Introduction
Esophageal cancer (ESCA) is the eighth leading cause
of cancer-associated death worldwide. Approximately 480,000 new
cases and >400,000 ESCA-associated deaths occur every year, of
which >80% of ESCA cases occur in developing countries, and the
incidence and mortality of ESCA is highest in China (1,2).
Traditional treatment of ESCA, such as chemotherapy, radiation
therapy and immunotherapy, does not result in an improved prognosis
due to drug resistance and recurrence (3). Therefore, it is necessary to
investigate the molecular pathogenesis of ESCA and identify
effective prognostic biomarkers and molecular targets for the
treatment of ESCA.
MicroRNAs (miRNAs/miRs), a type of short non-coding
RNA molecule, are 21–25 nucleotides in length (4). Mature miRNAs can function as oncogenes
or tumor suppressors and participate in the regulation of the cell
proliferation, migration, apoptosis, metabolism and other cellular
responses by binding to the 3′-untranslated region (UTR) of target
genes via complementary base pair interactions (5,6). Thus,
exploring the underlying molecular mechanisms of miRNA functions is
a promising strategy to improve our understanding of the
pathogenesis of ESCA, and to identify novel biomarkers for ESCA
diagnosis and treatment.
miR-454-3p is a subunit of miR-454 family that
functions in tumor initiation, invasion and metastasis. miR-454-3p
has been shown to negatively modulate invasion and metastasis in
bladder cancer by targeting the tumor promoter zinc finger
E-box-binding homeobox 2 (ZEB2) (7).
In addition, miR-454-3p inhibits BTG1 expression levels and
inhibits cell cycle progression and metastasis in renal carcinoma
cells (8). However, to the best of
our knowledge, it is still unclear whether miR-454-3p is involved
in the proliferation and invasion of ESCA cells.
Insulin-like growth factor 2 mRNA-binding protein 1
(IGF2BP1), a member of the RNA-binding IGF2BP protein family
containing IGF2BP1, 2 and 3 (9), has
been observed to be highly expressed in a variety of cancer types,
including breast, ovarian, lung, liver, pancreatic and esophageal
cancer (10), and functions as a
tumor-promoting factor associated with metastasis and overall poor
prognosis in colorectal carcinoma (11). The interaction between IGF2BP1 and
its corresponding target mRNAs, such as actin, cytoplasmic 1,
c-MYC, CD44 and MKI67 enable the modulation of cell
proliferation and migration at the post-transcriptional level
(12,13). It was reported that the binding of
IGF2BP1 to miRNA-140-5p results in the suppression of growth and
metastasis of cervical cancer (14).
However, whether this binding between IGF2BP1 and miR-454-3p plays
important role in the progression of ESCA is still unclear.
Therefore, the present study investigated whether
IGF2BP1 may be a promising direct target of miR-454-3p in ESCA, and
explored how miR-454-3p-overexpression downregulated the
proliferation, migration and invasion of ESCA cells by targeting
IGF2BP1 in vitro. Additionally, the current study aimed to
understand how miR-454-3p affected tumor growth and metastasis in
an in vivo ESCA tumor mouse model.
Materials and methods
Collection of clinical tumor
specimens
Patient information (including overall survival
rate), ESCA tissues and matched adjacent normal tissues (5 cm from
the tumor) were collected from patients with ESCA (n=40).
Tumor-Node-Metastasis (TNM) stage and invasion depth classification
were determined as previously described (15). All patients provided written informed
consent. The study was approved by The Ethics Committee of
Affiliated Haian Hospital of Nantong University (Nantong, China).
Surgically removed tissue samples by tumor resection were frozen in
liquid nitrogen and stored at −80°C until use.
Cell culture and transfections
The human esophageal epithelial cell (HEEC) line was
obtained from ScienCell Research Laboratories, Inc. (cat. no.
2720), and four ESCA cell lines, EC9706, ECA109, TE-1 and TE-8,
were obtained from the American Type Culture Collection. All cells
were cultured in DMEM or RPMI1640 medium (both Gibco; Thermo Fisher
Scientific, Inc.) with 10% of FBS (Gibco; Thermo Fisher Scientific,
Inc., or Bio-Rad Laboratories, Inc.) and 1% penicillin (100
µg/ml)/streptomycin (100 U/ml) mix at 37°C in an incubator under an
atmosphere of 5% CO2.
miR-454-3p mimics and negative control (miR-NC) were
synthesized by Shanghai Gene Pharma Co., Ltd. and transfected into
EC9706 and TE-1 cells using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.). miR-454-3p inhibitors
(miR-454-3p inh) and matched negative control (NC inh) were
synthesized by Shanghai Gene Pharma Co., Ltd. and transfected into
ECA109 and TE-8 cells using Lipofectamine® 2000. After
24 h of incubation, further experiments were performed. For
co-transfection, miR-454-3p mimics or miR-NC and short hairpin
(sh)RNA-IGF2BP1 (sh-IGF2BP1) were co-transfected into TE-1 cells
using Lipofectamine® 2000. miR-454-3p inh or NC inh and
sh-IGF2BP1 were co-transfected into TE-8 cells using
Lipofectamine® 2000. After 24 h of incubation, further
experiments were performed. The cDNA3.1-IGF2BP1 plasmid was
constructed in-house by inserting the open reading frame of IGF2BP1
into a pcDNA3.1 plasmid (Thermo Fisher Scientific, Inc.) and then
transfected into 293T cells, as previously described (16).
RNA extraction and quantitative
reverse transcription (RT-q)PCR
Total RNAs were extracted from patients' tissues and
cultured cell lines using TRIzol® reagent (Thermo Fisher
Scientific, Inc.) then converted into complementary DNA using a
RevertAid RT Reverse Transcription kit (Thermo Fisher Scientific,
Inc.), which was used according to the manufacturer's instructions.
mRNA expression levels were quantified using qPCR and a Standard
SYBR-Green RT-PCR kit (Takara Bio, Inc.) in accordance with the
manufacturer's protocols. The thermocycling conditions were as
follows: 50°C for 30 min, followed by 95°C for 10 min and 40 cycles
of 15 sec at 95°C and 60 sec at 60°C. GAPDH was used as the control
gene. All of the related miRNA reagents used were purchased from
RiboBio Co., Ltd. The relative primers sequences were prepared
(Sangon Biotech Co., Ltd.) as follows: miR-454-3p, Forward:
5′-ACCCTATCAATATTGTCTCTGC-3′ and reverse:
5′-GCGAGCACAGAATTAATACGAC-3′; IGF2BP1, forward:
5′-CAAAGGAGCCGGAAAATTCAAAT-3′ and reverse:
5′-CGTCTCACTCTCGGTGTTCA-3′; U6, forward,: 5′-CTCGCTTCGGCAGCACA-3′
and reverse: 5′-AACGCTTCACGAAYYYGCGT-3′ and GAPDH, forward:
5′-AGACACCATGGGGAAGGTGAA−3′ and reverse:
5′-ATTGCTGATGATCTTGAGGCTG-3′. The 2−ΔΔCq method
(17) was used to calculate the
relative expression levels of miR-454-3p expression levels after
normalization to GAPDH small nuclear RNA.
Cell proliferation assay
Cell proliferation was analyzed using a Cell
Counting Kit-8 (CCK-8) according to the manufacturer's protocol.
CCK-8 was purchased from Dojindo Molecular Technologies, Inc. A
total of 4×103 EC9706, TE-1, ECA109 and TE-8 cells were
separately seeded in each well in 96-well plates. After incubation
at 37°C for 2 h, 20 µl CCK-8 was added to each well and incubated
for 1 h at 37°C. Viable cells were counted by measuring the optical
density value using a microplate reader at the wavelength of 450
nm. Each measurement was performed in triplicate in all experiments
and the mean of the results was used to draw the growth curves.
Colony formation assay
A total of 5×102 cells (EC9706, TE-1,
ECA109 and TE-8) were separately seeded in each well in 6-well
plates. After incubation for 14 days in RPMI1640 medium containing
10% FBS in a 37°C atmosphere of 5% CO2, the cells were
fixed with 4% paraformaldehyde for 1 h at room temperature and
stained using methanol and 0.1% crystal violet in 20% methanol for
15 min at room temperature. Finally, the cell numbers were counted
manually under a light microscope (magnification, ×40; Leica
Biosystems).
Cell migration and invasion
assays
Cell migration was analyzed using a wound healing
assay. Firstly, ESCA cells (EC9706 and TE-1 cells transfected with
miR-454-3p mimics or miR-NC, as well as ECA109 and TE-8 cells
transfected with miR-454-3p inh or NC inh) were seeded in 6-well
plates with 5×105 cells/well. After 24 h of incubation
at 37°C, the cell monolayer was removed using a micropipette tip
and washed with phosphate-buffered saline (PBS). Then, a plastic
scriber was used to scratch the wound and the cell monolayer was
incubated at 37°C in serum-free medium for 48 h. Finally, wound
closure was observed under a light microscope (magnification, ×40;
Leica Biosystems). Triple detection was performed and the mean of
results was calculated. Cell invasion was estimated using a
Transwell assay. Firstly, 1×104 ESCA cells were seeded
into the upper chamber coated with or without Matrigel (BD
Biosciences) at 37°C for 24 h. RPMI1640 medium containing 10% FBS,
as a chemoattractant, was added into the bottom chamber. Then, the
chamber was removed after 24 h incubation in a 37°C atmosphere of
5% CO2, followed by gently removing the matrix glue and
cells from the upper chamber with a wet cotton swab. Finally, cells
were fixed using 4% paraformaldehyde for 30 min at room temperature
and stained with crystal violet for 1 h at room temperature. After
drying, five visual fields were selected under a light microscope
(magnification, ×40) and the total cells were manually counted.
