Introduction
Gastric cancer (GC) is a common cancer worldwide and
ranks third among the leading causes of cancer-associated
mortalities. Incidence rates are markedly elevated in Eastern Asia
(incidence rates are 24.7 per 100,000), when compared with Northern
America (incidence rates are 8.4 per 100,000) and Northern Europe
(incidence rates are 9.3 per 100,000) (1). Targeted therapy with biomarkers for
advanced GC has developed rapidly in recent years (2). Due to late diagnosis, patients with
extensive invasion and metastasis have poor prognoses (3). Even after a complete resection,
recurrence occurs in ~50% of patients (4). As the molecular mechanisms underlying
the metastasis and recurrence of GC have not been fully clarified,
identifying key GC-promoting molecules may contribute to the
understanding of GC pathogenesis and identification of new
therapeutic targets.
Long non-coding RNAs (lncRNAs) are a type of RNA
transcript that have >200 nucleotides and are not translated
into proteins (5). The myocardial
infarction associated transcript (MIAT) is first identified to play
a role in the pathogenesis of myocardial infarction (6). Recent studies have reported that MIAT
is upregulated in several types of cancers including papillary
thyroid (7), lung (8) and colorectal cancer (9). However, the underlying molecular
mechanism of MIAT in GC remains largely unknown.
MicroRNAs (miRNAs) are 20–23 nucleotides in length
and serve a negative regulatory role by binding to the
3′untranslated region (UTR) of target mRNAs, which results in
inhibition of mRNA translation or promotion of mRNA degradation
(10). lncRNAs can serve as sponges
of miRNAs and reduce their regulatory effects on target mRNAs
(11). miR-331-3p serves as a
potential tumour suppressor in multiple types of human cancers,
including pancreatic (12), ovarian
(13) and colorectal cancer
(14). miR-331-3p has also been
demonstrated to inhibit GC cell growth (15). Here, it was speculated that MIAT
functions via targeting miR-331-3p.
Ras-related protein Rab-5B (RAB5B), an isoform of
RAB5 (16). High expression of RAB5B
is associated with cancer progression and poor prognoses in
numerous cancer types, including pancreatic (17), breast (18) and ovarian cancer (19). However, the function of RAB5B in GC
is yet to be elucidated.
The present study investigated the effects and
mechanisms of the lncRNA MIAT on the proliferation, apoptosis and
metastasis of GC cells.
Materials and methods
The Cancer Genome Atlas (TCGA)
database
Tissues samples in TCGA database (https://cancergenome.nih.gov/) were divided into two
groups (MIAT high expression group: n=25; MIAT low expression
group: n=194; cut-off=9.96). A survival curve was generated to
analyze the association between MIAT expression and the overall
survival of patients with GC patients. A log-rank test was used to
compare survival times between two groups. The result was
considered statistically significant if P<0.05.
Patient samples and cell lines
GC tissue samples and paired normal tissue samples
were obtained from 47 patients who underwent surgery between March
2017 and March 2018 at Chengyang People's Hospital (Qingdao,
Shandong, China) with written informed consent. All samples were
frozen in liquid nitrogen and stored at −80°C until use. The
present study was approved by the Ethics Committee of Chengyang
People's Hospital. GC tissues were fixed in 4% buffered
paraformaldehyde for 24 h at 4°C, embedded in paraffin and then
sectioned to 5 µm. These sections were stained with hematoxylin for
15 min and eosin for 5 min at 25°C. Samples were examined under a
light microscope at a magnification of ×400. The human gastric
epithelial mucosa cell line GES-1 and GC cell lines HGC-27, AGS,
MKN45 and NCI-N87 were purchased from Nanjing Keygen Biotech Co.,
Ltd. HGC-27, AGS, MKN45 and NCI-N87 cells were maintained in
RPMI-1640 with 10% foetal bovine serum (FBS), 100 U/ml penicillin
and 100 µg/ml streptomycin (HyClone; GE Healthcare Life Sciences).
GES-1 cells were cultured in Dulbecco's modified Eagle's medium
(DMEM) with FBS, 100 U/ml penicillin and 100 µg/ml streptomycin
(HyClone; GE Healthcare Life Sciences). All cells were cultured in
a humidified chamber with 5% CO2 at 37°C.
Transfection
pcDNA 3.1-MIAT, pcDNA 3.1-RAB5B, pcDNA 3.1-negative
control (pcDNA 3.1-NC), small interfering RNA (si)-MIAT-1, si-RAB5B
and negative control siRNA (si-NC) were generated by Shanghai Gene
Pharma Company. miR-331-3p mimics, inhibitors and their respective
negative controls were purchased from Guangzhou RiboBio Co., Ltd.
The siRNA, miR-331-3p mimics and inhibitor sequences are listed in
Table I. HGC-27, AGS and GES-1 cells
were transfected using Lipofectamine 2000 transfection reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) at 37°C, and the fresh
medium was changed after 6 h. The final concentration for
transfection was 50 nM. HGC-27, AGS and GES-1 cells were harvested
at 48 h after transfection.
 | Table I.siRNA sequences used in the present
study. |
Table I.
siRNA sequences used in the present
study.
| Name | Sequences |
|---|
| si-MIAT-1 |
5′-GGUGUUAAGACUUGGUUUCTT-3′ |
| si-MIAT-2 |
5′-ACUUCUUCGUAUGUUCGGCTT-3′ |
| si-NC |
5′-UUCUCCGAACGUGUCACGUTT-3′ |
| miR-331-3p
mimics |
5′-GCCCCUGGGCCUAUCCUAGAA-3′ |
| miR-331-3p
inhibitors |
5′-UUCUAGGAUAGGCCCCAGGGGC-3′ |
Reverse transcription-quantitative PCR
(RT-qPCR) analysis
Total RNA from all cultured cells lines and human
tissue was extracted using TRIzol according to the manufacturer's
instructions (Invitrogen; Thermo Fisher Scientific, Inc.). cDNA was
synthesized from 1 µg total RNA using the PrimeScript®
RT reagent kit (Takara Bio, Inc.) according to the manufacturer's
instructions. qPCR was performed in an ABI 7500 instrument (Applied
Biosystems; Thermo Fisher Scientific, Inc.) using SYBR-Green
Real-Time PCR Master Mix (Takara Bio, Inc.). Amplification
conditions were 94°C for 7 min, followed by 40 cycles of 95°C for
15 sec, and 60°C for 30 sec. Relative expression levels were
quantitated using the 2−ΔΔCq method (20) with β-actin and U6 as reference
control. Primer pairs are listed in Table II.
| Table II.Primers used for reverse
transcription-quantitative PCR. |
Table II.
Primers used for reverse
transcription-quantitative PCR.
| Primer names | Sequences |
|---|
| lncRNA-MIAT | F:
5′-GGACGTTCACAACCACACTG-3′ |
| lncRNA-MIAT | R:
5′-TCCCACTTTGGCATTCTAGG-3′ |
| miR-331-3p | F:
5′-GCGCCCCTGGGCCTATC-3 |
| miR-331-3p | R:
5′-CGATGACCTATGAATTGACA-3′ |
| RAB5B | F:
5′-TTCCTCACCCAGTCCGTTTG-3′ |
| RAB5B | R:
5′-GCCTGTCGCTGTAGTTCCTT-3′ |
| U6 | F:
5′-CTCGCTTCGGCAGCACA-3′ |
| U6 | R:
5′-AACGCTTCACGAATTTGCGT-3′ |
| β-actin | F:
5′-TGACGTGGACATCCGCAAAG-3′ |
| β-actin | R:
5′-CTGGAAGGTGGACAGCGAGG-3′ |
Cell Counting Kit (CCK)-8 assay
The AGS and HGC-27 cells without transfection
(5×103 cells/well) were cultivated in RPMI-1640 complete
medium with 10% FBS or serum-free medium at 37°C for 1, 2 and 3
days. Then, according to the manufacturer's instructions, CCK-8
solution (10 µl; Dojindo Molecular Technologies, Inc.) was
supplemented into each well for incubating another 2 h. A
microplate reader (BioTek Instruments, Inc.) was used for examining
the absorbance at 450 nm. The GES-1 cells (5×103
cells/well) were cultivated in DMEM medium with 10% FBS at 37°C for
1, 2 and 3 days. The next steps were similar to the AGS and HGC-27
cells.
Cell proliferation assay
The HGC-27 and AGS cells at 48 h post transfection
were seeded into 24-well plates (2×104) and cultured
with 5-ethynyl-2′-deoxyuridine (EdU; 50 µM; Guangzhou RiboBio Co.,
Ltd.) at 37°C for 2 h. The cells were fixed in 4% paraformaldehyde
at 25°C for 30 min and incubated with glycine at 37°C for 4 h.
