Introduction
Gallbladder cancer (GBC) has the highest incidence
among malignant tumors of the biliary tract system, ranking fifth
among the common malignant diseases of the digestive system
worldwide (1), with an
age-standardized rate (ASR) of ~2.2 per 100,000 in 2012 (2). Due to the difficulty of early
diagnosis, rapid invasion and metastasis are common, while
treatment strategies are limited, resulting in patients with GBC
having a poor prognosis (3,4). Therefore, the development of an
effective agent that can prolong the survival of patients with GBC
is urgently required. There is a long history of the use of plants
to treat cancer. Studies have revealed a vast chemical diversity in
numerous plants, which remain a notable source of antitumor drugs
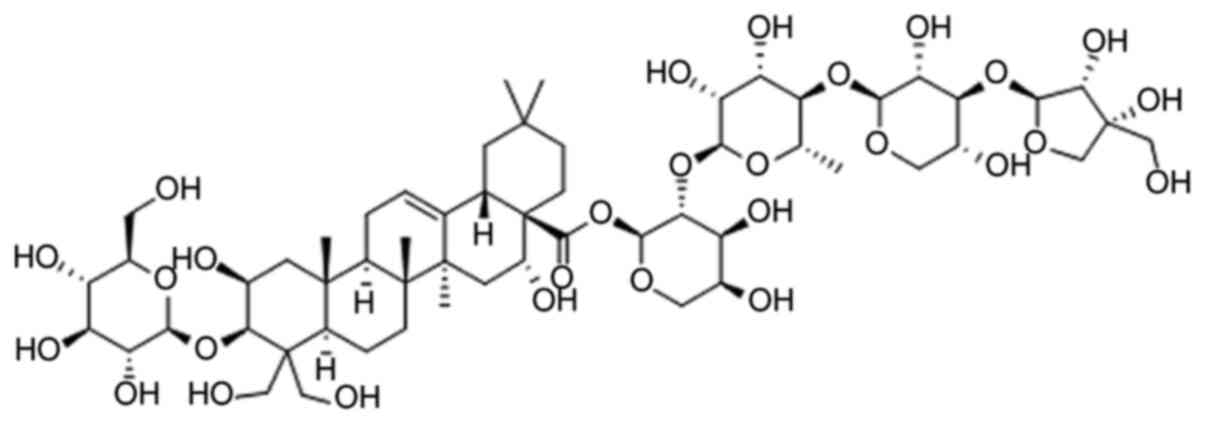
(5–8). Platycodin D (PD; Fig. 1) is a triterpenoid saponin extracted
from Platycodonis (9). Previous
studies have revealed that PD is a promising anticancer compound
that can inhibit various cancer cell lines, including prostate
cancer (10), glioma (11), leukemia (12), liver cancer (13), breast cancer (14) and gastric carcinoma (15), through various mechanisms including
proliferation and metastasis inhibition (16,17),
apoptosis induction (10), cell
cycle arrest (12) and autophagy
(18). However, the effect of PD on
GBC cell lines remains unknown. Therefore, the present study aimed
to explore the antitumor activity of PD on GBC cells and to
determine its potential molecular mechanisms of action, with the
goal of providing a promising drug for the future treatment of
GBC.
Materials and methods
Chemicals and reagents
PD (>98%, PubChem CID:132399081) was purchased
from Shanghai Yuanye Biotechnology Co., Ltd. It was dissolved in
PBS to obtain a stock solution (100 µmol/l), then stored at −20°C.
MTT and Hoechst 33342 were purchased from Sigma-Aldrich; Merck
KGaA. FITC Annexin V Apoptosis Detection kit I was purchased from
BD Biosciences. Primary antibodies against cleaved caspase-9
(1:1,000; cat. no. 9505), cleaved caspase-3 (1:1,000; cat. no.
9661), Bax (1:1,000; cat. no. 5023), Bcl-2 (1:1,000; cat. no.
4223), SAPK (stress-activated protein kinase)/JNK (1:1,000; cat.
no. 9252), phosphorylated-SAPK/JNK (p-JNK; 1:1,000; cat. no. 9251),
matrix metalloproteinase (MMP)-2 (1:1,000; cat. no. 87809), MMP-9
(1:1,000; cat. no. 3852), cyclin B1 (1:1,000; cat. no. 4138),
cytochrome c (1:1,000; cat. no. 4272) and β-tubulin
(1:1,000; cat. no. 2146), and all secondary antibodies (1:1,000;
cat. no. 7074) were purchased from Cell Signaling Technology, Inc.
The same secondary antibody was used for all primary antibodies.
The antibody against cyclin-dependent kinase 1 (CDK1; 1:1,000; cat.
no. ab133327) was purchased from Abcam.
Cell lines and cell culture
The human GBC NOZ, GBC-SD and SGC-996 cell lines
were obtained from the The Cell Bank of Type Culture Collection of
the Chinese Academy of Sciences. The cells were maintained in DMEM
supplemented with 10% FBS (both Gibco; Thermo Fisher Scientific,
Inc.), 100 µg/ml streptomycin and 100 U/ml penicillin. All cells
were cultured at 37°C in a humidified atmosphere with 5%
CO2.
MTT assay
GBC cells were added into 96-well plates at a
density of 2×103 cells/well and cultured overnight at
37°C and 5% CO2. Subsequently, different concentrations
of PD (0, 5, 10, 15, 20 and 25 µmol/l) were added to each well, and
the cells were cultured for 24, 48 or 72 h, separately. MTT (5
mg/ml) solution was added to the wells (10 µl/well) and incubated
at 37°C for 4 h. The culture medium was then replaced with DMSO
(100 µl/well) to dissolve the purple formazan and a microplate
reader (BioTek Instruments, Inc.) was used to measure the
absorbance at 490 nm.
Colony forming assay
NOZ and GBC-SD cells were collected and counted
manually. A total of 600 cells/well were added into 6-well plates
(Corning Inc.). Subsequently, PD at different concentrations (0, 5,
10 and 15 µmol/l) was used to treat the cells. The cells were
treated for ~14 days. After treatment, the cells were fixed with 4%
paraformaldehyde for 15 min and stained with 0.1% crystal violet
(Sigma-Aldrich; Merck KGaA) for 15 min at room temperature. All
colonies with >50 cells were recorded manually with a
fluorescence microscope (magnification ×40; Leica Microsystems
GmbH).