Bioinformatics analysis and luciferase
reporter assay
Bioinformatics analysis was performed using miRanda
(www.mirdb.org/), TargetScan (www.targetscan.org/vert_72/) and Pictar
(pictar.mdc-berlin.de/) to identify
potential target genes of miR-140-5p. To confirm the molecular
interaction between miR-454-3p and IGF2BP1, a luciferase reporter
assay was performed using human embryonic kidney 293T cells (Thermo
Fisher Scientific, Inc.). The pmirGLO-IGF2BP1-3′UTR wild-type
(IGF2BP1 3′UTR-WT) and pmirGLO-IGF2BP1-3′UTR mutant (IGF2BP1
3′UTR-MUT) were designed and obtained from Sangon Biotech Co., Ltd.
293T cells were seeded in 12-well plates with density of
1.5×105/well. Then, 30 nM miR-454-3p mimics or miR-NC
were co-transfected with 300 ng DNA (IGF2BP1 3′UTR-WT or IGF2BP1
3′UTR-MUT) into 293T cells using Lipofectamine 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) in accordance with the
manufacturer's instructions. After transfection for 24 h, the
relative luciferase activity was assessed using a Dual-Luciferase
Reporter Assay system (Promega Corporation). Relative firefly
luciferase activity was compared with Renilla luciferase
activity.
Western blot analysis
RIPA lysis buffer (9800; Cell Signaling Technology,
Inc.) was used to lyse and extract total protein from all ESCA cell
lines and patient tissues. After centrifugation at 16,128 × g for
10 min at 4°C, protein concentration was determined used a BCA
protein assay and proteins (20 ng/µl; 4 µl/lane) were separated via
10% SDS-PAGE. Subsequently, obtained samples were transferred to
polyvinylidene fluoride (PVDF) membranes and blocked with 5% milk
for 1 h at 37°C. Next, incubating the PVDF membrane with
anti-IGF2BP1 (cat. no. sc-166344; 1:100), anti-GAPDH (cat. no.
sc-47724; 1:1,000), anti-phosphorylated (p)-AKT (cat. no.
sc-101629; 1:2,000)/t-AKT (cat. no. sc-81434; 1:2,000),
anti-p-GSK3β (cat. no. sc-81496; 1:2,000)/t-GSK3β (cat. no.
sc-7291; 1:2,000) and anti-p-ERK1/2 (cat. no. sc-136521;
1:2,000)/t-ERK1/2 (cat. no. sc-135900; 1:2,000) antibodies (all
Santa Cruz Biotechnology, Inc.) at 37°C for 2 h. Then, PVDF
membranes were washed 3–5 times with 1X TBS-Tween buffer (1%
Tween-20) and co-incubated with HRP-conjugated anti-rabbit
secondary antibodies (cat. no. 7074; 1:5,000; Cell Signaling
Technology, Inc.) for 1 h at room temperature. ECL Western Blotting
Detection Reagents were purchased from Cytiva and used to detect
the visualized immunocomplexes. ImagePro software 6.0 (Media
Cybernetics, Inc.) was used to calculate the intensity of each
blots.
Immunohistochemistry (IHC) assay
Sections were fixed in 4% formaldehyde at room
temperature for 24 h and embedded into paraffin. Paraffin-embedded
sections (3-µm-thick) were immersed in xylene solution to dewax for
2 h at room temperature, and in 95% ethanol solution to gradient
rehydration. Then, 3% deionized water with hydrogen peroxide was
used for quenching. After recovering the antigens and blocking with
10% fetal bovine serum (cat. no. 26140087; Gibco; Thermo Fisher
Scientific, Inc.) in PBS at 37°C for 1 h, the sections were
incubated with anti-IGF2BP1 antibody (1:100; cat. no. HPA021367;
Sigma-Aldrich; Merck KGaA) overnight at 4°C, and subsequently
incubated with HRP-conjugated secondary antibody (cat. no. 7074;
1:5,000; Cell Signaling Technology, Inc.) for 30 min at 37°C.
Finally, the sections were stained with DAB for 1 h at 37°C, then
counterstained with hematoxylin for 1 h at 37°C. The section were
observed using confocal microscopy at a magnification of ×10.
Flow cytometry analysis for cell
apoptosis
Cell apoptosis was analyzed using a Dead Cell
Apoptosis kit (cat. no. V13242; Invitrogen; Thermo Fisher
Scientific, Inc.). Four transfected cell lines (EC9706, TE-1,
ECA109 and TE-8) were separately planted into 6-well plates with a
density of 8×104 in each well. After 48 h incubation at
37°C, the cells were harvested and subsequently re-suspended in 500
µl PBS buffer and stained with 5 µl Alexa Fluor 488-annexin V and 1
µl PI solution (100 µg/ml) in each 100 µl of cell suspension at
room temperature for 15 min. Apoptosis was measured using an Attune
NxT flow cytometer (Thermo Fisher Scientific, Inc.) and analyzed
with the Attune NxT software v2.6 (Thermo Fisher Scientific,
Inc.).
In vivo tumor xenograft in nude
mice
A total of 20 four-week old BALB/c-nude mice (male,
20–22 g) were purchased from the Shanghai Laboratory Animal Center.
There were five mice in each group. All mice had access to food and
water ad libitum, and the temperature in the cages was
maintained at 24±1°C with a 12/12 h light/dark cycle and 40–50%
humidity. All the procedures involved in experimental animals were
approved by The Ethics Committee of Affiliated Haian Hospital of
Nantong University. All the animal welfare considerations were
according to the international animal experiment standard. Next,
1×107 ESCA cells (TE-1 cell line) in 0.2 ml PBS buffer
were subcutaneously injected in the stomach of each mouse. Then the
mice were injected intraperitoneally with agomiR-miR-454-3p (equal
to miR-454-3p mimics) or agomiR-NC (equal to miR-NC) every 7 days
for 4 weeks. No mice died during the experiment. After successfully
obtaining a stable mouse model, the tumor size and volume [volume
(mm3)=width2 (mm2) × length
(mm)÷2] was measured every 3 days until the diameter of tumors were
~20 mm in control group (agomiR-NC treatment group). Then the mice
were euthanized by intravenous injection with 200 mg/kg
pentobarbital after 4 weeks of tumor growth, followed by cervical
dislocation. Finally, esophageal tissues were isolated and used for
the aforementioned tissue analyses.
Statistical analysis
All experiments were performed at least three times
and mean values were calculated. SPSS version 19.0 (IBM Corp.)
software was used to analyze the data, which was presented as the
mean ± SD. Kaplan-Meier analysis with log-rank test was performed
using GraphPad Prism 8.0 (GraphPad Software, Inc.) and was used to
determine the association between miR-454-3p expression and patient
survival rates. Spearman's correlation analysis was performed to
identify the correlation between miR-454-3p and IGF2BP1 expression.
The difference between groups was tested by unpaired or paired
(tumor vs. normal tissues) Student's t-test or one-way ANOVA with
Tukey's post hoc test. In addition, t-tests were used to determine
the associations between miR-454-3p expression and the
clinicopathological parameters. P<0.05 was considered to
indicate a statistically significant difference.
Results
Low miR-454-3p expression in human
ESCA tissues and association with patient survival
In the present study, the association between
miR-454-3p and the pathogenesis of ESCA and patient survival were
investigated. Histological staining was used for the isolated ESCA
tissues and normal tissues from 40 patients with ESCA. Two
high-power visual fields (magnification, ×200 and ×400) were
selected. It was observed that compared with normal tissues, ESCA
cells had larger volume and were pleomorphic, varying greatly in
size and shape, and the ratio of nucleus to cytoplasm in tumor
cells was different from that of normal cells (Fig. 1A). In addition, the mRNA expression
of miR-454-3p was significantly downregulated in tumor tissues
compared with that in adjacent normal tissues (Fig. 1B; P<0.001). Subsequently, the
association between clinicopathological characteristics and
different miR-454-3p expression in patients with ESCA were
analyzed. The results showed that either low or high expression of
miR-454-3p had no association with the characteristics of patients'
age, sex and cellular differentiation. However, miR-454-3p
expression was significantly associated with Tumor-Node-Metastasis
(TNM) stage (P=0.014), invasion depth classification (P=0.026) and
lymph node metastasis (P=0.029) (all Table I). Additionally, the association
between miR-454-3p expression and patients' survival rates was
analyzed. Kaplan-Meier analysis demonstrated that patients with
high miR-454-3p expression (n=17) had improved survival rates
compared with patients with low expression (n=23); this indicated
that miR-454-3p might be a tumor suppressor for ESCA (Fig. 1C). The expression differences of
miR-454-3p between normal human esophageal epithelial cells and
four ESCA cells (EC9706, ECA109, TE-1 and TE-8) were further
evaluated using RT-qPCR. Consistent with previous results, the
expression levels of miR-454-3p in normal HEECs were significantly
higher compared with those in four ESCA cell lines (Fig. 1D).