After being submerged into the Apollo reaction cocktail (deionized
water 469 µl, Apollo reaction buffer 25 µl, Apollo catalytic
solution 5 µl, Apollo 567 fluorescent dye solution 1.5 µl and
Apollo buffer additive 5 mg) at 37°C for 30 min in the dark, the
cells were washed with 0.5% Triton X-100. Cell nuclei were stained
using Hoechst 33342 at 37°C for 30 min. Cells were then imaged
using a Nikon fluorescence microscope (Eclipse Ti2-U; Nikon
Corporation; magnification, ×400).
Cell apoptosis analysis
Apoptosis analysis was conducted by flow cytometry
using the Annexin V/propidium iodide kit (Nanjing KeyGen Biotech
Co., Ltd). The HGC-27, AGS and GES-1 cells at 48 h post
transfection were detached from the plate using EDTA-free trypsin,
gathered into a cell suspension with 400 µl Annexin binding buffer
at a concentration of 106 cells/ml. Then, 5 µl Annexin
V-FITC and 10 µl propidium iodide were added, mixed and incubated
for 15 min in the dark at room temperature. Cell apoptosis was
tested using flow cytometry (A60-Micro, Apogee, UK and BD Accuri C6
Plus; BD Biosciences). The FlowJo software (version 10.0.6; FlowJo,
LLC) was applied for data analysis.
Wound healing assay
The HGC-27, AGS and GES-1 cells at 48 h post
transfection were seeded in a 6-well plate (3×105).
Subsequently, monolayer cells were scratched with a 200 µl pipette
tip to generate an artificial wound. After the cells were cultured
at 37°C for 24 h in serum-free medium, the wound healing rate was
calculated. Cells were imaged under a light microscope at a
magnification of ×100. Scratch healing was observed at the same
location at the 0 and 24 h. HGC-27, AGS and GES-1 cells migration
was assessed by calculating scratch width using the formula: The
relative scratch width=(number of cells at T24-number of cells at
T0)/number of cells at T0 ×100%, where T0 was 0 h and T24 was 24
h.
Invasion assay
The HGC-27, AGS and GES-1 cells at 48 h post
transfection in serum-free medium (3×105) were added to
the upper Transwell chambers which were pre-coated with Matrigel at
37°C for 60 min (BD Biosciences). The bottom chamber contained
medium with 10% FBS. After incubation at 37°C for 24 h, the cells
were stained with 0.1% crystal violet at 37°C for 30 min. Cell
invasion was quantified by counting the number of cells in 5 random
fields under a light microscope (magnification, ×100).
Luciferase reporter assay
Bioinformatics databases Starbase v3.0 (http://starbase.sysu.edu.cn/index.php)
and TargetScanHuman Release 7.2 (http://www.targetscan.org/vert_72/) were used to
identify the potential target sequences of miR-331-3p. HGC-27 and
AGS cells were seeded into 24-well plates at a density of
6×104 cells per well, followed by cotransfection with
MIAT 3′UTR wild-type (wt) or MIAT 3′UTR mutant type (mut) [RAB5B
3′UTR (wt) or RAB5B 3′UTR (mut)] and the miR-331-3p mimics or
mimics NC using Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). The wt and mut containing the putative binding
site of miR-331-3p were cloned and established in the firefly
luciferase-expressing pMIRREPORT vector (Obio Technology;
http://www.obiosh.com/kyfw/xb/sygsmjc/676.html?1574909908).
Luciferase activity was detected 48 h after transfection using the
dual-luciferase reporter gene assay kit (Promega Corporation) and
Renilla luciferase activity was used as the
normalization.
Western blotting
The HGC-27, AGS and GES-1 cells at 48 h post
transfection were lysed using ice-cold radioimmunoprecipitation
assay (RIPA) lysis buffer (Beyotime Institute of Biotechnology).
Protein concentration was quantified using an Enhanced BCA Protein
Assay kit (Beyotime Institute of Biotechnology). The total protein
extract (40 µg/lane) was separated via 10% SDS-PAGE and transferred
to nitrocellulose membranes. The membranes were blocked with 5%
non-fat milk for 1 h at room temperature. After blocking, the
membranes were incubated with the relevant antibodies for overnight
at 4°C. The following antibodies were used in the present study:
Bax (cat. no. 2774; 1:1,000; Cell Signaling Technology), Bcl-2
(cat. no. 4223; 1:1,000, Cell Signaling Technology), cleaved
caspase-3 (cat. no. 9661; 1:1,000; Cell Signaling Technology),
RAB5B (cat. no. ab72907; 1:1,000; Abcam) and β-actin (cat. no.
4967; 1:2,000; Cell Signaling Technology). The following day, the
membranes were incubated with horseradish peroxidase-conjugated
secondary antibody (cat. no. 7074; 1:5,000; Cell Signaling
Technology) for 2 h at room temperature. Proteins were normalized
to β-actin levels. Finally, bands were detected by enhanced
chemiluminescence (General Electric Company, Inc.) and quantified
using ImageJ software (National Institutes of Health).
Statistical analysis
Data analysis was performed using GraphPad Prism 7
(GraphPad Software, Inc.). All results were reported as the mean ±
standard deviation (SD) of three independent experiments.
Differences were assessed using paired Student's t-test or one-way
ANOVA followed by Tukey's multiple comparisons test. Pearson's
correlation was used to calculate the correlation between
expression of MIAT and miR-331-3p (miR-331-3p and RAB5B; MIAT and
RAB5B). Survival analysis was performed using the R package
‘survival’ (version 3.6.1) (21,22) and
the Kaplan-Meier curve method. A log-rank test was used to compare
survival times between two groups. The result was considered
statistically significant if P<0.05.
Results
MIAT expression is increased in GC
tissues and cell lines
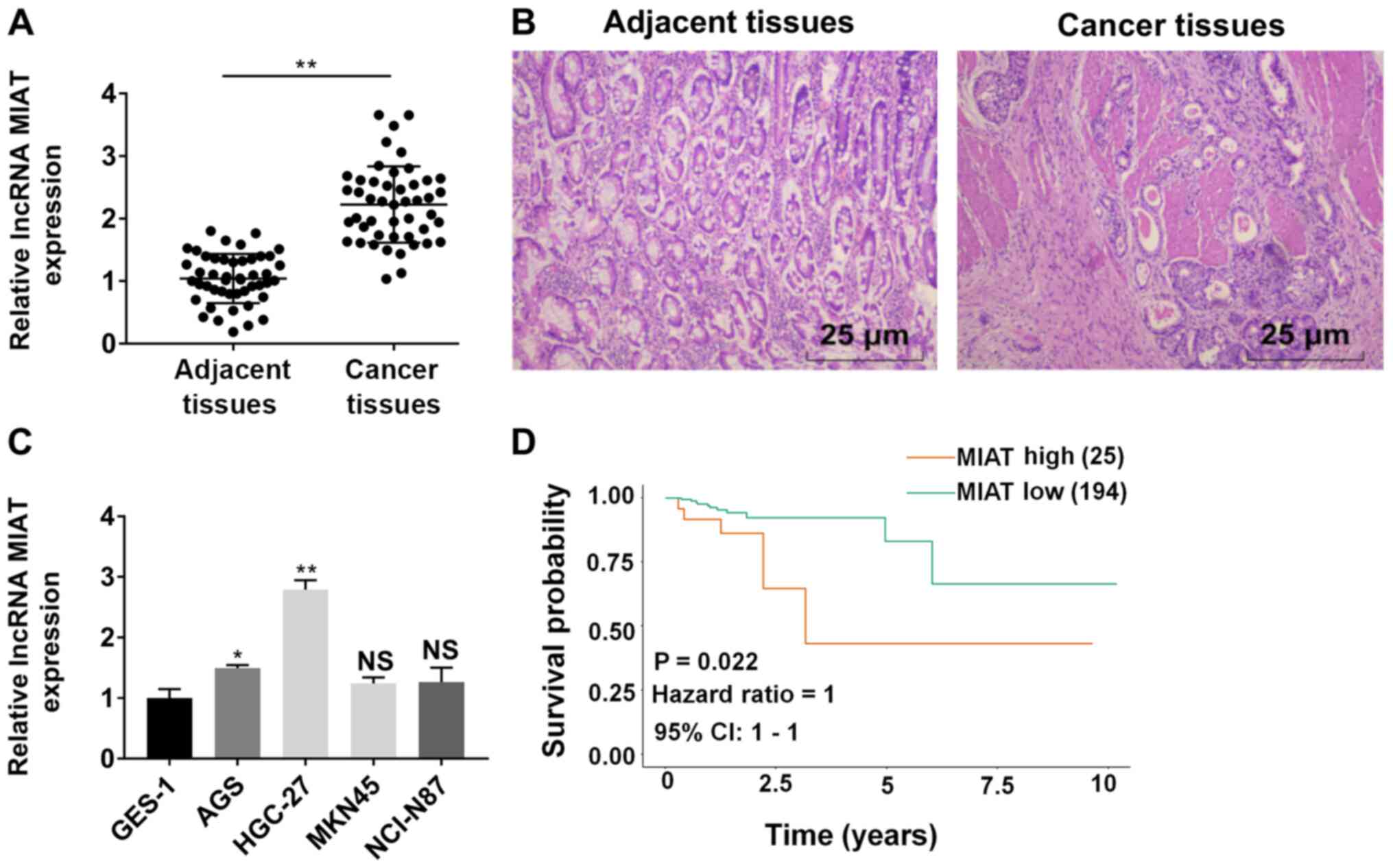
RT-qPCR results revealed that MIAT expression was
significantly increased in GC tissue samples (adenocarcinoma) as
compared with paratumor tissues (Fig.
