Introduction
Cervical cancer is one of the most common
gynecological malignancies, and it has become a crucial public
health problem (1). Globally, the
incidence of cervical cancer ranks second among all female
malignant tumors (2). At present,
surgery, chemotherapy and radiotherapy are the preferred treatments
for cervical cancer. However, chemotherapy drugs cannot achieve a
good therapeutic effect in cervical cancer due to drug resistance.
In addition, effective treatments for advanced and recurrent
cervical cancer are lacking (3).
However, targeted therapy for cervical cancer has been well
recognized. Specific molecular targeted drugs for tumors have
gradually been identified, such as drugs that target signal
transduction, cell receptors and angiogenesis (4,5). To the
best of our knowledge, the exact mechanism of cervical cancer
development is rarely studied, which greatly limits the
investigation of molecular targeted drug therapy. Therefore, it is
necessary to further study the molecular mechanism of cervical
cancer development.
With the progress of human genome sequencing
technology, >98% of sequences in the human genome are found to
be non-coding RNAs (6) Non-coding
RNAs are classified into short and long non-coding RNAs (lncRNAs)
based on their sequence lengths (7).
lncRNAs are >200 nucleotides in length and their structure is
similar to mRNAs (8,9). lncRNA is ubiquitous in eukaryotes,
however its sequence is poorly conserved among different species
(10). lncRNAs can form complex
secondary structures that provide space for binding to multiple
nucleic acids or proteins (11).
Functionally, lncRNA regulates gene expression at the
transcriptional and post-transcriptional level (12). lncRNAs are also involved in a variety
of cellular activities, such as signaling pathway regulation, gene
expression regulation, protein folding and cell activity (13). Studies have also reported that
lncRNAs are closely associated various tumors, such as colorectal
cancer and breast cancer (14–16).
Glucose-regulated protein 78 (GRP78) is a
multifunctional protein that is mainly distributed in the
endoplasmic reticulum (17). GRP78
acts as a molecular chaperone that controls protein folding and
assembly, prevents protein aggregation and regulates the
endoplasmic reticulum unfolded protein response pathway (18). As an essential stress sensor, GRP78
expression is altered by various factors in the tumor
microenvironment, such as hypoxia, glucose and nutrient deficiency,
acidosis, and inflammatory responses (19). A number of studies have shown that
GRP78 overexpression promotes proliferation, migration, invasion
and anti-apoptosis of cancer cells (20,21).
GRP78-specific antibody is capable of inhibiting tumor growth and
metastasis by neutralizing GRP78 protein level (22). Our previous studies demonstrated that
GRP78 is upregulated in cervical cancer, and GRP78-knockdown could
increase the sensitivity of chemotherapy drugs and improve the
cisplatin-induced apoptosis of cervical cancer cells (23,24).
In the present study, The Cancer Genome Atlas (TCGA)
and Gene Expression Omnibus (GEO) databases were used to screen
target lncRNAs that were associated with GRP78 in cervical cancer.
Subsequent experiments were performed to investigate the underlying
mechanism.
Materials and methods
Data collection
Two cervical cancer expression microarrays GSE26511
(https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=gse26511)
and GSE5787 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi) were
downloaded from the GEO database. The GSE26511 microarray contains
20 cases of cervical cancer without lymph node metastasis and 19
cases of cervical cancer with lymph node metastasis (25). GSE5787 contain 30 tumor samples from
11 patients with cervical cancer (26). TCGA http://cancergenome.nih.gov/) contained gene
expression profile data of cervical cancer. All data from TCGA were
downloaded in December 2017, and consisted of data for a total of
306 cervical cancer samples.
Data processing
Correlation analysis was performed between GRP78 and
lncRNAs extracted from GSE5787, GSE26511 and TCGA with the Cor R
package by R software. The top 200 lncRNAs with positive and
negative correlation with GRP78 in GSE26511, GSE5787 and TCGA were
extracted and analyzed in a Venn diagram (https://bioinfogp.cnb.csic.es/tools/venny/). Gene Set
Enrichment Analysis (GSEA) was conducted using GSEA 2.2.1 software
(27,28). The downloaded expression profile,
phenotype data and MsigDB microarray platform file were uploaded
into the GSEA program. Enrichment analysis was performed according
to the default weighted enrichment statistics method.
Cell culture and transfection
Human cervical cancer cell lines HeLa and SiHa were
obtained from American Type Culture Collection. HeLa and SiHa cells
were cultured in Dulbecco's modified Eagle's medium (Hyclone;
Cytiva) containing 10% fetal bovine serum (Hyclone; Cytiva), 100
U/ml penicillin and 100 µg/ml streptomycin (Hyclone; Cytiva). Cells
were incubated in a 5% CO2 incubator at 37°C.
For transfection, HeLa were seeded in the 6-well
plates, and when the confluence was 60–80%, the cells were
transfected with 50 nM small interfering RNAs (siRNAs and si-NC
used as control) using Lipofectamine 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
The culture medium was replaced 6 h later. After 24 h, the
transfected cells were harvested for subsequent experiments. The
sequences of siRNA used were as follows: si-negative control (NC)
sense, 5′-UUCUCCGAACGUGUCACGUTT-3′ and antisense,
5′-ACGUGACACGUUCGGAGAATT-3′; si-GRP78 sense,
5′-GGAGCGCAUUGAUACUAGATT-3′ and antisense,
5′-UCUAGUAUCAAUGCGCUCCTT-3′; si-LINC00294-1 sense,
5′-CCACAAAGUUAUCAGGAAATT-3′ and antisense,
5′-UUUCCUGAUAACUUUGUGGTT-3′; si-LINC00294-2 sense,
5′-CCUGGAAUCUCAUAGGAUUTT-3′ and antisense,
5′-AAUCCUAUGAGAUUCCAGGTT-3′; and si-LINC00294-3 sense,
5′-GCGAACAUGUAACCCUCUATT-3′ and antisense,
5′-UAGAGGGUUACAUGUUCGCTT-3′.
