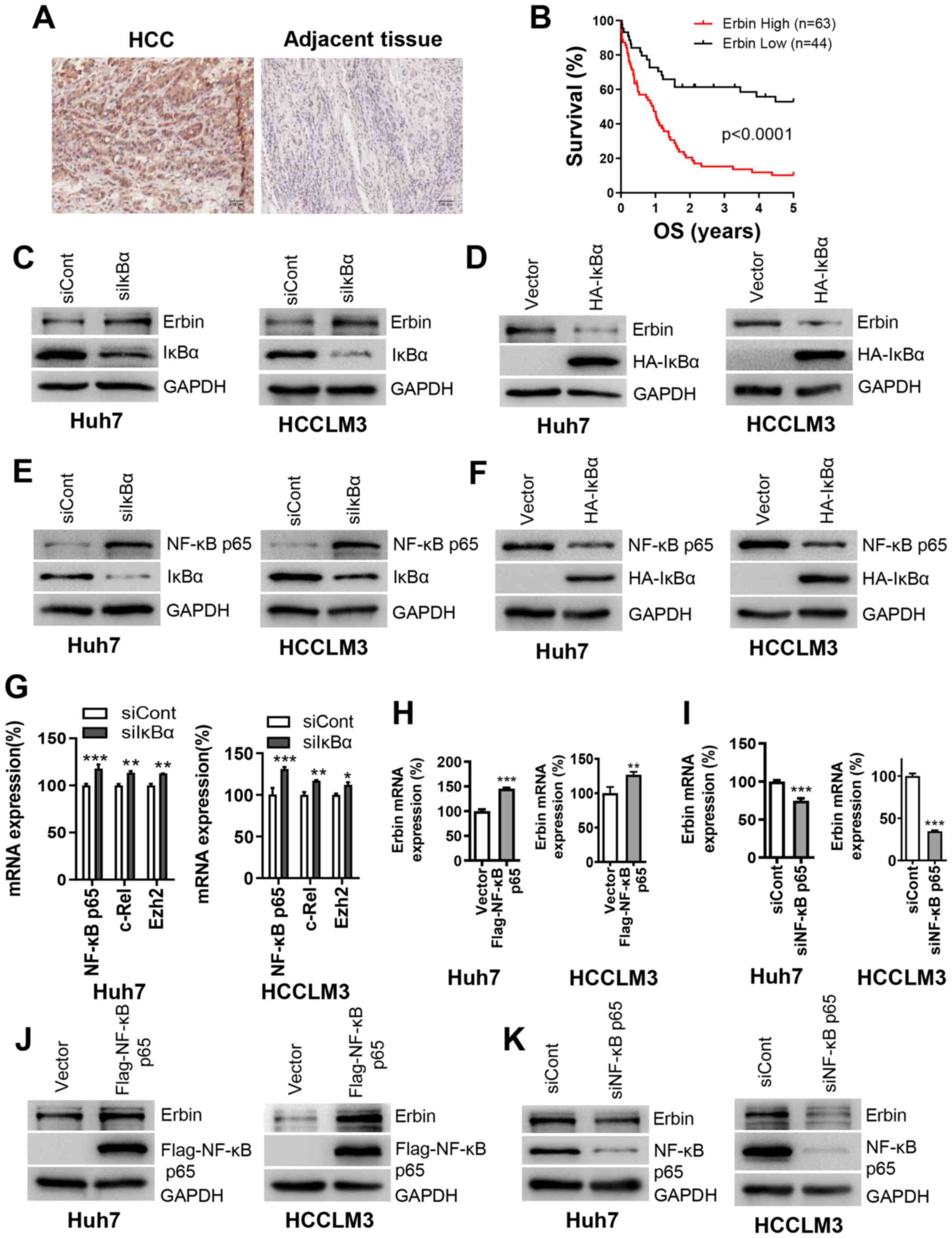 | Figure 2.IκBα negatively regulates Erbin
expression via NF-κB. (A) Representative images of HCC and adjacent
liver tissues from 107 cases stained with anti-Erbin antibody. (B)
Association between OS time of 107 patients with HCC and Erbin
expression. Cells were transfected with (C) siIκBα or (D) HA-IκBα
for 48 h and Erbin expression was determined using western blot
analysis. Cells were transfected with (E) siIκBα or (F) HA-IκBα for
48 h and NF-κB p65 expression was determined using western blot
analysis. (G) Cells were transfected with siIκBα for 48 h and mRNA
expression levels of c-Rel and Ezh2 were assessed using RT-qPCR.