1A). Representative staining images are displayed in Fig. 1B. The high expression of MIAT was
correlated with late TNM stage and lymph node involvement (Table III). In Fig. 1C, MIAT expression was detected in
GES-1 cells and four GC (AGS, HGC-27, MNK45 and NCI-N87) cell
lines. The expression of MIAT in AGS, HGC-27, MNK45 and NCI-N87
cells was higher compared with GES-1 cells. Among the four cell
lines, MIAT expression was significantly higher in AGS and HGC-27
cells, compared with GES-1 cells. Therefore, AGS and HGC-27 cells
were selected to perform subsequent experiments. The expression of
MIAT in HGC-27 cells was higher compared with in AGS cells
(Fig. S1A). As expected, the
proliferation, migration and invasion ability of HGC-27 cells was
higher compared with AGS cells (Fig.
S1B and C). Kaplan-Meier analysis revealed that high expression
of MIAT was associated with poor overall survival time based on
data from TCGA database (Fig.
1D).
| Table III.The association between lncRNA MIAT
expression and clinicopathological variables of patients with
gastric cancer. |
Table III.
The association between lncRNA MIAT
expression and clinicopathological variables of patients with
gastric cancer.
|
|
| lncRNA MIAT
expression |
|
|---|
|
|
|
|
|
|---|
| Variables | Value (n) | High (n=23) | Low (n=24) | P-value |
|---|
| Sex |
|
|
| 0.1351 |
|
Male | 30 | 18 | 12 |
|
|
Female | 17 | 5 | 12 |
|
| Age, years |
|
|
| 0.2124 |
|
≤60 | 15 | 5 | 10 |
|
|
>60 | 32 | 18 | 14 |
|
| Tumor size, cm |
|
|
| 0.3852 |
| ≤5 | 22 | 9 | 13 |
|
|
>5 | 25 | 14 | 11 |
|
| Depth of
invasion |
|
|
| 0.1468 |
|
T1+T2 | 22 | 8 | 14 |
|
|
T3+T4 | 25 | 15 | 10 |
|
|
Differentiation |
| Well or
moderate | 21 | 8 | 13 | 0.2443 |
| Poor or
other | 26 | 15 | 11 |
|
| Lymph node
involvement |
|
|
| 0.0355a |
|
Absence | 18 | 6 | 12 |
|
|
Presence | 29 | 17 | 12 |
|
| TNM stage |
|
|
| 0.0415a |
|
I–II | 26 | 9 | 17 |
|
|
III–IV | 21 | 14 | 7 |
|
MIAT promotes HGC-27 and AGS cell
proliferation and inhibits apoptosis
Overexpression of MIAT was induced in HGC-27 and AGS
cells following transfection with pcDNA3.1-MIAT (Fig. 2A). The results revealed that both
si-MIAT-1 and si-MIAT-2 downregulated MIAT expression levels
(Fig. 2B). Cell proliferation was
significantly increased in cells transfected with pcDNA3.1-MIAT
compared with cells transfected with pcDNA3.1-NC. Knockdown of MIAT
significantly suppressed cell proliferation in HGC-27 and AGS cells
(Fig. 2C). The apoptotic rate of
HGC-27 and AGS cells was decreased by overexpression of MIAT and
increased by knockdown of MIAT (Fig.
2D). The expression of Bax and cleaved caspase-3 were
significantly decreased following pcDNA3.1-MIAT transfection and
increased by transfection withsi-MIAT-1. By contrast, the
expression of Bcl-2 was increased by pcDNA3.1-MIAT transfection and
decreased by transfection with si-MIAT-1 (Fig. 2E).
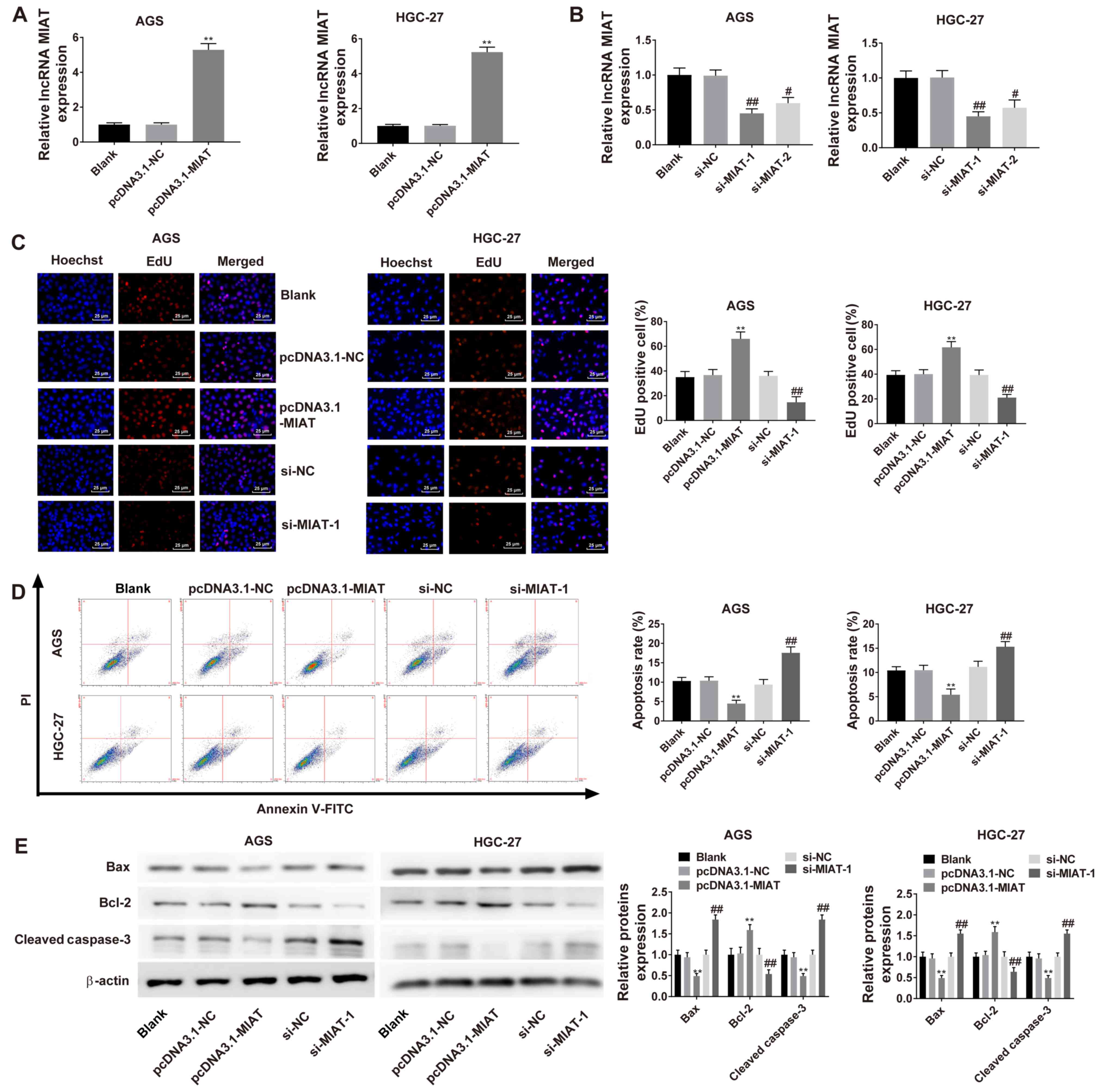 | Figure 2.MIAT promotes GC cell proliferation
by inhibiting apoptosis. (A) RT-qPCR analysis of MIAT expression in
HGC-27 and AGS cells transfected with pcDNA3.1-NC or pcDNA3.1-MIAT.