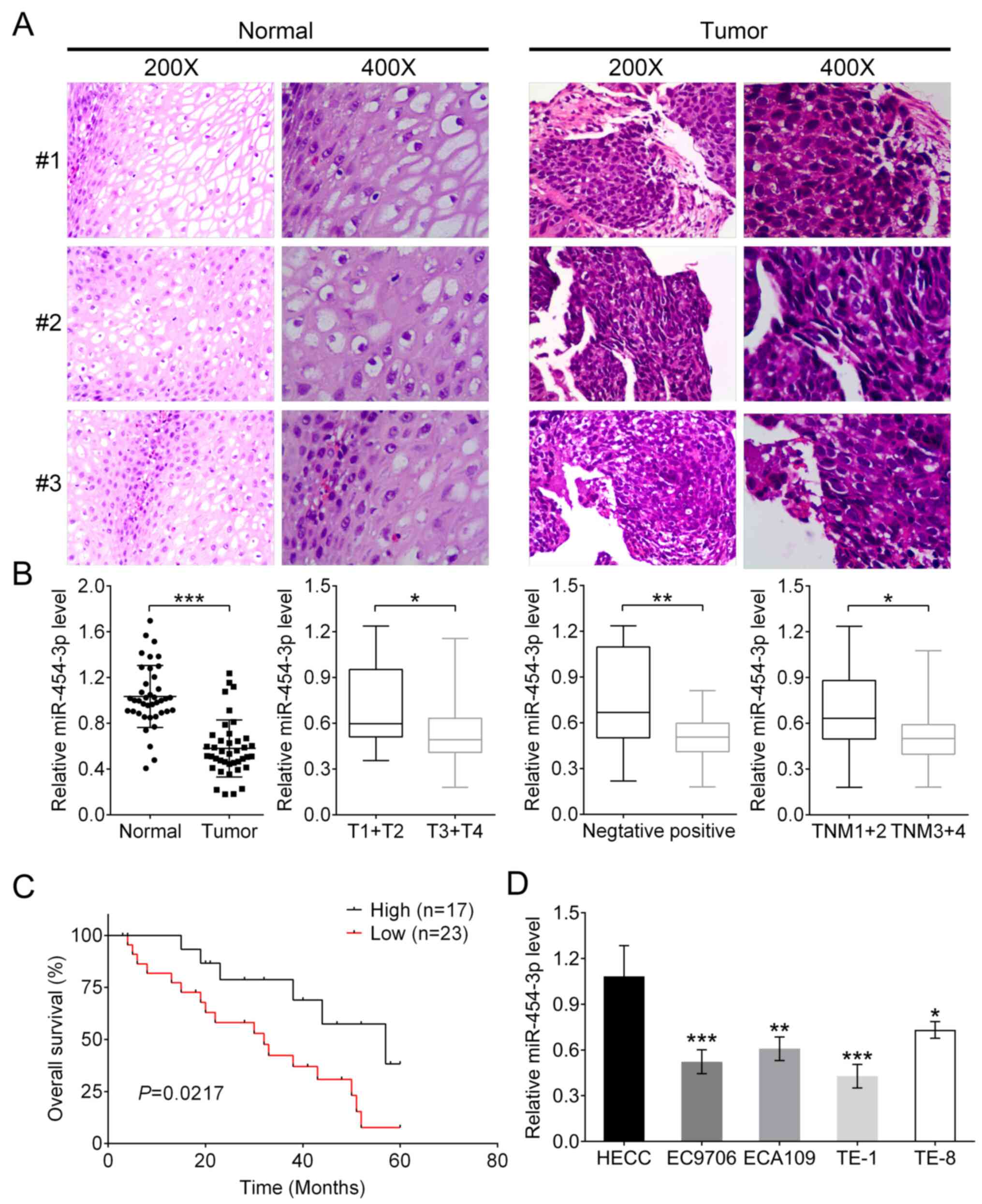 | Figure 1.miR-454-3p expression is low in ESCA
tissues and cells and associated with the overall survival of
patients. (A) Representative images of hematoxylin and eosin
stained ESCA tissues and matched adjacent normal tissues. (B)
RT-qPCR was used to analyze the expression difference of miR-454-3p
between tumor tissues and adjacent normal tissues in 40 patients as
well as the association between various clinicopathological
parameters, including invasive depth, lymph node metastasis and TNM
stages, and miR-454-3p expression. (C) Overall survival analysis of
miR-454-3p expression in patients with ESCA. (D) RT-qPCR was used
to analyze the expression difference of miR-454-3p in normal human
esophageal epithelial cells and four ESCA cell lines (EC9706,
ECA109, TE-1 and TE-8). All of the aforementioned results are
presented as mean ± SD. *P<0.05, **P<0.01 and ***P<0.001.
miR, microRNA; ESCA, esophageal cancer; RT-qPCR, reverse
transcription-quantitative PCR; TNM, Tumor-Node-Metastasis. |
 | Table I.The correlation between
clinicopathological characteristics and miR-454-3p expression in
ESCA patients (n=40). |
Table I.
The correlation between
clinicopathological characteristics and miR-454-3p expression in
ESCA patients (n=40).
| Characteristic | Value, % | miR-454-3p
expression | P-value |
|---|
| Age, years |
|
|
|
|
≥60 | 24 | 5.45±1.03 | 0.821 |
|
<60 | 16 | 5.53±1.17 |
|
| Sex |
|
|
|
|
Male | 23 | 5.31±1.07 | 0.248 |
|
Female | 17 | 5.71±1.06 |
|
|
Differentiation |
|
|
|
|
Well/Moderate | 18 | 5.36±1.22 | 0.551 |
|
Poor | 22 | 5.57±0.99 |
|
| TNM stage |
|
|
|
| Stage
I/II | 16 | 4.98±1.04 | 0.014a |
| Stage
III/IV | 24 | 5.81±0.97 |
|
| Invasive depth |
|
|
|
|
T1/T2 | 14 | 4.97+0.96 | 0.026a |
|
T3/T4 | 26 | 5.75±1.04 |
|
| Lymph node
metastasis |
|
|
|
|
Negative | 13 | 5.09±1.09 | 0.029a |
|
Positive | 27 | 5.67±1.03 |
|
Overexpression of miR-454-3p
suppresses ESCA cell proliferation and promotes apoptosis in
vitro
To investigate the effect of miR-454-3p on the
proliferation and apoptosis of ESCA cells, EC9706, ECA109, TE-1 and
TE-8 cells were stably transfected with either miR-454-3p mimics or
inhibitors. RT-qPCR analysis revealed that EC9706 and TE-1 cells
that transfected with miR-454-3p mimics displayed higher expression
of miR-454-3p compared with that of miR-NC-transfected cells, and
that ECA109 and TE-8 cells treated with miR-454-3p inhibitors
possessed lower expression levels of miR-454-3p compared with those
of NC inh-transfected cells (Fig.
2A). In addition, the effect of miR-454-3p on the cell
proliferation was detected using a CCK-8 and colony formation
assay. The results of the CCK-8 assay indicated that the cell
proliferation rate in miR-454-3p mimics-transfected EC9706 and TE-1
cells was significantly decreased (P<0.05). Meanwhile, the cell
proliferation rate of ECA109 and TE-8 cells treated with miR-454-3p
inhibitors was significantly increased (P<0.05) (both Fig. 2B). The colony formation assay
indicated that the number of colonies formed by EC9706 and TE-1
cells transfected with miR-454-3p mimics and ECA109 and TE-8 cells
transfected with NC inhibitors were less compared with those in the
control groups (Fig. 2C). Therefore,
it was speculated that miR-454-3p negatively regulated cell
proliferation in ESCA. However, whether miR-454-3p affected the
apoptosis of ESCA cells needed to be verified. Flow cytometry was
performed to detect the rate of apoptosis in EC9706, TE-1, ECA109
and TE-8 cells transfected with miR-454-3p mimics or miR-454
inhibitors (Fig. 2D). The data
demonstrated that miR-454-3p-overexpression significantly promoted
apoptosis. In conclusion, these results showed that miR-454-3p
inhibited the proliferation of ESCA cells.