Cells were transfected with (H) Flag-NF-κB p65 or (I) siNF-κB p65
for 48 h and then the mRNA expression of Erbin was measured using
RT-qPCR. Cells were transfected with (J) Flag-NF-κB p65 or (K)
siNF-κB p65 for 48 h, then Erbin expression was determined using
western blot analysis. Scale bar, 100 µm. The error bars represent
SEM from three independent experiments. *P<0.05, **P<0.01 and
***P<0.001 vs. corresponding control group. siRNA, small
interfering RNA; IκBα, NF-κB inhibitor α; Cont, control; HCC,
hepatocellular carcinoma; OS, overall survival; Erbin, Erbb2
interacting protein; RT-qPCR, reverse transcription-quantitative
PCR; Rel, Rel proto-oncogene, NF-kB subunit; Ezh2, enhancer of
zeste 2 polycomb repressive complex 2 subunit. |
| Table II.Association between Erbin expression
and clinicopathological variables of 107 patients with HCC. |
Table II.
Association between Erbin expression
and clinicopathological variables of 107 patients with HCC.
|
| Erbin
expression |
|
|
|---|
|
|
|
|
|
|---|
| Parameter | High (n=63) | Low (n=44) | χ2 | P-value |
|---|
| Sex |
|
| 0.195 | 0.659 |
|
Male | 54 | 39 |
|
|
|
Female | 9 | 5 |
|
|
| Age, years |
|
| 0.009 | 0.926 |
|
≤50 | 39 | 27 |
|
|
|
>50 | 24 | 16 |
|
|
| TNM stage |
|
| 4.541 | 0.033a |
| I | 20 | 23 |
|
|
|
II–IV | 43 | 21 |
|
|
| Tumor thrombi |
|
| 4.739 | 0.029a |
|
Yes | 22 | 7 |
|
|
| No | 41 | 37 |
|
|
| Histological
grade |
|
| 0.000 | 0.988 |
| I | 7 | 4 |
|
|
|
II–III | 56 | 40 |
|
|
| Intrahepatic
metastasis |
|
| 2.640 | 0.014 |
|
Yes | 15 | 5 |
|
|
| No | 48 | 39 |
|
|
| Tumor
recurrence |
|
| 9.165 | 0.002a |
|
Yes | 48 | 21 |
|
|
| No | 15 | 23 |
|
|
NF-κB/Rel transcription factors present in the
cytosol in an inactive state form a complex with inhibitory IκB
proteins (7). It has been suggested
that NF-κB p65 is released from combination with IκBα and nuclear
translocation occurs when IκBα is downregulated in HCC (6). The results indicated that knockdown of
IκBα increased NF-κB p65 expression (Fig. 2E), while overexpression of IκBα
inhibited NF-κB p65 expression (Fig.
2F). Subsequently, NF-κB activation upon IκB knockdown was
examined, as indicated by activation of canonical and non-canonical
transcriptional targets of NF-κB, c-Rel and Ezh2 after transfection
of siIκBα (Fig. 2G).
To assess whether the transcription factor NF-κB
regulates Erbin, the effect of knockdown or overexpression of NF-κB
on Erbin at the mRNA and protein levels was measured. In contrast
to IκBα, overexpression of NF-κB enhanced both mRNA and protein
expression levels of Erbin, whereas knockdown of NF-κB inhibited
these expression levels (Fig. 2H-K).