(B) RT-qPCR analysis of MIAT expression in HGC-27 and AGS cells
transfected with si-NC, si-MIAT-1 or si-MIAT-2. (C) EdU analysis of
cell proliferation in HGC-27 and AGS cells transfected with
pcDNA3.1-NC, pcDNA3.1-MIAT, si-NC or si-MIAT-1. (D) Flow cytometric
analyses of HGC-27 and AGS cells transfected with pcDNA3.1-NC,
pcDNA3.1-MIAT, si-NC or si-MIAT-1. (E) The protein levels of Bax,
Bcl-2 and cleaved caspase-3 in HGC-27 and AGS cells were measured
via western blotting. n=3. **P<0.01 vs. the blank or pcDNA3.1-NC
group; #P<0.05, ##P<0.01 vs. the blank
or si-NC group. GC, gastric cancer; MIAT, myocardial infarction
associated transcript; si, small interfering; NC, negative control;
RT-qPCR, reverse transcription-quantitative PCR; PI, propridium
iodide. |
MIAT promotes HGC-27 and AGS cell
migration and invasion
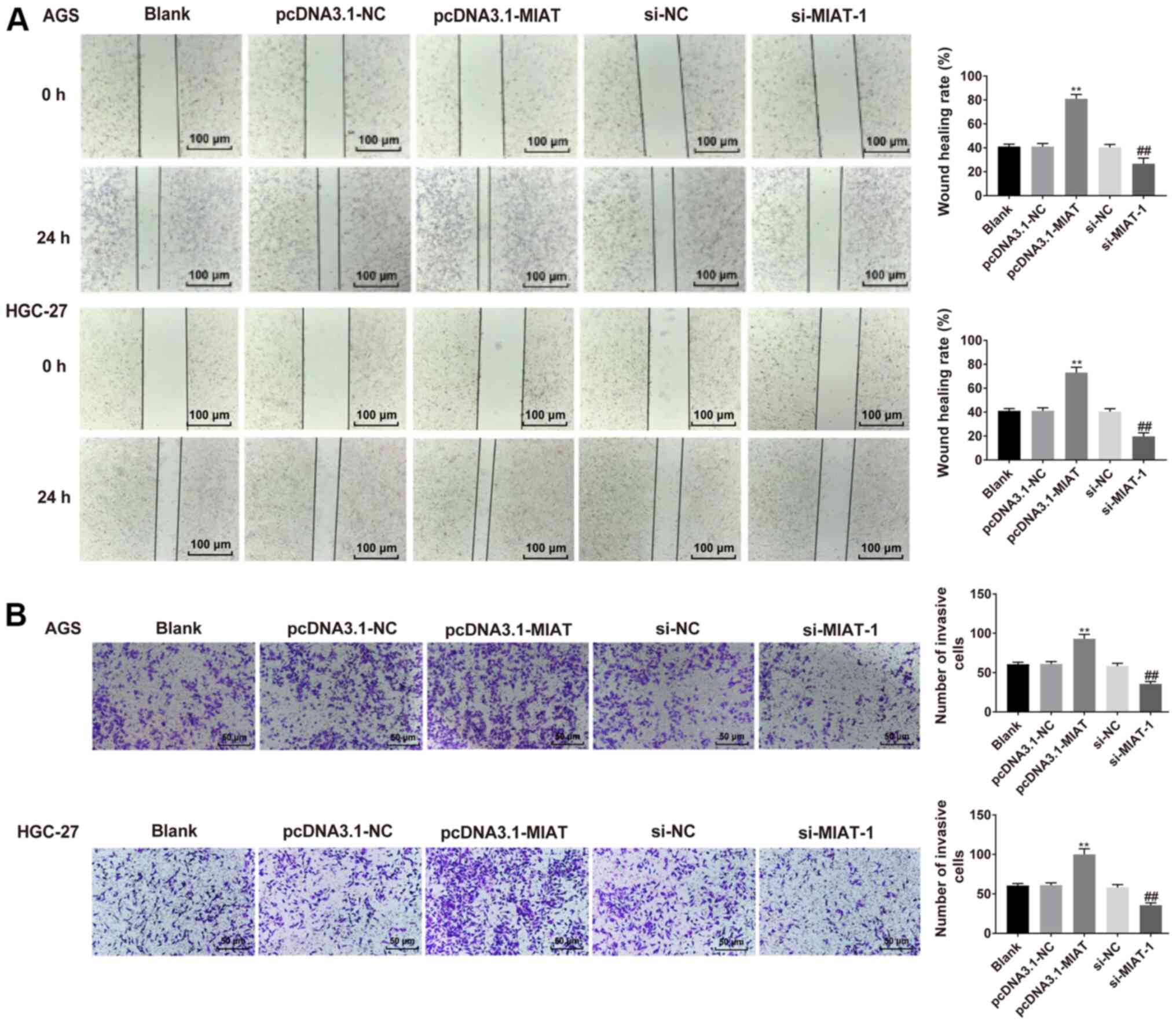
To determine the effect of MIAT on HGC-27 and AGS
cell migration and invasion, wound healing and Transwell assays
were performed. HGC-27 and AGS cell proliferation in serum-free
medium was assessed via the CCK-8 assay. HGC-27 and AGS cell
proliferation in serum-free medium was slightly lower compared with
in medium with 10% FBS (Fig. S2).
It was demonstrated that HGC-27 and AGS cells overexpressing MIAT
exhibited a faster closing of the scratch wound compared with the
NC group (Fig. 3A). MIAT
overexpression significantly promoted cell invasion relative to the
NC group. The opposite results were observed following MIAT
knockdown (Fig. 3B).
MIAT serves as a miR-331-3p sponge in
HGC-27 and AGS cells
Using StarBase 3.0, it was predicted that miR-331-3p
had binding sites complementary to MIAT (Fig. 4A). Overexpression of miR-331-3p
suppressed the luciferase activity of MIAT WT 3′-UTR in the HGC-27
and AGS cells (Fig. 4B). The
expression of miR-331-3p was downregulated in GC tissues compared
with adjacent tissues (Fig. 4C), and
miR-331-3p expression was negatively correlated with the expression
of MIAT in GC tissues (r=−0.5638; P<0.0001; Fig. 4D). miR-331-3p was also significantly
decreased in HGC-27 and AGS cells compared with the GES-1 cells
(Fig. 4E). Overexpression of MIAT
suppressed the expression of miR-331-3p, while MIAT knockdown
increased the expression of miR-331-3p in both AGS and HGC-27 cells
(Fig. 4F).
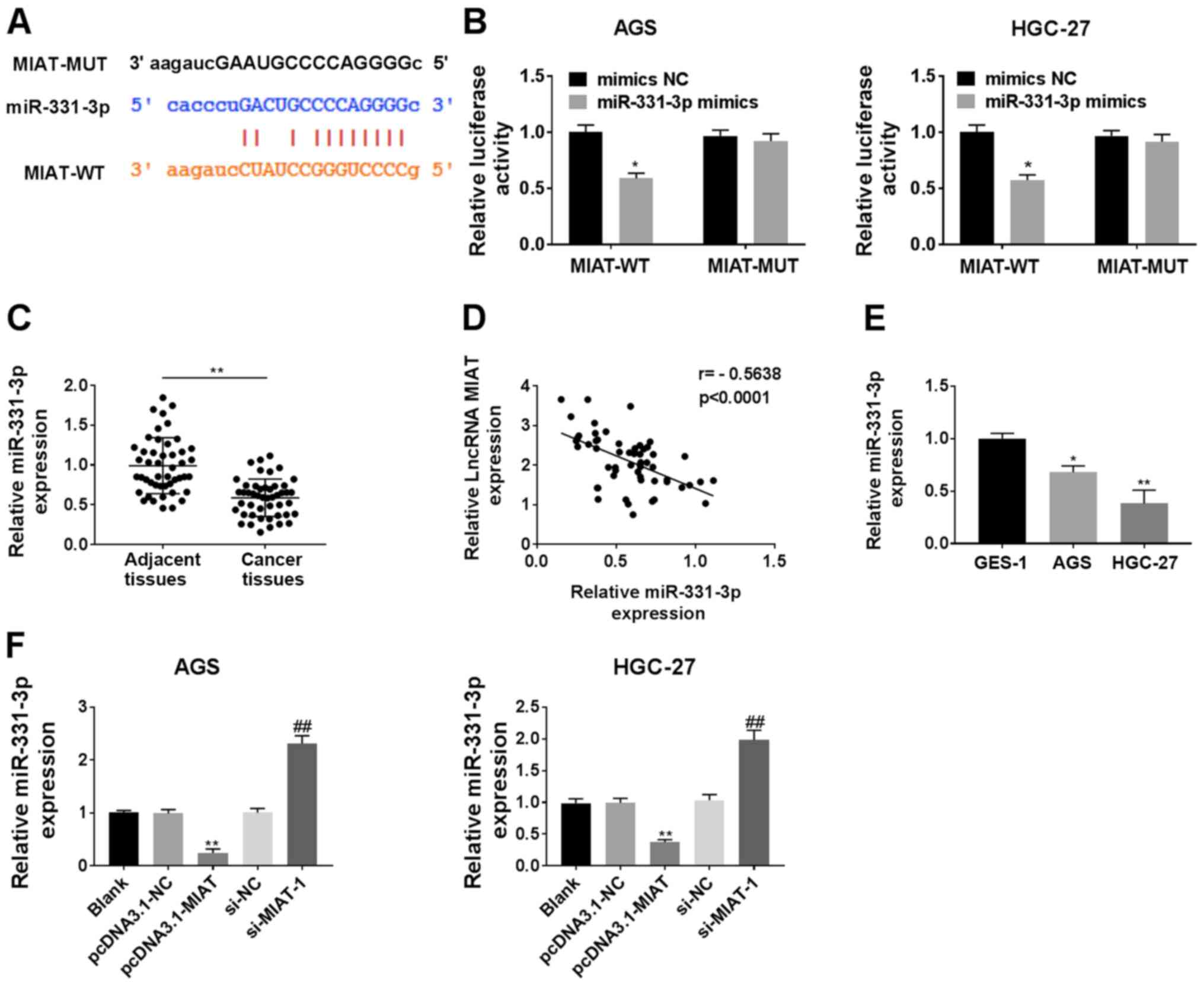 | Figure 4.MIAT serves as a miR-331-3p sponge in
GC cells. (A) The predicted binding sites between miR-331-3p and
MIAT 3′UTR. (B) The luciferase activities of the MIAT-WT or
MIAT-MUT reporter were detected via a dual-luciferase reporter
assay. *P<0.05 vs. mimics NC/MIAT-WT. (C) miR-331-3p levels were
measured cia RT-qPCR in 47 pairs of GC tissues and adjacent