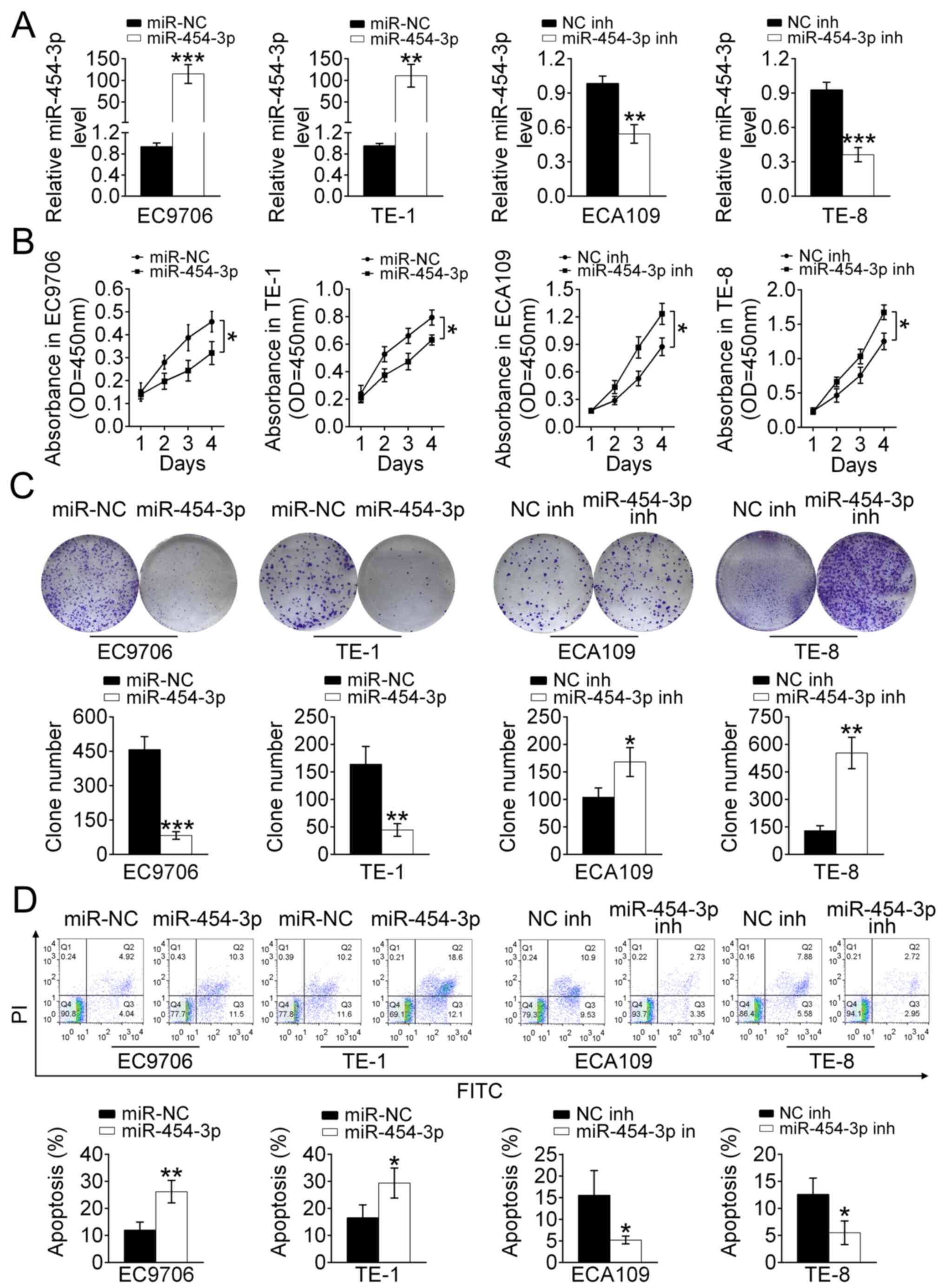 | Figure 2.Overexpression of miR-454-3p
suppresses the proliferation and promotes the apoptosis of ESCA
cells in vitro. (A) Reverse transcription-quantitative PCR
was used to analyze the relative expression levels of miR-454-3p in
EC9706 and TE-1 cells transfected with miR-454-3p mimics/miR-NC as
well as in ECA109 and TE-8 cells transfected with miR-454-3p inh/NC
inhibitors (NC inh). (B) A Cell Counting Kit-8 assay was used to
evaluate the proliferation of EC9706, TE-1, ECA109 and TE-8 cells
transfected with miR-454-3p plasmid/NC mimics or miR-454 inhs/NC
inh for 1, 2, 3 and 4 days. (C) Colony formation assays
demonstrated the effect of miR-454-3p on the proliferation of
EC9706, TE-1, ECA109 and TE-8 cells transfected with miR-454-3p
mimics/miR-NC or miR-454-3p inh/NC inh and the colony number was
quantified after transfection. (D) The apoptotic EC9706, TE-1,
ECA109 and TE-8 cells transfected with miR-454-3p mimics/miR-NC or
miR-454-3p inh/NC inh were determined using flow cytometry
analysis. Results are presented as mean ± SD. *P<0.05,
**P<0.01 and ***P<0.001 vs. respective control. miR,
microRNA; ESCA, esophageal cancer; NC, negative control; inh,
inhibitor; miR-NC, negative control mimic. |
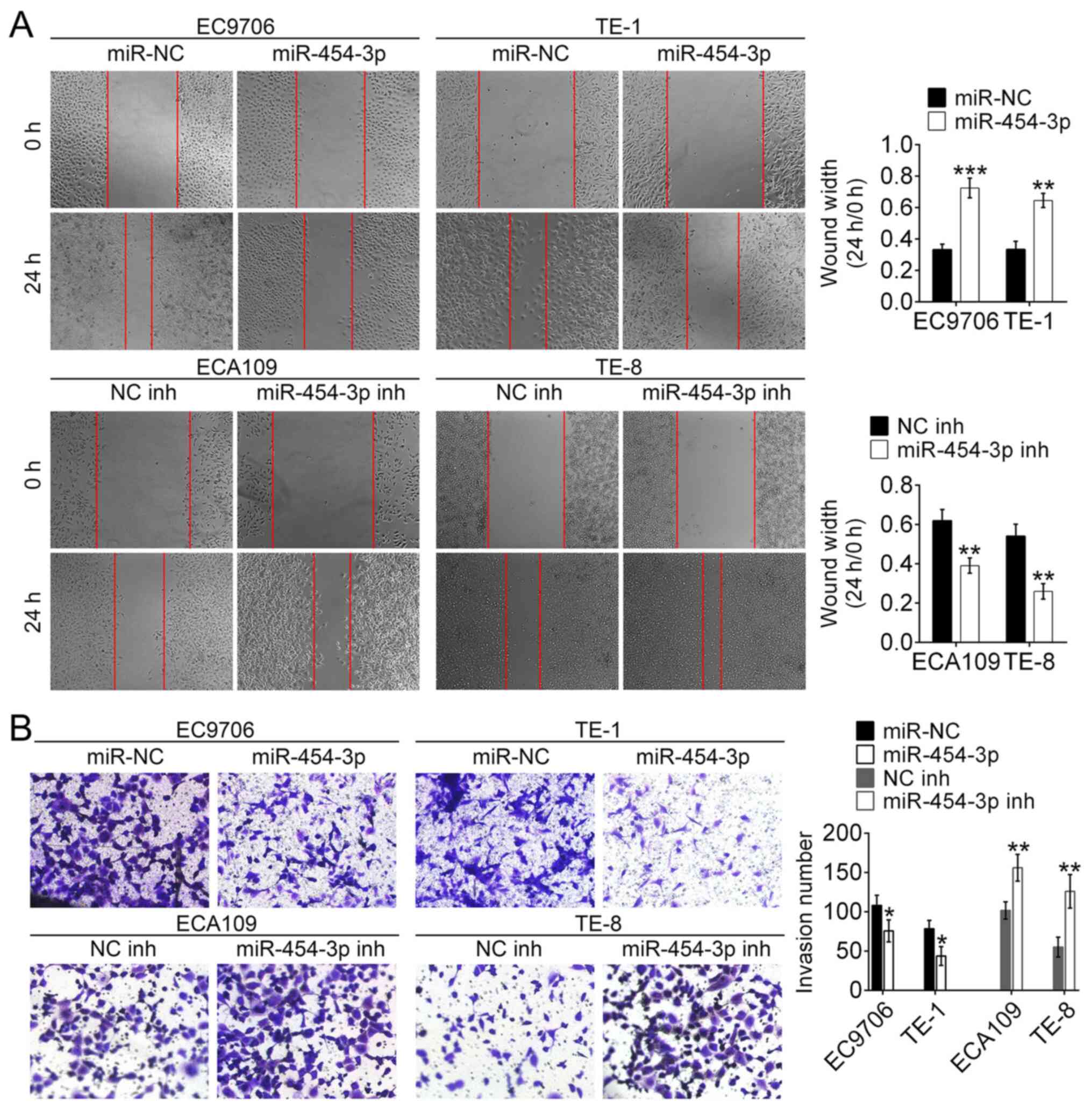
miR-454-3p inhibits the migration and
invasion in ESCA cells in vitro
To identify the role of miR-454-3p in the migration
and invasion of ESCA cells, wound healing and Matrigel assays were
conducted. The wound healing assay revealed that the wound width of
EC9706 and TE-1 cells transfected with miR-454-3p mimics was
significantly wider compared with that of miR-NC-transfected cells.
This was the opposite to the results of ECA109 and TE-8 cells
transfected with miR-454-3p inhibitors/NC inhibitors (Fig. 3A). These results demonstrated that
miR-454-3p overexpression could inhibit migration of ESCA cells.