Collectively, the results indicated that reduced IκBα promoted
NF-κB-mediated Erbin expression at both mRNA and protein
levels.
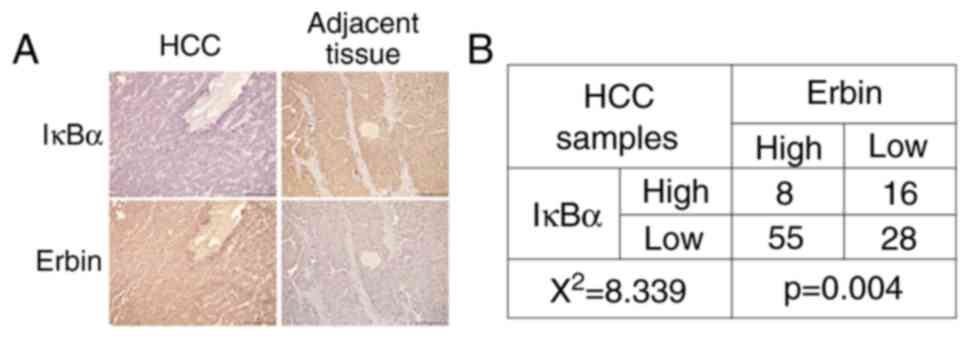
IκBα is negatively associated with
Erbin in patients with HCC
To further evaluate the association between IκBα and
Erbin, the expression levels of IκBα and Erbin were analyzed in 107
HCC tissues using IHC staining. It was determined that 77.6% of HCC
samples were IκBα-negative or low expression, demonstrating active
NF-κB signaling in the majority of cases (Fig. 3A and B). In line with this finding,
Erbin protein expression in 58.9% (63/107) of cases was upregulated
compared with the adjacent tissues. Thus, there was a negative
association between IκBα and Erbin expression levels in HCC tissues
(Fig. 3).
Erbin is responsible for
NF-κB-mediated HCC cell proliferation and migration
Based on the oncogenic function of NF-κB (22) and its positive regulation over Erbin,
it was evaluated whether Erbin was responsible for NF-κB-mediated
cell proliferation and migration. Overexpression of NF-κB
upregulated Erbin expression, and overexpression of Erbin exerted
oncogenic effects on the proliferation and migration of HCC cells.
However, knockdown of Erbin attenuated the aforementioned cellular
abilities (Fig. 4A-F). In addition,
increased proliferation and migration of HCC cells by NF-κB
overexpression was reversed by Erbin knockdown (Fig. 4G-I), indicating that NF-κB promoted
its activities via Erbin signaling. In healthy liver tissues,
expression of IκBα limited the activity of NF-κB (6) and, in turn, its downstream target Erbin
according to the present results. However, in HCC, NF-κB was
abnormally activated due to the absence of IκBα, and
transcriptionally upregulated expression of Erbin stimulated the
proliferation and migration of HCC cells.