Cell apoptosis assay
NOZ and GBC-SD cells were cultured with PD at
various concentrations (0, 5, 10 and 15 µmol/l) for 48 h at 37°C
and 5% CO2. After culturing, the cells were collected
and washed with PBS. Next, the cells were diluted to the
appropriate density (106 cells/ml) using a Annexin V
binding buffer (BD Biosciences). The cell suspension (200 µl) was
gently mixed with Annexin V-FITC (5 µl) (BD Biosciences) and PI (5
µl) (BD Biosciences) and incubated for 15 min in the dark at room
temperature, these were a part of the kit mentioned earlier and
were used according to the manufacturer's protocol. Subsequently,
300 µl of the binding buffer was added. Flow cytometry using a BD
FACSCanto II (BD Biosciences) was used to analyze the sample within
1 h and BD FACSDiva Software v6.1.3 (BD Biosciences) was used to
analyze the results.
Hoechst 33342 staining
NOZ and GBC-SD cells were added into 12-well plates
and incubated overnight at 37°C and 5% CO2.
Subsequently, PD at 0, 5, 10 and 15 µmol/l was added to the wells,
and the plates were incubated for 48 h at 37°C and 5%
CO2. After treatment, the cells were stained with
Hoechst 33342 for 30 min in the dark at 37°C and then washed with
PBS. The cells were observed using a fluorescence microscope
(magnification, ×200; Leica Microsystems GmbH).
Mito-Tracker green staining
NOZ and GBC-SD cells were treated with different
concentrations (0, 5, 10 and 15 µmol/l) of PD for 48 h at 37°C and
5% CO2. Subsequently, the cells were stained with
Mito-Tracker green (Beyotime Institute of Biotechnology) at 37°C
for 30 min in the dark. The cells were observed using a
fluorescence microscope (magnification ×100; Leica Microsystems
GmbH).
Cell migration and invasion assay
Transwell plates with 24 wells (Corning Inc.) were
used to perform cell migration and invasion assays. The upper
chambers with or without Matrigel® (1 mg/ml) were dried
at 37°C for 30 min. NOZ and GBC-SD cells treated with PD (0, 5, 10
and 15 µmol/l) for 48 h were collected and diluted in serum-free
DMEM at a density of 2×105 cells/ml. Subsequently, 100
µl of the cell suspension was added to the upper chambers.
Simultaneously, 500 µl of DMEM supplemented with 10% FBS was added
to the lower chambers. After 24 h, the cells on top of the membrane
were removed using a cotton swab. The cells on the lower membrane
were fixed with 4% paraformaldehyde for 15 min at room temperature,
stained with crystal violet for 15 min at room temperature, and
observed using a phase-contrast microscope (magnification, ×100;
Olympus Corporation). Five fields of vision were randomly selected
per well.
Cell cycle analysis
For cell cycle analysis, the Cell Cycle and
Apoptosis Analysis kit was used (Beyotime Institute of
Biotechnology). NOZ and GBC-SD cells were seeded into 6-well plates
and incubated overnight at 37°C and 5% CO2. The
supernatant was replaced with different concentrations (0, 5, 10
and 15 µmol/l) of PD. After 48 h of incubation at 37°C and 5%
CO2, the cell layer was digested using trypsin, washed
with cold PBS and fixed with 70% ethanol at 4°C for at least 2 h
before storing at 4°C overnight. Subsequently, the cells were
washed with cold PBS and resuspended in staining buffer. Next, 0.1
mg/ml RNase A and 0.1 mg/ml PI were added to the cell suspension.
The staining buffer, RNase A and PI were part of the Cell Cycle and
Apoptosis Analysis kit (Beyotime Institute of Biotechnology) and
were used according to the manufacturer's instructions. The cells
were analyzed using flow cytometry after incubation at 37°C in the
dark for 30 min. BD FACSDiva Software v6.1.3 (BD Biosciences) was
used to analyze the distribution of the cell cycle phases.
Western blot analysis
NOZ and GBC-SD cells treated with PD (0, 5, 10 and
15 µmol/l) for 48 h were collected and washed with PBS. Total
protein was extracted from the cells using RIPA buffer (Beyotime
Institute of Biotechnology). After sonication (15–25 KHz, 25 sec on
ice), the mixture was centrifuged at 14,000 × g for 20 min at 4°C.
Protein concentration was quantified using a bicinchoninic acid
assay kit (Beyotime Institute of Biotechnology). Subsequently, the
proteins (30 µg/lane) were separated via 10% SDS-PAGE and
transferred to PVDF membranes (EMD Millipore). After blocking with
5% skimmed milk for 1 h at room temperature, the membranes were
incubated with the corresponding aforementioned primary antibodies
at 4°C overnight. The membranes were washed three times with TBS
supplemented with 0.1% Tween-20 (TBST) and incubated with
horseradish peroxidase-conjugated goat anti-rabbit secondary
antibodies (Cell Signaling Technology, Inc.) for 1 h at room
temperature. Subsequently, the membranes were washed again with
TBST three times and Millipore Western Blot Chemiluminescence HRP
Substrate ECL Luminescent solution was added (EMD Millipore). The
membranes were observed using the Gel Doc 2000 (Bio-Rad
Laboratories, Inc.) and ImageJ software version 1.48 (National
Institutes of Health) was used to quantify the densitometric values
of the detected bands.
Statistical analysis
Each experiment was performed three times. The data
was expressed as the mean ± SD. Shapiro-Wilk normality tests were
used to assess normality of quantitative variable distributions. If
the variables followed a normal distribution, one-way ANOVAs
followed by Dunnett's multiple comparisons test was performed using
GraphPad Prism version 7.0 (GraphPad Software, Inc.) to analyze the
results. Alternatively, Kruskal-Wallis test followed by Dunn's
multiple comparisons test was used in cases where the samples were
not normally distributed. P<0.05 was considered to indicate a
statistically significant difference.