Additionally, the Matrigel assay revealed the number of invasive
EC9706 and TE-1 cells treated with miR-454-3p mimics were fewer
compared with the number of invasive EC9706 and TE-1 cells
transfected with miR-NC. Conversely, the number of invasive EC9706
and TE-1 cells treated with miR-454-3p inhibitors were higher
compared with the number of invasive ECA109 and TE-8 cells
transfected with NC-inh (Fig. 3B).
These results illustrated that overexpression of miR-454-3p could
significantly suppress the invasion and migration of ESCA cells.
Taken together, these data suggested that miR-454-3p functions as
an inhibitor of the migration and invasion of ESCA cells.
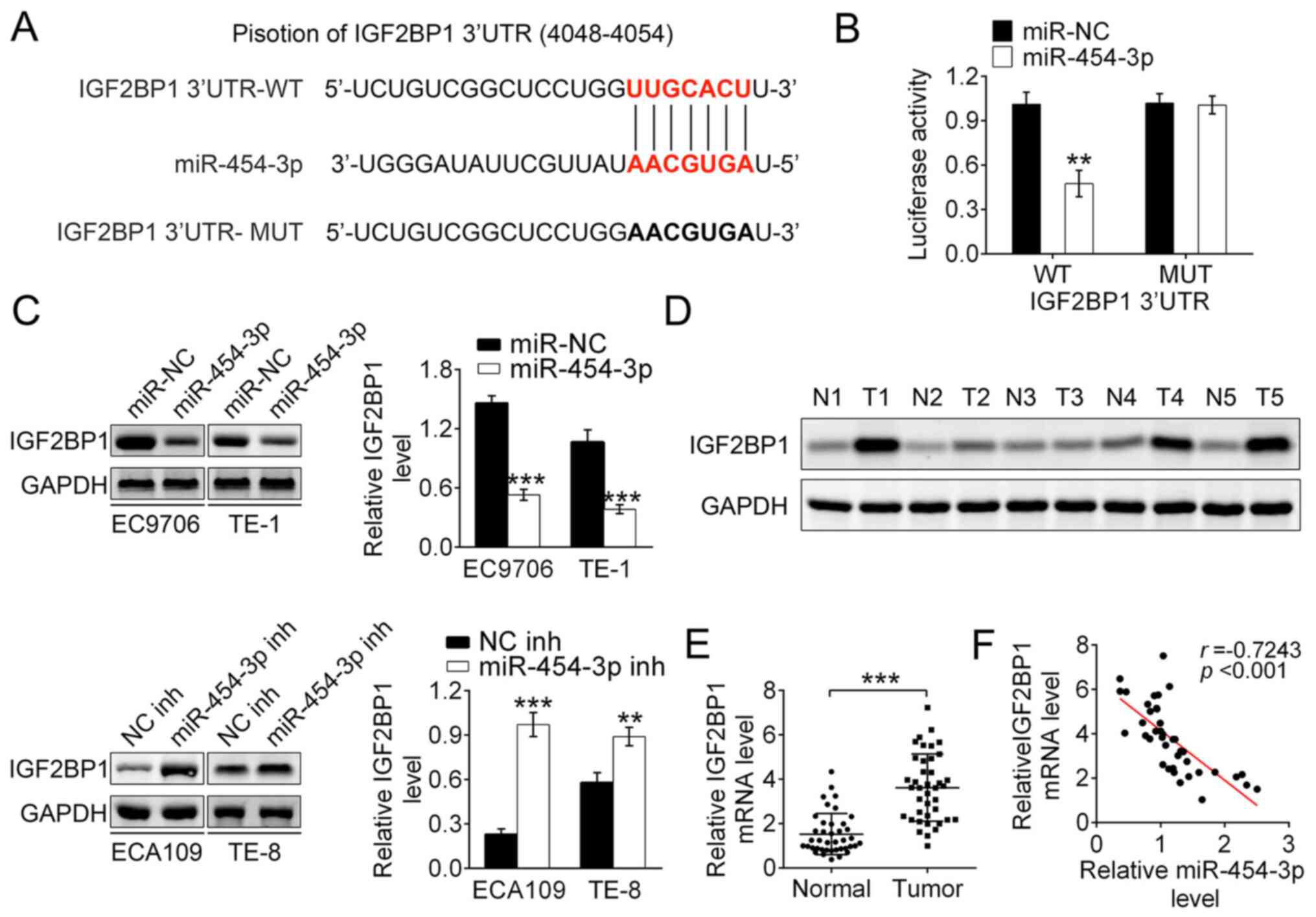
IGF2BP1 is the direct target of
miR-454-3p in ESCA
According to bioinformatics predication with
miRanda, TargetScan and Pictar, the IGF2BP1 gene was identified as
the target gene of miR-140-5p (Fig.
4A). Then luciferase reporter assays were used to detect the
binding affinity of miR-454-3p to the wild-type 3′UTR of IGF2BP1 in
293T cells. The results showed that the luciferase activity in
miR-454-3p mimics-transfected 293T cells was significantly
decreased. Notably, miR-454-3p-overexpression did not change the
luciferase activity of mutant IGF2BP1 3′UTR (Fig. 4B). Therefore, it was speculated that
IGF2BP1 3′UTR-WT was the direct target of miR-454-3p. To explore
how miR-454-3p regulated the expression of IGF2BP1, western blot
analysis was performed. The results showed that the miR-454-3p
mimics transfection group had significantly decreased expression of
IGF2BP1 both in EC9706 and TE-1 cells. Conversely, EC109 and TE-8
cells transfected with miR-454-3p inhibitors had increased
expression of IGF2BP1. Thus, these data demonstrated that
miR-454-3p negatively regulated the expression of IGF2BP1 (Fig. 4C). Additionally, the expression
differences of IGF2BP1 between tumor tissues and normal adjacent
tissues in five patients with ESCA were explored using western
blotting. It was reported that the protein expression of IGF2BP1 in
tumor tissue was significantly higher compared with those in normal
tissues, which indicated that upregulated IGF2BP1 promoted the
development of ECSA (Fig. 4D).
RT-qPCR analysis reported that the mRNA expression of
IGF2BP1 was higher in tumor tissues compared with that in
adjacent normal tissues, which was consistent with previous results
(Fig. 4E). The data showed a
significant inverse correlation between IGF2BP1 and miR-454-3p
expression in tumor tissues using Spearman's correlation analysis
(r=0.7243, P<0.001; Fig. 4F).
Taken together, these results demonstrated that IGF2BP1 is the
downstream target gene of miR-454-3p and that downregulated
miR-454-3p leads to the upregulation of IGF2BP1 in ESCA.
miR-454-3p suppresses ESCA cell
proliferation, migration and apoptosis by targeting IGF2BP1 in
vitro
To explore whether the tumor inhibitor function of
miR-454-3p is mediated by IGF2BP1, TE-1 and TE-8 cells were stably
transfected with IGF2BP1, miR-454-3p or their inhibitors,
respectively.
The CCK-8 and colony formation assays revealed that
transfection with miR-454-3p mimics significantly inhibited the
proliferation of both TE-1 and TE-8 cells, but this inhibition was
reversed by IGF2BP1-overexpression. (Fig. 5A and B). Hence, it was hypothesized
that miR-454-3p inhibited proliferation by targeting IGF2BP1 in
ESCA cells. Apoptosis was detected using flow cytometry analysis of
stably transfected TE-1 and TE-8 cells, and the results showed that
miR-454-3p-overexpression promoted apoptosis and
IGF2BP1-overexpression suppressed the apoptosis (Fig. 5C). These results suggested that
miR-454-3p may function as a tumor suppressor by targeting IGF2BP1.
The wound healing assay showed that high expression of miR-454-3p
maintained a significantly wider wound width and this results was
reversed by IGF2BP1-overexpression in both TE-1 and TE-8 cells.
These results suggested that miR-454-3p-overexpression inhibited
cell migration by targeting ICF2BP1 (Figs. 5D and S1). Additionally, the Transwell assay
demonstrated that miR-454-3p-overexpression significantly decreased
the invasive cell number and that this effect could be reversed by
additional IGF2BP1 transfection in both TE-1 and TE-8 cells
(Fig. 5E). Therefore, it was
revealed that miR-140-5p suppressed ESCA cell migration and
invasion in vitro by targeting ICF2BP1. A previous study
reported the inhibition effect of miR-9 on the ERK and AKT pathways
was mediated by targeting IGF2BP1 in hepatocellular carcinoma
(17), thus the present study
explored whether miR-454-3p acted as a suppressor of these pathways
through targeting IGF2BP1 in ESCA. Western blot analysis revealed a
significant downregulation of phosphorylated proteins, such as
p-AKT, p-GSK3 and p-ERK1/2, in TE-1 cells that were transfected
with miR-454-3p mimics, and this decrease was recovered by
IGF2BP1-overexpression. Moreover, the expression levels of these
phosphorylated proteins were increased by transfection with
miR-454-3p inhibitors in TE-8 cells, and this tendency was reversed
by IGF2BP1-overexpression (Fig. 5F).