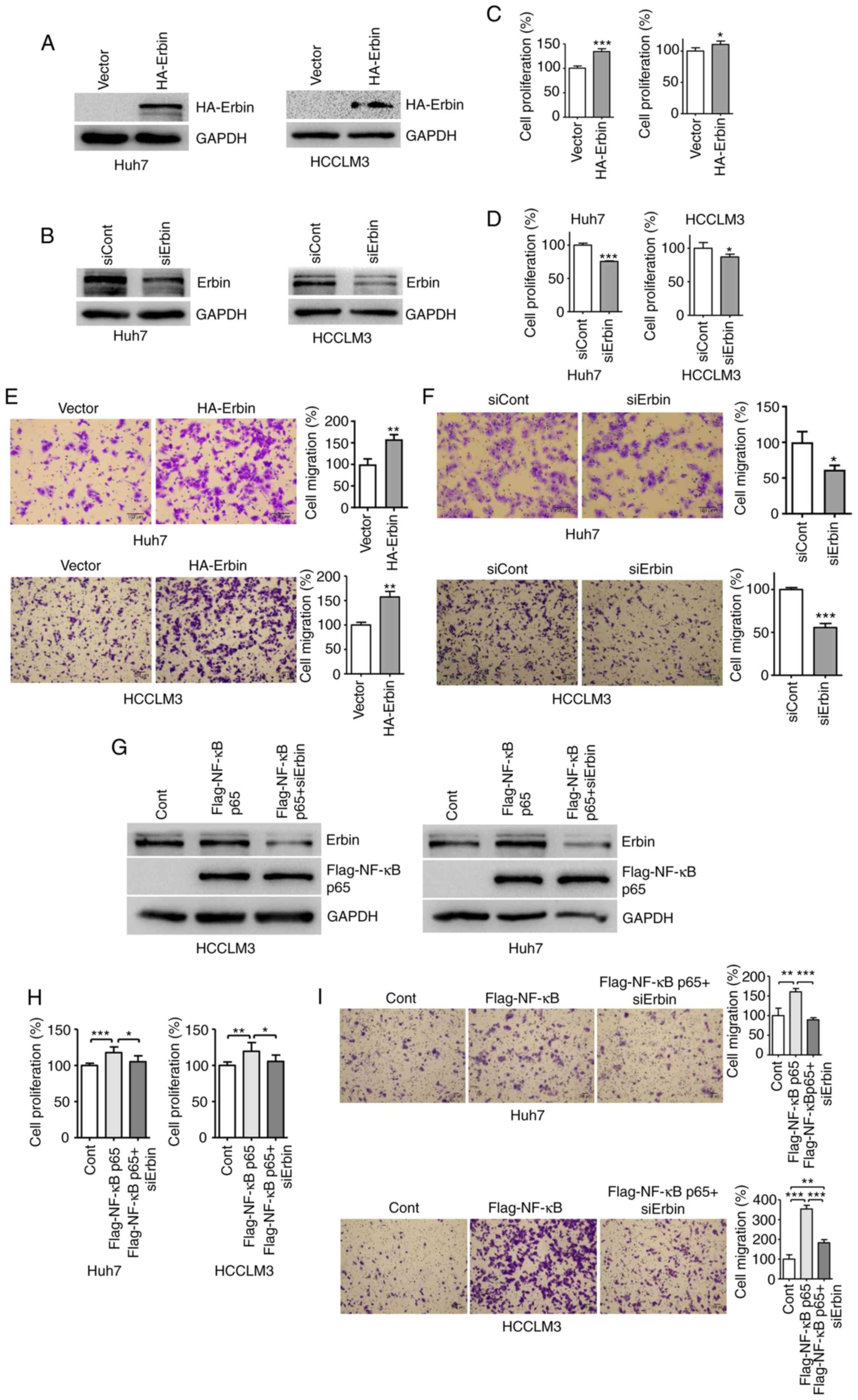 | Figure 4.Erbin is responsible for
NF-κB-mediated HCC cell proliferation and migration. Cells were
transfected with (A) HA-Erbin or (B) siErbin for 48 h and Erbin
expression was determined by western blotting. Cells were
transfected with (C) HA-Erbin or (D) siErbin and cell proliferation
was assessed after 24 h of transfection using CCK-8 assay. After
transfection with (E) HA-Erbin or (F) siErbin for 48 h, cells were
seeded into Transwell plate for 24 h and stained with 0.1% crystal
violet staining solution. Cells were transfected with Flag-NF-κB
p65, with or without Erbin siRNA, and analyzed using (G) western
blotting, (H) CCK-8 assay or (I) Transwell migration assay. Scale
bar, 100 µm. The error bars represent SEM from three independent
experiments. *P<0.05, **P<0.01 and ***P<0.001 vs.
corresponding control group. siRNA, small interfering RNA; IκBα,
NF-κB inhibitor α; Cont, control; Erbin, Erbb2 interacting protein;
CCK-8, Cell Counting Kit-8. |
Discussion
As HCC is a serious and lethal disease worldwide
(1), and thus it is important to
determine a novel therapeutic target. The present study identified
that in HCC tissues, there was low expression of IκBα, the
inhibitor of NF-κB, and decreased expression of IκBα promoted
cancer cell proliferation and migration. Once IκBα undergoes
proteasomal degradation, NF-κB activity is initiated (6), and positively affects cell
proliferation and migration by promoting the expression of Erbin.