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from transfected cells using
TRIzol (Invitrogen; Thermo Fisher Scientific, Inc.) and reversed
transcribed using PrimeScript RT Master mix (Takara Bio, Inc.)
according to the manufacturer's protocol. RNA concentration was
detected using a spectrometer and samples with an A260/A280 ratio
of 1.8–2.0 were selected for the following qPCR reaction. qPCR was
then performed using SYBR® Green Master mix (Takara Bio,
Inc.), according to the manufacturer's protocol. RT-qPCR was
carried out at 94°C for 5 min and 40 cycles at 94°C for 30 sec,
55°C for 30 sec and 72°C for 90 sec following the protocol of SYBR
Premix Ex Taq™ (Takara). The primers used for qPCR were as follows:
GAPDH forward, 5′-CACCCACTCCTCCACCTTTG-3′ and reverse,
5′-CCACCACCCTGTTGCTGTAG-3′; HLA complex 5 forward,
5′-CTGTGGATGACATGGCCTTA-3′ and reverse, 5′-GATGCCAGCTTTGAGTGGA-3′;
HOXA transcript antisense RNA (HOTAIR), myeloid specific 1 forward,
5′-AGTGCTGGAGCGAAGAAGAG-3′ and reverse, 5′-CCTCTCGCCAGTTCATCTTT-3′;
HOXA11 antisense RNA forward, 5′-CGGCTAACAAGGAGATTTGG-3′ and
reverse, 5′-GCAAAGGCTGTGGAAAGAAG-3′; LINC00162 forward,
5′-CTCAGAAACACCCACCATGC-3′ and reverse, 5′-GAAAGTCCAGGCAGTTCAGC-3′;
LINC00294 forward, 5′-TGTGTTGTCCTCCAGAATCG-3′ and reverse,
5′-CCAACCAAGAGCCAACAAAG-3′; LINC00888 forward,
5′-TTGGCCCTTGAAAGAATCAG-3′ and reverse, 5′-ACTGGCATTTCCTCCACTGT-3′;
ST3GAL4 antisense RNA 1 forward, 5′-ACCTTCCTGCTGTCCTTCCT-3′ and
reverse, 5′-CCCTCAGAGCCTTGATGTCT-3′; TOPORS antisense RNA 1
forward, 5′-CTCTCACCTCCTGTTGTACGC-3′ and reverse,
5′-CGTGGTCTGTTTAGGGAGGA-3′; P16 forward, 5′-TTATTTGAGCTTTGGTTCTG-3′
and reverse, 5′-CCGGCTTTCGTAGTTTTCAT-3′; P21 forward,
5′-GAGTCTCCAGGTCCACCTGG-3′ and reverse, 5′-CGTGGTCTGTTTAGGGAGGA-3′;
cyclin D forward, 5′-ATGTTCGTGGCCTCTAAGATGA-3′ and reverse,
5′-CAGGTTCCACTTGAGCTTGTTC-3′; cyclin E forward,
5′-GTTATAAGGGAGACGGGGAG-3′, and reverse,
5′-TGCTCTGCTTCTTACCGCTC-3′; and cyclin-dependent kinase 4 (CDK4)
forward, 5′-CATGTAGACCAGGACCTAAGG-3′, and reverse,
5′-AACTGGCGCATCAGATCCTAG-3′. Relative gene expression was analyzed
using 2−ΔΔCt method (29).
Cell cycle detection
HeLa and SiHa cells were digested with trypsin
(Gibco; Thermo Fisher Scientific, Inc.) and prepared into a cell
suspension. After cells were washed with Hanks buffer (Gibco;
Thermo Fisher Scientific, Inc.), cells were centrifuged at 1,000 ×
g for 5 min at room temperature. Cells were then resuspended and
incubated with pre-cooled 70% ethanol overnight. Finally, cells
were stained with propidium iodide (30 µg/ml) for 30 min at room
temperature (Beyotime Institute of Biotechnology), followed by cell
cycle detection using a flow cytometer (BD LSRFFortessa; BD
Biosciences). The results were analyzed using FlowJo software
(version 10.6.2; BD Biosciences).
Western blotting
Total protein was extracted from treated cells using
RIPA solution at 4°C (Beyotime Institute of Biotechnology) and the
protein concentration was quantified using BCA kit (Beyotime
Institute of Biotechnology). Protein samples (60 µg) were separated
by 10% SDS gel with electrophoresis and transferred to a PVDF
membrane. Membranes were blocked with 5% skimmed milk for 1 h at
room temperature, followed by incubation with the primary
antibodies against GRP78 (1:1,000; cat. no. 3177T), p16 (1:1,000;
cat. no. 80772), p21 (1:1,000; cat. no. 2947), Cyclin D (1:1,000;
cat. no. 55506), Cyclin E (1:1,000; cat. no. 4129), CDK4 (1:1,000;
cat. no. 12790), Gli1 (1:1,000; cat. no. 3538T), Sonic (1:1,000;
cat. no. 2207T) and GAPDH (1:1,000; cat. no. 5174T; Cell Signaling
Technology, Inc.) overnight at 4°C. Subsequently, the membranes
were incubated with anti-rabbit IgG HRP-linked antibody (1:1,000;
cat. no. 7074; Cell Signaling Technology, Inc.) at room temperature
for 1 h. The protein blots on the membrane were exposed by
chemiluminescence reagent (Thermo Fisher Scientific, Inc.).
Relative expression levels were normalized to endogenous control
GAPDH using Image J software (1.52a; National Institutes of
Health).