Therefore, it was speculated that miR-454-3p suppressed the ERK and
AKT pathways through targeting IGF2BP1 in ESCA. In a summary, these
results showed that miR-454-3p suppressed ESCA cell proliferation,
migration and apoptosis by targeting IGF2BP1 via the ERK and AKT
pathways in vitro.
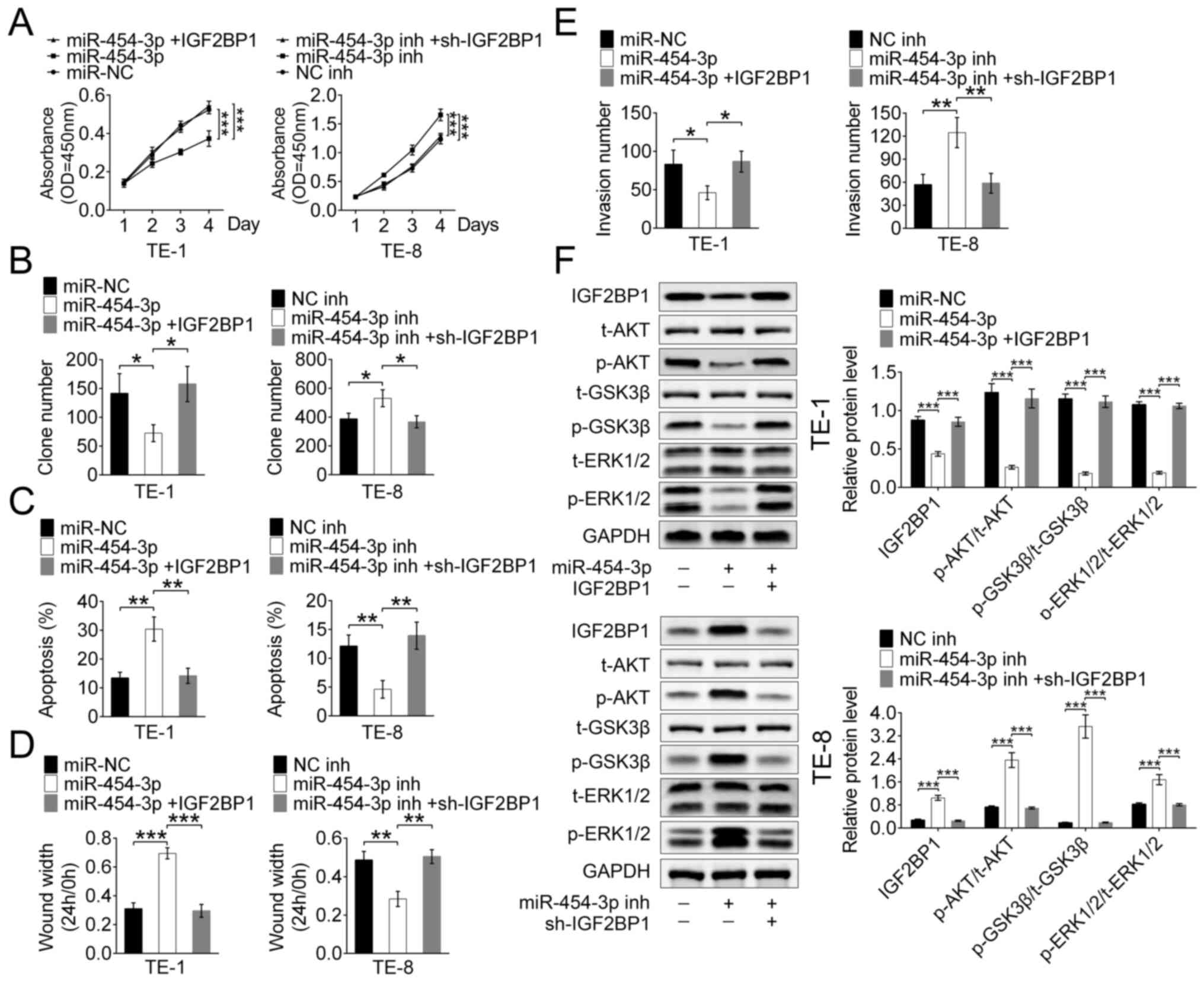 | Figure 5.miR-454-3p inhibited IGF2BP1
expression via the ERK and P13K/AKT signaling pathways in
vitro. (A) A Cell Counting Kit-8 assay and (B) colony formation
assay were used to evaluate the proliferation in TE-1 and TE-8
cells transfected with miR-454-3p mimics IGF2BP1 and their
inhibitors. (C) Flow cytometry was used to analyze the apoptosis
TE-1 and TE-8 cells transfected with miR-454-3p mimics, IGF2BP1 or
their inhibitors. (D) Wound healing and (E) Transwell assays were
used to detected the cell migration and invasion of TE-1 and TE-8
cells transfected with miR-454-3p mimics, IGF2BP1 and their
inhibitors. (F) Western blot analysis of the expression difference
of related protein levels, including t-AKT, p-AKT, p-GSK3β, t-GSK3,
p-ERK1/2 and t-ERK1/2, of the ERK and AKT signaling pathways in
TE-1 and TE-8 cells transfected with miR-454-3p mimics, IGF2BP1 or
their inhibitors. Results are presented as mean ± SD. *P<0.05,
**P<0.01 and ***P<0.001. miR, microRNA; IGF2BP1, insulin-like
growth factor 2 mRNA-binding protein 1; t-total; p-,
phosphorylated. |
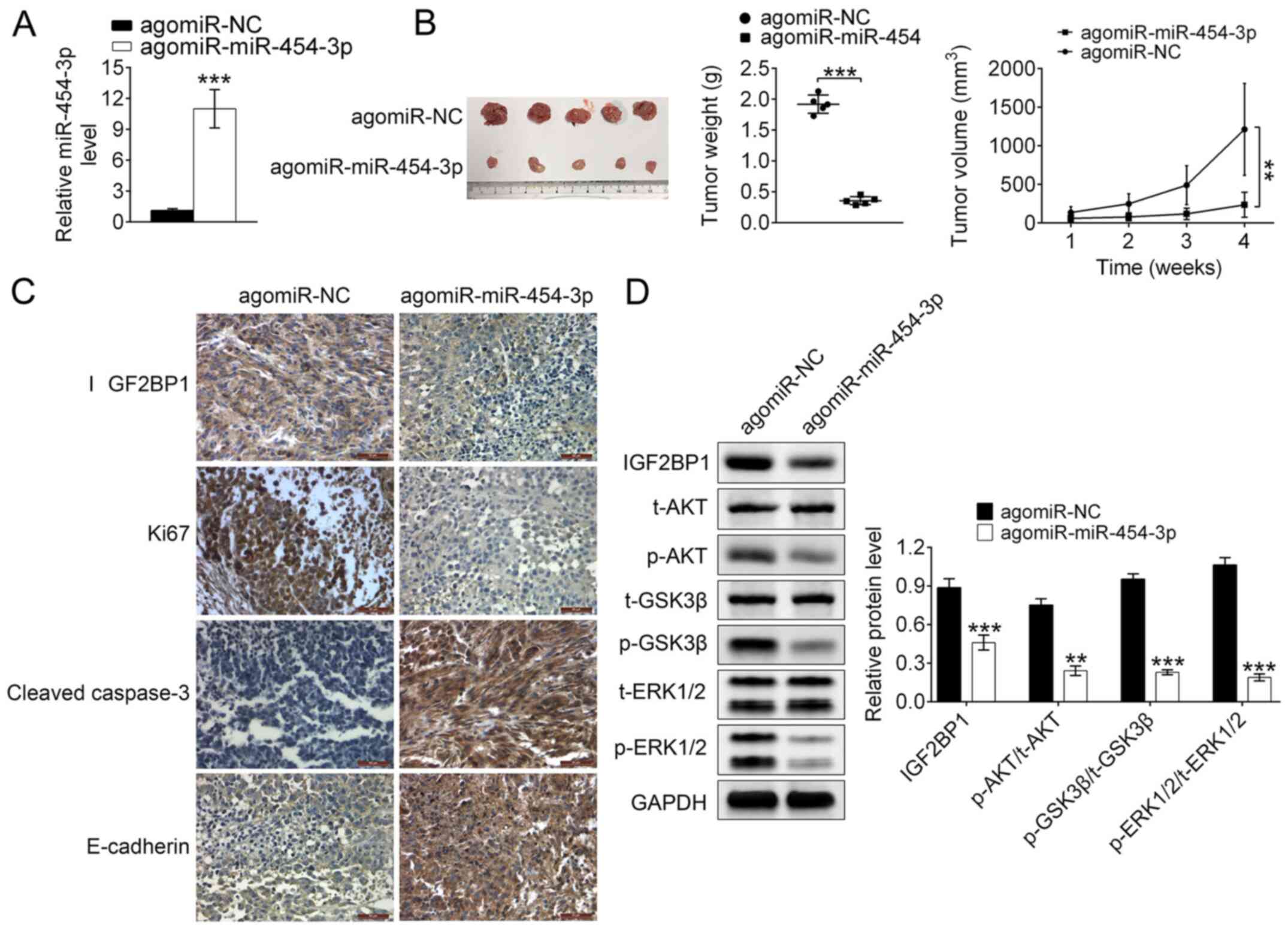
Studying the suppression of miR-140-5p
on tumor proliferation and metastasis in vivo
In addition to in vitro studies, the in
vivo suppressive effects of miR-140-5p on tumor proliferation
and metastasis were investigated using a xenograft tumor mouse
model. RT-qPCR analysis reported that the relative expression
levels of miR-454-3p were significantly higher in
TE-1-angmiR-miR-454-3p mice compared with the control (P<0.001;
Fig. 6A), which demonstrated that
the xenograft tumor mouse model was successfully constructed.
