Introduction
Gastric cancer (GC) is one of the most common
digestive malignancies throughout the world (1). It remains an important cancer worldwide
and was responsible for over 1,000,000 new cases in 2018 and an
estimated 783,000 mortalities, making it the fifth most frequently
diagnosed cancer and the third leading cause of cancer-associated
mortality (1). Despite the declining
morbidity and mortality rates among patients with GC in the
majority of developed countries, the disease remains largely
incurable and is still the second leading cause of
cancer-associated death worldwide (2,3).
Meanwhile, the treatment options available to patients with GC
remain limited. Due to the lack of obvious early symptoms of GC and
insufficient popularization of routine gastroscopy test, about 80%
of GC cases have been diagnosed at an advanced stage. For GC
treatment, besides surgical treatment, radiotherapy and
chemotherapy are still frequently used adjuvant treatment methods
(4). Immunotherapy has received
great interest in recent years; however, it has not become a major
treatment option for GC (5). Tumor
immunotherapy is gaining significant popularity in the field of
cancer treatment, benefiting from the in-depth study of cancer
immunosuppression in recent years (5). However, few tumor immunosuppression
studies have focused on the role of mast cells (MCs) in this
process. Therefore, the present study aimed to investigate the
mechanism by which MCs regulate cancer immunosuppression, in the
hope that improving our understanding of these functions efforts
may provide novel insights and possibly a new strategy for tumor
immunotherapy by targeting MCs.
MCs are a group of long-lived heterogeneous cells
originating from bone marrow (6).
Their effects on tumor development are numerous and complex as MCs
were originally identified by their roles in angiogenesis (7) and inflammation. For example,
accumulation of MCs has been found to accelerate inflammation and
aggravate immunosuppression in the tumor microenvironment via the
stem cell factor (SCF)/c-kit signaling pathway (8). However, previous work has also revealed
a role for MCs in regulating the adaptive immune response (6).
Tumor-induced immune suppression hinders the
cytotoxic responses of T lymphocytes, as well as natural killer
cells, and promotes tumor progression (9). There are various immunosuppressive
mechanisms in which tumors and regulatory T cells (Tregs) play a
vital role (9,10). Treg cells, which are characterized by
expression of the transcription factor forkhead box protein (Foxp)3
in the nucleus, are a functionally distinct subset of T lymphocytes
with immunosuppressive capacities necessary for maintaining immune
tolerance (11,12). Through the interaction between MCs
and Tregs influence the intensity of tumor-induced inflammation,
and can result in either the promotion or inhibition of tumor
growth (6). An increase in MCs has
been observed in numerous experimental animal tumor models, as well
as human tumor specimens (13,14). Our
previous study provided evidence of a clear correlation between the
number of MCs and expression of Foxp3 in human GC (15), but the mechanism underlying this
correlation remains unclear. Therefore, the present study aimed to
resolve these mechanisms in GC.
OX40, the receptor of OX40L, could be expressed by
effector and memory CD4+ T cells (16). Studies have shown that inhibition of
OX40/OX40L signaling pathway can regulate inflammation and immune
response and thereby promote GC patient recovery (17), suggesting OX40L may be related to the
progression of GC, and can act as a marker to determine the
malignancy and prognosis of GC. However, no research on this has
been reported so far. Therefore, the results presented in this
study may help to elucidate a new strategy for GC immunotherapy
targeting MCs.
Materials and methods
Patients and specimens
Samples were collected from 40 patients with GC at
the Ningbo Medical Center of Lihuili Hospital (Ningbo, China)
following approval from the Ethics Committee of Lihuili Hospital
(approval no. KY2020PJ020). Written consent was obtained from the
patients prior to the collection of samples. The inclusion and
exclusion criteria were as follows: i) Diagnosis of GC confirmed by
pathology; ii) without anticancer treatment before surgery; iii)
underwent curative resection for GC between January 2015 and
December 2017 (patients in stage IV underwent resection only); iv)
with complete clinicopathological and follow-up data.
Postoperatively, all the specimens were stored using paraffin. The
specimens were fixed in 4% paraformaldehyde at 25°C for 4 h, and
then transferred to 70% ethanol. The individual lobes of tumor
biopsy material were placed in processing cassettes, dehydrated
through a serial alcohol gradient, embedded in paraffin wax blocks,
and then subjected to paraffin section with a thickness of 5 µm.
Clinical stages were classified according to the 7th
Tumor-Node-Metastasis (TNM) staging system (18). The clinical characteristics of all
patients are summarized in Table
I.
 | Table I.Clinical characteristics and the
stages of the patients with gastric cancer. |
Table I.
Clinical characteristics and the
stages of the patients with gastric cancer.
|
Characteristics | Value |
|---|
| Sex,
male/femalea | 23/17
(57.5/42.5) |
| Age,
yearsb | 30-75 |
| TNM
stagea |
|
| I | 10 (20) |
| II | 10 (20) |
|
III | 10 (20) |
| IV | 10 (20) |
Immunofluorescence
Primary rabbit anti-Mast Cell Tryptase (MCT) (1:500;
cat. no. bs-2725R; BIOSS) and mouse anti-FoxP3 (1:500; cat. no.
sc-166212; Santa Cruz Biotechnology, Inc.) antibodies were used to
detect MCT+ cells and FoxP3 expression, respectively.