Statistical analysis
All experiments were performed in triplicate. SPSS
20.0 statistical software (IBM Corp.) was used for statistical
analysis, and GraphPad 5.0 (GraphPad Software, Inc.) was used to
generate figures. Quantitative data are expressed as mean ±
standard deviation. Comparisons among multiple groups were
performed by one-way analysis of variance. Comparisons among four
groups were analyzed by one-way analysis of variance followed by
Tukey's post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
GRP78 regulates LINC00294
expression
Cervical cancer gene expression profile data were
downloaded from GEO and TCGA databases. Specifically, GSE26511
contained 39 cervical cancer samples, GSE5787 contained 30 cervical
cancer samples and TCGA contained 306 cervical cancer samples.
Subsequently, correlation analysis between the identified lncRNAs
and GRP78 was performed, the top 200 lncRNAs that were positively
correlated with GRP78 and the top 200 lncRNAs that were negatively
correlated with GRP78 were selected. The overlapping lncRNAs that
were positively correlated with GRP78 in the GSE26511, GSE5787 and
TCGA datasets were obtained from the Venn diagram. A total of eight
overlapping lncRNAs were finally screened out to be positively
correlated with GRP78 (Fig. 1A).
Whereas, only one lncRNA was negatively correlated with GRP78 in
all datasets (Fig. 1B). These eight
positively corrected lncRNAs (LINC00294, HCP5, HOTAIRM1, HOXA11-AS,
LINC00162, LINC00888) were selected for the following
experiments.
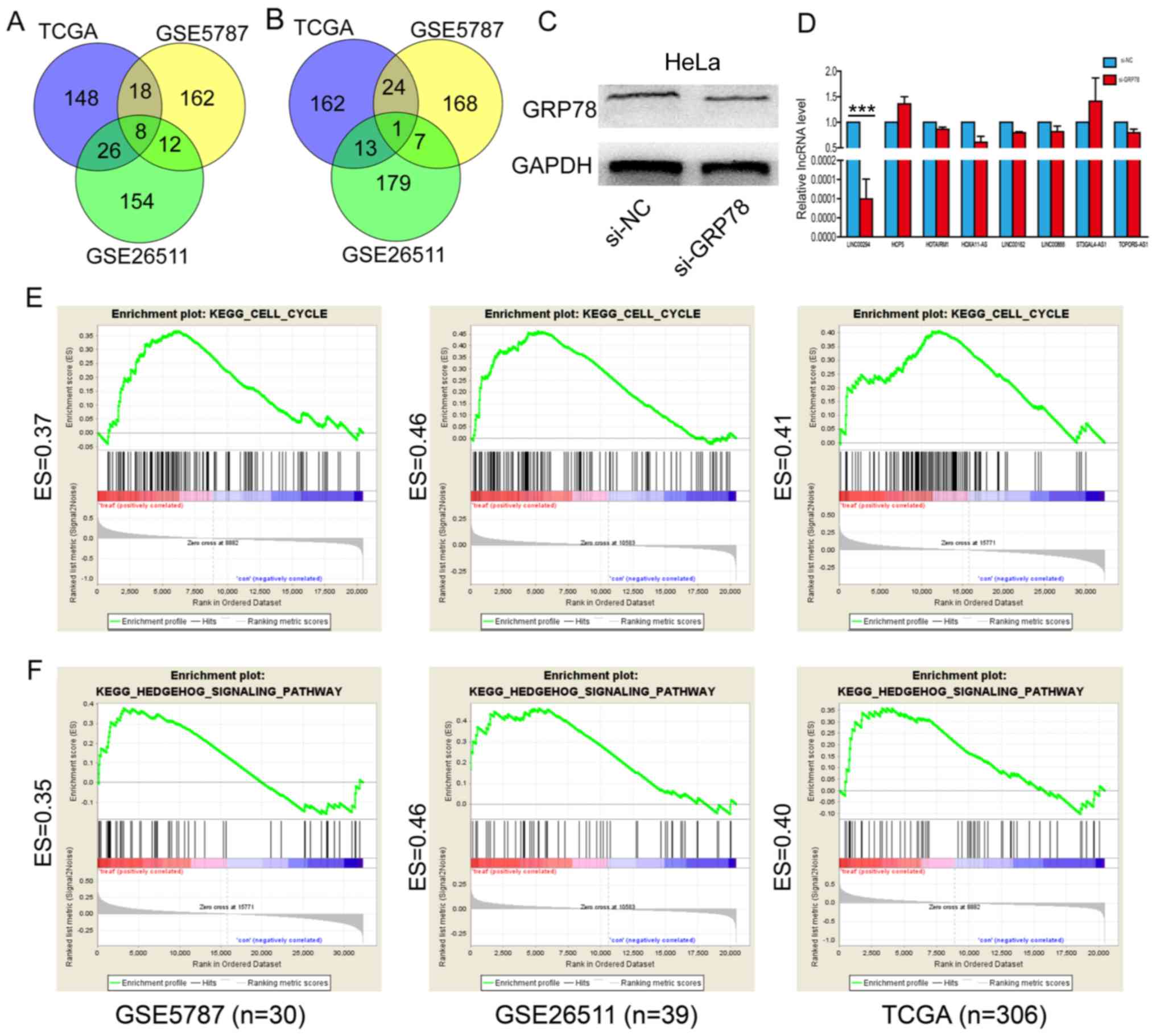 | Figure 1.GRP78 regulates LINC00294 expression.
(A) The top 200 lncRNAs that were positively correlated with GRP78
in the GSE26511, GSE5787 and TCGA datasets. (B) The top 200 lncRNAs
that were negatively correlated with GRP78 in the GSE26511, GSE5787
and TCGA datasets. (C) The GRP78 protein expression level was
decreased by transfection with si-GRP78. (D) LINC00294 expression
was decreased following GRP78-knockdown. (E) LINC00294 is
associated with the cell cycle according to GSEA. (F) GSEA
demonstrated that LINC00294 is enriched in the Hedgehog pathway.