Additionally, compared with TE-1-agomiR-NC mice, the tumor size,
weight and volume were significantly decreased in
TE-1-angmiR-miR-454-3p mice compared with the control (Fig. 6B), which indicated that
miR-454-3p-overexpression could inhibit the tumor growth in ESCA.
After raising for 4 weeks, the mice were sacrificed and the
esophagus was isolated. At the time of sacrifice, compared to
initial treatment, the weight of each mouse in the agomiR-NC group
was 24.2, 23.1, 22.3, 22.2 and 21.8 g, while, compared to initial
treatment, the weight of each mouse in the agomiR-454 group was
19.7, 16.4, 17.9, 18.2 and 20.6 g. Additionally, the percentages of
the tumor weight/animal body weight in each mouse were calculated.
For the agomiR-NC mice group, the percentages of the tumor
weight/animal body weight in each mouse were 7.1, 5.8, 7.1, 8.2 and
8.5%. For the agomiR-454 group, the data were 0.7, 0.9, 0.7, 1.0
and 1.0. At the time of sacrifice, compared with the control group
(not initial treatment) due to cachexia/ascites, the weight
increase in each mouse in the agomiR-NC group was +2.52, +2.01,
+2.24, +2.15 and +1.63 g, and no weight loss was observed in the
agomiR-NC group. However, compared with the control group (not
initial treatment) due to cachexia/ascites, the weight loss in each
mouse in the agomiR-454 group was-2.11, −2.30, −1.97, −2.01 and
−1.32 g, and no weight increase was observed in the agomiR-454
group.
IHC staining analysis revealed that isolated ESCA
tumor issues from TE-1-angmiR-miR-454-3p mice exhibited higher
expression of cleaved caspase-3 and E-cadherin as well as lower
expression of Ki-67 and IGF2BP1 when compared with the
TE-1-agomiR-NC mice. The results strongly illustrated
miR-454-3p-overexpression could inhibit the tumor cell
proliferation and invasion in vivo (Fig. 6C). It was then explored whether
miR-454-3p could inhibit the ESCA cell proliferation, migration and
apoptosis by targeting IGF2BP1 via ERK and AKT pathways in
vivo. Western blot data demonstrated TE-1-angmiR-miR-454-3p
mice could induce significantly lower expression of phosphorylated
proteins, such as p-AKT, p-GSK3 and p-ERK1/2, which demonstrated
that miR-454-3p suppressed the ERK and AKT pathways through
targeting IGF2BP1 in vivo (Fig.
6D). Taken together, the xenograft tumor mouse model studies
demonstrated miR-454-3p-overexpression served as an inhibitor for
ESCA tumor proliferation and metastasis in vivo.
Discussion
ESCA is one of the most common tumors and a notable
threat to human health (18).
Despite traditional therapies having improved greatly, the
prognosis and 5-year survival rates of patients with ESCAs remain
poor (19). Due to the lack of
obvious early symptoms and precise tumor markers, the vast majority
of patients with ESCA are diagnosed at a late stage before
treatment (20). Therefore, to
improve the treatment ESCA, our understanding of its pathogenesis
needs to improve to allowed the identification of effective
therapeutic targets.
Several studies have reported that miRNAs are
involved in promoting or suppressing tumor initiation and
progression in various human malignancies, such as cervical cancer
(21), gastric cancer (22), ESCA (23) and hepatocellular carcinoma (24). Therefore, exploring the functional
targets of miRNAs as well as illustrating the molecular mechanism
underlying their function is a promising strategy for the cancer
treatment.
miR-454-3p, a widely studied miRNA, was the focus of
the present study as it participates in tumorigenesis. For example,
abnormally expressed miR-454-3p is involved in regulating tumor
progression in bladder cancer (7),
renal carcinoma (8), chondrosarcoma
(25) and non-small cell lung cancer
adenocarcinoma (26). In the current
study, by analyzing the association between clinicopathological
characteristics and miR-454-3p expression, it was revealed that
miR-454-3p was associated with the prognosis of patients with
esophageal cancer. Therefore, it was demonstrated that miR-454-3p
functions in the initiation and progression of ESCA. It is known
that the molecular pathogenesis of miRNAs in modulating the
metastasis of different types of cancer are quite different. For
example, one study reported that miR-454-3p suppressed tumor cell
growth and invasion by interacting with ZEB2 in bladder cancer
(7), while another study
demonstrated miR-454-3p suppressed the progression of lung cancer
by targeting Calbindin 1 (27).
Besides, other target genes, such as BTG1, Stat3 and
Atg12, were also found to be regulated by miR-454-3p in
chondrosarcoma initiation and progression (25). The aforementioned studies have
documented that miR-454-3p regulated tumor cell growth and invasion
by targeting different downstream genes via different signaling
pathways. However, the precise molecular mechanisms of miR-454-3P
as well as its direct target in ESCA still remain unclear.
In the present study, RT-qPCR and western blot
assays revealed that the gene and protein expression levels of
miR-454-3p were lower in tumor tissues from ESCA patients and ESCA
cells compared with healthy tissues and cells. Moreover, survival
rate analysis indicated that low miR-454-3p expression improved the
survival time of patients with ESCA compared with those with high
expression. Therefore, it was hypothesized that miR-454-3p was an
important biomarker for diagnosis of ESCA patients and its
overexpression could induce the tumor growth inhibition. The
evaluations of cell proliferation, migration and invasion were
detected using CCK-8, colony formation, wound healing and Transwell
assays, respectively, with four ESCA cells (EC9706, ECA109, TE-1
and TE-8). The in vitro results demonstrated that
miR-454-3p-overexpression inhibited ESCA cell proliferation, and
decreased the number of migrating and invasive cells; however,
these effects could be reversed by deregulating the miR-454-3p
expression. Besides, the apoptosis assay for EC9706, ECA109, TE-1
and TE-8 cells indicated that miR-454-3P was an apoptosis promotor
in tumor cell metabolism. The in vitro results were
consistent with a previous study (25) regarding the suppressive function of
miR-454-3p in tumor proliferation, migration and invasion.
IGF2BP1, a post-transcriptional regulator,
has been identified in numerous types of cancer, such as lung
(28), liver (29) and breast cancer (30), and functions as a carcinogenic gene
to accelerate cell proliferation and invasion (31). IGF2BP1 is regulated by a variety of
miRNAs, thus affecting the occurrence and development of tumors.
For example, the upregulation of miR-150 (32) or miR-491-5p (28) suppressed osteosarcoma and lung cancer
cells proliferation, respectively, by directly targeting IGF2BP1.
The present bioinformatics prediction revealed miR-454-3p could
directly bind to the 3′UTR of wild-type IGF2BP1 and
miR-454-3p-overexpression significantly inhibited the expression of
IGF2BP1 in ESCA cells in vitro, which is similar with
the aforementioned previous studies. In addition, RT-qPCR and
western blot assays demonstrated that the overexpression or
inhibition of miR-454-3p resulted in the downregulation or
upregulation of IGF2BP1, respectively. Meanwhile, Spearman's
correlation analysis also demonstrated a significant negative
correlation between IGF2BP1 and miR-454-3p expression in tumor
tissues from 40 patients with ESCA. It was therefore hypothesized
that IGF2BP1 was a direct downstream target for miR-454-3p.
To further confirm the function of miR-454-3p in
ESCA, the proliferation rate of TE-1 ESCA cancer cells with or
without antagomiR-454-3p was calculated after subcutaneous
implantation into male BALB/c-nude mice. The RT-qPCR assay for all
experimental mice showed that miR-454-3p-overexpression inhibited
tumor cell proliferation and migration, and this was consistent
with the results in vitro. Immunohistochemical staining
illustrated that tumor tissues derived from mice treated with
antag-miR-454-3p ESCA cancer cells showed higher expression levels
of activated caspase-3 and E-cadherin as well as lower expression
levels of Ki-67 compared with those in the control group. Notably,
other studies have reported that miRNAs directly targeted IGF2BP1
via suppressing the phosphorylation of AKT pathway or both AKT and
ERK pathways to accelerate tumor cells proliferation and migration
(33,34). The present western blot analysis
reported that the overexpression of miR-454-3p decreased IGF2BP1
expression and activated the protein expression of downstream
target genes p-AKT, p-GSK3 and p-ERK1/2, both in the ERK and AKT
signaling pathways. Taken together, these data demonstrated that
miR-454-3p suppressed ESCA cell proliferation, migration and
apoptosis by targeting IGF2BP1 via the ERK and AKT signaling
pathways in vivo.