tissues. n=3. **P<0.01 vs. adjacent tissues group. (D)
Correlation analysis between miR-331-3p and MIAT in 47 GC tissue
samples. (E) miR-331-3p levels were detected via RT-qPCR in
different cell lines (GES-1, HGC-27 and AGS). n=3. *P<0.05,
**P<0.01 vs. the GES-1 group. (F) HGC-27 and AGS cells were
transfected with pcDNA3.1-NC, pcDNA3.1-MIAT, si-NC or si-MIAT-1,
followed by the measurement of miR-331-3p levels. n=3. **P<0.01
vs. the blank or pcDNA3.1-NC group; ##P<0 .01 vs. the
blank or si-NC group. GC, gastric cancer; MIAT, myocardial
infarction associated transcript; si, small interfering; NC,
negative control; miR, micro RNA; RT-qPCR, reverse
transcription-quantitative PCR. |
MIAT promotes RAB5B expression via
miR-331-3p
Bioinformatics analysis using the TargetScan 7.2
revealed that miR-331-3p potentially binds to the 3′UTR of RAB5B
mRNA, which was significantly reduced by transfection of miR-331-3p
mimics (Fig. 5A and B). The
expression of RAB5B was markedly higher in GC tissues and HGC-27
and AGS cell lines (Fig. 5C and F,
respectively). A correlation analysis revealed that the mRNA
expression of RAB5B in GC tissues was inversely correlated with
miR-331-3p levels (r=−0.6545; P<0.0001) and positively
correlated with MIAT levels (r=0.6543; P<0.0001) (Fig. 5D and E, respectively). The expression
of miR-331-3p was upregulated in AGS and HGC-27 cells transfected
with miR-331-3p mimics, whereas it was downregulated in cells
transfected with miR-331-3p inhibitors (Fig. 5G). The mRNA and protein expression
levels of RAB5B were downregulated in HGC-27 and AGS cells
transfected with miR-331-3p mimics, whereas they were was
upregulated in AGS and HGC-27 transfected with miR-331-3p
inhibitors (Fig. 5H and I,
respectively). The mRNA and protein expression of RAB5B was
upregulated in AGS and HGC-27 transfected with pcDNA-RAB5B
(Fig. 5J and K, respectively).
Subsequently, the mRNA and protein expression of RAB5B were
determined after transfection of si-MIAT and pcDNA-RAB5B into
HGC-27 and AGS cells and it was demonstrated that the decreased
expression of RAB5B in response to si-MIAT was partially reversed
by RAB5B overexpression (Fig. 5L and
M).
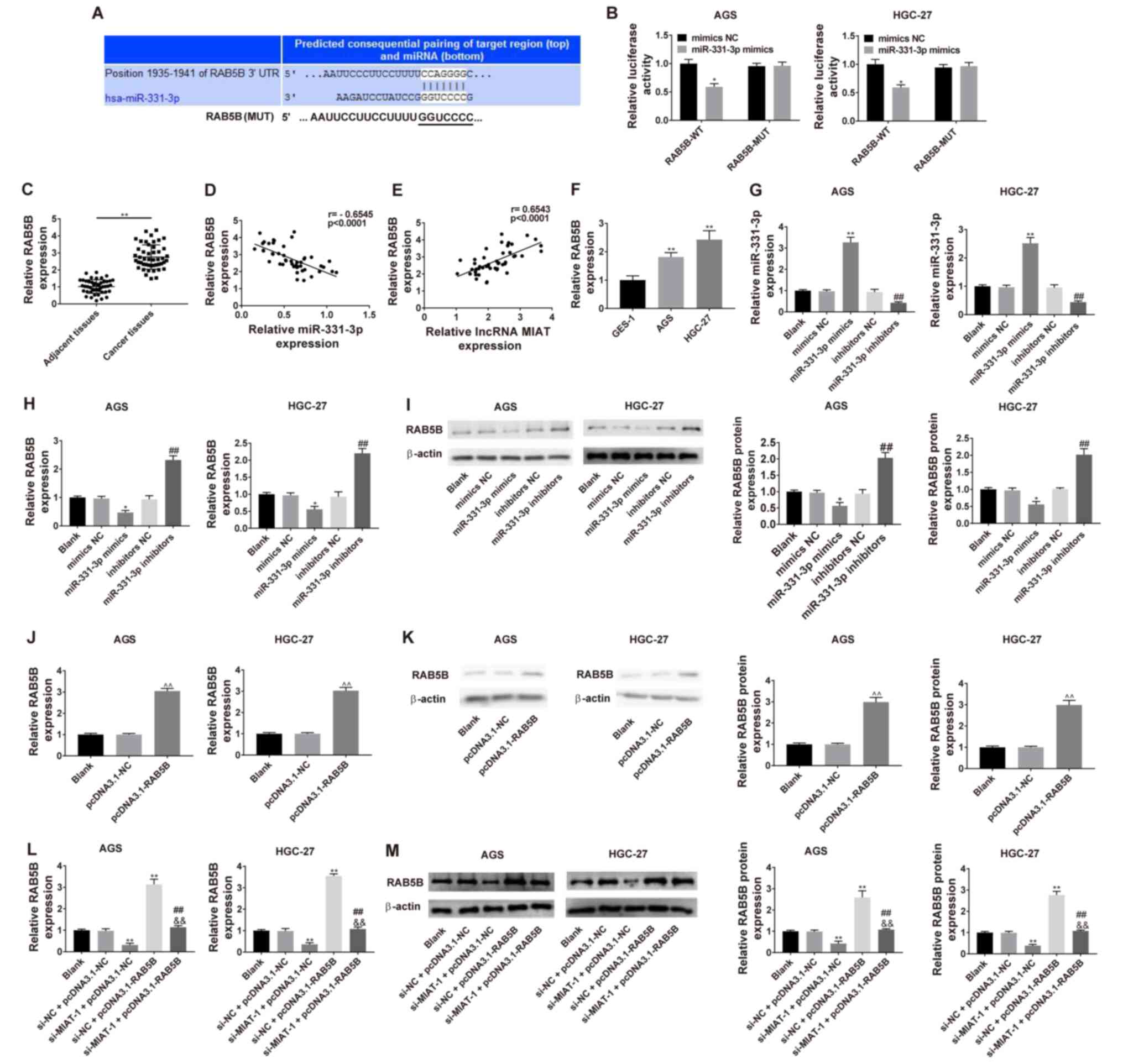 | Figure 5.Dual-luciferase reporter system
detection of the effect of miR-331-3p on RAB5B in HGC-27 and AGS
cells. (A) Predicted binding between miR-331-3p and RAB5B. (B)
Luciferase activities of RAB5B-WT or RAB5B-MUT reporter were
detected via dual-luciferase reporter assay. *P<0.05 vs. mimics
NC/RAB5B-WT. (C) RAB5B levels were measured via RT-qPCR in 47 pairs
of GC tissues and adjacent tissues. (D) Correlation analysis
between miR-331-3p and RAB5B in 47 GC tissue samples. n=3.