***P<0.001. GRP78, glucose-regulated protein 78; lncRNA, long
non-coding RNA; TCGA, The Cancer Genome Atlas; si, small
interfering RNA; NC, negative control; HCP5, HLA complex 5;
HOTAIRM1, HOXA transcript antisense RNA, myeloid specific 1;
HOXA11-AS, HOXA11 antisense; ST3GAL4-AS1, ST3GAL4 antisense RNA 1;
TOPORs-AS1, TOPORS antisense RNA 1; ES, enrichment score; KEGG,
Kyoto Encyclopedia of Genes and Genomes; GSEA, Gene Set Enrichment
Analysis. |
For the following experiments, si-GRP78 was
transfected into HeLa cells and the expression levels of the eight
positively correlated lncRNAs were detected. Western blotting was
used to confirm that GRP78 was successfully reduced in HeLa cells
transfected with si-GRP78 compared with cells transfected with the
si-NC (Fig. 1C). Subsequently, the
eight positively corrected lncRNAs were confirmed by RT-qPCR
following GRP78 knockdown. The results demonstrated that only
LINC00294 was downregulated following GRP78-knockdown in HeLa cells
(Fig. 1D). Subsequently, GSEA was
performed to predict the biological processes that LINC00294 is
associated with. The results demonstrated that the LINC00294 is
associated with the cell cycle (Fig.
1E) and Hedgehog signaling pathway (Fig. 1F). These results indicated that
LINC00294 may participate in cervical cancer development via
GRP78.
LINC00294-knockdown arrests the cell
cycle of cervical cancer cells
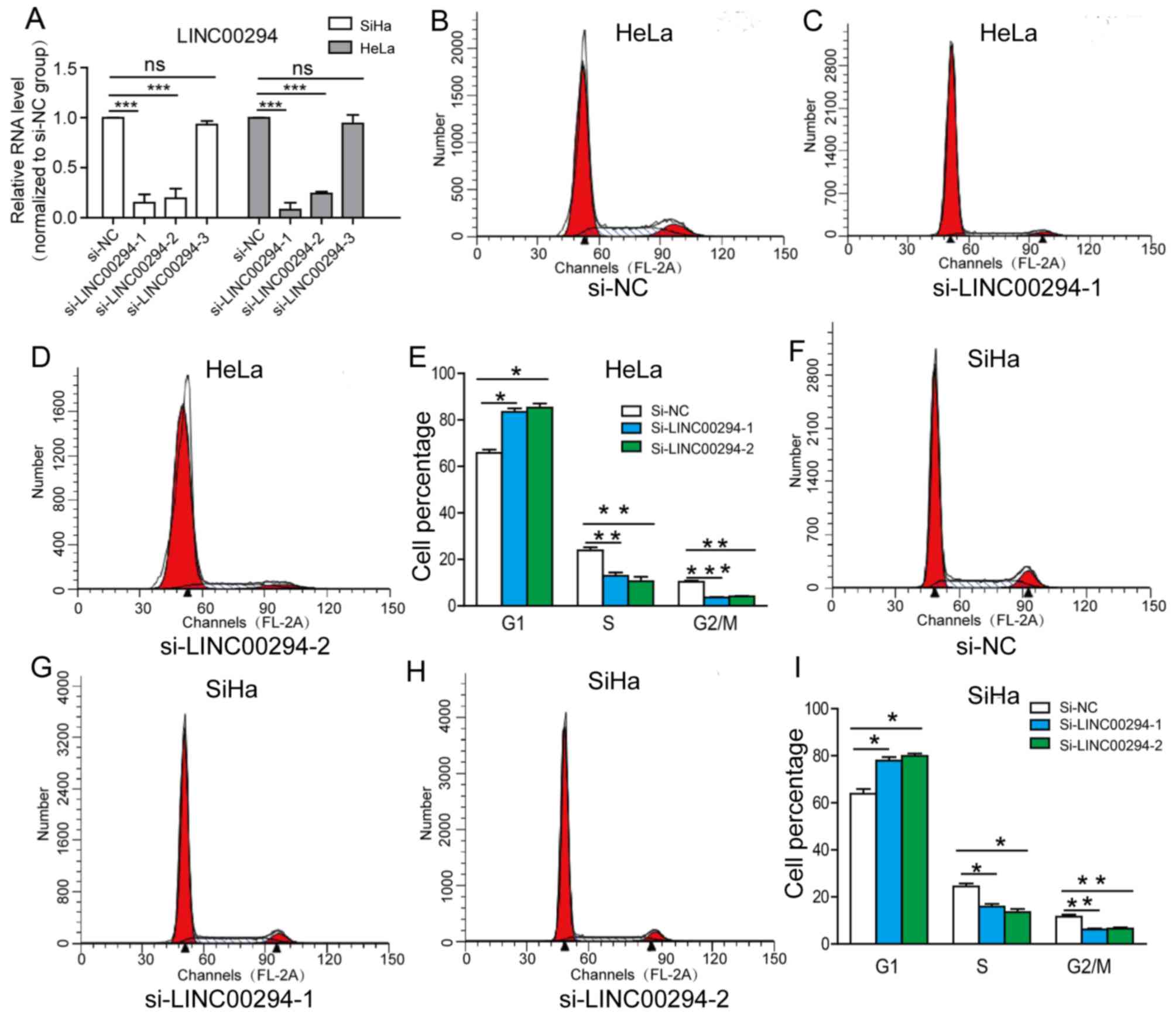
To investigate whether LINC00294 can regulate the
cell cycle of HeLa and SiHa cells, three siRNA sequences targeting
LINC00294 were transfected into the cells. It was identified that
si-LINC00294-1 and si-LINC00294-2 significantly reduced the mRNA
level of LINC00294, whereas the efficacy of si-LINC00294-3 was
insignificant (Fig. 2A). Following
transfection of HeLa and SiHa cells with si-LINC00294-1 and
si-LINC00294-2, the percentage of cells in the G0/G1 phase was
significantly increased, whereas the percentages of cells in the S
and G2/M phases were significantly decreased (Fig. 2B-D). These results demonstrated that
LINC00294-knockdown arrests the cell cycle at the G0/G1 phase,
thereby inhibiting the proliferative ability of cervical cancer
cells.