In summary, it was reported that miR-454-3p had
lower expression in ESCA tissues and cells. The results further
confirmed that miR-454-3p was associated with clinical features and
prognosis of patients with ESCA. Notably, miR-129-5p could suppress
the proliferation, migration, and invasion and accelerate apoptosis
in ESCA cells in vitro. More importantly, further in
vitro and in vivo studies revealed that miR-454-3p
suppressed ESCA cell proliferation, migration and apoptosis by
targeting IGF2BP1 via the ERK and AKT pathways. Taking these finds
together, the present study demonstrated miR-454 may be a useful
biomarker and may improve our understanding of the molecular
pathogenesis of ESCA. This may enable further development of novel
therapies for patients with ESCA.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was funded by the Municipal
Science and Technology Plan (Guidance) Project of Nantong City in
2014 (grant no. HS149147).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YZ conceived and designed the experiments. AY
performed the statistical analysis. CW and LZ analyzed and
interpreted the results of the experiments. JZ and YZ performed the
experiments. YZ and AY wrote the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by The Ethics
Committee of Affiliated Haian Hospital of Nantong University
(Nantong, China). All animal experiment protocols were approved by
the Ethics Committee of Affiliated Haian Hospital of Nantong
University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ESCA
|
esophageal cancer
|
|
IGF2BP1
|
insulin-like growth factor 2
mRNA-binding protein 1
|
|
miR
|
microRNA
|
References
|
1
|
Thrift AP and Whiteman DC: The incidence
of esophageal adenocarcinoma continues to rise: Analysis of period
and birth cohort effects on recent trends. Ann Oncol. 23:3155–3162.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kim T, Grobmyer SR, Smith R, Ben-David K,
Ang D, Vogel SB and Hochwald SN: Esophageal cancer-the five year
survivors. J Surg Oncol. 103:179–183. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Iorio MV and Croce CM: MicroRNA
dysregulation in cancer: Diagnostics, monitoring and therapeutics.
A comprehensive review. EMBO Mol Med. 4:143–159. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang S, Zhang G, Zheng W, Xue Q, Wei D,
Zheng Y and Yuan J: MiR-454-3p and miR-374b-5p suppress migration
and invasion of bladder cancer cells through targetting ZEB2.
Biosci Rep. 38:BSR201814362018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wu X, Ding N, Hu W, He J, Xu S, Pei H, Hua
J, Zhou G and Wang J: Down-regulation of BTG1 by miR-454-3p
enhances cellular radiosensitivity in renal carcinoma cells. Radiat
Oncol. 9:1792014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yisraeli JK: VICKZ proteins: A
multi-talented family of regulatory RNA-binding proteins. Biol
Cell. 97:87–96. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bell JL, Wachter K, Muhleck B, Pazaitis N,
Kohn M, Lederer M and Huttelmaier S: Insulin-like growth factor 2
mRNA-binding proteins (IGF2BPs): Post-transcriptional drivers of
cancer progression? Cell Mol Life Sci. 70:2657–2675. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Vainer G, Vainer-Mosse E, Pikarsky A,
Shenoy SM, Oberman F, Yeffet A, Singer RH, Pikarsky E and Yisraeli
JK: A role for VICKZ proteins in the progression of colorectal
carcinomas: Regulating lamellipodia formation. J Pathol.
215:445–456. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Huttelmaier S, Zenklusen D, Lederer M,
Dictenberg J, Lorenz M, Meng X, Bassell GJ, Condeelis J and Singer
RH: Spatial regulation of beta-actin translation by Src-dependent
phosphorylation of ZBP1. Nature. 438:512–515. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Stohr N, Kohn M, Lederer M, Glass M,
Reinke C, Singer RH and Huttelmaier S: IGF2BP1 promotes cell
migration by regulating MK5 and PTEN signaling. Genes Dev.
26:176–189. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Su Y, Xiong J, Hu J, Wei X, Zhang X and
Rao L: MicroRNA-140-5p targets insulin like growth factor 2 mRNA
binding protein 1 (IGF2BP1) to suppress cervical cancer growth and
metastasis. Oncotarget. 7:68397–68411. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bando E, Makuuchi R, Irino T, Tanizawa Y,
Kawamura T and Terashima M: Validation of the prognostic impact of
the new tumor-node-metastasis clinical staging in patients with
gastric cancer. Gastric Cancer. 22:123–129. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen J, Ren X, Li L, Lu S, Chen T, Tan L,
Liu M, Luo Q, Liang S, Nie Q, et al: Integrative analyses of mRNA
expression profile reveal the involvement of IGF2BP1 in chicken
adipogenesis. Int J Mol Sci. 20:29232019. View Article : Google Scholar
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lang B and Zhao S: miR-486 functions as a
tumor suppressor in esophageal cancer by targeting CDK4/BCAS2.
Oncol Rep. 39:71–80. 2018.PubMed/NCBI
|
|
19
|
Jung HK, Tae CH, Lee HA, Lee H, Don Choi
K, Park JC, Kwon JG, Choi YJ, Hong SJ, Sung J, et al: Treatment
pattern and overall survival in esophageal cancer during a 13-year
period: A nationwide cohort study of 6,354 Korean patients. PLoS
One. 15:e02314562020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lin Y, Totsuka Y, Shan B, Wang C, Wei W,
Qiao Y, Kikuchi S, Inoue M, Tanaka H and He Y: Esophageal cancer in
high-risk areas of China: Research progress and challenges. Ann
Epidemiol. 27:215–221. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gomez-Gomez Y, Organista-Nava J and
Gariglio P: Deregulation of the miRNAs expression in cervical
cancer: Human papillomavirus implications. Biomed Res Int.
2013:4070522013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Riquelme I, Letelier P, Riffo-Campos AL,
Brebi P and Roa JC: Emerging role of miRNAs in the drug resistance
of gastric cancer. Int J Mol Sci. 17:4242016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hu Y, Correa AM, Hoque A, Guan B, Ye F,
Huang J, Swisher SG, Wu TT, Ajani JA and Xu XC: Prognostic
significance of differentially expressed miRNAs in esophageal
cancer. Int J Cancer. 128:132–143. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Murakami Y, Yasuda T, Saigo K, Urashima T,
Toyoda H, Okanoue T and Shimotohno K: Comprehensive analysis of
microRNA expression patterns in hepatocellular carcinoma and
non-tumorous tissues. Oncogene. 25:2537–2545. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bao X, Ren T, Huang Y, Sun K, Wang S, Liu
K, Zheng B and Guo W: Knockdown of long non-coding RNA HOTAIR
increases miR-454-3p by targeting Stat3 and Atg12 to inhibit
chondrosarcoma growth. Cell Death Dis. 8:e26052017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shao Y, Liang B, Long F and Jiang SJ:
Diagnostic MicroRNA biomarker discovery for non-small-cell lung
cancer adenocarcinoma by integrative bioinformatics analysis.
Biomed Res Int. 2017:25630852017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jin C, Lin T and Shan L: Downregulation of
calbindin 1 by miR-454-3p suppresses cell proliferation in nonsmall
cell lung cancer in vitro. Cancer Biother Radiopharm. 34:119–127.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gong F, Ren P, Zhang Y, Jiang J and Zhang
H: MicroRNAs-491-5p suppresses cell proliferation and invasion by
inhibiting IGF2BP1 in non-small cell lung cancer. Am J Transl Res.
8:485–495. 2016.PubMed/NCBI
|
|
29
|
Zhou X, Zhang CZ, Lu SX, Chen GG, Li LZ,
Liu LL, Yi C, Fu J, Hu W, Wen JM and Yun JP: miR-625 suppresses
tumour migration and invasion by targeting IGF2BP1 in
hepatocellular carcinoma. Oncogene. 35:50782016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fakhraldeen SA, Clark RJ, Roopra A, Chin
EN, Huang W, Castorino J, Wisinski KB, Kim T, Spiegelman VS and
Alexander CM: Two isoforms of the RNA binding protein, coding
region determinant-binding protein (CRD-BP/IGF2BP1), are expressed
in breast epithelium and support clonogenic growth of breast tumor
cells. J Biol Chem. 290:13386–13400. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Stohr N and Huttelmaier S: IGF2BP1: A
post-transcriptional ‘driver’ of tumor cell migration. Cell Adh
Migr. 6:312–318. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Qu Y, Pan S, Kang M, Dong R and Zhao J:
MicroRNA-150 functions as a tumor suppressor in osteosarcoma by
targeting IGF2BP1. Tumour Biol. 37:5275–5284. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang J, Cheng J, Zeng Z, Wang Y, Li X,
Xie Q, Jia J, Yan Y, Guo Z, Gao J, et al: Comprehensive profiling
of novel microRNA-9 targets and a tumor suppressor role of
microRNA-9 via targeting IGF2BP1 in hepatocellular carcinoma.
Oncotarget. 6:42040–42052. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Qin X, Sun L and Wang J: Restoration of
microRNA-708 sensitizes ovarian cancer cells to cisplatin via
IGF2BP1/Akt pathway. Cell Biol Int. 41:1110–1118. 2017. View Article : Google Scholar : PubMed/NCBI
|