**P<0.01 vs. the adjacent tissues group. (E) Correlation
analysis between MIAT and RAB5B in 47 GC tissue samples. (F) RAB5B
levels were detected via RT-qPCR in different cell lines (GES-1,
HGC-27 and AGS). n=3. **P<0.01 vs. the GES-1 group. (G) HGC-27
and AGS cells were transfected with mimics NC, miR-331-3p mimics,
inhibitors NC or miR-331-3p inhibitors, followed by the measurement
of miR-331-3p level. HGC-27 and AGS cells were transfected with
mimics NC, miR-331-3p mimics, inhibitors NC or miR-331-3p
inhibitors, followed by the measurement of RAB5B level at the (H)
mRNA and (I) protein levels. n=3. *P<0.05 vs. the blank or
mimics NC group; ##P<0.01 vs. the blank or inhibitors
NC group. (J) HGC-27 and AGS cells were transfected with
pcDNA3.1-NC or pcDNA3.1-NC, followed by (K) the measurement of
RAB5B level. n=3. ^^P<0.01, compared with the blank
or pcDNA3.1-NC group. (L) Transfection efficiency in co-transfected
AGS and HGC-27 cells. (M) Measurement of RAB5B level at 48 h after
co-transfection. n=3. *P<0.05, **P<0.01 vs. the blank or
si-NC + pcDNA3.1-NC group; ##P<0.01 vs. the si-MIAT-1
+ pcDNA3.1-NC group; &&P<0.01 vs. the si-NC +
pcDNA3.1-RAB5B group. GC, gastric cancer; MIAT, myocardial
infarction associated transcript; si, small interfering; NC,
negative control; miR, micro RNA; WT, wild-type; MUT, mutant;
RT-qPCR, reverse transcription-quantitative PCR. |
MIAT knockdown effects are reversed by
miR-331-3p inhibitor or RAB5B overexpression in HGC-27 and AGS
cells
The results revealed that MIAT knockdown inhibited
cell proliferation (Fig. S3A) and
invasion (Fig. S3B) in HGC-27 and
AGS cells, while these functions were abrogated by miR-331-3p
inhibitor. It was also demonstrated that MIAT knockdown inhibited
cell proliferation (Fig. 6A),
induced cell apoptosis (Fig. 6B),
and inhibited migration (Fig. 6C)
and invasion (Fig. 6D) in HGC-27 and
AGS cells, while these functions were reversed by RAB5B
overexpression.
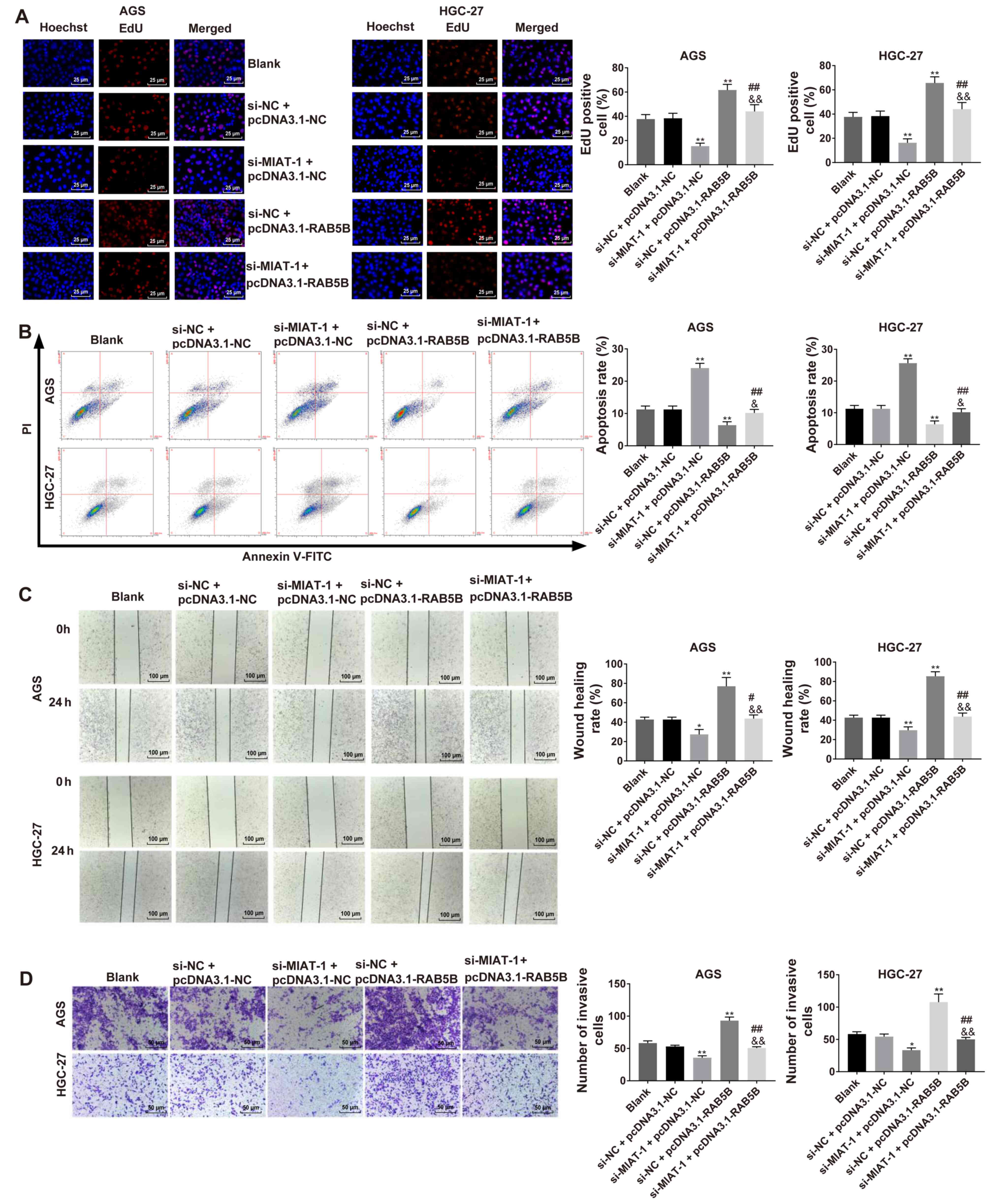 | Figure 6.Regulatory functions of MIAT
knockdown on regulating cell proliferation, apoptosis, migration
and invasion were reversed by RAB5B overexpression in GC cells. (A)
Cell proliferation was assessed via EdU assay. (B) Cell apoptosis
was evaluated via flow cytometry. (C) Cell migration capacity was
evaluated using a wound healing assay. (D) Cell invasion capacity
was evaluated by the Transwell invasion assay. n=3. *P<0.05,
**P<0.01 vs. the blank or si-NC + pcDNA3.1-NC group;
#P<0.05, ##P<0.01 vs. the si-MIAT-1 +
pcDNA3.1-NC group; &P<0.05,
&&P<0.01 vs. the si-NC + pcDNA3.1-RAB5B
group. GC, gastric cancer; MIAT, myocardial infarction associated
transcript; si, small interfering; NC, negative control; PI,
propidium iodide. |
Influence of MIAT overexpression on
cell proliferation, apoptosis, migration and invasion are reversed
by RAB5B knockdown in GES-1 cells
Overexpression of MIAT was induced in GES-1 cells
via transfection with pcDNA3.1-MIAT (Fig. 7A). The results revealed that
pcDNA3.1-MIAT downregulated miR-331-3p expression (Fig. 7B). Overexpression of MIAT upregulated
RAB5B mRNA and protein expression, and knockdown of RAB5B
downregulated RAB5B mRNA and protein expression (Fig. 7C and D, respectively). Subsequently,
RAB5B mRNA and protein expression levels were assessed after
transfection of the pcDNA3.1-MIAT and si-RAB5B into GES-1 cells and
it was revealed that the increased mRNA and protein expression of
RAB5B in response to pcDNA3.1-MIAT could be decreased by RAB5B
knockdown (Fig. 7E and F). It was
also demonstrated that MIAT overexpression induced cell viability
(Fig. 7G), inhibited cell apoptosis
(Fig. 7H), and promoted migration
(Fig. 7I) and invasion (Fig. 7J) in GES-1 cells, while these
functions were significantly reversed following RAB5B
knockdown.