Other primary antibodies including rat anti-OX40 (1:500; cat. no.
sc-71768; Santa Cruz Biotechnology, Inc.), mouse anti-IL-9R (1:500;
cat. no. sc-515622; Santa Cruz Biotechnology, Inc.), rabbit
anti-IL-9 (1:500; cat. no. bs-10435R; BIOSS) and rabbit anti-TGF β1
(1:500; cat. no. bs-0086R; BIOSS) antibodies were also used. Before
immunofluorescence analysis, the sections were blocked with donkey
serum albumin (1:50; cat. no. BMS0140; Abbkine Scientific Co.,
Ltd.) in PBS for 1 h at room temperature and then incubated at 4°C
with the aforementioned primary antibodies. Then the sections were
incubated with secondary goat anti-rat IgG antibody (1:500; cat.
no. bs-0293G), goat anti-rabbit IgG antibody (1:500; cat. no.
bs-0295G) or goat anti-Mouse IgG antibody (1:500; bs-0296G) (all
BIOSS) at 37°C for 1 h. Slides were then mounted with Vectashield
containing DAPI (Vector Laboratories, Inc.; Maravai LifeSciences)
and visualized and images captured using a fluorescence microscope
(Olympus Corporation; magnification, ×200) coupled to a CCD camera
(Nikon Corporation). Negative controls, in which PBS was used in
place of primary antibodies, were included for each marker. The
mean intensity of fluorescence was analyzed using ImageJ (version
1.45r; National Institutes of Health).
Statistical analysis
All statistical analyses were performed using SPSS
version 14.0 software (SPSS Inc.). The results are expressed as
mean ± standard error of the mean of three independent experiments.
The means of the tumor and non-tumor groups were compared using a
paired Student's t-test. Multiple comparisons were performed using
a one-way analysis of variance followed by Tukey's post hoc test,
which was used to analyze clinical stages in relation to mean
levels of OX40L. P<0.05 was considered to indicate a
statistically significant difference. Relationships between
tryptase and Foxp3, and tryptase and TGFβ1 in GC tissue or from The
Cancer Genome Atlas database (https://tcga-data.nci.nih.gov/tcga) were analyzed with
the Spearman's rank correlation coefficient.
Results
The role of MCs in regulating
microenvironment immunity of GC
MCs play an important role in the tumor
microenvironment of GC (15),
therefore, to clarify the role of MCs in GC, an immunofluorescence
assay to detect the expression of MCs in 40 human paired GC and
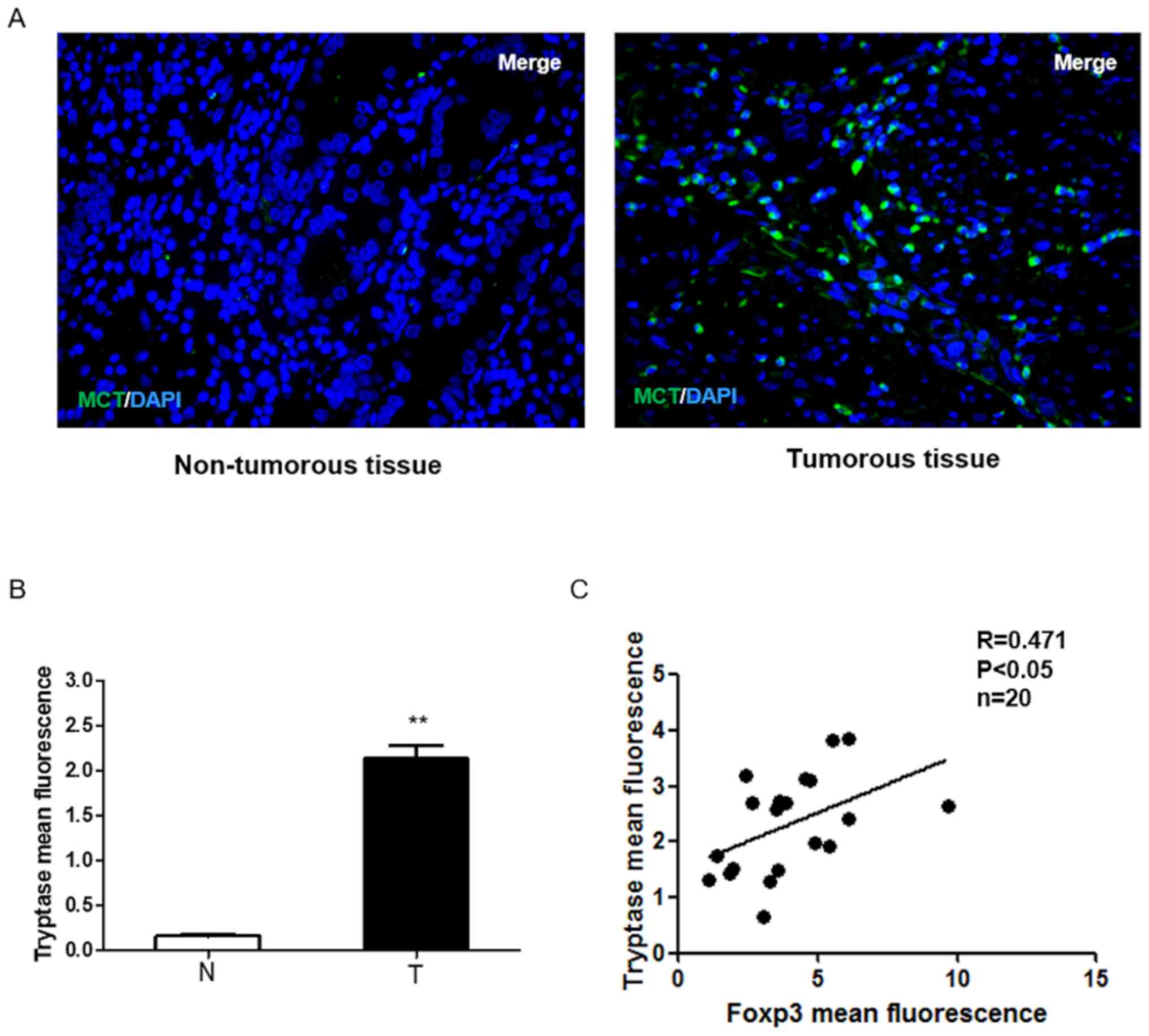
non-tumor tissue samples. As shown in Fig. 1A and B, tryptase was upregulated in
GC specimens compared with the surrounding non-tumorous tissues. In
total, 20 of the 40 samples were used to detect Foxp3 expression in
GC tissues. The results showed that MCs displayed high levels of
Foxp3 in GC samples and tryptase and Foxp3 expressions were
positively correlated in human GC (P<0.05; Fig. 1C), which suggests that the
infiltration of MCs in GC may increase the number of Tregs and MCs
may play an important role in regulating tumor immunity.