Results
PD inhibits proliferation and colony
formation in GBC cells
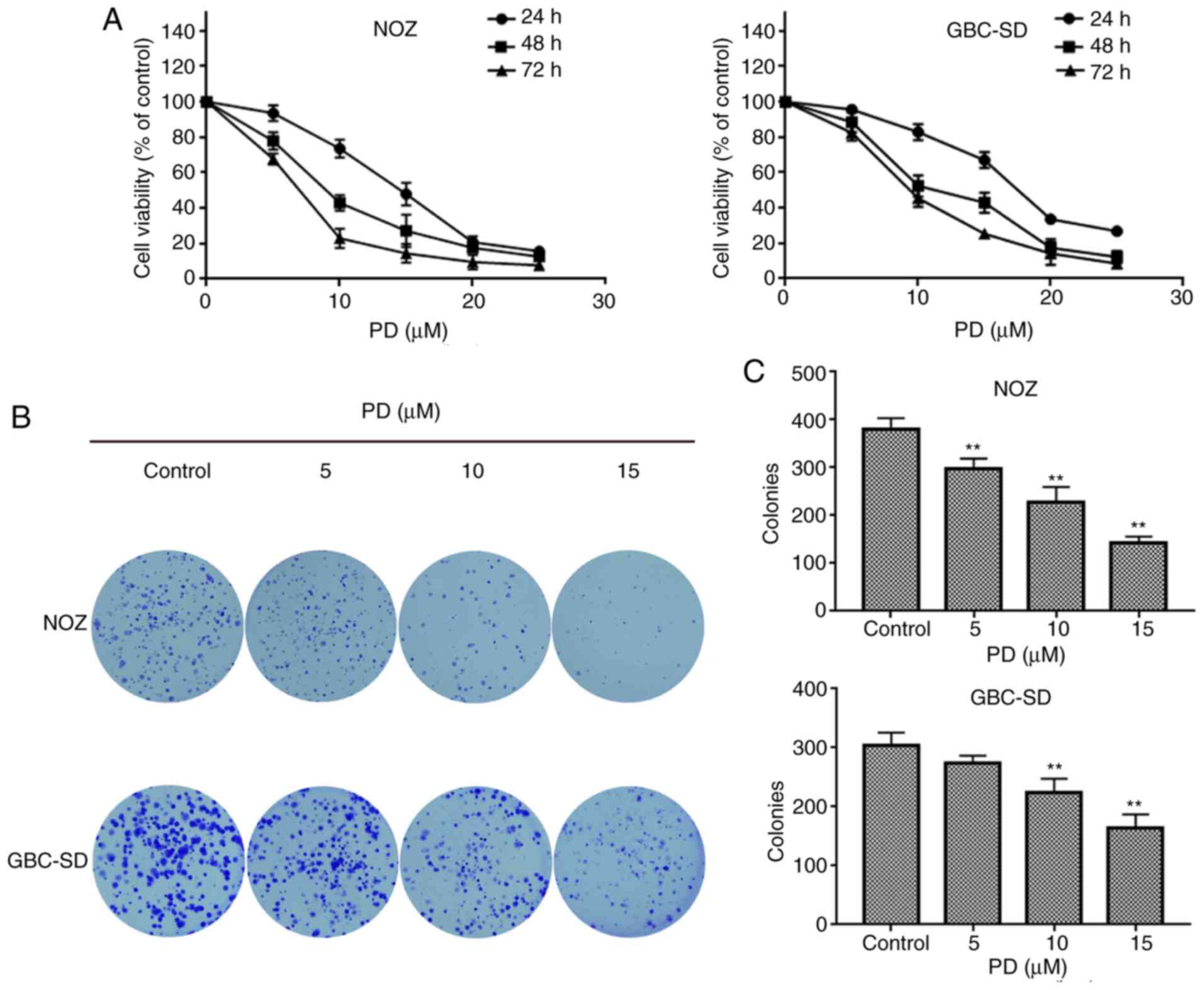
The MTT assay was used to detect the proliferative
ability of PD-treated GBC cells. The results indicated that the
proliferation of GBC cells decreased with an increased
concentration of PD in a time-dependent manner. In NOZ and GBC-SD
cells, the IC50 of PD was ~10 µmol/l at 48 h (Fig. 2A). PD had a stronger inhibitory
effect on NOZ and GBC-SD cells than on SGC-996 cells (Fig. S1); therefore, NOZ and GBC-SD cells
were selected for subsequent experiments. A colony forming assay
was performed to further investigate the colony forming ability of
PD-treated GBC cells. The results indicated that PD had a negative
effect on the ability of GBC cells to form colonies (Fig. 2B). Additionally, the PD-treated
groups developed fewer clones than those in the control group
(Fig. 2C). When treated with 5
µmol/l of PD, a significant decrease in colonies was observed in
NOZ cells, while a significant decrease in colonies was observed in
GBC-SD cells when the cells were treated with 10 µmol/l of PD
(Fig. 2C).
PD induces apoptosis, chromatin
condensation and mitochondrial dysfunction in GBC cells
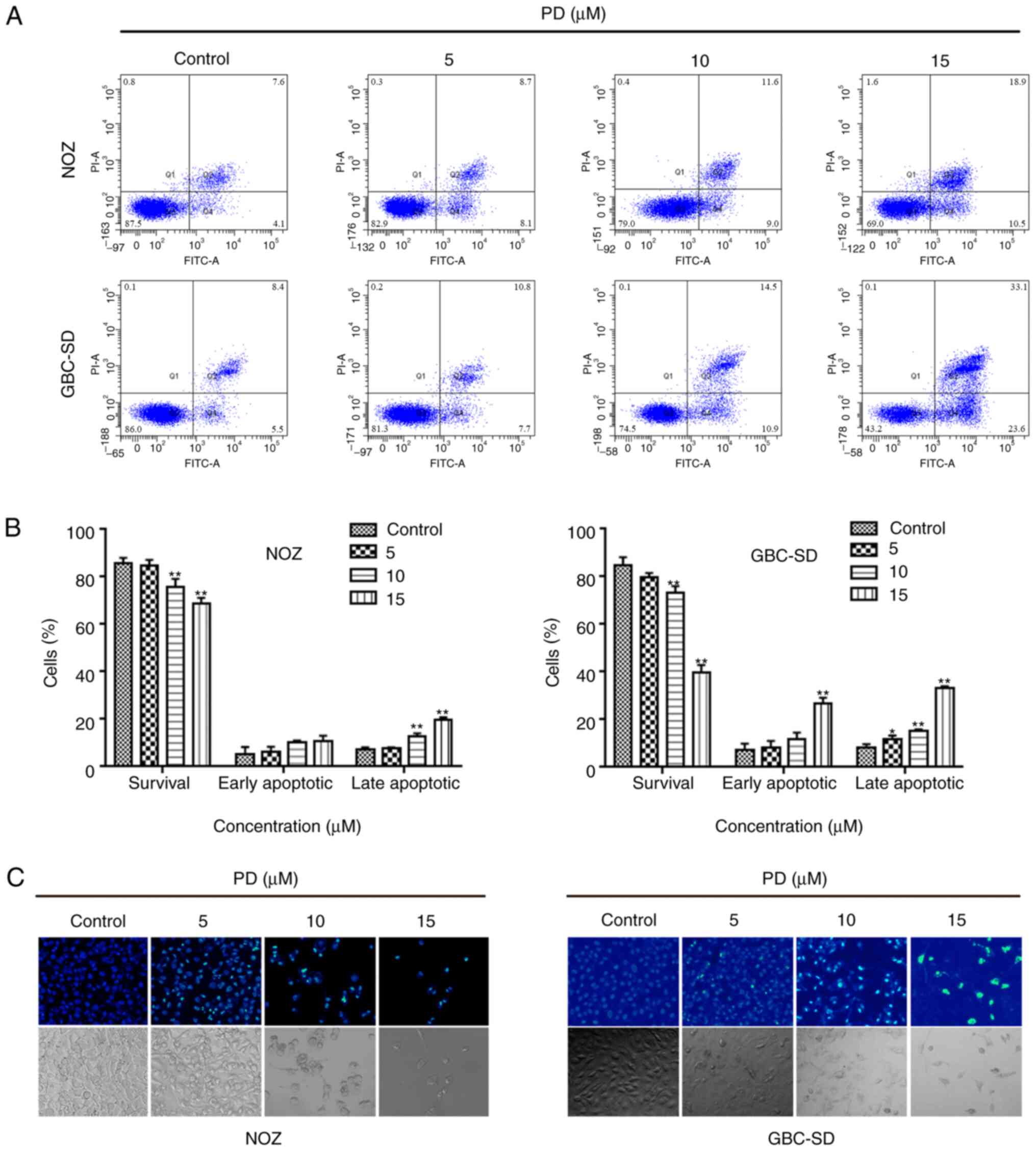
To assess the potential mechanisms of action behind
PD-mediated growth inhibition, PD-treated GBC cells were analyzed
using flow cytometry (Fig. 3A). As
the drug concentration increased, the proportion of apoptotic cells
also increased, while the percentage of surviving cells was reduced
(Fig. 3B). Subsequently, the
morphology of the nuclei was analyzed using Hoechst 33342 staining.