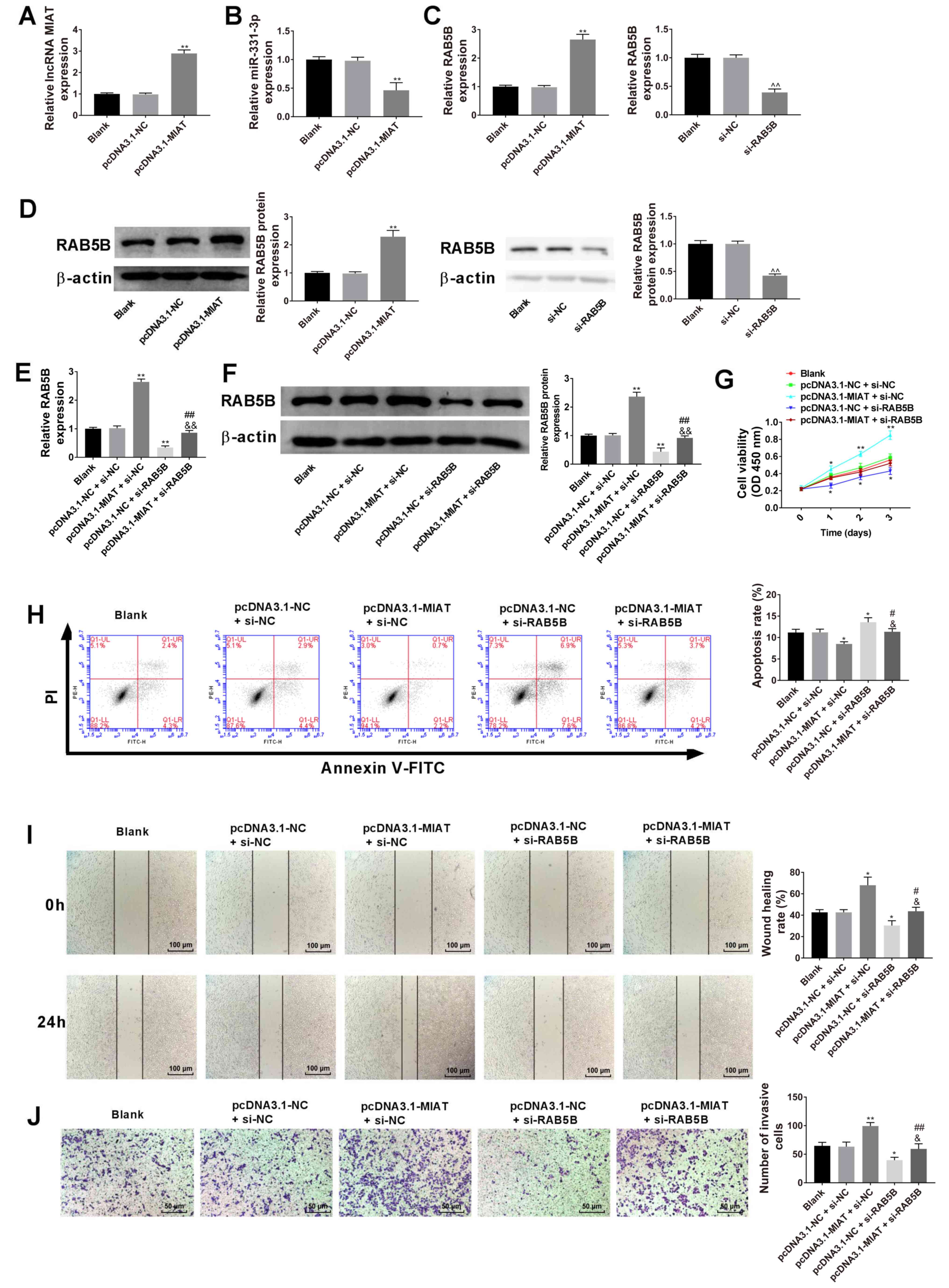 | Figure 7.Influence of MIAT-overexpression on
GES-1 cell proliferation, apoptosis, migration and invasion are
reversed by RAB5B-knockdown. (A) RT-qPCR analysis of MIAT
expression in the gastric epithelial mucosa cell line GES-1. (B)
RT-qPCR analysis of miR-331-3p expression in GES-1 cells. (C)
RT-qPCR and (D) western blot analysis of RAB5B expression in GES-1
cells, n=3. **P<0.01 vs. the blank or pcDNA3.1-NC group,
^^P<0.01 vs. the blank or si-NC group. (E) HGC-27 and
AGS cells were transfected with pcDNA3.1-NC, pcDNA3.1-MIAT, si-NC,
or si-RAB5B, followed by (F) the measurement of RAB5B level. (G)
Cell proliferation ability was assessed by the Cell Counting Kit-8
assay. (H) Cell apoptosis was evaluated by flow cytometry. (I) Cell
migration capacity was evaluated using a wound healing assay. (J)
Cell invasion capacity was evaluated by the Transwell invasion
assay. n=3. *P<0.05, **P<0.01 vs. the blank or pcDNA3.1-NC +
si-NC group; #P<0.05, ##P<0.01 vs. the
pcDNA3.1-MIAT + si-NC group; &P<0.05,
&&P<0.01 vs. the pcDNA3.1-NC + si-RAB5B
group. RT-qPCR, reverse transcription-quantitative PCR; GC, gastric
cancer; MIAT, myocardial infarction associated transcript; si,
small interfering; NC, negative control. |
Discussion
lncRNAs regulate gene expression via various
mechanisms, including transcriptional and post-transcriptional
processing, and have extensive regulatory functions in tumour
initiation and progression (23).
MIAT, a recently identified oncogenic lncRNA, has been reported to
be upregulated in several types of cancers, including papillary
thyroid cancer (7), lung cancer
(8) and acute myeloid leukemia
(24). However, the detailed role
and molecular mechanisms of MIAT in GC remain to be elucidated.
MIAT was revealed to be upregulated in GC tissues, which is
consistent with previous reports (25). In the present study, it was also
demonstrated that high expression of MIAT was correlated with late
TNM stage, lymphatic metastasis and a poor prognosis. Similarly,
upregulation of lncRNA LINC00858 is associated with a poor
prognosis in patients with GC (26).
High expression of LINC00858 is positively associated with TNM
stage and lymphatic metastasis (26). In the present study, functional
experiments revealed that MIAT knockdown inhibited HGC-27 and AGS
cell proliferation, induced GC cell apoptosis and inhibited HGC-27
and AGS cell migration and invasion. These results suggest that
MIAT serves an important role in the GC tumorigenesis and
metastasis.
Emerging evidence suggests that lncRNAs can serve as
competing endogenous RNAs (ceRNAs) to regulate miRNAs, subsequently
regulating expression of target genes (27). For instance, HLA-F-AS1 promotes
colorectal cancer progression by sponging miR-330-3p to upregulate
PFN1 expression (28).
Exosome-transmitted lncARSR functions as a sponge of miR-34/miR-449
to induce c-MET and AXL expression and mediates sunitinib
resistance in renal cell carcinoma (29). In the present study, it was revealed
that MIAT shares miR-331-3p response elements with RAB5B and
facilitates RAB5B expression via sponging miR-331-3p. RAB5B was
experimentally validated as a genuine target of miR-331-3p.
Functional inhibition of miR-331-3p effectively rescued the
decreased expression of RAB5B protein that was induced by MIAT
knockdown in HGC-27 and AGS cells, indicating that MIAT serves as a
ceRNA. Two other targets have been reported to serve roles in
gastric cancer downstream of MIAT. Knockdown of MIAT suppresses
cell biological behaviours in gastric cancer via a mechanism
involving the miR-29a-3p/HDAC4 axis (30). Moreover, MIAT promotes gastric cancer
growth and metastasis via regulation of the miR-141/DDX5 pathway
(25). The current results also
extend the regulatory mechanism of MIAT function.
Recent studies have reported the involvement of
miR-331-3p in cancer progression. miR-331-3p inhibits cell
proliferation and induces cell apoptosis in nasopharyngeal
carcinoma via targeting elF4B and blocks the PI3K-AKT signalling
pathway (31). Reduced expression of
miR-331-3p in ovarian cancer promotes proliferation and invasion,
due to upregulation of its target RCC2 (13). Guo et al (15) reported that miR-331-3p suppresses GC
cell growth via inhibiting E2F1. Zhao et al (32) revealed that miR-331-3p suppresses
cell proliferation in triple-negative breast cancer cells via
downregulating NRP2 (32). The
current results also supported the regulatory role of
miR-331-3p.
RAB5 has three isoforms (RAB5A, B and C) (33). Rab5B is a member of the Ras
superfamily of small Rab GTPases (34). RAB5B is localized at the plasma
membrane and early endosomes, and functions as a key regulator of
vesicular trafficking during early endocytosis (35). Inhibition of RAB5/7 efficiently
eliminates colorectal cancer stem cells and disrupts cancer foci
(36). RAB5B expression is elevated
in melanoma cells (37). RAB5B
regulates cell adhesion and migration by promoting Rac1 activation
and cancer cell migration (38).
Kong et al (39) reported
that RAB5B is directly downregulated by miR-130a-3p, and knockdown
of RAB5B inhibits cell proliferation, migration and invasion of
breast cancer cells. Wang et al (40) demonstrated that lncRNA-APC1
expression inhibits colorectal carcinoma cell growth, metastasis
and tumour angiogenesis via suppressing exosome production through
the direct binding of Rab5b mRNA. The present results further
confirmed that RAB5B serves a critical role in the progression of
GC.
In conclusion, the present study revealed that MIAT
is upregulated in GC, which is associated with poor clinical
outcomes. MIAT promotes HGC-27 and AGS cell proliferation via
RAB5B. MIAT promotes RAB5B activity via sponging miR-331-3p to
upregulate RAB5B expression. The present findings provide insight
into the MIAT/RAB5B pathway, and indicate it as a promising
potential therapeutic target in GC, suggesting important
translational implications.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JZ designed the study. XML, YYJ, BHL, HXW and RRW
performed the research and analyzed data. XML wrote the study. All
authors read and approved the final version of the manuscript.