MCs regulate Treg cells in the GC
tumor microenvironment by secreting TGFβ1
It has been previously reported that GC cells induce
the increased human CD4+ Foxp3+ Tregs via the
production of TGFβ1 (19), but
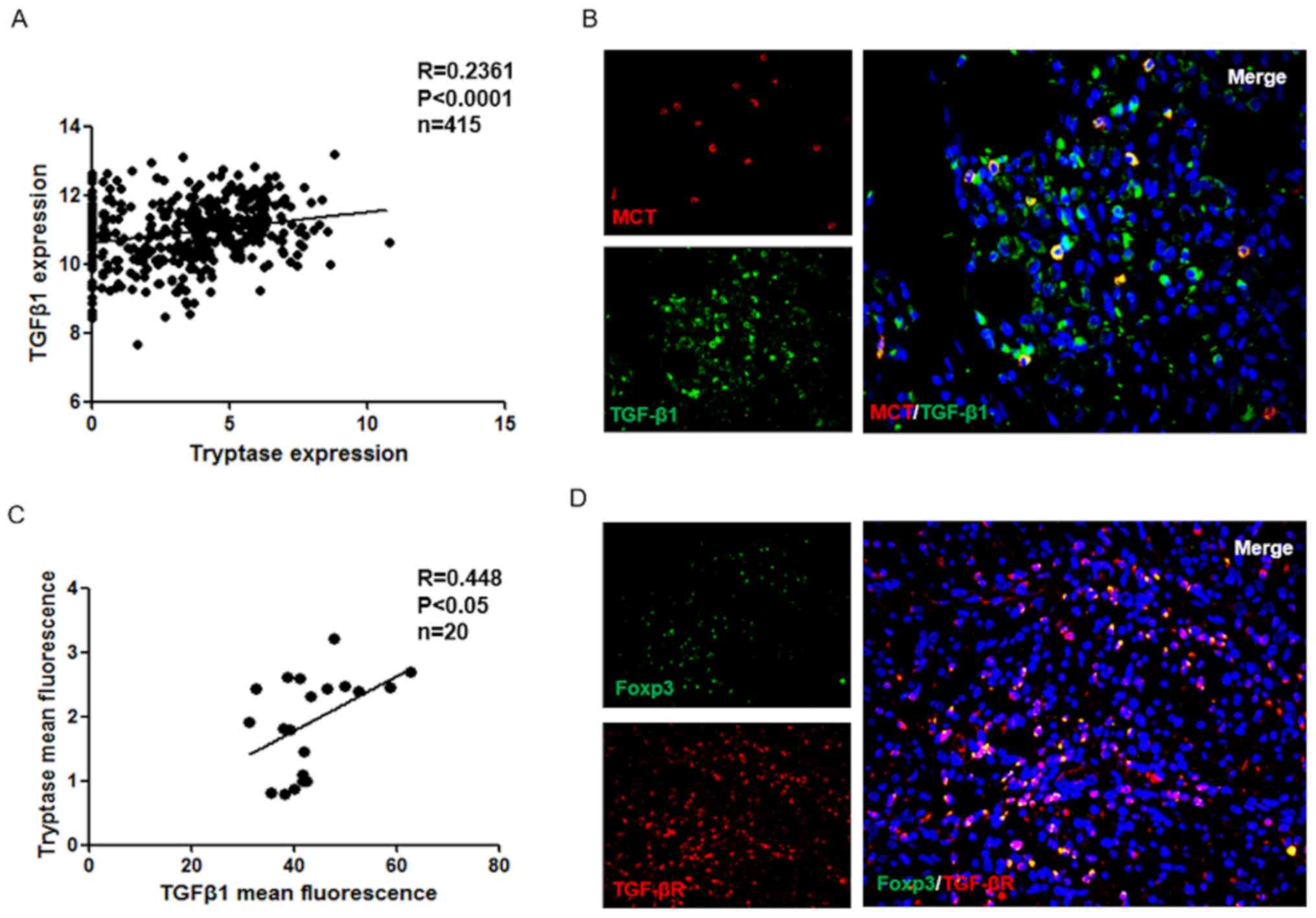
whether mast cells can secrete TGFβ1 in GC is unclear. Using TCGA
database, it was demonstrated that TGFβ1 expression has a positive
correlation with MCs in the GC group (P<0.001; Fig. 2A). In addition, immunofluorescence
was used to analyze the localization of MCs and TGFβ1 in GC tissues
(Fig. 2B), and the results were
examined using correlation analysis. As presented in Fig. 2C, the TGFβ1 expression exhibited a
positive correlation with MCs in GC tissues (P<0.05).
Furthermore, the expression of TGFβR in GC tissues was also
examined. The results indicated that TGFβR is expressed in Treg
cells (Fig. 2D). In summary, these
data suggested that the cytokine TGFβ1 may be a molecule involved
in crosstalk between MC and Treg cells.