In the control group, the morphology of the cells was normal and
the chromatin was uniformly distributed, while the chromatin in the
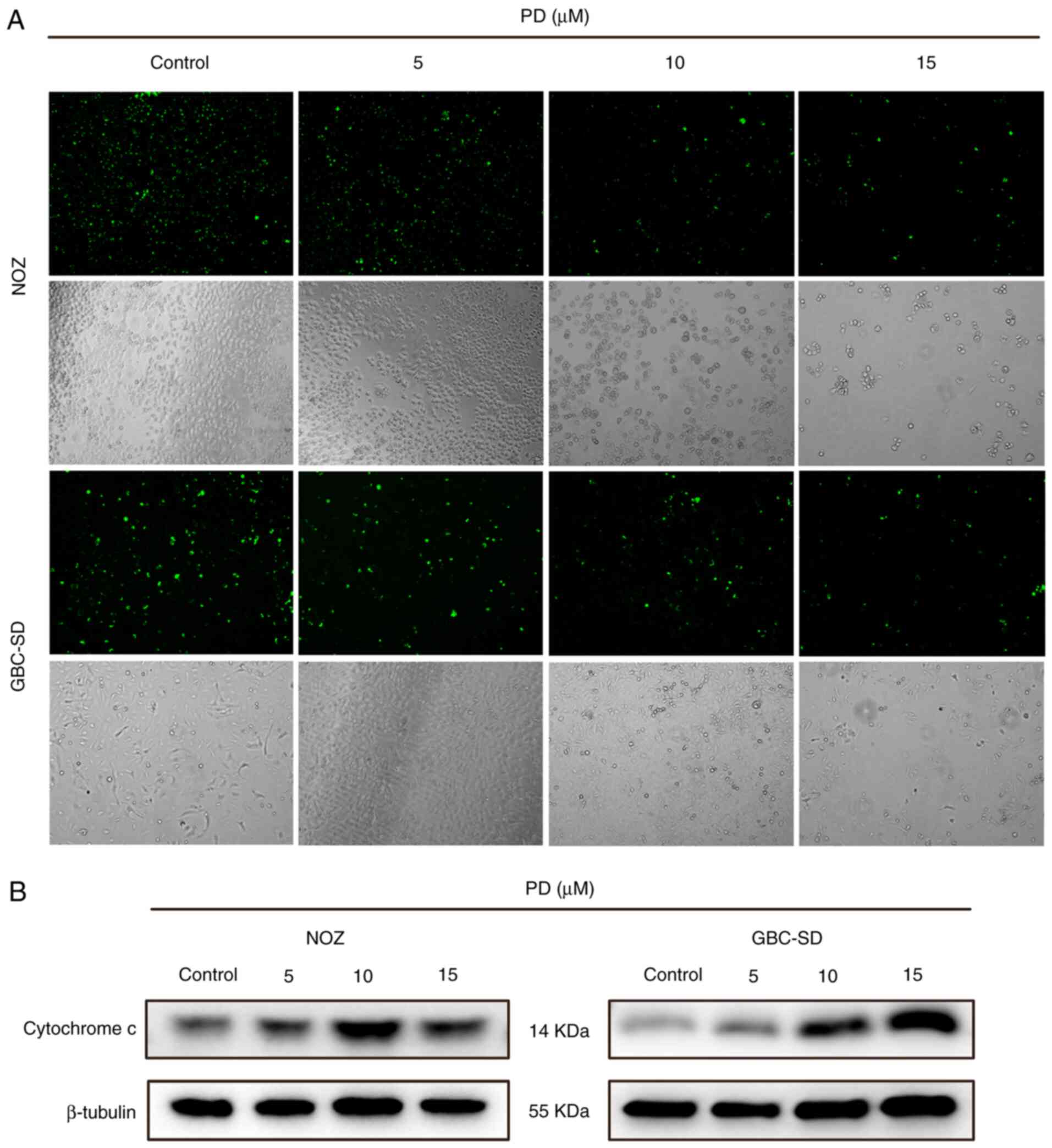
PD-treated group were markedly aggregated and broken (Fig. 3C). Subsequently, Mito-Tracker green
was used to stain the mitochondria. After treatment, bright green
fluorescence was observed in the control group, while the
fluorescence intensity of the PD-treated group was visibly reduced
(Fig. 4A). Additionally, cytochrome
c expression was higher in the PD-treated groups than that
in the control group (Fig. 4B).