LINC00294 regulates the cell cycle of
cervical cancer cells via the Hedgehog pathway
Expression levels of cell cycle-related genes were
detected by RT-qPCR and western blotting. The results indicated
that following LINC00294-knockdown in HeLa and SiHa cells the mRNA
levels of p16 and p21 were significantly increased compared with
the control cells, whereas the mRNA levels of cyclin D, cyclin E
and CDK4 were significantly decreased (Fig. 3A). Similar results were obtained by
western blotting (Fig. 3B and C).
Based on GSEA data, the Hedgehog pathway was predicted to be
involved in cervical cancer development. Therefore, key genes in
Hedgehog pathway were detected by western blotting. Protein
expression levels of Sonic and Gli1 were significantly decreased
following transfection with si-LINC00294 (Fig. 3D and E), indicating a role of the
Hedgehog pathway in regulating the cell cycle of cervical cancer
cells.
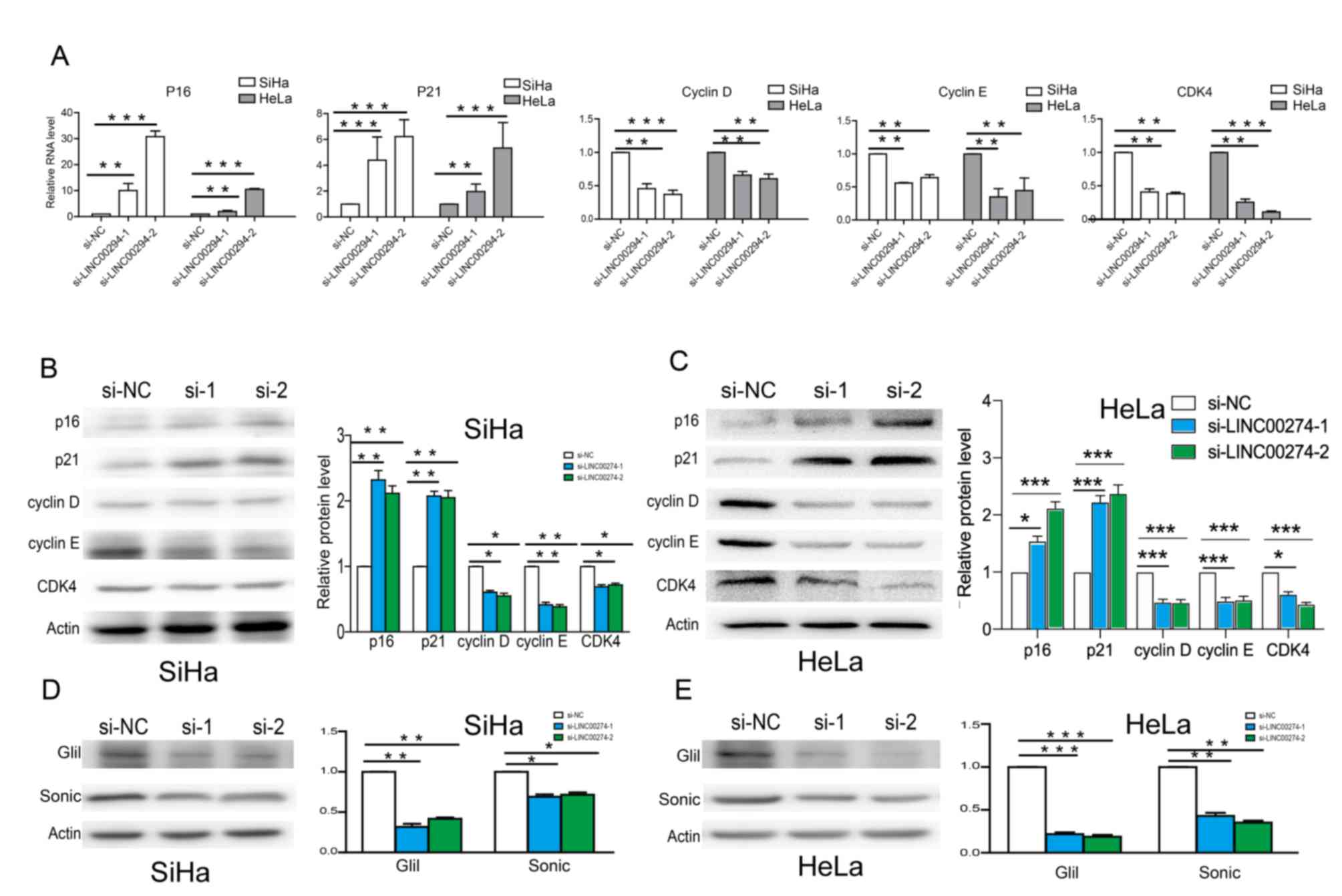 | Figure 3.LINC00294 regulates the cell cycle
via the Hedgehog pathway. (A) The mRNA levels of p16, p21, cyclin
D, cyclin E and CDK in HeLa and SiHa cells that were transfected
with si-LINC00294. The protein levels of p16, p21, cyclin D, cyclin
E and CDK in (B) SiHa and (C) HeLa cells following
LINC00294-knockdown. The protein levels of Sonic and Gli1 in (D)
SiHa and (E) HeLa cells following LINC00294-knockdown. *P<0.05,
**P<0.01, ***P<0.001. si, small interfering RNA; NC, negative
control; CDK4, cyclin-dependent kinase 4; Gli1, GLI family zinc
finger 1. |
Discussion
Cervical cancer is one of the most common types of
malignant tumor in women, and it is a continuous process from a
benign lesion, cervical intraepithelial neoplasia, carcinoma in
situ and invasive carcinoma (30). Therefore, early detection and
treatment are essential to reduce the morbidity and mortality of
cervical cancer (31). Whole genome
sequencing results have demonstrated that the majority of the
stably transcribed RNAs are non-coding RNAs (32). Among them, lncRNAs possess functions
in multiple cellular processes, including cell proliferation,
differentiation and metabolism (33). lncRNAs are involved in various
diseases, such as tumors, diabetes and immune diseases (34,35). For
example, overexpressed HOTAIR in cervical cancer is associated with
lymph node metastasis, survival rate and postoperative recurrence
(36). Maternally expressed 3 is
downregulated in cervical cancer tissues, which promotes
proliferation of cervical cancer (37). In addition, decreased expression of
lncRNA growth arrest specific 5 can serve as an unfavorable
prognostic factor for cervical cancer (38). Our previous study demonstrated that
GRP78 is upregulated in cervical cancer tissues compared with
paracancerous tissues. In the present study, LINC00294 was
identified to be positively associated with GRP78 in. In
vitro experiments demonstrated that LINC00294 may regulate the
cell cycle of cervical cancer cells via the Hedgehog pathway.