Tregs regulate MCs in the tumor
microenvironment by producing IL-9
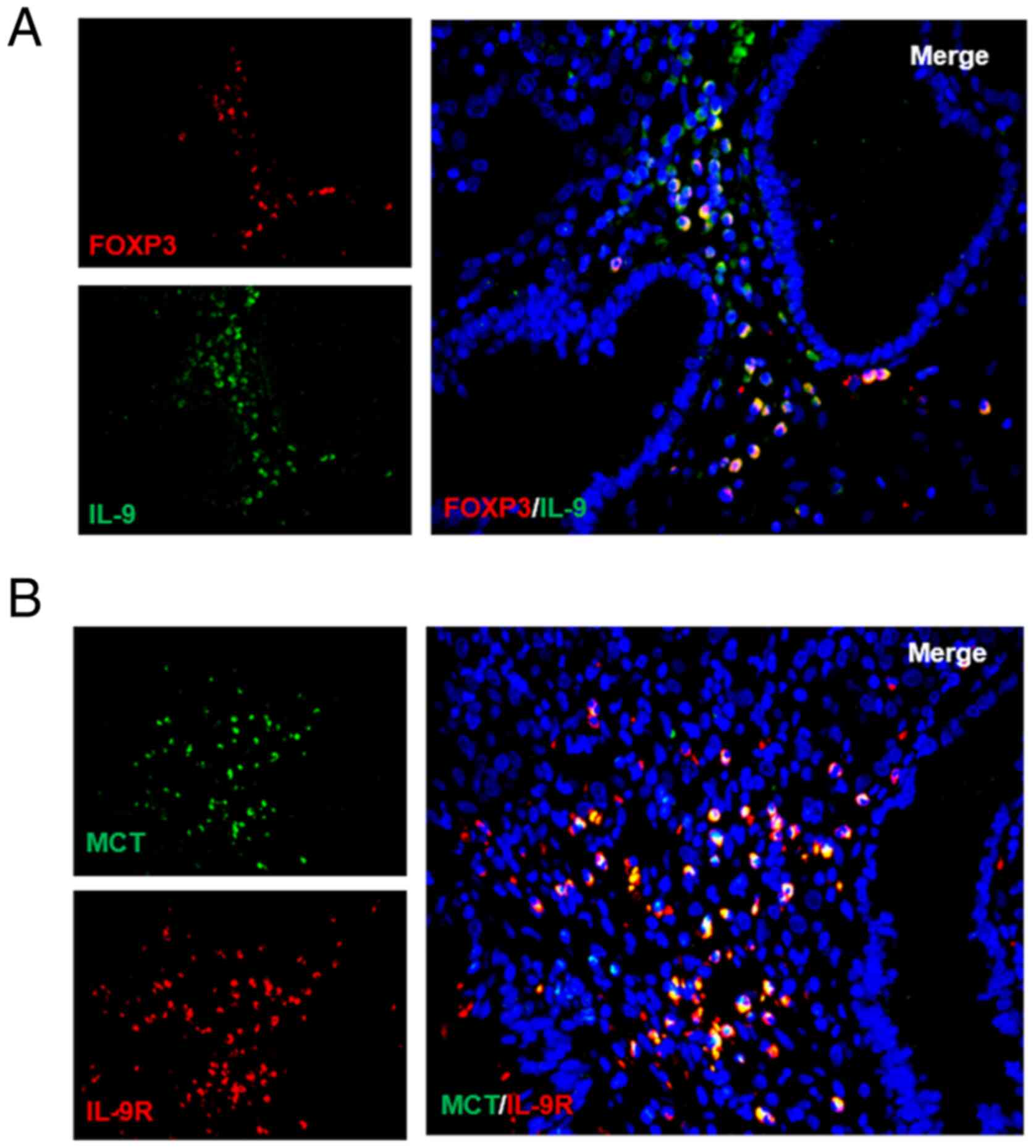
Considering that IL-3, IL-4 and IL-9 affect MC
proliferation but only IL-9 is able to promote the growth of MCs
from bone marrow and MC progenitors (20,21), it
was hypothesized that Tregs may regulate MCs through IL-9. To test
this hypothesis, IL-9 and Treg cells were probed for using
immunofluorescence. As expected, it was reported that Tregs
secreted IL-9 (Fig. 3A). High IL-9R
expression was also detected on the surface of MCs for the first
time (Fig. 3B). Together, these
results showed that a positive feedback regulation system may exist
between MCs and Treg cells that operates through TGFβ1 and
IL-9.