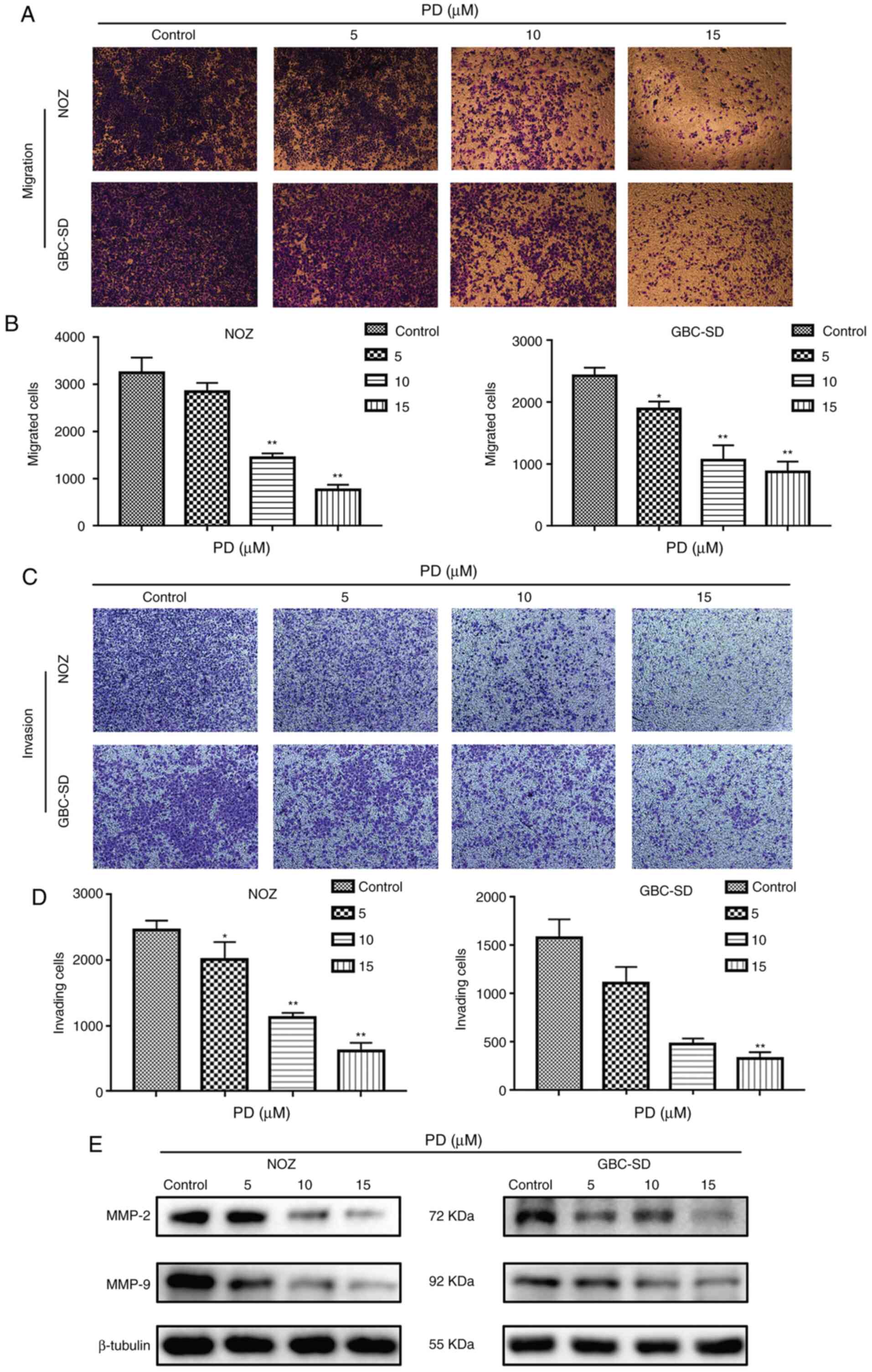
PD inhibits the migration and invasion
of GBC cells
To explore the influence of PD on the migratory and
invasive abilities of GBC cells, cell migration and invasion assays
were performed. Compared with the control group, PD treatment (10
and 15 µmol/l) markedly suppressed the migration numbers of NOZ and
GBC-SD cells (Fig. 5A) and the
difference was statistically significant (P<0.01; Fig. 5B). In addition, in the cell invasion
assay, PD treatment led to a significant anti-invasion effect in
GBC cells and this effect was dose-dependent (Fig. 5C and D). Additionally, western
blotting analysis confirmed that MMP-2 and MMP-9 had low expression
in PD-treated GBC cells compared with the control group (Fig. 5E). These data indicated that PD
inhibited the migration and invasion of GBC cells.
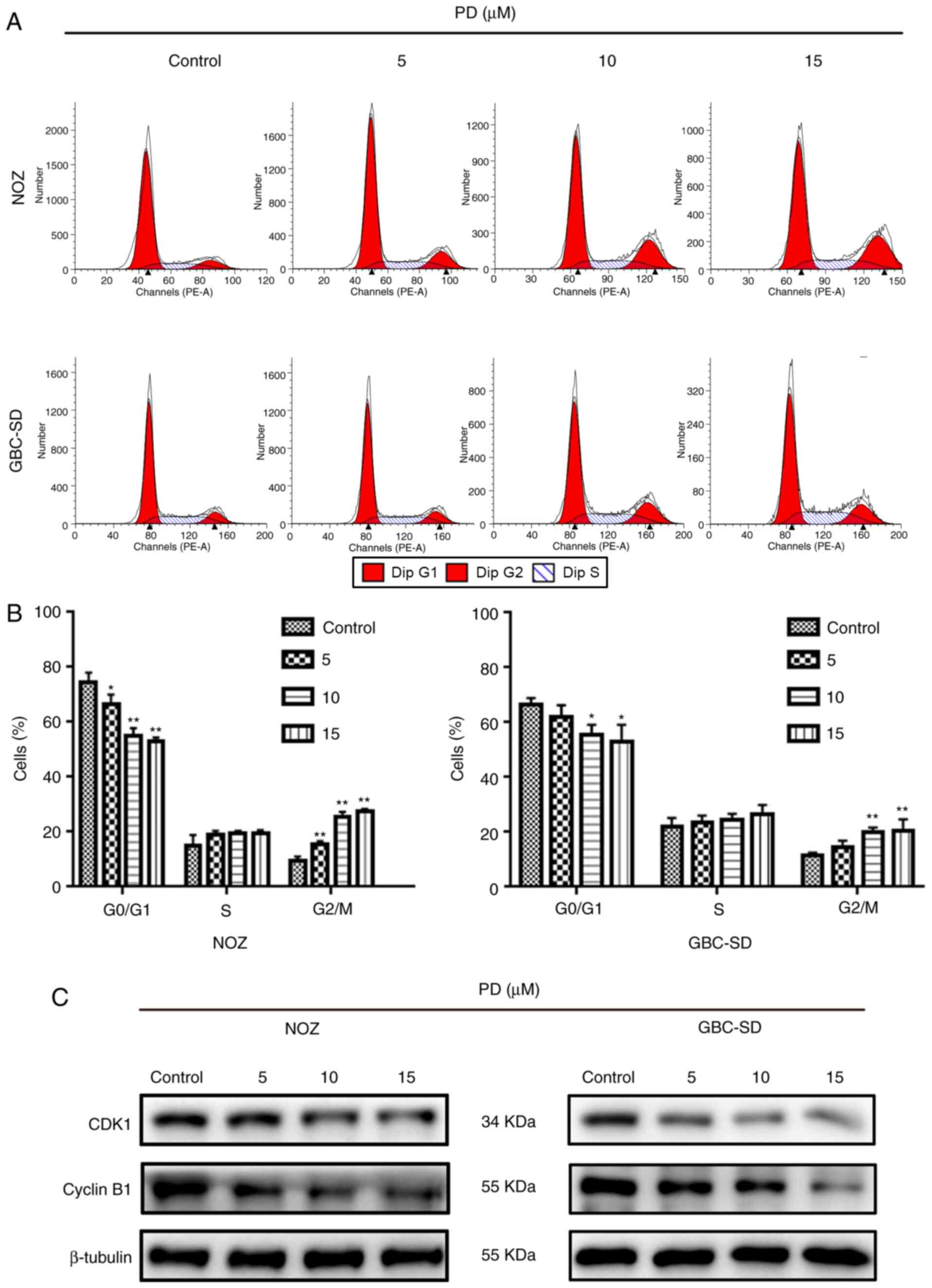
PD induces G2/M phase
arrest via regulation of the expression levels of proteins
associated with the cell cycle in GBC cells
To further understand how PD affected the
proliferation of GBC cells, flow cytometry was performed to measure
the proportion of PD-treated GBC cells at different cell cycle
stages (Fig. 6A). When cells were
treated with 10 µmol/l PD, the proportion of cells in
G2/M phase increased significantly, while the proportion
of cells in G0/G1 phase decreased in both