Cell cycle disorder is one of the main causes of
tumorigenesis (39). Under normal
circumstances, the cell cycle transitions between the G1, S, G2 and
M phases via precise regulation by the cell cycle molecular network
system of cyclins, CDKs and cyclin-depending kinase inhibitors
(CKIs) (40). Specifically, cyclin
D1, cyclin E, CDK4, p16 and p21 are key factors that regulate the
cell cycle (39,40). At different phases of the cell cycle,
cyclins bind to the corresponding CDKs to form cyclin/CDK
complexes, which activate CDKs and promote cell cycle
transformation. However, CDKs or cyclin/CDK complexes also inhibit
CDK activities via binding to the corresponding CKIs, thereby
inhibiting the transition of the cell cycle (39,40). The
present study demonstrated that LINC00294-knockdown in cervical
cancer significantly arrests cell cycle in G0/G1 phase, thereby
inhibiting cell cycle progression and cell proliferation. Western
blotting demonstrated that LINC00294-knockdown downregulated cyclin
D, cyclin E and CDK4, whereas it upregulated p16 and p21 in HeLa
and SiHa cells.
The Hedgehog gene was first discovered in
drosophila in 1980 (41).
Previous studies have reported that the Hedgehog pathway plays a
key role in animal embryonic development, including in the
formation of lungs, skin, bones, limbs, neural tube and the
gastrointestinal system (42–44). In
addition, the Hedgehog pathway is involved in the regulation of
cell growth, proliferation, migration and differentiation (45,46).
Hedgehog is closely associated with the occurrence and progression
of malignant tumors, such as basal cell carcinoma, breast cancer,
prostate cancer and multiple digestive system cancers (47,48).
Abnormal activation of the Hedgehog pathway results in invasion and
metastasis of tumor cells (49–51). In
the present study, the Hedgehog pathway was found to be involved in
the occurrence of cervical cancer. Preliminary mechanism studies
demonstrated that LINC00294-knockdown inhibited the expression
levels of Gli1 and Sonic, which are key genes in the Hedgehog
pathway.
In summary, the present study first identified
LINC00294 to be strongly correlated with GRP78. Subsequently,
RT-qPCR demonstrated that LINC00294 was regulated by GRP78. In
addition, GSEA revealed that LINC00294 was mainly enriched in
mediating the cell cycle and the Hedgehog pathway. An in
vitro assay confirmed that LINC00294 could regulate the cell
cycle. Furthermore, western blotting demonstrated that following
knockdown of LINC00294, the CKD family was significantly
downregulated and CDK were inhibitors upregulated. Additionally,
Hedgehog pathway-associated proteins were also expressed at lower
levels following knockdown of LINC00294. The key findings of the
present study were that LINC00294 was regulated by GRP78 and
knockdown of LINC00294 could arrest the cell cycle at the G0/G1
phrase. However, there were some limitations of the study. In order
to further confirm the regulatory relationship between GRP78 and
LINC00294, whether overexpression of GRP78 can increase the
expression of LINC00294 should be investigated, as well as the
effects of LINC00294 on cell proliferation, invasion and migration
using in vivo experiments, which will be performed in future
studies. In conclusion, LINC00294 is positively correlated with and
regulated by GRP78, which promotes the progression of cervical
cancer through arresting the cell cycle at the G0/G1 phase via the
Hedgehog pathway.
Acknowledgements
Not applicable.
Funding
This study was supported by the Jiangsu Maternal and
Child Health Key Program (grant no. F2012110).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JQ and SZ performed the experiments and collected
the data. WC analyzed the data. CL conceived and designed the
experiments, and wrote the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nunes de Arruda F, da Costa S, Bonadio R,
Dornellas A, Pereira D, de Bock GH and Del Pilar Estevez Diz M:
Quality of life of locally advanced cervical cancer patients after
neoadjuvant chemotherapy followed by chemoradiation versus
chemoradiation alone (CIRCE trial): A randomized phase II trial.
Int J Gynecol Cancer. 30:749–756. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Diaz-Padilla I, Monk BJ, Mackay HJ and
Oaknin A: Treatment of metastatic cervical cancer: Future
directions involving targeted agents. Crit Rev Oncol Hematol.