Expression of OX40L in MCs is an
important marker for determining GC malignancy and prognosis
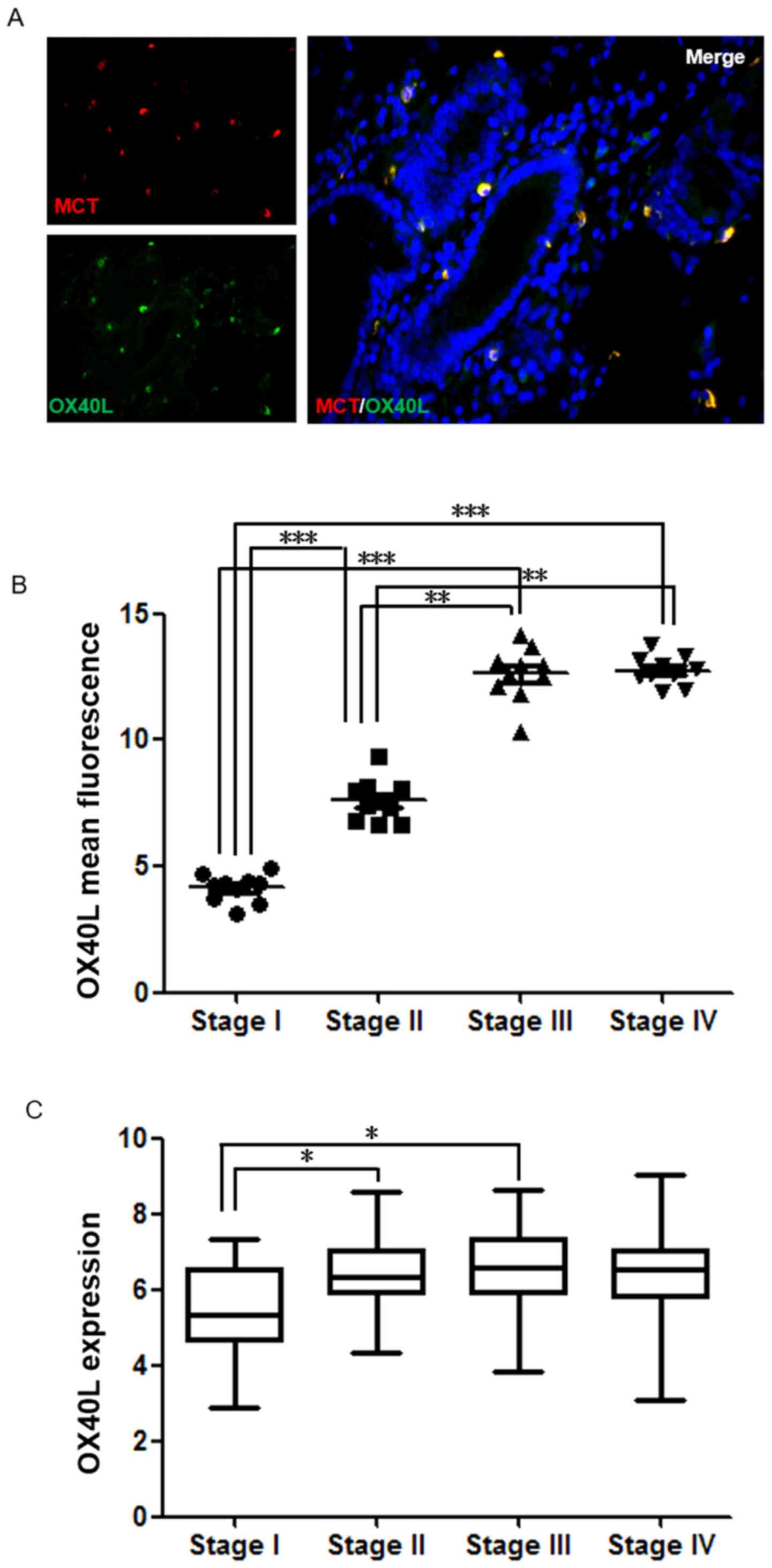
OX40L was also a protein of interest. Notably, in GC
tissues, it was demonstrated that the majority of OX40L was
expressed on MCs, which has not been reported previously to the
best of our knowledge (Fig. 4A). By
analyzing clinical samples, it was found that OX40L expression in
GC was significantly associated with TNM stage. As presented in
Fig. 4B, there was a significant
difference in OX40L levels between Stage I and Stages II–IV
(P<0.001). To further confirm whether the expression of OX40L is
an important marker for determining GC malignancy and prognosis,
TCGA data were analyzed and there was a significant difference in
OX40L levels between Stages I and II (P<0.05; Fig. 4C). This suggests that OX40L expressed
by MCs may be an important marker for determining GC malignancy and
prognosis.
Discussion
GC is the most common gastrointestinal tumor
(1). Although the medical
understanding of this disease is improving, the rates of incidence
and mortality of GC are still high, especially in China. In 2018,
42.6% of new GC cases and 45% of GC-induced deaths occurred in
China (22). The research on tumor
immunity observed GC has led to several important discoveries
(23), yet further exploration is
needed, despite the progress made.
Our previous study focused on the roles of MCs in
GC, reporting that the frequency of MCs is higher in tumors
compared with in healthy tissues, and MC levels are correlated with
TNM stage (13). Ribatti et
al (14) also demonstrated that
MC density is correlated with the progression gastric carcinoma and
the density of MCs is positively correlated with the development of
the disease from stage I to stage IV. Some studies have suggested
that MCs play a protective role in human cancer (24–27);
however, in GC, the overall role of MCs in promoting cancer is
unclear. The tumor promoting mechanism of MCs is exceedingly
complicated, and involves tissue remodeling, angiogenesis and
immune regulation (14). Therefore,
conducting research on MCs can be very difficult, leading the
present study to focus on the immunoregulatory function of MCs.
Several studies have demonstrated increased presence of
intratumoral and circulating Treg cells in gastric adenocarcinomas
(28–30). In gastric tumors, Treg abundance is
also correlated with decreased overall survival (28,31),
and, in particular, a high Foxp3 level is an independent factor
associated with worse overall survival time and rate (30). In our previous study a positive
correlation was identified between MCs and Tregs using flow
cytometry (15). In the present
study, immunofluorescence was used to further confirm this
correlation and to further explore the possible mechanisms of
interaction.
It is widely recognized that TGFβ plays several
central roles in carcinogenesis (31–33). The
TGFβ family of proteins regulates numerous cellular functions,
including cell growth, differentiation, adhesion, migration and
apoptosis. TGFβ is also able to manipulate the tumor
microenvironment to promote carcinogenesis. Malignant tumor cells
secrete a large amount of TGFβ protein, which not only accelerates
the proliferation and migration of cancer cells, but also enables
cancer cells to evade the immune system. TGFβ also induces Tregs to
inhibit effector T cells, a set of cells which have the capability
of recognizing and killing cancer cells like Cytotoxic T lymphocyte
(34). In addition, TGFβ can act
directly on effector T cells, natural killer cells and B cells, to
inhibit their immunological activities (34). Yuan et al (19) found that GC cells could induce human
CD4+ Foxp3+ Tregs through the production of
TGFβ1. TGFβ1 is also an important product of MCs (35), but whether MCs can regulate Tregs in
human GC by secreting TGFβ1 has not been previously reported. The
present study demonstrated that there is a correlation between
TGFβ1 and MCs in GC (Fig. 2),
suggesting that MCs are also involved in TGFβ1 secretion, which can
have a positive effect on Tregs.