cell lines (Fig. 6B). This suggested
that PD inhibited the cell cycle progression of GBC cells. Next,
the levels of proteins that are closely associated with the cell
cycle, such as cyclin B1 and CDK1, were examined. The expression
levels of these proteins in GBC cells were markedly decreased
compared with the control group (Fig.
6C). Therefore, PD may induce G2/M phase arrest in
GBC cells to inhibit cell proliferation.
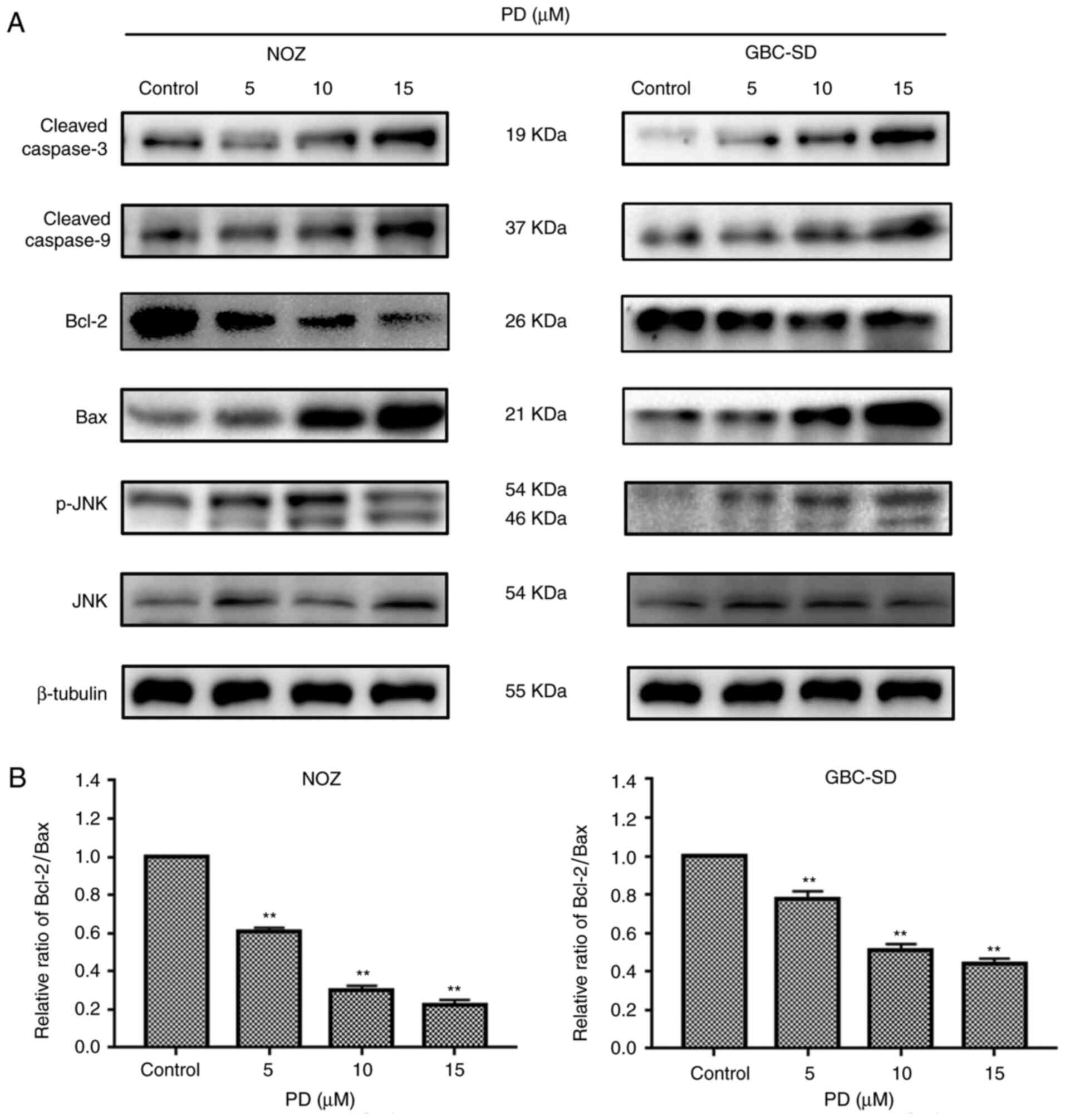
PD may induce apoptosis through the
JNK signaling pathway in NOZ and GBC-SD cells
Caspase family and Bcl-2 family proteins are both
important in the regulation of apoptosis (19). To further elucidate how PD induced
apoptosis in GBC cells, the expression levels of
apoptosis-associated proteins were investigated. It was observed
that the expression levels of Bax, cytochrome c, cleaved
caspase-9 and cleaved caspase-3 were increased with an increased
concentration of PD, while Bcl-2 expression was decreased with an
increased concentration of PD compared with the control group
(Figs. 4B and 7A). According to a previous study, the
Bax/Bcl-2 ratio is a key factor in the regulation of the apoptotic
process (20). In the present study,
PD decreased the expression levels of the anti-apoptotic protein
Bcl-2 and increased those of the pro-apoptotic protein Bax, thereby
decreasing the ratio of Bcl-2/Bax (Fig.