In the current study, consistent with previous reports of increased
Erbin expression in HCC tissues obtained from Shanghai (18), the results indicated Erbin expression
was upregulated in HCC tissues from 107 patients in southwest of
China and exerted oncogenic functions. Moreover, downregulation of
IκBα and upregulation of Erbin were closely associated with poor
prognosis of patients with HCC. Therefore, the present results
demonstrated the clinical importance of IκBα and Erbin, identified
a novel association between IκBα/NF-κB and indicated the role of
the downstream oncoprotein Erbin in the promotion of HCC
tumorigenesis.
IκBα is considered to be a tumor-suppressing factor
in different types of cancer, such as breast (23,24),
ovarian (25), gastric (26) and colorectal (27), mainly due to its inhibition of NF-κB
activity; however, the underlying mechanism is yet to be
elucidated. Liver tumorigenesis is usually attributed to
inflammation (28). Cell
proliferation increases during inflammation, in which accelerated
DNA replication results in the increased occurrence of oncogenic
mutation (29). Moreover, NF-κB, as
a hallmark of inflammation, not only participates in the
initiation, but also maintains inflammation via blocking apoptosis,
together with TNFα to activate anti-apoptosis genes (30). Thus, NF-κB serves a crucial role in
promoting inflammation and contributing to malignancy (31).
Erbin has been reported to exert a range of diverse
functions. For example, Erbin suppresses the activity of the
Ras/Raf pathway to inhibit proliferation and stop
epithelial-mesenchymal transition progress in colorectal cancer
(32). Deficiency of Erbin
significantly promotes ERK phosphorylation in ERBB2-overexpressed
breast cancer (33). Furthermore,
accumulation of Erbin attenuates Raf activation to halt autophagy
and senescence in KRAS proto-oncogene, GTPase-induced skin
tumorigenesis (34). Erbin also
exerts a pro-tumorigenesis role by negatively regulating ERα
(18). Therefore, these findings
suggest that the function of Erbin depends closely on cell types or
a certain cancer microenvironment. The current study demonstrated a
novel signaling via which NF-κB promotes tumorigenesis. It was
indicated that in HCC, Erbin is highly expressed and associated
with poor prognosis of patients. In HCC cells, it was identified
that NF-κB activated the expression of Erbin at both the mRNA and
protein levels, and overexpression of Erbin enhanced cell
proliferation and migration, while its knockdown reduced these
abilities.
In conclusion, the present study identified that: i)
IκBα was decreased in HCC and associated with poor survival of
patients with HCC; ii) knockdown of IκBα upregulated NF-κB
signaling to promote HCC cell proliferation and migration; and iii)
NF-κB-mediated Erbin upregulation contributes to HCC cell
proliferation and migration and a poor outcome of patients with
HCC. Therefore, the present results indicated the important
clinical implications of IκBα and Erbin, as well as the novel
NF-κB/Erbin oncogenic pathway. Thus, disruption of this signaling
pathway may be a novel strategy to slow the progression of HCC.
Acknowledgements
Not applicable.
Funding
This work was supported by the Introduction of
Special Funds for Talents from Army Medical University (grant no.
4174C6), Natural Science Foundation of Chongqing (grant no.
cstc2019jcyjmsxmX0519) and the Natural Science Foundation (grant
no. 4241212I62).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
CMX designed and supervised the study. YT, XTL, YDL,
JZ, LF, YYZ, HQY and LDZ performed the experiments. YT, YYZ, HQY
and CMX contributed to immunohistochemistry analysis and
pathological analysis. YJ and LS contributed to the clinical sample
collection, fixing and embedding, and pathological information
processing. YT, PB and CMX analyzed and interpreted the data. YT
and CMX drafted the manuscript. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
Southwest Hospital. The informed consent forms were obtained from
the patients with HCC.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
HCC
|
hepatocellular carcinoma
|
|
IκBα
|
NF-κB inhibitor α
|
|
IKK
|
IκB kinase
|
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Negro F: Natural history of NASH and HCC.