85:303–314. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zagouri F, Sergentanis TN, Chrysikos D,
Filipits M and Bartsch R: Molecularly targeted therapies in
cervical cancer. A systematic review. Gynecol Oncol. 126:291–303.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Djebali S, Davis CA, Merkel A, Dobin A,
Lassmann T, Mortazavi A, Tanzer A, Lagarde J, Lin W, Schlesinger F,
et al: Landscape of transcription in human cells. Nature.
489:101–108. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Esteller M: Non-coding RNAs in human
disease. Nat Rev Genet. 12:861–874. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Marques AC and Ponting CP: Catalogues of
mammalian long noncoding RNAs: Modest conservation and
incompleteness. Genome Biol. 10:R1242009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ørom UA, Derrien T, Beringer M, Gumireddy
K, Gardini A, Bussotti G, Lai F, Zytnicki M, Notredame C, Huang Q,
et al: Long noncoding RNAs with enhancer-like function in human
cells. Cell. 143:46–58. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kutter C, Watt S, Stefflova K, Wilson MD,
Goncalves A, Ponting CP, Odom DT and Marques AC: Rapid turnover of
long noncoding RNAs and the evolution of gene expression. PLoS
Genet. 8:e10028412012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Perez DS, Hoage TR, Pritchett JR,
Ducharme-Smith AL, Halling ML, Ganapathiraju SC, Streng PS and
Smith DI: Long, abundantly expressed non-coding transcripts are
altered in cancer. Hum Mol Genet. 17:642–655. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hung T and Chang HY: Long noncoding RNA in
genome regulation: Prospects and mechanisms. RNA Biol. 7:582–585.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yan Y, Zhang L, Jiang Y, Xu T, Mei Q, Wang
H, Qin R, Zou Y, Hu G, Chen J and Lu Y: LncRNA and mRNA interaction
study based on transcriptome profiles reveals potential core genes
in the pathogenesis of human glioblastoma multiforme. J Cancer Res
Clin Oncol. 141:827–838. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liao Q, He W, Liu J, Cen Y, Luo L, Yu C,
Li Y, Chen S and Duan S: Identification and functional annotation
of lncRNA genes with hypermethylation in colorectal cancer. Gene.
572:259–265. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Richards EJ, Zhang G, Li ZP, Permuth-Wey
J, Challa S, Li Y, Kong W, Dan S, Bui MM, Coppola D, et al: Long
non-coding RNAs (LncRNA) regulated by transforming growth factor
(TGF) β: LncRNA-hit-mediated TGFβ-induced epithelial to mesenchymal
transition in mammary epithelia. J Biol Chem. 290:6857–6867. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang R, Du L, Yang X, Jiang X, Duan W, Yan
S, Xie Y, Zhu Y, Wang Q, Wang L, et al: Identification of long
noncoding RNAs as potential novel diagnosis and prognosis
biomarkers in colorectal cancer. J Cancer Res Clin Oncol.
142:2291–2301. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ramirez MU, Hernandez SR, Soto-Pantoja DR
and Cook KL: Endoplasmic reticulum stress pathway, the unfolded
protein response, modulates immune function in the tumor
microenvironment to impact tumor progression and therapeutic
response. Int J Mol Sci. 21:1692019. View Article : Google Scholar
|
|
18
|
Bánhegyi G, Baumeister P, Benedetti A,
Dong D, Fu Y, Lee AS, Li J, Mao C, Margittai E, Ni M, et al:
Endoplasmic reticulum stress. Ann N Y Acad Sci. 1113:58–71. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li Z and Li Z: Glucose regulated protein
78: A critical link between tumor microenvironment and cancer
hallmarks. Biochim Biophys Acta. 1826:13–22. 2012.PubMed/NCBI
|
|
20
|
Misra UK, Payne S and Pizzo SV: Ligation
of prostate cancer cell surface GRP78 activates a proproliferative
and antiapoptotic feedback loop: A role for secreted
prostate-specific antigen. J Biol Chem. 286:1248–1259. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huang P, Guo Y, Zhao Z, Ning W, Wang H, Gu
C, Zhang M, Qu Y, Zhang H and Song Y: UBE2T promotes glioblastoma
invasion and migration via stabilizing GRP78 and regulating EMT.
Aging (Albany NY). 12:10275–10289. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Misra UK, Mowery Y, Kaczowka S and Pizzo
SV: Ligation of cancer cell surface GRP78 with antibodies directed
against its COOH-terminal domain up-regulates p53 activity and
promotes apoptosis. Mol Cancer Ther. 8:1350–1362. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Luo C, Fan W, Jiang Y, Zhou S and Cheng W:
Glucose-related protein 78 expression and its effects on
cisplatin-resistance in cervical cancer. Med Sci Monit.
24:2197–2209. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Luo C and Qiu J: miR-181a inhibits
cervical cancer development via downregulating GRP78. Oncol Res.
25:1341–1348. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Noordhuis MG, Fehrmann RS, Wisman GB,
Nijhuis ER, van Zanden JJ, Moerland PD, van Themaat EVL, Volders
HH, Kok M, ten Hoor KA, et al: Involvement of the TGF-beta and
beta-catenin pathways in pelvic lymph node metastasis in
early-stage cervical cancer. Clin Cancer Res. 17:1317–1330. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bachtiary B, Boutros PC, Pintilie M, Shi
W, Bastianutto C, Li JH, Schwock J, Zhang W, Penn LZ, Jurisica I,
et al: Gene expression profiling in cervical cancer: An exploration
of intratumor heterogeneity. Clin Cancer Res. 12:5632–5640. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mootha VK, Lindgren CM, Eriksson KF,
Subramanian A, Sihag S, Lehar J, Puigserver P, Carlsson E,
Ridderstråle M, Laurila E, et al: PGC-1alpha-responsive genes
involved in oxidative phosphorylation are coordinately
downregulated in human diabetes. Nat Genet. 34:267–273. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Subramanian A, Tamayo P, Mootha VK,
Mukherjee S, Ebert BL, Gillette MA, Paulovich A, Pomeroy SL, Golub
TR, Lander ES and Mesirov JP: Gene set enrichment analysis: A
knowledge-based approach for interpreting genome-wide expression
profiles. Proc Natl Acad Sci USA. 102:15545–15550. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–440. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Polman NJ, Ebisch RMF, Heideman DAM,
Melchers WJG, Bekkers RLM, Molijn AC, Meijer CJLM, Quint WGV,
Snijders PJF, Massuger LFAG, et al: Performance of human
papillomavirus testing on self-collected versus clinician-collected
samples for the detection of cervical intraepithelial neoplasia of
grade 2 or worse: A randomised, paired screen-positive,
non-inferiority trial. Lancet Oncol. 20:229–238. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Halaska M, Robova H, Pluta M and Rob L:
The role of trachelectomy in cervical cancer.