IL-9 was also investigated, another important factor
linking Tregs and MCs, which has been shown to promote MC
proliferation and function (36).
Previous work has shown that the number of basal MCs was normal
without IL-9 (37); however the
presence of IL-9, in combination with Stem cell factor, was able to
promote the proliferation of MCs from bone marrow and MC
progenitors (36). The primary
source of IL-9 is T lymphocytes, including natural Tregs and
inducible Tregs, both of which are Foxp3+ populations
that are able to secrete IL-9 (38,39).
However, there is conflicting evidence regarding the production of
IL-9 from human Treg cells (40,41).
Additionally, in human donors the co-expression of Foxp3 and IL-9
has not been reported either. There are several pieces of evidence
connecting IL-9 and MCs (42). IL-9
is a key proliferation or differentiation factor and
chemoattractant for MCs (43,44) and
has been previously implicated as a key cytokine important for
regulating the interactions between Tregs and MCs in other systems,
such as inflammation (39).
Furthermore, IL-9 production by Tregs recruits MCs that are
essential for Treg-induced immune-suppression (45). Thus, the present study sought to
analyze the functional role of IL-9 in Treg-MC interactions in GC.
The current study analyzed Foxp3 and IL-9 expression in GC, and
reported that these were co-expressed, indicating that Treg cells
can secrete IL-9 in GC tissues. In addition, we discovered for the
first time that IL-9R is expressed on the surface of MC. Based on
these results, it is reasonable to speculate that Tregs can
regulate MCs by secreting IL-9 in GC. These results suggested that
a positive feedback regulation system exists between Treg cells and
MCs that operates through TGFβ1 and IL-9. This feedback may have an
inhibitory effect and result in cancer-mediated
immunosuppression.
The present study also investigated the important
molecule OX40L. It was found that patients with high expression of
OX40L had a worse prognosis (Fig.
4). However, the expression of OX40L did not significantly
affect the survival curve (data not shown). Further analysis showed
that OX40L expression was also correlated with TNM stage (Fig. 4), and OX40L expression was higher in
GC patients at late-stage (stages II–IV) compared with that in
stage I patients. Previous studies have found an association
between MC and TNM staging of GC (13,15),
leading to the speculation that changes in OX40L expression could
be caused by MCs. Previous work has shown that OX40L expression is
upregulated in response to antigen presentation on multiple
antigen-presenting cells (46). The
type of cells that can be induced to express OX40L is broader
compared with that for OX40, and studies have reported expression
of OX40L on MCs (47,48), as well as vascular endothelial cells
(49). In addition, MCs can also
promote angiogenesis (24),
suggesting that the increased expression of OX40L is likely caused
by neovascularization. The results of the present study
demonstrated a significant difference in OX40L levels between stage
I and stages II–IV GC patients, suggesting that OX40L can be used
as a potential novel GC marker for clinical evaluation of
occurrence, development and metastasis of GC. Although OX40L has
not been clinically used as a prognostic factor for GC and the
immune-promoting effect of OX40L may not be dominant in GC, the
present study does provide a new perspective on the unique role of
MCs in GC.
In summary, the present results showed increased
expression of both MCs and Foxp3 in GC samples compared with normal
tissues. The significant correlation between MCs and Foxp3 supports
the hypothesis that MCs play a role in the immune suppression seen
in GC and may, at least partially, affect the prognosis. The
mechanism of action between these two cell types was further
investigated, revealing that there be a regulatory feedback
mechanism involving TGFβ1 and IL-9. TGFβ1 has been shown to play an
important role in GC (50). The
current study showed that MCs are involved in the secretion of
TGFβ1, and can promote Tregs through TGFβ1. Tregs can also
positively regulate MCs by producing IL-9 to promote MC function.
OX40L may serve as a potential prognostic indicator of GC and could
provide a new perspective to study the angiogenesis in this
disease. However, the present study has some limitations. First,
the study only focused on tumor immunity and did not thoroughly
analyze detailed clinicopathological data, hence the present data
lack some important clinicopathological characteristics, such as
lymph node metastasis and liver metastasis. In addition, the
present study only provided preliminarily evidence to support the
hypothesis that there may be a positive feedback regulation system
between Treg cells and MCs operating through TGFβ1 and IL-9.
Therefore, further research is needed to validate the current
results. Ultimately, further experiments could further improve our
understanding of the mechanistic interactions between Tregs and
MCs.
Acknowledgements
The authors of the present study would like to thank
Professor Wu Ke (Wuhan Union Hospital) for critically reading this
manuscript.
Funding
The present study was funded by the Natural Science
Foundation of Ningbo (grant no. 2015A610223).
Availability of data and materials
All data generated or analyzed during this study are
included within the article.
Authors' contributions
HY conceived the study. YZ and HY designed the
study. YZ searched the literature and collected the data. YZ, SY
and KD performed the experiments and interpreted the data. SY, JS,
KD, QL, YW and WC performed the statistical analysis. All authors
have read and approved the final manuscript.
Ethics approval and consent to
participate
Human samples were obtained with informed written or
oral consent from the donors. The study was approved by the Ethical
committee of Ningbo Medical Center Lihuili Hospital (Ningbo, China;
approval no. approval no. KY2020PJ020).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang LH, Su L and Wang JT: Correlation
between elevated FOXP3 expression and increased lymph node
metastasis of gastric cancer. Chin Med J (Engl). 123:3545–3549.