Liver Int. 40 (Suppl 1):S72–S76. 2020. View Article : Google Scholar
|
|
3
|
Nahon P, Vibert E, Nault JC, Ganne-Carrie
N, Ziol M and Seror O: Optimizing curative management of
hepatocellular carcinoma. Liver Int. 40 (Suppl 1):S109–S115. 2020.
View Article : Google Scholar
|
|
4
|
Refolo MG, Messa C, Guerra V, Carr BI and
D'Alessandro R: Inflammatory mechanisms of HCC development. Cancers
(Basel). 12:6412020. View Article : Google Scholar
|
|
5
|
Mohs A, Kuttkat N, Reissing J, Zimmermann
HW, Sonntag R, Proudfoot A, Youssef SA, de Bruin A, Cubero FJ and
Trautwein C: Functional role of CCL5/RANTES for HCC progression
during chronic liver disease. J Hepatol. 66:743–753. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hayden MS and Ghosh S: Shared principles
in NF-kappaB signaling. Cell. 132:344–362. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hoffmann A, Levchenko A, Scott ML and
Baltimore D: The IkappaB-NF-kappaB signaling module: Temporal
control and selective gene activation. Science. 298:1241–1245.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen J and Chen ZJ: Regulation of NF-κB by
ubiquitination. Curr Opin Immunol. 25:4–12. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li Q and Verma IM: NF-kappaB regulation in
the immune system. Nat Rev Immunol. 2:725–734. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li Y, Lu L, Tu J, Zhang J, Xiong T, Fan W,
Wang J, Li M, Chen Y, Steggerda J, et al: Reciprocal regulation
between forkhead Box M1/NF-kB and methionine adenosyltransferase 1A
drives liver cancer. Hepatology (Baltimore, Md.). Feb 21–2020.doi:
10.1002/hep.31196. Online ahead of print. View Article : Google Scholar
|
|
11
|
Ghosh S and Karin M: Missing pieces in the
NF-kappaB puzzle. Cell. 109 (Suppl):S81–S96. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gupta SC, Awasthee N, Rai V, Chava S,
Gunda V and Challagundla KB: Long non-coding RNAs and nuclear
factor-κB crosstalk in cancer and other human diseases. Biochim
Biophys Acta Rev Cancer. 1873:1883162020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Borg JP, Marchetto S, Le Bivic A,
Ollendorff V, Jaulin-Bastard F, Saito H, Fournier E, Adélaïde J,
Margolis B and Birnbaum D: ERBIN: A basolateral PDZ protein that
interacts with the mammalian ERBB2/HER2 receptor. Nat Cell Biol.
2:407–414. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dan L, Shi M, Duan H, Han C and Guo N:
Erbin, a negative regulator in diverse signal pathways. Curr
Protein Pept Sci. 11:759–764. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huang H, Song Y, Wu Y, Guo N, Ma Y and
Qian L: Erbin loss promotes cancer cell proliferation through
feedback activation of Akt-Skp2-p27 signaling. Biochem Biophys Res
Commun. 463:370–376. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Huang YZ, Zang M, Xiong WC, Luo Z and Mei
L: Erbin suppresses the MAP kinase pathway. J Biol Chem.