7B). Additionally, it was observed that p-JNK expression was
upregulated, while JNK expression was not markedly changed. In
summary, the present results suggested that PD may induce apoptosis
by initiating mitochondrial destruction through the JNK signaling
pathway in GBC cells.
Discussion
Previous studies have suggested that PD can suppress
the proliferation and development of tumor cells, including
prostate cancer (10), lung cancer
(21) and liver cancer (22) cells, through various mechanisms, such
as inhibiting cancer cell proliferation (16), inducing cell apoptosis (10), arresting cell cycle (12), inhibiting metastasis (17) and autophagy (18). However, to the best of our knowledge,
there is currently no research on the influence of PD on GBC cells.
The present study assessed the antitumor activity of PD using MTT
and clone forming assays, and revealed that PD significantly
inhibited the proliferation of GBC cells in a time- and
concentration-dependent manner. Subsequently, several experiments
were performed, including flow cytometry and nuclear staining with
Hoechst 33342, which revealed that the amount of PD-induced
apoptotic cells increased in a dose-dependent manner.
Invasiveness is an important feature of malignant
tumors. With the help of cell migration and invasion assays, the
present study demonstrated that the migratory and invasive
abilities of GBC cells were inhibited by PD. MMP-2 and MMP-9 are
closely associated with the migratory and invasive abilities of
tumor cells, mainly through the hydrolysis of basement membrane
components (23). Subsequently,
western blotting revealed that the expression levels of MMP-2 and
MMP-9 were decreased in PD-treated compared with the untreated GBC
cells.
Apoptosis is a complex pathophysiological process
involving several factors and proteins, such as the Bcl-2 (24) and caspase family (25). The imbalance between the
anti-apoptotic protein Bcl-2 and the pro-apoptotic protein Bax is
particularly important (26). A
previous study has demonstrated that Bcl-2 and Bax usually exist as
heterodimers and jointly regulate apoptosis (27). Changes in the Bcl-2/Bax ratio serve a
key role in mitochondrial-dependent apoptosis (28). When Bcl-2 is upregulated, it forms a
large number of heterodimers with Bax, inhibiting the activity of
Bax and slowing down the cell cycle (29,30).
With the upregulation of Bax, several Bax homodimers are formed,
which induce the release of cytochrome c into the cytoplasm;
activate caspase-9 and caspase-3; promote the cleavage of
caspase-3; and initiate the caspase cascade, resulting in apoptosis
(31,32). Therefore, the detection of cleaved
caspase-3 is considered a reliable marker of cell death and
apoptosis (33). In the present
study, it was demonstrated that elevated PD levels increased the
expression levels of Bax, cytochrome c, cleaved caspase-9
and cleaved caspase-3 in NOZ and GBC-SD cells. The mechanism of
action by which PD induces the apoptosis of GBC cells may be
associated with the regulation of the apoptosis signaling pathways
of Bax, Bcl-2 and caspase-3. The present study revealed that p-JNK
expression levels were increased in PD-treated GBC cells, while JNK
protein expression levels were unchanged, suggesting that
PD-treated GBC cells were activated by the JNK signaling pathway
and that this pathway may mediate mitochondrial apoptosis (34), further confirming the hypothesis of
the present study. In addition to the G1/S checkpoint,
the G2/M checkpoint is an important cell cycle
checkpoint (35). The CDK1-cyclin B1
complex is a protein kinase complex necessary for G2/M
phase transition (36). With the
help of cell cycle analysis, the present study revealed that PD
arrested GBC cells in the G2/M phase. Western blot
analysis indicated that the expression levels of CDK1 and cyclin B1
protein were decreased in PD-treated compared with untreated
cells.
There are several limitations in the present study.
Firstly, although the antitumor effects of PD on GBC cells were
investigated in vitro, whether PD may serve the same role
in vivo remains unknown. Secondly, the present study
hypothesized that the JNK signaling pathway may be involved in the
regulation of PD on GBC cell apoptosis, but how PD may activate the
JNK signaling pathway and its upstream genes remains unclear.
In summary, the present results demonstrated the
ability of PD to suppress the invasion of GBC cells and to induce
apoptosis through the JNK signaling pathway and G2/M
phase arrest, resulting in marked antitumor effects. Therefore, PD
represents a novel chemotherapy drug that may potentially be used
for the treatment of GBC.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was funded by the National Natural
Science Foundation of China (grant no. 81372642).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JW and TZ designed the study. XZ performed the
literature search. XZ and TZ performed the experiments. ZH, DZ, LJ
and CH performed the data analysis and revised the manuscript. XZ
was a major contributor in writing the manuscript. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kanthan R, Senger JL, Ahmed S and Kanthan
SC: Gallbladder cancer in the 21st century. J Oncol.
2015:9674722015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Are C, Ahmad H, Ravipati A, Croo D, Clarey
D, Smith L, Price RR, Butte JM, Gupta S, Chaturvedi A and Chowdhury
S: Global epidemiological trends and variations in the burden of
gallbladder cancer. J Surg Oncol. 115:580–590. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ma MZ, Li CX, Zhang Y, Weng MZ, Zhang MD,
Qin YY, Gong W and Quan ZW: Long non-coding RNA HOTAIR, a c-myc
activated driver of malignancy, negatively regulates miRNA-130a in
gallbladder cancer. Mol Cancer. 13:1562014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Weng M, Gong W, Ma M, Chu B, Qin Y, Zhang
M, Lun X, McFadden G, Forsyth P, Yang Y and Quan Z: Targeting
gallbladder cancer: Oncolytic virotherapy with myxoma virus is
enhanced by rapamycin in vitro and further improved by hyaluronan
in vivo. Mol Cancer. 13:822014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Millimouno FM, Dong J, Yang L, Li J and Li
X: Targeting apoptosis pathways in cancer and perspectives with
natural compounds from mother nature. Cancer Prev Res (Phila).