2010.PubMed/NCBI
|
|
3
|
Yoshii M, Tanaka H, Ohira M, Muguruma K,
Iwauchi T, Lee T, Sakurai K, Kubo N, Yashiro M, Sawada T and
Hirakawa K: Expression of forkhead box P3 in tumour cells causes
immunoregulatory function of signet ring cell carcinoma of the
stomach. Br J Cancer. 106:1668–1674. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tan Z: Recent advances in the surgical
treatment of advanced gastric cancer: A review. Med Sci Monit.
25:3537–3541. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Magalhães H, Fontes-Sousa M and Machado M:
Immunotherapy in advanced gastric cancer: An overview of the
emerging strategies. Can J Gastroenterol Hepatol.
2018:27324082018.PubMed/NCBI
|
|
6
|
Khazaie K, Blatner NR, Khan MW, Gounari F,
Gounaris E, Dennis K, Bonertz A, Tsai FN, Strouch MJ, Cheon E, et
al: The significant role of mast cells in cancer. Cancer Metastasis
Rev. 30:45–60. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ribatti D and Crivellato E: Mast cells,
angiogenesis, and tumour growth. Biochim Biophys Acta. 1822:2–8.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huang B, Lei Z, Zhang GM, Li D, Song C, Li
B, Liu Y, Yuan Y, Unkeless J, Xiong H and Feng ZH: SCF-mediated
mast cell infiltration and activation exacerbate the inflammation
and immunosuppression in tumor microenvironment. Blood.
112:1269–1279. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rabinovich GA, Gabrilovich D and Sotomayor
EM: Immunosuppressive strategies that are mediated by tumor cells.
Annu Rev Immunol. 25:267–296. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Piconese S and Colombo MP: Regulatory T
cells in cancer. Blood. 108:804–811. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fontenot JD, Gavin MA and Rudensky AY:
Foxp3 programs the development and function of CD4+CD25+ regulatory
T cells. Nat Immunol. 4:330–336. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kryczek I, Liu R, Wang G, Wu K, Shu X,
Szeliga W, Vatan L, Finlayson E, Huang E, Simeone D, et al: FOXP3
defines regulatory T cells in human tumor and autoimmune disease.
Cancer Res. 69:3995–4000. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ribatti D and Crivellato E: Chapter 4 the
controversial role of mast cells in tumor growth. Int Rev Cell Mol
Biol. 275:89–131. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ribatti D, Guidolin D, Marzullo A, Nico B,
Annese T, Benagiano V and Crivellato E: Mast cells and angiogenesis
in gastric carcinoma. Int J Exp Pathol. 91:350–356. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhao Y, WU K, Cai K, Zhai R, Tao K, Wang G
and Wang J: Increased numbers of gastric-infiltrating mast cells
and regulatory T cells are associated with tumor stage in gastric
adenocarcinoma patients. Oncol Lett. 4:755–758. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
De Smedt T, Smith J, Baum P, Fanslow W,
Butz E and Maliszewski C: Ox40 costimulation enhances the
development of T cell responses induced by dendritic cells in vivo.
J Immunol. 168:661–670. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huang L, Wang M, Yan Y, Gu W, Zhang X, Tan
J, Sun H, Ji W and Chen Z: OX40L induces helper T cell
differentiation during cell immunity of asthma through PI3K/AKT and
P38 MAPK signaling pathway. J Transl Med. 16:742018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Edge SB and Compton CC: The American joint
committee on cancer: The 7th edition of the AJCC cancer staging
manual and the future of TNM. Ann Surg Oncol. 17:1471–1474. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yuan XL, Chen L, Zhang TT, Ma YH, Zhou YL,
Zhao Y, Wang WW, Dong P, Yu L, Zhang YY and Shen LS: Gastric cancer
cells induce human CD4+Foxp3+ regulatory T cells through the
production of TGF-β1. World J Gastroenterol. 17:2019–2027. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang J, Zhang Y, Zeng Y, Ge S, Sun X, Jia
M, Wu Y and Wang N: Isoimperatorin reduces the effective dose of
dexamethasone in a murine model of asthma by inhibiting mast cell
activation. Phytother Res. 2020.(Epub ahead of print). View Article : Google Scholar
|
|
21
|
Feng LL, Gao JM, Li PP and Wang X: IL-9
contributes to immunosuppression mediated by regulatory T cells and
mast cells in B-cell non-hodgkin's lymphoma. J Clin Immunol.
31:1084–1094. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yu F, Tian T, Deng B, Wang T, Qi Q, Zhu M,
Yan C, Ding H, Wang J, Dai J, et al: Multi-marker analysis of
genomic annotation on gastric cancer GWAS data from Chinese
populations. Gastric Cancer. 22:60–68. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Muro K, Chung HC, Shankaran V, Geva R,
Catenacci D, Gupta S, Eder JP, Golan T, Le DT, Burtness B, et al:
Pembrolizumab for patients with PD-L1-positive advanced gastric
cancer (KEYNOTE-012): A multicentre, open-label, phase 1b trial.
Lancet Oncol. 17:717–726. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lis R, Touboul C, Mirshahi P, Ali F,
Mathew S, Nolan DJ, Maleki M, Abdalla SA, Raynaud CM, Querleu D, et
al: Tumor associated mesenchymal stem cells protects ovarian cancer
cells from hyperthermia through CXCL12. Int J Cancer. 128:715–725.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ogino S, Shima K and Baba Y: Colorectal
cancer expression of peroxisome proliferator-activated
receptor-gamma (PPARG, PPARgamma)is associated with good prognosis.
Gastroenterology. 136:1242–1250. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sinnamon MJ, Carter KJ, Sims LP, Lafleur
B, Fingleton B and Matrisian LM: A protective role of mast cells in
intestinal tumorigenesis. Carcinogenesis. 29:880–886. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhao YB, Wang JL and Wang GB: The function
of mast cells in gastric cancer. Gastroenterology. 19:2246–2250.