Ecancermedicalscience. 9:5062015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mercer TR, Dinger ME and Mattick JS: Long
non-coding RNAs: Insights into functions. Nat Rev Genet.
10:155–159. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yuan L, Xu ZY, Ruan SM, Mo S, Qin JJ and
Cheng XD: Long non-coding RNAs towards precision medicine in
gastric cancer: Early diagnosis, treatment, and drug resistance.
Mol Cancer. 19:962020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lee C and Kikyo N: Strategies to identify
long noncoding RNAs involved in gene regulation. Cell Biosci.
2:372012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Novikova IV, Hennelly SP and Sanbonmatsu
KY: Sizing up long non-coding RNAs: Do lncRNAs have secondary and
tertiary structure? Bioarchitecture. 2:189–199. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kim HJ, Lee DW, Yim GW, Nam EJ, Kim S, Kim
SW and Kim YT: Long non-coding RNA HOTAIR is associated with human
cervical cancer progression. Int J Oncol. 46:521–530. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Qin R, Chen Z, Ding Y, Hao J, Hu J and Guo
F: Long non-coding RNA MEG3 inhibits the proliferation of cervical
carcinoma cells through the induction of cell cycle arrest and
apoptosis. Neoplasma. 60:486–492. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cao S, Liu W, Li F, Zhao W and Qin C:
Decreased expression of lncRNA GAS5 predicts a poor prognosis in
cervical cancer. Int J Clin Exp Pathol. 7:6776–6783.
2014.PubMed/NCBI
|
|
39
|
Murphy PJ and Campbell SS: Physiology of
the circadian system in animals and humans. J Clin Neurophysiol.
13:2–16. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lim S and Kaldis P: Cdks, cyclins and
CKIs: Roles beyond cell cycle regulation. Development.
140:3079–3093. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Nusslein-Volhard C and Wieschaus E:
Mutations affecting segment number and polarity in
Drosophila. Nature. 287:795–801. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
van Dop WA, Rosekrans SL, Uhmann A, Jaks
V, Offerhaus GJ, van den Bergh WM, Kasper M, Heijmans J, Hardwick
JC, Verspaget HW, et al: Hedgehog signalling stimulates precursor
cell accumulation and impairs epithelial maturation in the murine
oesophagus. Gut. 62:348–357. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhang L, Yang Y, Liao Z, Liu Q, Lei X, Li
M, Saijilafu, Zhang Z, Hong D, Zhu M, et al: Genetic and
pharmacological activation of Hedgehog signaling inhibits
osteoclastogenesis and attenuates titanium particle-induced
osteolysis partly through suppressing the JNK/c-Fos-NFATc1 cascade.
Theranostics. 10:6638–6660. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sun Q, Zeng J, Liu Y, Chen J, Zeng QC,
Chen YQ, Tu LL, Chen P, Yang F and Zhang M: microRNA-9 and −29a
regulate the progression of diabetic peripheral neuropathy via
ISL1-mediated sonic hedgehog signaling pathway. Aging (Albany NY).
12:11446–11465. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Bufalieri F, Severini LL, Caimano M,
Infante P and Di Marcotullio L: DUBs activating the hedgehog
signaling pathway: A promising therapeutic target in cancer.
Cancers (Basel). 12:15182020. View Article : Google Scholar
|
|
46
|
Lecca S, Namboodiri VMK, Restivo L,
Gervasi N, Pillolla G, Stuber GD and Mameli M: Heterogeneous
habenular neuronal ensembles during selection of defensive
behaviors. Cell Rep. 31:1077522020. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wu J, Zhu P, Lu T, Du Y, Wang Y, He L, Ye
B, Liu B, Yang L, Wang J, et al: The long non-coding RNA LncHDAC2
drives the self-renewal of liver cancer stem cells via activation
of Hedgehog signaling. J Hepatol. 70:918–929. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Hanna A and Shevde LA: Hedgehog signaling:
Modulation of cancer properies and tumor mircroenvironment. Mol
Cancer. 15:242016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang K, Pan L, Che X, Cui D and Li C:
Sonic Hedgehog/GLI(1) signaling pathway inhibition restricts cell
migration and invasion in human gliomas. Neurol Res. 32:975–980.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Onishi H, Kai M, Odate S, Iwasaki H,
Morifuji Y, Ogino T, Morisaki T, Nakashima Y and Katano M: Hypoxia
activates the hedgehog signaling pathway in a ligand-independent
manner by upregulation of Smo transcription in pancreatic cancer.
Cancer Sci. 102:1144–1150. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Karhadkar SS, Bova GS, Abdallah N, Dhara
S, Gardner D, Maitra A, Isaacs JT, Berman DM and Beachy PA:
Hedgehog signalling in prostate regeneration, neoplasia and
metastasis. Nature. 431:707–712. 2004. View Article : Google Scholar : PubMed/NCBI
|