7:1081–1107. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cragg GM and Newman DJ: Plants as a source
of anti-cancer agents. J Ethnopharmacol. 100:72–79. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Balunas MJ and Kinghorn AD: Drug discovery
from medicinal plants. Life Sci. 78:431–441. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chin YW, Balunas MJ, Chai HB and Kinghorn
AD: Drug discovery from natural sources. AAPS J. 8:E239–E253. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tada A, Kaneiwa Y, Shoji J and Shibata S:
Studies on the saponins of the root of platycodon grandiflorum A.
De candolle. I. Isolation and the structure of platycodin-D. Chem
Pharm Bulletin (Tokyo). 23:2965–2972. 1975. View Article : Google Scholar
|
|
10
|
Zhou R, Lu Z, Liu K, Guo J, Liu J, Zhou Y,
Yang J, Mi M and Xu H: Platycodin D induces tumor growth arrest by
activating FOXO3a expression in prostate cancer in vitro and in
vivo. Curr Cancer Drug Targets. 14:860–871. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xu C, Sun G, Yuan G, Wang R and Sun X:
Effects of platycodin D on proliferation, apoptosis and PI3K/akt
signal pathway of human glioma U251 cells. Molecules.
19:21411–21423. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kim MO, Moon DO, Choi YH, Lee JD, Kim ND
and Kim GY: Platycodin D induces mitotic arrest in vitro, leading
to endoreduplication, inhibition of proliferation and apoptosis in
leukemia cells. Int J Cancer. 122:2674–2681. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li T, Xu WS, Wu GS, Chen XP, Wang YT and
Lu JJ: Platycodin D induces apoptosis, and inhibits adhesion,
migration and invasion in hepG2 hepatocellular carcinoma cells.
Asian Pac J Cancer Prev. 15:1745–1749. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yu JS and Kim AK: Platycodin D induces
apoptosis in MCF-7 human breast cancer cells. J Med Food.
13:298–305. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chun J, Joo EJ, Kang M and Kim YS:
Platycodin D induces anoikis and caspase-mediated apoptosis via p38
MAPK in AGS human gastric cancer cells. J Cell Biochem.
114:456–470. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ahn KS, Noh EJ, Zhao HL, Jung SH, Kang SS
and Kim YS: Inhibition of inducible nitric oxide synthase and
cyclooxygenase II by platycodon grandiflorum saponins via
suppression of nuclear factor-kappaB activation in RAW 264.7 cells.
Life Sci. 76:2315–2328. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chun J and Kim YS: Platycodin D inhibits
migration, invasion, and growth of MDA-MB-231 human breast cancer
cells via suppression of EGFR-mediated akt and MAPK pathways. Chem
Biol Interact. 205:212–221. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li T, Tang ZH, Xu WS, Wu GS, Wang YF,
Chang LL, Zhu H, Chen XP, Wang YT, Chen Y and Lu JJ: Platycodin D
triggers autophagy through activation of extracellular
signal-regulated kinase in hepatocellular carcinoma HepG2 cells.
Eur J Pharmacol. 749:81–88. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cosulich SC, Savory PJ and Clarke PR:
Bcl-2 regulates amplification of caspase activation by cytochrome
c. Curr Biol. 9:147–150. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nuñez G and Clarke MF: The Bcl-2 family of
proteins: Regulators of cell death and survival. Trends Cell Biol.
4:399–403. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li T, Chen X, Chen X, Ma DL, Leung CH and
Lu JJ: Platycodin D potentiates proliferation inhibition and
apoptosis induction upon AKT inhibition via feedback blockade in
non-small cell lung cancer cells. Sci Rep. 6:379972016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li T, Xu XH, Tang ZH, Wang YF, Leung CH,
Ma DL, Chen XP, Wang YT, Chen Y and Lu JJ: Platycodin D induces
apoptosis and triggers ERK- and JNK-mediated autophagy in human
hepatocellular carcinoma BEL-7402 cells. Acta Pharmacol Sin.
36:1503–1513. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bauvois B: New facets of matrix
metalloproteinases MMP-2 and MMP-9 as cell surface transducers:
Outside-in signaling and relationship to tumor progression. Biochim
Biophys Acta. 1825:29–36. 2012.PubMed/NCBI
|
|
24
|
Cory S and Adams JM: The Bcl2 family:
Regulators of the cellular life-or-death switch. Nat Rev Cancer.
2:647–656. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Thornberry NA and Lazebnik Y: Caspases:
Enemies within. Science. 281:1312–1316. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Campbell KJ and Tait SW: Targeting BCL-2
regulated apoptosis in cancer. Open Biol. 8:1800022018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang W, Guo Q, You Q, Zhang K, Yang Y, Yu
J, Liu W, Zhao L, Gu H, Hu Y, et al: Involvement of bax/bcl-2 in
wogonin-induced apoptosis of human hepatoma cell line SMMC-7721.
Anticancer Drugs. 17:797–805. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jin CY, Moon DO, Choi YH, Lee JD and Kim
GY: Bcl-2 and caspase-3 are major regulators in agaricus
blazei-induced human leukemic U937 cell apoptosis through
dephoshorylation of akt. Biol Pharm Bull. 30:1432–1437. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Vaux DL, Cory S and Adams JM: Bcl-2 gene
promotes haemopoietic cell survival and cooperates with c-myc to
immortalize pre-B cells. Nature. 335:440–442. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cheng N, Janumyan YM, Didion L, Van
Hofwegen C, Yang E and Knudson CM: Bcl-2 inhibition of T-cell
proliferation is related to prolonged T-cell survival. Oncogene.
23:3770–3780. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Pistritto G, Trisciuoglio D, Ceci C,
Garufi A and D'Orazi G: Apoptosis as anticancer mechanism: Function
and dysfunction of its modulators and targeted therapeutic
strategies. Aging (Albany NY). 8:603–619. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Elmore S: Apoptosis: A review of
programmed cell death. Toxicol Pathol. 35:495–516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zaman S, Wang R and Gandhi V: Targeting
the apoptosis pathway in hematologic malignancies. Leuk Lymphoma.
55:1980–1992. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xia Z, Dickens M, Raingeaud J, Davis RJ
and Greenberg ME: Opposing effects of ERK and JNK-p38 MAP kinases
on apoptosis. Science. 270:1326–1331. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Stark GR and Taylor WR: Control of the
G2/M transition. Mol Biotechnol. 32:227–248. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Malumbres M: Cyclin-dependent kinases.
Genome Biol. 15:1222014. View
Article : Google Scholar : PubMed/NCBI
|