Ethics approval and consent to
participate
The protocol of this research has been approved by
the Ethics Committee of Qing Dao Cheng Yang People's Hospital
(approval no. 20170106). All patients have signed written informed
consent.
Patient consent for publication
All patients agreed to the publication of the
article.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Selim JH, Shaheen S, Sheu WC and Hsueh CT:
Targeted and novel therapy in advanced gastric cancer. Exp Hematol
Oncol. 8:252019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Digklia A and Wagner AD: Advanced gastric
cancer: Current treatment landscape and future perspectives. World
J Gastroenterol. 22:2403–2414. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Maehara Y, Hasuda S, Koga T, Tokunaga E,
Kakeji Y and Sugimachi K: Postoperative outcome and sites of
recurrence in patients following curative resection of gastric
cancer. Br J Surg. 87:353–357. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nagano T and Fraser P: No-nonsense
functions for long noncoding RNAs. Cell. 145:178–181. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ishii N, Ozaki K, Sato H, Mizuno H, Saito
S, Takahashi A, Miyamoto Y, Ikegawa S, Kamatani N, Hori M, et al:
Identification of a novel non-coding RNA, MIAT, that confers risk
of myocardial infarction. J Hum Genet. 51:1087–1099. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang R, Zhao L, Ji L, Bai L and Wen Q:
Myocardial infarction associated transcript (MIAT) promotes
papillary thyroid cancer progression via sponging miR-212. Biomed
Pharmacother. 118:1092982019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lin D, Xu HP, Lin JH, Hu HH, Wang Q and
Zhang J: Long non-coding RNA MIAT promotes non-small cell lung
cancer progression by sponging miR-1246. Eur Rev Med Pharmacol Sci.
23:5795–5801. 2019.PubMed/NCBI
|
|
9
|
Liu Z, Wang H, Cai H, Hong Y, Li Y, Su D
and Fan Z: Long non-coding RNA MIAT promotes growth and metastasis
of colorectal cancer cells through regulation of miR-132/Derlin-1
pathway. Cancer Cell Int. 18:592018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Thomson DW and Dinger ME: Endogenous
microRNA sponges: Evidence and controversy. Nat Rev Genet.
17:272–283. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen X, Luo H, Li X, Tian X, Peng B, Liu
S, Zhan T, Wan Y, Chen W, Li Y, et al: miR-331-3p functions as an
oncogene by targeting ST7L in pancreatic cancer. Carcinogenesis.
39:1006–1015. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Buranjiang G, Kuerban R, Abuduwanke A, Li
X and Kuerban G: MicroRNA-331-3p inhibits proliferation and
metastasis of ovarian cancer by targeting RCC2. Arch Med Sci.
15:1520–1529. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhao D, Sui Y and Zheng X: MiR-331-3p
inhibits proliferation and promotes apoptosis by targeting HER2
through the PI3K/Akt and ERK1/2 pathways in colorectal cancer.
Oncol Rep. 35:1075–1082. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Guo X, Guo L, Ji J, Zhang J, Zhang J, Chen
X, Cai Q, Li J, Gu Q, Liu B, et al: miRNA-331-3p directly targets
E2F1 and induces growth arrest in human gastric cancer. Biochem
Biophys Res Commun. 398:1–6. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Daitoku H, Isida J, Fujiwara K, Nakajima T
and Fukamizu A: Dimerization of small GTPase Rab5. Int J Mol Med.
8:397–404. 2001.PubMed/NCBI
|
|
17
|
Igarashi T, Araki K, Yokobori T, Altan B,
Yamanaka T, Ishii N, Tsukagoshi M, Watanabe A, Kubo N, Handa T, et
al: Association of RAB5 overexpression in pancreatic cancer with
cancer progression and poor prognosis via E-cadherin suppression.
Oncotarget. 8:12290–12300. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang PS, Yin PH, Tseng LM, Yang CH, Hsu
CY, Lee MY, Horng CF and Chi CW: Rab5A is associated with axillary
lymph node metastasis in breast cancer patients. Cancer Sci.
102:2172–2178. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhao Z, Liu XF, Wu HC, Zou SB, Wang JY, Ni
PH, Chen XH and Fan QS: Rab5a overexpression promoting ovarian
cancer cell proliferation may be associated with APPL1-related
epidermal growth factor signaling pathway. Cancer Sci.
101:1454–1462. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
R Core Team, . R: A language and
environment for statistical computing. R Foundation for Statistical
Computing; Vienna: 2011
|
|
22
|
RStudio Team, . Integrated development
environment for R. RStudio, PBC; Boston, MA: 2015
|
|
23
|
Camacho CV, Choudhari R and Gadad SS: Long
noncoding RNAs and cancer, an overview. Steroids. 133:93–95. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang G, Li X, Song L, Pan H, Jiang J and
Sun L: Long noncoding RNA MIAT promotes the progression of acute
myeloid leukemia by negatively regulating miR-495. Leuk Res.
87:1062652019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sha M, Lin M, Wang J, Ye J, Xu J, Xu N and
Huang J: Long non-coding RNA MIAT promotes gastric cancer growth
and metastasis through regulation of miR-141/DDX5 pathway. J Exp
Clin Cancer Res. 37:582018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ai W, Li F, Yu HH, Liang ZH and Zhao HP:
Up-regulation of long noncoding RNA LINC00858 is associated with
poor prognosis in gastric cancer. J Gene Med. 22:e31492020.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chan JJ and Tay Y: Noncoding RNA:RNA
regulatory networks in cancer. Int J Mol Sci. 19:13102018.
View Article : Google Scholar
|
|
28
|
Huang Y, Sun H, Ma X, Zeng Y, Pan Y, Yu D,
Liu Z and Xiang Y: HLA-F-AS1/miR-330-3p/PFN1 axis promotes
colorectal cancer progression. Life Sci. 254:1171802020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Qu L, Ding J, Chen C, Wu ZJ, Liu B, Gao Y,
Chen W, Liu F, Sun W, Li XF, et al: Exosome-transmitted lncARSR
promotes sunitinib resistance in renal cancer by acting as a
competing endogenous RNA. Cancer Cell. 29:653–668. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li Y, Wang K, Wei Y, Yao Q, Zhang Q, Qu H
and Zhu G: lncRNA-MIAT regulates cell biological behaviors in
gastric cancer through a mechanism involving the miR-29a-3p/HDAC4
axis. Oncol Rep. 38:3465–3472. 2017.PubMed/NCBI
|
|
31
|
Xuefang Z, Ruinian Z, Liji J, Chun Z,
Qiaolan Z, Jun J, Yuming C and Junrong H: miR-331-3p inhibits
proliferation and promotes apoptosis of nasopharyngeal carcinoma
cells by targeting elf4B-PI3K-AKT pathway. Technol Cancer Res
Treat. 19:15330338198922512020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhao M, Zhang M, Tao Z, Cao J, Wang L and
Hu X: miR-331-3p suppresses cell proliferation in TNBC cells by
downregulating NRP2. Technol Cancer Res Treat.
19:15330338209058242020. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chua CE and Tang BL: The role of the small
GTPase Rab31 in cancer. J Cell Mol Med. 19:1–10. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wilson DB and Wilson MP: Identification
and subcellular localization of human rab5b, a new member of the
ras-related superfamily of GTPases. J Clin Invest. 89:996–1005.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Stenmark H: Rab GTPases as coordinators of
vesicle traffic. Nat Rev Mol Cell Biol. 10:513–525. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Takeda M, Koseki J, Takahashi H, Miyoshi
N, Nishida N, Nishimura J, Hata T, Matsuda C, Mizushima T, Yamamoto
H, et al: Disruption of endolysosomal RAB5/7 efficiently eliminates
colorectal cancer stem cells. Cancer Res. 79:1426–1437. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Peinado H, Alečković M, Lavotshkin S,
Matei I, Costa-Silva B, Moreno-Bueno G, Hergueta-Redondo M,
Williams C, García-Santos G, Ghajar C, et al: Melanoma exosomes
educate bone marrow progenitor cells toward a pro-metastatic
phenotype through MET. Nat Med. 18:883–891. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Díaz J, Mendoza P, Ortiz R, Díaz N, Leyton
L, Stupack D, Quest AFG and Torres VA: Rab5 is required in
metastatic cancer cells for Caveolin-1-enhanced Rac1 activation,
migration and invasion. J Cell Sci. 127:2401–2406. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kong X, Zhang J, Li J, Shao J and Fang L:
MiR-130a-3p inhibits migration and invasion by regulating RAB5B in
human breast cancer stem cell-like cells. Biochem Biophys Res
Commun. 501:486–493. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang FW, Cao CH, Han K, Zhao YX, Cai MY,
Xiang ZC, Zhang JX, Chen JW, Zhong LP, Huang Y, et al:
APC-activated long noncoding RNA inhibits colorectal carcinoma
pathogenesis through reduction of exosome production. J Clin
Invest. 129:727–743. 2019. View Article : Google Scholar : PubMed/NCBI
|