2011.
|
|
28
|
Sasada T, Kimura M, Yoshida Y, Kanai M and
Takabayashi A: CD4+CD25+ regulatory T cells
in patients with gastrointestinal malignancies: Possible
involvement of regulatory T cells in disease progression. Cancer.
98:1089–1099. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shen Z, Zhou S, Wang Y, Li RL, Zhong C,
Liang C and Sun Y: Higher intratumoral infiltrated
Foxp3+ Treg numbers and
Foxp3+/CD8+ ratio are associated with adverse
prognosis in resectable gastric cancer. J Cancer Res Clin Oncol.
136:1585–1595. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mizukami Y, Kono K, Kawaguchi Y, Akaike H,
Kamimura K, Sugai H and Fujii H: Localisation pattern of
Foxp3+ regulatory T cells is associated with clinical
behaviour in gastric cancer. Br J Cancer. 98:148–153. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bataller A, Montalban-Bravo G, Soltysiak
KA and Garcia-Manero G: The role of TGFβ in hematopoiesis and
myeloid disorders. Leukemia. 33:1076–1089. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhu H, Luo H, Shen Z, Hu X, Sun L and Zhu
X: Transforming growth factor-β1 in carcinogenesis, progression,
and therapy in cervical cancer. Tumour Biol. 37:7075–7083. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Syed V: TGF-β signaling in cancer. J Cell
Biochem. 117:1279–1287. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Schwartz M, Zhang Y and Rosenblatt JD: B
cell regulation of the anti-tumor response and role in
carcinogenesis. J Immunother Cancer. 4:402016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Elieh Ali Komi D and Grauwet K: Role of
mast cells in regulation of T cell responses in experimental and
clinical settings. Clin Rev Allergy Immunol. 54:432–445. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Matsuzawa S, Sakashita K, Kinoshita T, Ito
S, Yamashita T and Koike K: IL-9 enhances the growth of human mast
cell progenitors under stimulation with stem cell factor. J
Immunol. 170:3461–3467. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Townsend JM, Fallon GP, Matthews JD, Smith
P, Jolin EH and McKenzie NA: IL-9-deficient mice establish
fundamental roles for IL-9 in pulmonary mastocytosis and goblet
cell hyperplasia but not T cell development. Immunity. 13:573–583.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Schmitt E, Van Brandwijk R, Van Snick J,
Siebold B and Rüde E: Tcgfiii/p40 is produced by naive murine
cd4+ t cells but is not a general t cell growth factor.
Eur J Immunol. 19:2167–2170. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lu LF, Lind EF, Gondek DC, Bennett KA,
Gleeson MW, Pino-Lagos K, Scott ZA, Coyle AJ, Reed JL, Van Snick J,
et al: Mast cells are essential intermediaries in regulatory T-cell
tolerance. Nature. 442:997–1002. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Beriou G, Bradshaw EM, Lozano E,
Costantino CM, Hastings WD, Orban T, Elyaman W, Khoury SJ, Kuchroo
VK, Baecher-Allan C and Hafler DA: TGF-beta induces IL-9 production
from human Th17 cells. J Immunol. 185:46–54. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Putheti P, Awasthi A, Popoola J, Gao W and
Strom TB: Human CD4+ memory T cells can become
CD4+IL-9+ T cells. PLoS One. 5:e87062010.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sehra S, Yao W, Nguyen ET, Glosson-Byers
NL, Akhtar N, Zhou B and Kaplan MH: TH9 cells are required for
tissue mast cell accumulation during allergic inflammation. J
Allergy Clin Immunol. 136:433–40.e1. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hauber HP, Bergeron C and Hamid Q: IL-9 in
allergic inflammation. Int Arch Allergy Immunol. 134:79–87. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Renga G, Moretti S, Oikonomou V, Borghi M,
Zelante T, Paolicelli G, Costantini C, De Zuani M, Villella VR,
Raia V, et al: IL-9 and mast cells are key players of candida
albicans commensalism and pathogenesis in the gut. Cell Rep.
23:1767–1778. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Eller K, Wolf D, Huber JM, Metz M, Mayer
G, McKenzie ANJ, Maurer M, Rosenkranz AR and Wolf AM: IL-9
production by regulatory T cells recruits mast cells that are
essential for regulatory T cell-induced immune-suppression. J
Immunol. 186:83–91. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Webb GJ, Hirschfield GM and Lane PJL:
OX40, OX40L and autoimmunity: A comprehensive review. Clin Rev
Allergy Immunol. 50:312–332. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kashiwakura J, Yokoi H, Saito H and
Okayama Y: T cell proliferation by direct cross-talk between OX40
ligand on human mast cells and OX40 on human T cells: Comparison of
gene expression profiles between human tonsillar and lung-cultured
mast cells. J Immunol. 173:5247–5257. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Nakae S, Suto H, Kakurai M, Sedgwick JD,
Tsai M and Galli SJ: Mast cells enhance T cell activation:
Importance of mast cell-derived TNF. Proc Natl Acad Sci USA.
102:6467–6472. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Imura A, Hori T, Imada K, Ishikawa T,
Tanaka Y, Maeda M, Imamura S and Uchiyama T: The human OX40/gp34
system directly mediates adhesion of activated T cells to vascular
endothelial cells. J Exp Med. 183:2185–2195. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Ebert MP, Yu J, Miehlke S, Fei G,
Lendeckel U, Ridwelski K, Stolte M, Bayerdörffer E and
Malfertheiner P: Expression of transforming growth factor beta-1 in
gastric cancer and in the gastric mucosa of first-degree relatives
of patients with gastric cancer. Br J Cancer. 82:1795–1800. 2000.
View Article : Google Scholar : PubMed/NCBI
|