278:1108–1114. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hu Y, Chen H, Duan C, Liu D, Qian L, Yang
Z, Guo L, Song L, Yu M, Hu M, Shi M and Guo N: Deficiency of Erbin
induces resistance of cervical cancer cells to anoikis in a
STAT3-dependent manner. Oncogenesis. 2:e522013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wu H, Yao S, Zhang S, Wang JR, Guo PD, Li
XM, Gan WJ, Mei L, Gao TM and Li JM: Elevated expression of Erbin
destabilizes ERalpha protein and promotes tumorigenesis in
hepatocellular carcinoma. J Hepatol. 66:1193–1204. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
McDonald C, Chen FF, Ollendorff V, Ogura
Y, Marchetto S, Lécine P, Borg JP and Nuñez G: A role for Erbin in
the regulation of Nod2-dependent NF-kappaB signaling. J Biol Chem.
280:40301–40309. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Department of Medical Administration,
National Health Commission of the People's Republic of China:
Guidelines for diagnosis and treatment of primary liver cancer in
China (2019 edition). Zhonghua Gan Zang Bing Za Zhi. 28:112–128.
2020.(In Chinese). PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dolcet X, Llobet D, Pallares J and
Matias-Guiu X: NF-κB in development and progression of human
cancer. Virchows Arch. 446:475–482. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu B, Sun L, Liu Q, Gong C, Yao Y, Lv X,
Lin L, Yao H, Su F, Li D, et al: A cytoplasmic NF-κB interacting
long noncoding RNA blocks IκB phosphorylation and suppresses breast
cancer metastasis. Cancer Cell. 27:370–381. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ma C, Zuo W, Wang X, Wei L, Guo Q and Song
X: Lapatinib inhibits the activation of NF-κB through reducing
phosphorylation of IκB-α in breast cancer cells. Oncol Rep.
29:812–818. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ataie-Kachoie P, Badar S, Morris DL and
Pourgholami MH: Minocycline targets the NF-κB Nexus through
suppression of TGF-β1-TAK1-IκB signaling in ovarian cancer. Mol
Cancer Res. 11:1279–1291. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rhoads MG, Kandarian SC, Pacelli F,
Doglietto GB and Bossola M: Expression of NF-kappaB and IkappaB
proteins in skeletal muscle of gastric cancer patients. Eur J
Cancer. 46:191–197. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lu YX, Ju HQ, Wang F, Chen LZ, Wu QN,
Sheng H, Mo HY, Pan ZZ, Xie D, Kang TB, et al: Inhibition of the
NF-κB pathway by nafamostat mesilate suppresses colorectal cancer
growth and metastasis. Cancer Lett. 380:87–97. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Feld JJ and Krassenburg LAP: What comes
first: Treatment of viral hepatitis or liver cancer? Dig Dis Sci.
64:1041–1049. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Clevers H: At the crossroads of
inflammation and cancer. Cell. 118:671–674. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Pikarsky E, Porat RM, Stein I, Abramovitch
R, Amit S, Kasem S, Gutkovich-Pyest E, Urieli-Shoval S, Galun E and
Ben-Neriah Y: NF-kappaB functions as a tumour promoter in
inflammation-associated cancer. Nature. 431:461–466. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Elsharkawy AM and Mann DA: Nuclear
factor-kappaB and the hepatic inflammation-fibrosis-cancer axis.
Hepatology. 46:590–597. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Stevens PD, Wen YA, Xiong X, Zaytseva YY,
Li AT, Wang C, Stevens AT, Farmer TN, Gan T, Weiss HL, et al: Erbin
suppresses KSR1-mediated RAS/RAF signaling and tumorigenesis in
colorectal cancer. Cancer Res. 78:4839–4852. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu D, Shi M, Duan C, Chen H, Hu Y, Yang
Z, Duan H and Guo N: Downregulation of Erbin in Her2-overexpressing
breast cancer cells promotes cell migration and induces trastuzumab
resistance. Mol Immunol. 56:104–112. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xie CM, Wei D, Zhao L, Marchetto S, Mei L,
Borg JP and Sun Y: Erbin is a novel substrate of the Sag-βTrCP E3
ligase that regulates KrasG12D-induced skin tumorigenesis. J Cell
Biol. 209:721–737. 2015. View Article : Google Scholar : PubMed/NCBI
|